The Disturbance of Hepatic and Serous Lipids in Aristolochic Acid Ι Induced Rats for Hepatotoxicity Using Lipidomics Approach

Abstract
1. Introduction
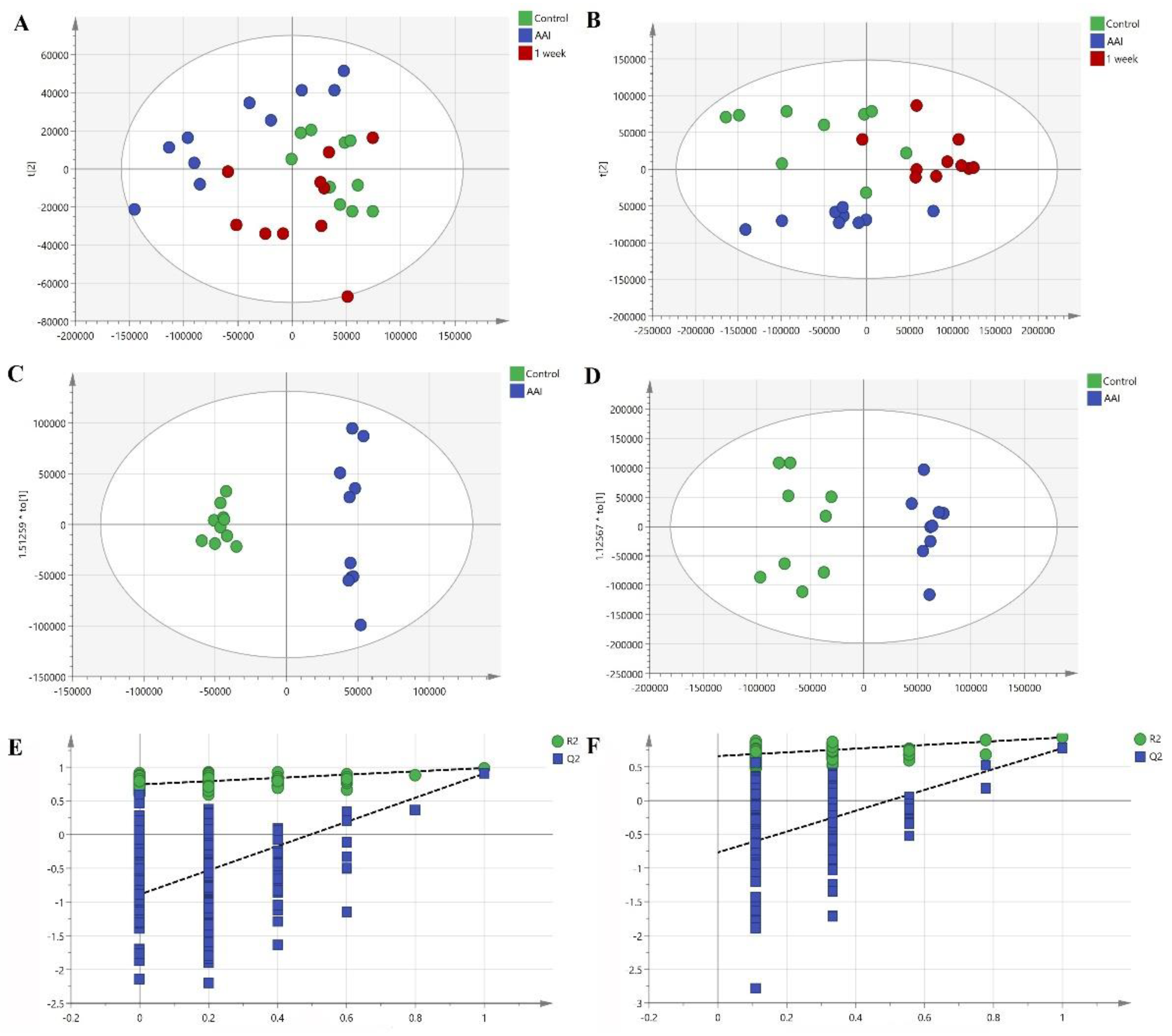
2. Results and Discussion
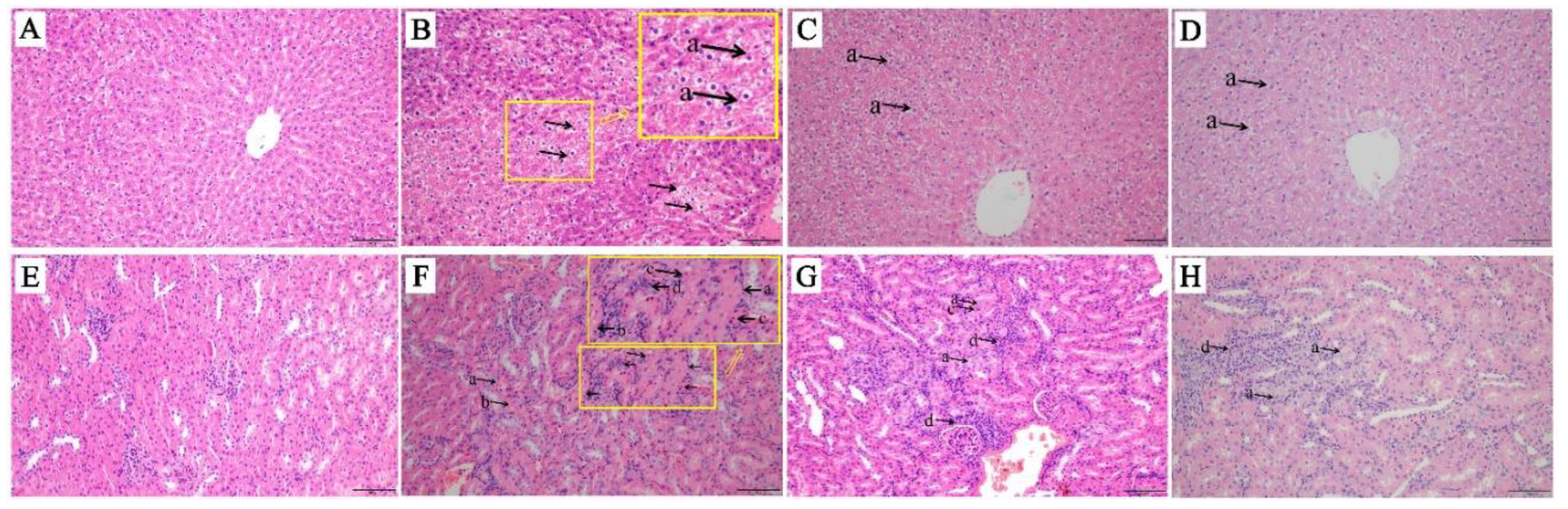
2.1. Tissue Histological Characteristics
2.2. Serum Biochemical Index Results
2.3. Examination of Concentration of AAΙ and ALΙ in Liver and Serum
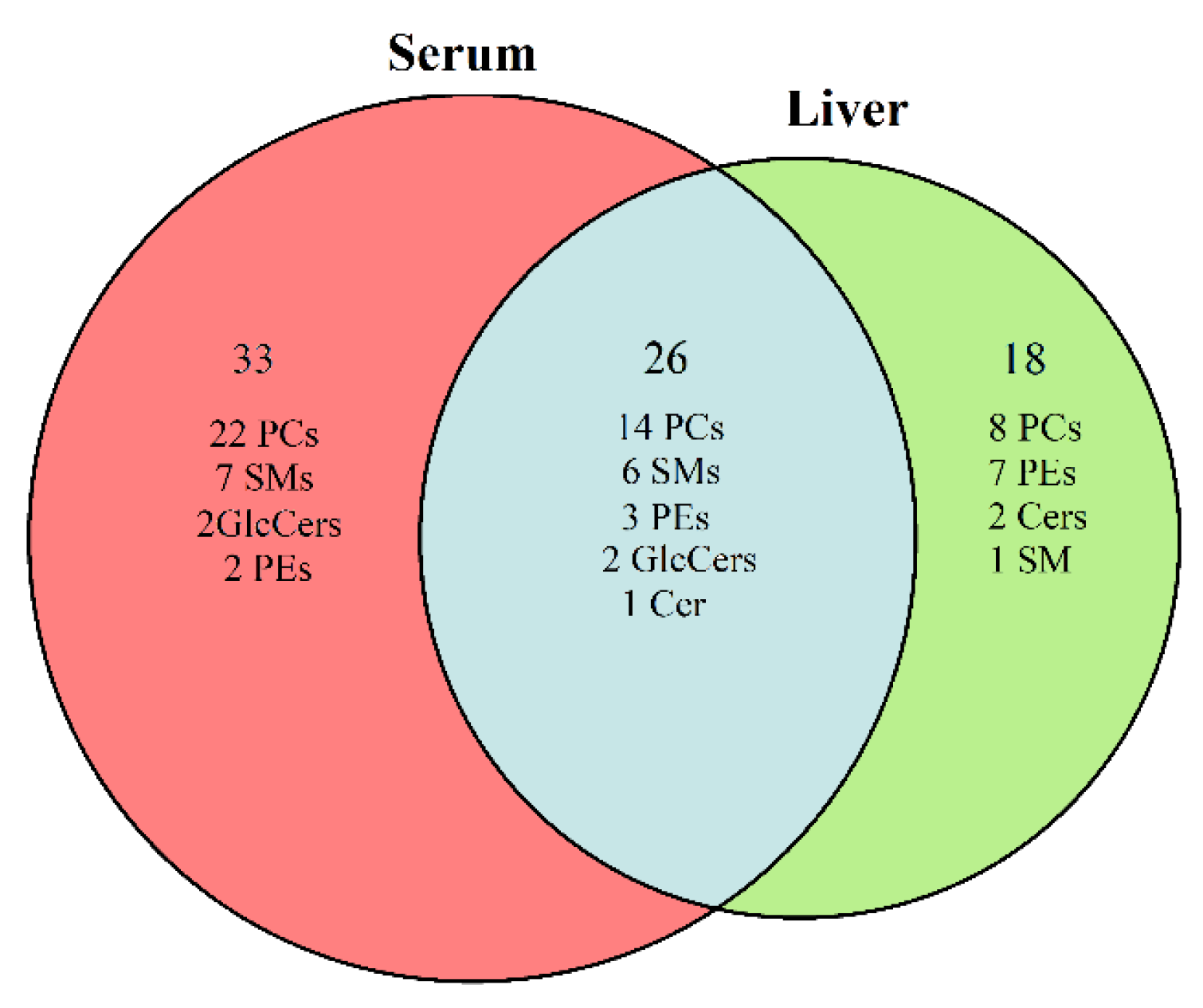
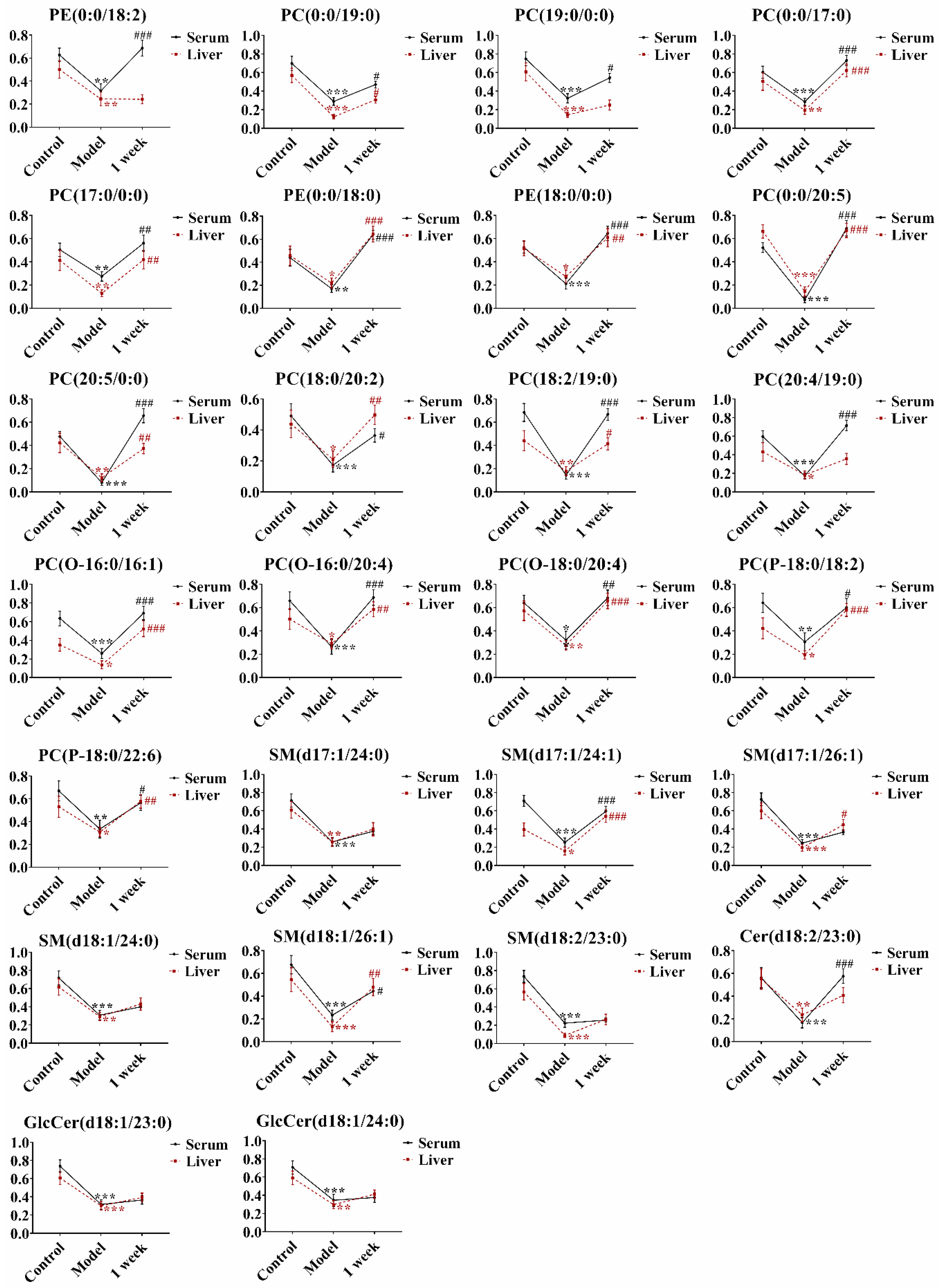
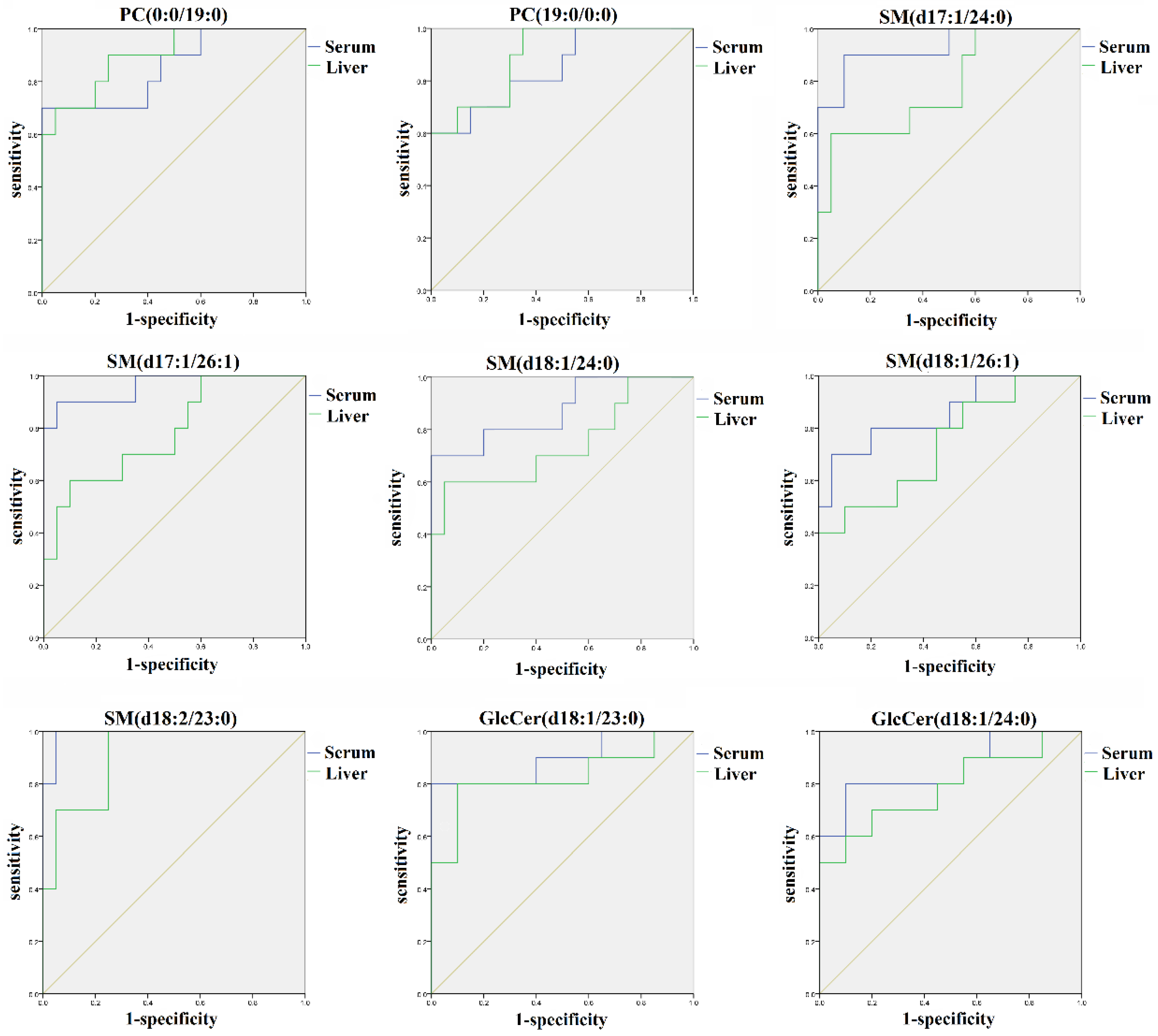
2.4. Identification of the Potential Lipid Biomarkers
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Animals and Sample Collection
3.3. Biochemical Analysis
3.4. Histological Evaluation
3.5. Sample Preparation for Lipidomics
3.5.1. Treatment of Serum Samples
3.5.2. Treatment of Liver Samples
3.6. Quantitative Analysis of AAI and ALI in Liver and Serum
3.6.1. Preparation of Stock Solutions, Calibration Standards, and Internal Standard Solutions
3.6.2. Preparation for the Serum and Liver Samples
3.6.3. UPLC-MS/MS Conditions
3.6.4. Analytical Validation
3.7. UPLC/Q-Exacitve/MS Analysis for Lipidomics
3.8. Data processing and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, J.; Zhang, L.; Jiang, Z.; Shu, B.; Li, F.; Bao, Q.; Zhang, L. Toxicities of aristolochic acid I and aristololactam I in cultured renal epithelial cells. Toxicol. In Vitro 2010, 24, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Z.; Bao, Q.L.; Sun, L.X.; Huang, X.; Wang, T.; Zhang, S.; Li, H.; Zhang, L.Y. Possible role of mtDNA depletion and respiratory chain defects in aristolochic acid I-induced acute nephrotoxicity. Toxicol. Appl. Pharmacol. 