UHPLC-HR-MS/MS-Guided Recovery of Bioactive Flavonol Compounds from Greco di Tufo Vine Leaves

 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
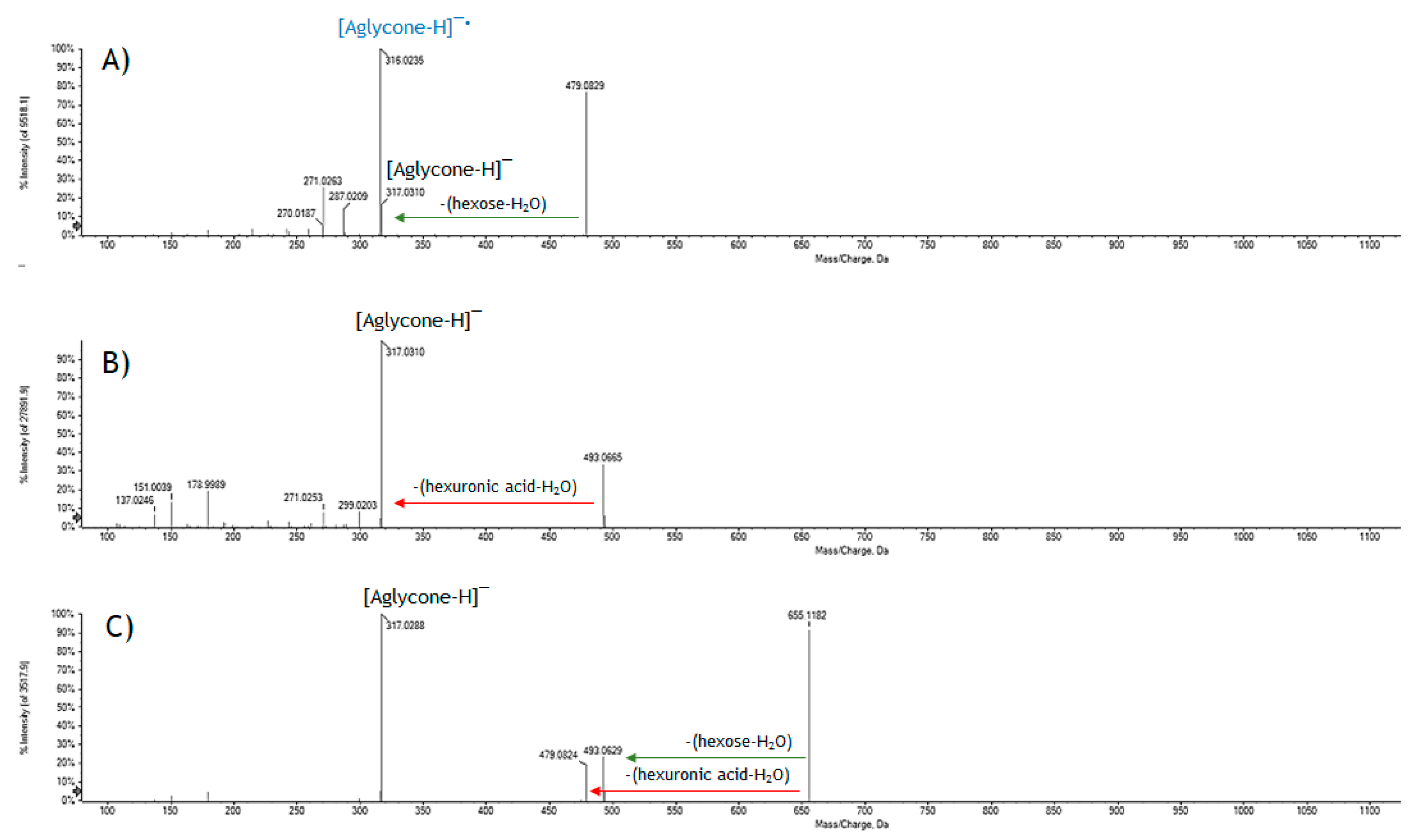
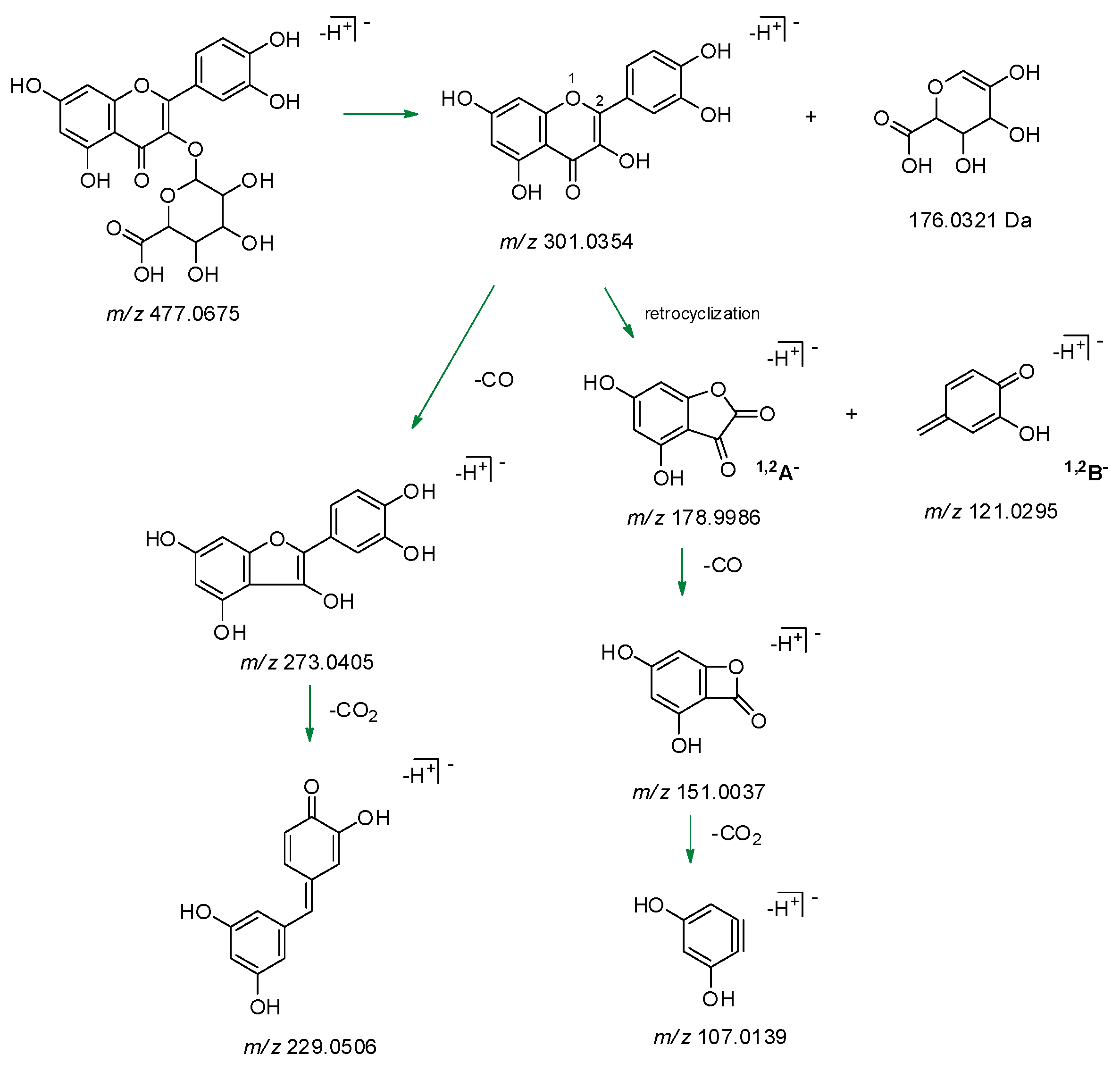
2.1. GrM Chemical Composition
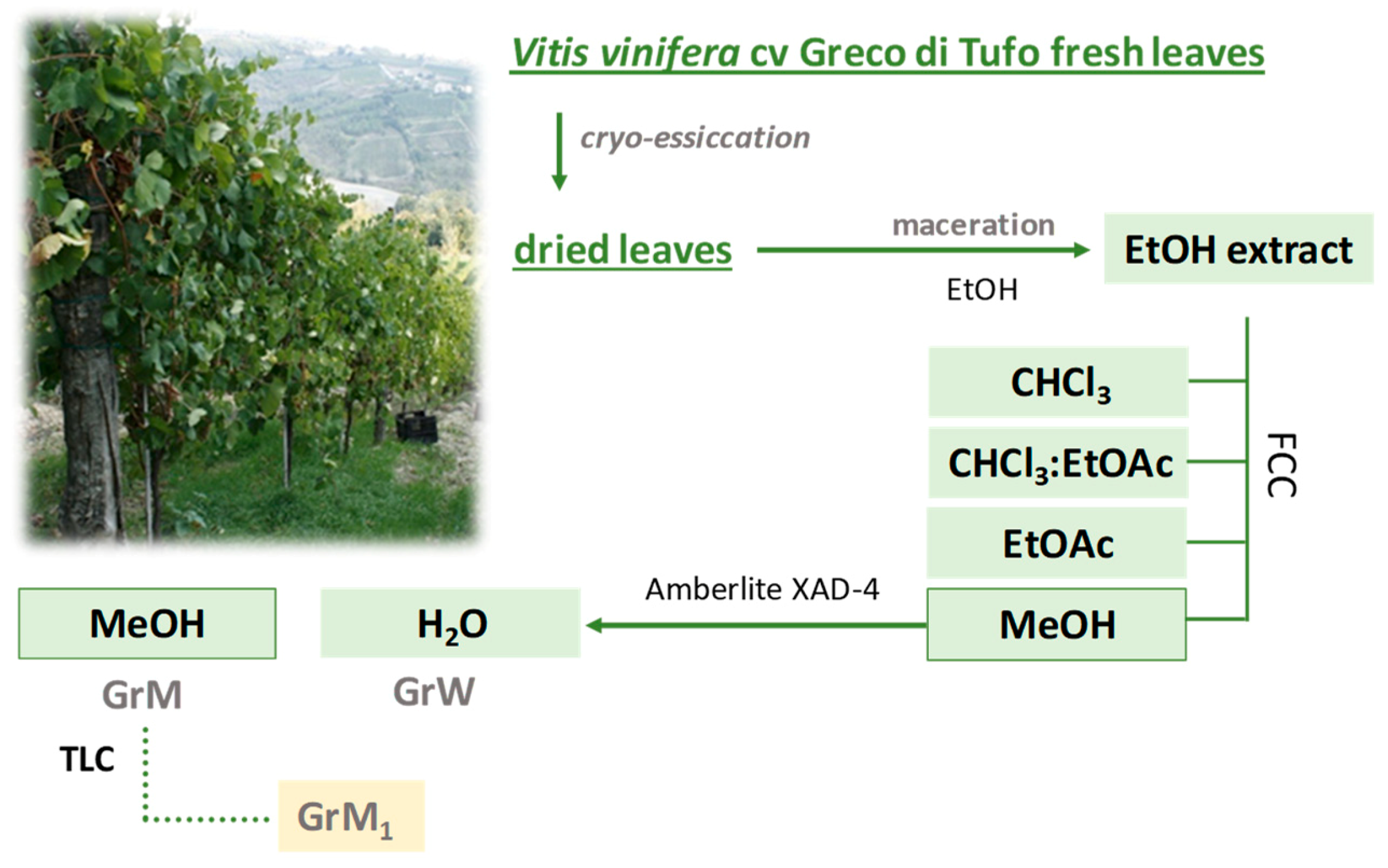
2.2. GrM1 Purification
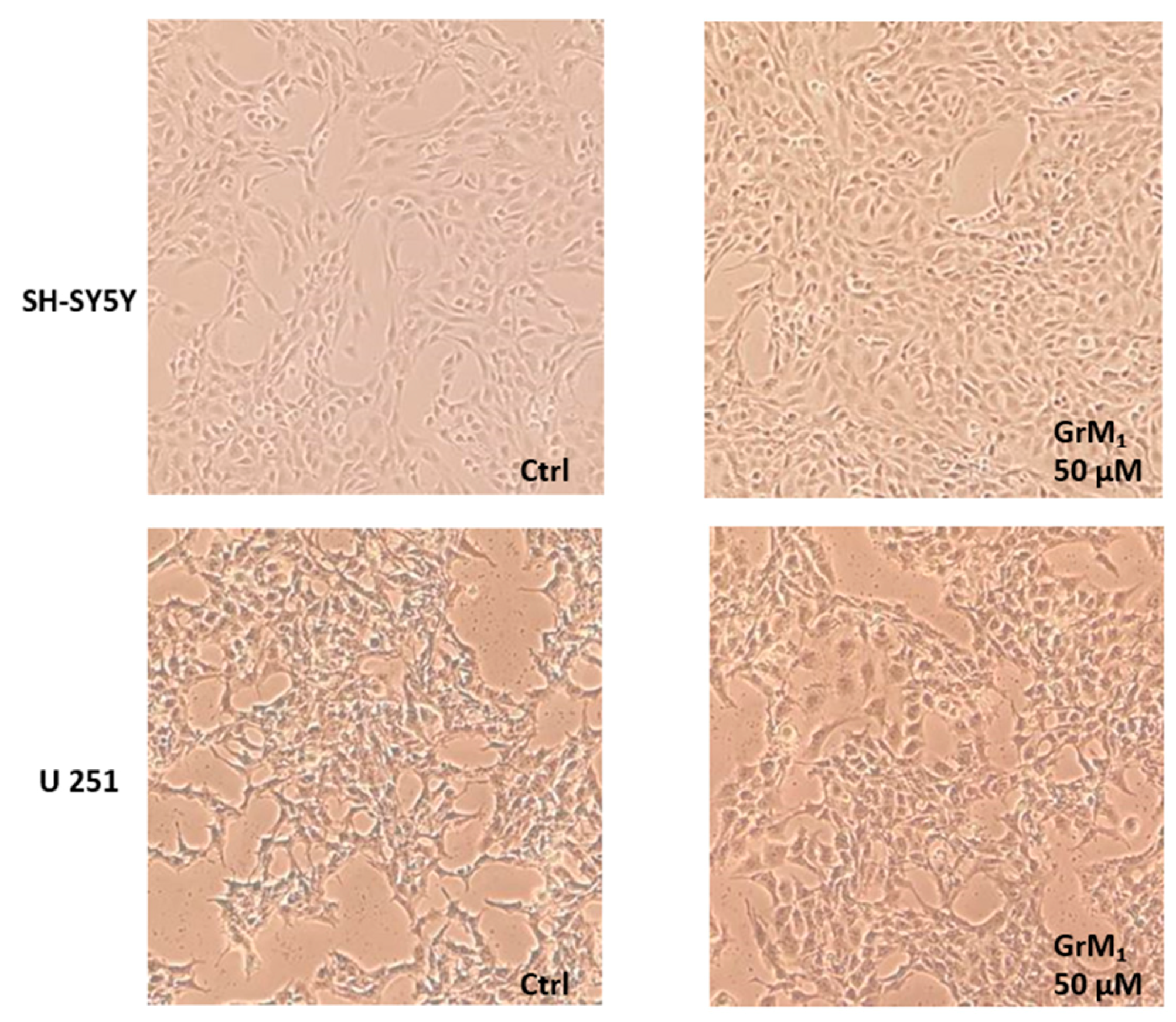
2.3. Cytotoxicity of GrM
2.4. Cytotoxicity of GrM1
3. Materials and Methods
3.1. Plant Extraction and Fractionation
3.2. UHPLC-HR-MS and UV–Vis Analyses
3.3. Cell Culture and Cytotoxicity Assessment
3.3.1. MTT (3-(4, 5-Dimethylthiazolyl-2)-2,5-Diphenyltetrazolium Bromide) Cell Viability Test
3.3.2. SRB (Sulforhodamine B) Cell Viability Test
3.3.3. Anti-Acetylcholinesterase (AChE) Activity Assay
3.3.4. Cell Metabolomic Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Parfitt, J.; Barthel, M.; Macnaughton, S. Food waste within food supply chains: Quantification and potential for change to 2050. Philos. Trans. Royal Soc. B 2010, 465, 3065–3081. [Google Scholar] [CrossRef] [PubMed]
- Piccolella, S.; Bianco, A.; Crescente, G.; Santillo, A.; Chieffi Baccari, G.; Pacifico, S. Recovering Cucurbita pepo cv. ‘Lungo Fiorentino’ wastes: UHPLC-HRMS/MS metabolic profile, the basis for establishing their nutra- and cosmeceutical valorisation. Molecules 2019, 24, 1479. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Lopez Garcia, I.; Kookos, I.K.; Papanikolaou, S.; Kwanb, T.H.; Sze Ki Lin, C. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef] [PubMed]