2013, 266, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wang, Y.; Jin, J.; Guan, C.; Li, M.; Xi, C.; Ouyang, Z.; Chen, M.; Qiu, Y.; Huang, M.; et al. Endoplasmic reticulum stress mediates aristolochic acid I-induced apoptosis in human renal proximal tubular epithelial cells. Toxicol. In Vitro 2012, 26, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Tsai, S.H.; Chen, S.M.; Chang, Y.M.; Huang, T.C.; Huang, Y.P.; Chang, C.T.; Lee, J.A. Aristolochic acid-induced accumulation of methylglyoxal and nepsilon-(carboxymethyl) lysine: An important and novel pathway in the pathogenic mechanism for aristolochic acid nephropathy. Biochem. Biophys. Res. Commun. 2012, 423, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Stiborová, M.; Arlt, V.M.; Schmeiser, H.H. DNA Adducts Formed by Aristolochic Acid Are Unique Biomarkers of Exposure and Explain the Initiation Phase of Upper Urothelial Cancer. Int. J. Mol. Sci. 2017, 18, 2144. [Google Scholar] [CrossRef]
- Lu, Z.N.; Luo, Q.; Zhao, L.N.; Shi, Y.; Wang, N.; Wang, L.; Han, Z.G. Mutational Features of Aristolochic Acid–Induced Mouse and Human Liver Cancers. Hepatology 2019. [Google Scholar] [CrossRef]
- Rolim, A.E.; Henrique-Araújo, R.; Ferraz, E.G.; de Araújo Alves Dultra, F.K.; Fernandez, L.G. Lipidomics in the study of lipid metabolism: Current perspectives in the omic sciences. Gene 2015, 554, 131–139. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Ogundare, M.; Williams, C.M.; Wang, Y. On the future of “omics”: Lipidomics. J. Inherit. Metab. Dis. 2011, 34, 583–592. [Google Scholar] [CrossRef]
- Han, X. Lipidomics: Developments and applications. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2009, 877, 2663. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, Y.; Chai, L.; Wang, H. Histological changes, lipid metabolism and oxidative stress in the liver of, Bufo gargarizans, exposed to cadmium concentrations. Chemosphere 2017, 179, 337–346. [Google Scholar] [CrossRef]