- Varricchio, E.; Coccia, E.; Orso, G.; Lombardi, V.; Imperatore, R.; Vito, P.; Paolucci, M. Influence of polyphenols from olive mill wastewater on the gastrointestinal tract, alveolar macrophages and blood leukocytes of pigs. Ital. J. Anim. Sci. 2019, 18, 574–586. [Google Scholar] [CrossRef]
- Parrillo, L.; Coccia, E.; Volpe, M.G.; Siano, F.; Pagliarulo, C.; Scioscia, E.; Varricchio, E.; Safari, O.; Eroldogan, T.; Paolucci, M. Olive mill wastewater-enriched diet positively affects growth, oxidative and immune status and intestinal microbiota in the crayfish, Astacus leptodactylus. Aquaculture 2017, 473, 161–168. [Google Scholar] [CrossRef]
- Michalska, A.; Wojdyło, A.; Majerska, J.; Lech, K.; Brzezowska, J. Qualitative and quantitative evaluation of heat-induced changes in polyphenols and antioxidant capacity in Prunus domestica L. by-products. Molecules 2019, 24, 3008. [Google Scholar] [CrossRef] [PubMed]
- Ramón-Gonçalves, M.; Gómez-Mejía, E.; Rosales-Conrado, N.; León-González, M.E.; Madrid, Y. Extraction, identification and quantification of polyphenols from spent coffee grounds by chromatographic methods and chemometric analyses. Waste Manag. 2019, 96, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Galanakis, C.M. Recovery of high added-value components from food wastes: Conventional, emerging technologies and commercialized applications. Food. Sci. Technol. 2012, 26, 68–87. [Google Scholar] [CrossRef]
- Volpe, M.G.; Santagata, G.; Coccia, E.; Di Stasio, M.; Malinconico, M.; Paolucci, M. Pectin based pellets for crayfish aquaculture: Structural and functional characteristics and effects on redclaw Cherax quadricarinatus performances. Aquac. Nutr. 2014, 21, 814–823. [Google Scholar] [CrossRef]
- Brezoiu, A.M.; Matei, C.; Deaconu, M.; Stanciuc, A.M.; Trifan, A.; Gaspar-Pintiliescu, A.; Berger, D. Polyphenols extract from grape pomace. Characterization and valorisation through encapsulation into mesoporous silica-type matrices. Food Chem. Toxicol. 2019, 133, 110787. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, M.; Shi, T.; Guo, C.; Huang, Y.; Chen, Y.; Xie, M. Recovery of dietary fiber and polyphenol from grape juice pomace and evaluation of their functional properties and polyphenol compositions. Food Funct. 2017, 8, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Federici, F.; Fava, F.; Kalogerakis, N.; Mantzavinos, D. Valorisation of agro- industrial by- products, effluents and waste: Concept, opportunities and the case of olive mill wastewaters. J. Chem. Technol. Biot. 2009, 84, 895–900. [Google Scholar] [CrossRef]
- Rondeau, P.; Gambier, F.; Jolibert, F.; Brosse, N. Compositions and chemical variability of grape pomaces from French vineyard. Ind. Crop. Prod. 2013, 43, 251–254. [Google Scholar] [CrossRef]
- Torres, J.L.; Varela, B.; García, M.T.; Carilla, J.; Matito, C.; Centelles, J.J.; Cascante, M.; Sort, X.; Bobet, R. Valorization of grape (Vitis vinifera) byproducts. Antioxidant and biological properties of polyphenolic fractions differing in procyanidin composition and flavonol content. J. Agric. Food Chem. 2002, 50, 7548–7555. [Google Scholar] [CrossRef] [PubMed]
- Cosme, F.; Pinto, T.; Vilela, A. Oenology in the kitchen: The sensory experience offered by culinary dishes cooked with alcoholic drinks, grapes and grape leaves. Beverages 2017, 3, 42. [Google Scholar] [CrossRef]