- Khattab, H.; Fouad, A.; Hamza, M.; Mohey, M.A.; El-Akel, W.; Ghoneim, H.; Abul-Fotouh, A.; Esmat, G. Relation of ALT and AST levels to the histopathological changes in liver biopsies of patients with chronic hepatitis C genotype 4. Arab. J. Gastroenterol. 2015, 16, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P.; Keach, J.C.; Batts, K.P.; Lindor, K.D. Independent predictors of liver fibrosis in patients with nonalcoholic steatohepatitis. Hepatology 1999, 30, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Lin, B.P.; Ngu, M.C.; Jones, D.B.; Katelaris, P.H. Aspartate aminotransferase: Alanine aminotransferase ratio in chronic hepatitis C infection: Is it a useful predictor of cirrhosis? J. Gastroenterol. Hepatol. 2000, 15, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.S. The clinical application value of biochemical test in the diagnosis of patients with cirrhosis. Guide China Med. 2016, 14, 148. [Google Scholar]
- Yang, Q.H. The clinical application value analysis of biochemical test in the diagnosis of liver cirrhosis. JACM 2018, 11, 47–48. [Google Scholar]
- Park, E.S.; Lee, J.H.; Hong, J.H.; Park, Y.K.; Lee, J.W.; Lee, W.J.; Lee, J.W.; Kim, K.P.; Kim, K.H. Phosphatidylcholine Alteration Identified Using MALDI Imaging MS in HBV-Infected Mouse Livers and Virus-Mediated Regeneration Defects. PLoS ONE 2014, 9, e103955. [Google Scholar] [CrossRef]
- Li, Z.; Guan, M.; Lin, Y.; Cui, X.; Zhang, Y.; Zhao, Z.; Zhu, J. Aberrant Lipid Metabolism in Hepatocellular Carcinoma Revealed by Liver Lipidomics. Int. J. Mol. Sci. 2017, 18, 2550. [Google Scholar] [CrossRef]