- Bell, D.R.; Gochenaur, K. Direct vasoactive and vasoprotective properties of anthocyanin-rich extracts. J. Appl. Physiol. (1985) 2006, 100, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Xu, B.; Hu, M. Regioselective glucuronidation of flavonols by six human UGT1A isoforms. Pharm. Res. 2011, 28, 1905–1918. [Google Scholar] [CrossRef][Green Version]
- Pacifico, S.; D’Abrosca, B.; Scognamiglio, M.; Gallicchio, M.; Galasso, S.; Monaco, P.; Fiorentino, A. Antioxidant polyphenolic constituents of Vitis × labruscana cv. ‘Isabella’. Open Nat. Prod. J. 2013, 6, 5–11. [Google Scholar] [CrossRef][Green Version]
- Faugno, S.; Piccolella, S.; Sannino, M.; Principio, L.; Crescente, G.; Baldi, G.M.; Fiorentino, N.; Pacifico, S. Can agronomic practices and cold-pressing extraction parameters affect phenols and polyphenols content in hempseed oils? Ind. Crop. Prod. 2019, 130, 511–519. [Google Scholar] [CrossRef]
- Fabre, N.; Rustan, I.; de Hoffmann, E.; Quetin-Leclercq, J. Determination of flavone, flavonol, and flavanone aglycones by negative ion liquid chromatography electrospray ion trap mass spectrometry. J. Am. Soc. Mass Spectrom. 2001, 12, 707–715. [Google Scholar] [CrossRef]
- Singh, R.; Wu, B.; Tang, L.; Liu, Z.; Hu, M. Identification of the position of mono-O-glucuronide of flavones and flavonols by analyzing shift in online UV spectrum (lambdamax) generated from an online diode array detector. J. Agric Food Chem. 2010, 58, 9384–9395. [Google Scholar] [CrossRef] [PubMed]
- Docampo, M.; Olubu, A.; Wang, X.; Pasinetti, G.; Dixon, R.A. Glucuronidated flavonoids in neurological protection: Structural analysis and approaches for chemical and biological synthesis. J. Agric. Food Chem. 2017, 65, 7607–7623. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Mingo-Chornet, H.; Pérez-Alonso, J.J.; Di Paola-Naranjo, R.; González-Paramás, A.M.; Santos-Buelga, C. Preparation of quercetin glucuronides and characterization by HPLC–DAD–ESI/MS. Eur. Food Res. Technol. 2008, 227, 1069–1076. [Google Scholar] [CrossRef]
- Gao, C.; Zhou, Y.; Li, H.; Cong, X.; Jiang, Z.; Wang, X.; Cao, R.; Tian, W. Antitumor effects of baicalin on ovarian cancer cells through induction of cell apoptosis and inhibition of cell migration in vitro. Mol. Med. Rep. 2017, 16, 8729–8734. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gao, Y.; Wu, J.; Chen, Y.; Chen, B.; Hu, J.; Zhou, J. Exploring therapeutic potentials of baicalin and its aglycone baicalein for hematological malignancies. Cancer Lett. 2014, 354, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Piccolella, S.; Nocera, P.; Carillo, P.; Woodrow, P.; Greco, V.; Manti, L.; Fiorentino, A.; Pacifico, S. An apolar Pistacia lentiscus L. leaf extract: GC-MS metabolic profiling and evaluation of cytotoxicity and apoptosis inducing effects on SH-SY5Y and SK-N-BE(2)C cell lines. Food Chem. Toxicol. 2016, 95, 64–74. [Google Scholar] [CrossRef]
- Pacifico, S.; Piccolella, S.; Lettieri, A.; Nocera, P.; Bollino, F.; Catauro, M. A metabolic profiling approach to an Italian sage leaf extract (SoA541) defines its antioxidant and anti-acetylcholinesterase properties. J. Funct. Foods 2017, 29, 1–9. [Google Scholar] [CrossRef]
- Sak, K.; Illes, P. Neuronal and glial cell lines as model systems for studying P2Y receptor pharmacology. Neurochem. Int. 2005, 47, 401–412. [Google Scholar] [CrossRef]