- Ming, Y.N.; Zhang, J.Y.; Wang, X.L.; Li, C.M.; Ma, S.C.; Wang, Z.Y.; Liu, X.L.; Li, X.B.; Mao, Y.M. Liquid chromatography mass spectrometry-based profiling of phosphatidylcholine and phosphatidylethanolamine in the plasma and liver of acetaminophen-induced liver injured mice. Lipids Health Dis. 2017, 16, 153. [Google Scholar] [CrossRef]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef]
- Zhao, L.L. UPLC-Q/TOF MS Analysis of GP Metabolism in RAW264.7 Cell Inflammation Model Interfering by NSAIDs; GDPU: Guangzhou, China, 2012. [Google Scholar]
- Na, J.Y.; Song, K.; Kim, S.; Kwon, J. Hepatoprotective effect of phosphatidylcholine against carbon tetrachloride liver damage in mice. Biochem. Biophys. Res. Commun. 2015, 460, 308–313. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Polizzi, A.; Guillou, H.; Loiseau, N. Sphingolipid metabolism in non-alcoholic fatty liver diseases. Biochimie 2019, 159, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Freeman, C.M.; Gulbins, E.; Lentsch, A.B. Sphingolipids in liver injury, repair and regeneration. Biol. Chem. 2015, 396, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Qu, F.; Zheng, S.J.; Ren, J.Y.; Wu, H.L.; Liu, M.; Liu, H.; Ren, F.; Chen, Y.; Zhang, J.L.; et al. Plasma sphingolipids as potential indicators of hepatic necroinflammation in patients with chronic hepatitis C and normal alanine aminotransferase level. PLoS ONE 2014, 9, e95095. [Google Scholar] [CrossRef] [PubMed]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, S91–S96. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Futerman, A.H. The metabolism and function of sphingolipids and glycosphingolipids. Cell Mol. Life Sci. 2007, 64, 2270–2284. [Google Scholar] [CrossRef] [PubMed]