- Araújo, K.C.F.; Costa, E.M.M.B.; Pazini, F.; Valadares, M.C.; Oliveira, V. Bioconversion of quercetin and rutin and the cytotoxicity activities of the transformed products. Food Chem. Toxicol. 2013, 51, 93–96. [Google Scholar] [CrossRef]
- Pacifico, S.; Piccolella, S.; Papale, F.; Nocera, P.; Lettieri, A.; Catauro, M. A polyphenol complex from Thymus vulgaris L. plants cultivated in the Campania Region (Italy): New perspectives against neuroblastoma. J. Funct. Foods 2016, 20, 253–266. [Google Scholar] [CrossRef]
- Pacifico, S.; Gallicchio, M.; Lorenz, P.; Duckstein, S.M.; Potenza, N.; Galasso, S.; Marciano, S.; Fiorentino, A.; Stintzing, F.C.; Monaco, P. Neuroprotective potential of Laurus nobilis antioxidant polyphenol enriched leaf extracts. Chem. Res. Toxicol. 2014, 27, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Dietmar, S.; Timmins, N.E.; Gray, P.P.; Nielsen, L.K.; Kromer, J.O. Towards quantitative metabolomics of mammalian cells: Development of a metabolite extraction protocol. Anal. Biochem. 2010, 404, 155–164. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min) | Tentative Assignment | Formula | [M − H]– Found (m/z) | [M − H]– calc. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) | |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.308 | Gallic acid hexoside | C13H16O10 | 331.0666 | 331.0671 | –1.3 | 6 | 169.014; 125.0248 |
| 2 | 3.129 | Benzyl O-[arabinofuranosyl-(1-6)-glucoside] | C18H26O10 | 401.1460 437.1232 [M + Cl]– | 401.1453 | 1.7 | 6 | 269.1030; 193.0486; 101.0238; 85.0279 |
| 3 | 4.204 | Lariciresinol hexoside | C26H34O11 | 521.2043 | 521.2028 | 2.8 | 10 | 359.1507; 344.1273; 329.1035; 313.1078; 299.0936; 255.0657; 255.0687; 241.0509 |