- Breier, C.; Lisch, H.J.; Braunsteiner, H. Lipoproteins, HDL-apolipoproteins, activities of hepatic lipase and lecithin-cholesterol acyltransferase in the plasma of patients with post-alcoholic end-stage liver cirrhosis. Klin. Wochenschr. 1983, 61, 929–931. [Google Scholar] [CrossRef]
- Vere, C.C.; Streba, C.T.; Streba, L.; Rogoveanu, I. Rogoveanu, Lipid serum profile in patients with viral liver cirrhosis. Med. Princ. Pract. 2012, 21, 566–568. [Google Scholar] [CrossRef]
- Krautbauer, S.; Eisinger, K.; Wiest, R.; Liebisch, G.; Buechler, C. Systemic saturated lysophosphatidylcholine is associated with hepatic function in patients with liver cirrhosis. Prostaglandins Other Lipid Mediat. 2016, 124, 27–33. [Google Scholar] [CrossRef]
- Krautbauer, S.; Wiest, R.; Liebisch, G.; Buechler, C. Associations of systemic sphingolipids with measures of hepatic function in liver cirrhosis are related to cholesterol. Prostaglandins Other Lipid Mediat. 2017, 131, 25–32. [Google Scholar] [CrossRef]
- Margalit, M.; Shalev, Z.; Pappo, O.; Sklair-Levy, M.; Alper, R.; Gomori, M.; Engelhardt, D.; Rabbani, E.; Ilan, Y. Glucocerebroside ameliorates the metabolic syndrome in OB/OB mice. J. Pharmacol. Exp. Ther. 2006, 319, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, E.; Zangen, S.W.; Pappo, O.; Sklair-Levy, M.; Lalazar, G.; Zolotaryova, L.; Raz, I.; Ilan, Y. β-Glycosphingolipids improve glucose intolerance and hepatic steatosis of the Cohen diabetic rat. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E72–E78. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Zheng, S.J.; Wang, L.L.; Liu, S.; Ren, F.; Chen, Y.; Bai, L.; Liu, M.; Duan, Z.P. Glucosylceramide synthase regulates the proliferation and apoptosis of liver cells in vitro by Bcl-2/Bax pathway. Mol. Med. Rep. 2017, 16, 7355–7360. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.C.; Radin, N.S. Stimulation of liver growth and DNA synthesis by glucosylceramide. Lipids 1988, 23, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Marsh, N.L.; Elias, P.M.; Holleran, W.M. Glucosylceramides stimulate murine epidermal hyperproliferation. J. Clin. Investig. 1995, 95, 2903–2909. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Wang, H.L.; Cheng, X.L.; Wei, F.; Bai, X.; Lin, R.C.; Vaziri, N.D. Metabolomics analysis reveals the association between lipid abnormalities and oxidative stress, inflammation, fibrosis, and Nrf2 dysfunction in aristolochic acid-induced nephropathy. Sci Rep. 2015, 5, 12936. [Google Scholar] [CrossRef]
- Kulkarni, H.; Meikle, P.J.; Mamtani, M.; Weir, J.M.; Barlow, C.K.; Jowett, J.B.; Bellis, C.; Dyer, T.D.; Johnson, M.P.; Rainwater, D.L.; et al. Variability in associations of phosphatidylcholine molecular species with metabolic syndrome in Mexican-American families. Lipids 2013, 48, 497–503. [Google Scholar] [CrossRef]
- Syme, C.; Czajkowski, S.; Shin, J.; Abrahamowicz, M.; Leonard, G.; Perron, M.; Richer, L.; Veillette, S.; Gaudet, D.; Strug, L.; et al. Glycerophosphocholine metabolites and cardiovascular disease risk factors in adolescents: A cohort study. Circulation 2016, 134, 1629–1636. [Google Scholar] [CrossRef]
- Zeng, H.; Tong, R.; Tong, W.; Yang, Q.; Qiu, M.; Xiong, A.; Sun, S.; Ding, L.; Zhang, H.; Yang, L. Metabolic biomarkers for prognostic prediction of pre-diabetes: Results from a longitudinal cohort study. Sci. Rep. 2017, 7, 6575. [Google Scholar] [CrossRef]
- Chen, S.L.; Yin, P.Y.; Zhao, X.J.; Xing, W.B.; Hu, C.X.; Zhou, L.N.; Xu, G.W. Serum lipid profiling of patients with chronic hepatitis B, cirrhosis, and hepatocellular carcinoma by ultra fast LC/IT-TOF MS. Electrophoresis 2013, 34, 2848–2856. [Google Scholar] [CrossRef]
- Gui, S.X.; Li, B.Y.; Zhao, X.Y.; Sheng, L.; Hong, J.; Yu, X.H.; Sang, X.Z.; Sun, Q.Q.; Ze, Y.G.; Wang, L.; et al. Renal Injury and Nrf2 Modulation in Mouse Kidney Following Chronic Exposure to TiO2 Nanoparticles. J. Agric. Food Chem. 2013, 61, 8959. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ALT (U/L) | AST (U/L) | TBA (μmol/L) | ALB (g/L) | GLB (g/L) | CHE (U/L) |
| Control | 52 ± 6.07 | 93.25 ± 25.47 | 28.22 ± 17.41 | 31.53 ± 0.89 | 20.60 ± 0.48 | 102.75 ± 19.48 |
| AA | 64.14 ± 24.39 | 112.2 ± 34.19 | 30.16 ± 12.99 | 29.63 ± 0.69 ### | 19.87 ± 1.76 | 84.44 ± 17.30 # |
| 1 week | 100.63 ± 7.58 *** | 132.88 ± 27.05 | 35.77 ± 18.06 | 30.29 ± 1.27 | 18.82 ± 1.15 | 107.10 ± 17.80 ** |
| 2 week | 55.40 ± 7.04 | 107.22 ± 14.13 | 20.98 ± 11.88 | 31.71 ± 0.66 *** | 19.45 ± 0.90 | 117.00 ± 15.40 *** |
| Group | TP (g/L) | LDH (U/L) | Crea (μmol/L) | BUN (mmol/L) | UA (μmol/L) | |
| Control | 52.43 ± 1.71 | 367.71 ± 127.57 | 15.78 ± 1.72 | 4.82 ± 0.45 | 27.63 ± 8.28 | |
| AA | 48.54 ± 1.42 ### | 643.00 ± 315.26 ## | 19.80 ± 6.03 # | 5.19 ± 0.94 | 28.17 ± 19.16 | |
| 1 week | 49.05 ± 1.60 | 349.89 ± 133.01 ** | 21.50 ± 6.09 | 5.37 ± 0.63 | 15.89 ± 7.72 | |
| 2 week | 51.46 ± 1.82 *** | 305.67 ± 96.07 ** | 22.33 ± 1.75 | 5.54 ± 0.78 | 14.00 ± 4.12* |
| Analyte | m/z | DP (V) | CE (V) | |
|---|---|---|---|---|
| Precursor | Product | |||
| AAΙ | 359.1 | 298.8 | 66 | 15 |
| ALΙ | 294.1 | 279.2 | 146 | 39 |
| Buspirone (IS) | 386.2 | 122.2 | 91 | 49 |
| Analyte | Equation | Correlation Coefficient (r) | Linear Range (ng/mL) | LOQ (ng/mL) |
|---|---|---|---|---|
| AAΙ | y = 0.000512 x + 0.000337 | 0.9954 | 5–1000 | 5 |
| ALΙ | y = 0.000425 x − 0.000135 | 0.9928 | 5–1000 | 5 |
| Analyte | Equation | Correlation Coefficient (r) | Linear Range (ng/mL) | LOQ (ng/mL) |