| 4 | 5.086 | Myricetin derivative | C23H16O14 | 515.0456 | 515.0467 | –2.2 | 16 | 339.0125; 317.0300; 316.0225; 271.0225; 178.9976; 151.0018 |
| 5 | 5.091 | Myricetin hexoside 1 | C21H20O13 | 479.0840 | 479.0831 | 1.8 | 12 | 479.0829; 317.0310; 316.0235; 287.0209; 271.0263; 270.0187; 259.0268; 214.0280 |
| 6 | 5.094 | Myricetin hexuronide | C21H18O14 | 493.0630 | 493.0624 | 1.3 | 13 | 493.0665; 317.0310 (178.9980; 151.0042; 137.0243; 109.0291); 299.0203; 271.0253; 227.0347; 178.9989; 151.0039; 137.0246 |
| 7 | 5.224 | Myricetin hexoside 2 | C21H20O13 | 479.0841 | 479.0831 | 2.1 | 12 | 317.0310; 316.0228; 287.0202; 271.0253; 270.0185; 259.0248; 242.0219; 214.0291; 178.9968; 151.0043 |
| 8 | 5.323 | Furan/piran linalool oxide pentosyl hexoside | C21H36O11 | 463.2198 509.2253 [M + FA]– | 463.2185 | 2.8 | 4 | 463.0896; 331.1754; 161.0447; 101.0240; 85.0289 |
| 9 | 5.424 | Myricetin hexosyl hexuronide | C27H28O19 | 655.1143 | 655.1152 | –1.4 | 14 | 655.1182; 493.0629; 479.0824; 317.0288 |
| 10 | 5.580 | Caffeic acid derivative | C22H36O12 | 491.2190 | 491.2134 | 5.9 | 5 | 329.1618; 227.1272; 101.0242 |
| 11 | 5.731 | Hydroxygeraniol pentosyl hexoside | C21H38O11 | 465.2353 501.2113 [M + Cl]– | 465.2341 | 2.5 | 3 | 251.0794; 191.0578; 149.0447; 131.0352; 101.0241; 89.0246 |
| 12 | 6.270 | Rutin | C27H30O16 | 609.1486 | 609.1461 | 4.1 | 13 | 609.1498; 301.0343; 300.0268; 271.0227; 255.0285 |
| 13 | 6.288 | Quercetin hexuronide | C21H18O13 | 477.0685 955.1420 [2M – H]– | 477.0675 | 3.2 | 13 | 477.0690; 301.0351; 283.0244; 273.0399; 255.0293; 245.0450; 211.0395; 178.9979; 151.0029; 121.0297; 107.0136 |
| 14 | 6.494 | Quercetin hexoside | C21H20O12 | 463.0896 | 463.0882 | 3 | 12 | 463.0896; 301.0345; 300.0268; 271.0236; 255.0281; 243.0281; 227.0327; 178.9926; 151.0017 |