|---|---|---|---|---|
| AAΙ | y = 0.000403x + 0.000775 | 0.9993 | 5–1000 | 5 |
| ALΙ | y = 0.00045 x + 0.000265 | 0.9937 | 5–1000 | 5 |
| Analyte | Serum | Serum | Serum | Serum | Serum | Serum | Mean ± SEM |
| AAΙ | 75.1 | 67.6 | 65.5 | 81.5 | 55.5 | 52.4 | 66.27 ± 4.55 |
| ALΙ | - | - | - | - | - | - | - |
| Analyte | Liver | Liver | Liver | Liver | Liver | Liver | Mean ± SEM |
| AAΙ | 98.6 | 50.2 | 97.0 | 95.8 | 62.5 | 79.4 | 80.58 ± 8.32 |
| ALΙ | 42.7 | 61.2 | 32.5 | 37.4 | 86.4 | 68.3 | 54.75 ± 8.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Yang, Y.; Wang, H.; Bian, B.; Yang, J.; Wei, X.; Zhou, Y.; Si, N.; Zhao, H. The Disturbance of Hepatic and Serous Lipids in Aristolochic Acid Ι Induced Rats for Hepatotoxicity Using Lipidomics Approach. Molecules 2019, 24, 3745. https://doi.org/10.3390/molecules24203745
Zhou J, Yang Y, Wang H, Bian B, Yang J, Wei X, Zhou Y, Si N, Zhao H. The Disturbance of Hepatic and Serous Lipids in Aristolochic Acid Ι Induced Rats for Hepatotoxicity Using Lipidomics Approach. Molecules. 2019; 24(20):3745. https://doi.org/10.3390/molecules24203745
Chicago/Turabian StyleZhou, Junyi, Yifei Yang, Hongjie Wang, Baolin Bian, Jian Yang, Xiaolu Wei, Yanyan Zhou, Nan Si, and Haiyu Zhao. 2019. "The Disturbance of Hepatic and Serous Lipids in Aristolochic Acid Ι Induced Rats for Hepatotoxicity Using Lipidomics Approach" Molecules 24, no. 20: 3745. https://doi.org/10.3390/molecules24203745
APA StyleZhou, J., Yang, Y., Wang, H., Bian, B., Yang, J., Wei, X., Zhou, Y., Si, N., & Zhao, H. (2019). The Disturbance of Hepatic and Serous Lipids in Aristolochic Acid Ι Induced Rats for Hepatotoxicity Using Lipidomics Approach. Molecules, 24(20), 3745. https://doi.org/10.3390/molecules24203745