| 15 | 7.998 | Kaempferol rutinoside | C27H30O15 | 593.1534 | 593.1512 | 3.7 | 13 | 593.1523; 285.0383; 284.0305; 255.0278; 229.0497; 227.0326 |
| 16 | 8.187 | Kaempferol hexuronide | C21H18O12 | 461.0738 | 461.0725 | 2.7 | 13 | 461.0757; 285.0406; 257.0452; 229.0503 |
| 17 | 8.722 | Isorhamnetin rutinoside | C28H32O16 | 623.1620 | 623.1618 | 0.4 | 13 | 623.1643; 315.0492; 314.0421; 300.0258; 271.0233 |
| 18 | 9.305 | Isorhamnetin hexuronide | C22H20O13 | 491.0845 | 491.0831 | 2.8 | 13 | 491.0834; 315.0482; 301.0333; 300.0250; 271.0221; 255.0271; 243.0271 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piccolella, S.; Crescente, G.; Volpe, M.G.; Paolucci, M.; Pacifico, S. UHPLC-HR-MS/MS-Guided Recovery of Bioactive Flavonol Compounds from Greco di Tufo Vine Leaves. Molecules 2019, 24, 3630. https://doi.org/10.3390/molecules24193630
Piccolella S, Crescente G, Volpe MG, Paolucci M, Pacifico S. UHPLC-HR-MS/MS-Guided Recovery of Bioactive Flavonol Compounds from Greco di Tufo Vine Leaves. Molecules. 2019; 24(19):3630. https://doi.org/10.3390/molecules24193630
Chicago/Turabian StylePiccolella, Simona, Giuseppina Crescente, Maria Grazia Volpe, Marina Paolucci, and Severina Pacifico. 2019. "UHPLC-HR-MS/MS-Guided Recovery of Bioactive Flavonol Compounds from Greco di Tufo Vine Leaves" Molecules 24, no. 19: 3630. https://doi.org/10.3390/molecules24193630
APA StylePiccolella, S., Crescente, G., Volpe, M. G., Paolucci, M., & Pacifico, S. (2019). UHPLC-HR-MS/MS-Guided Recovery of Bioactive Flavonol Compounds from Greco di Tufo Vine Leaves. Molecules, 24(19), 3630. https://doi.org/10.3390/molecules24193630