Polyphenol Composition and (Bio)Activity of Berberis Species and Wild Strawberry from the Argentinean Patagonia

 , and
, and
Abstract
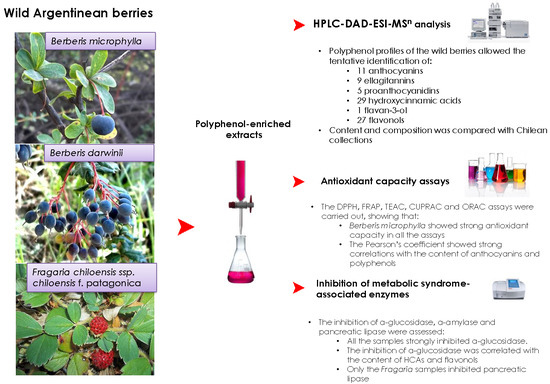
:
1. Introduction
2. Results and Discussion
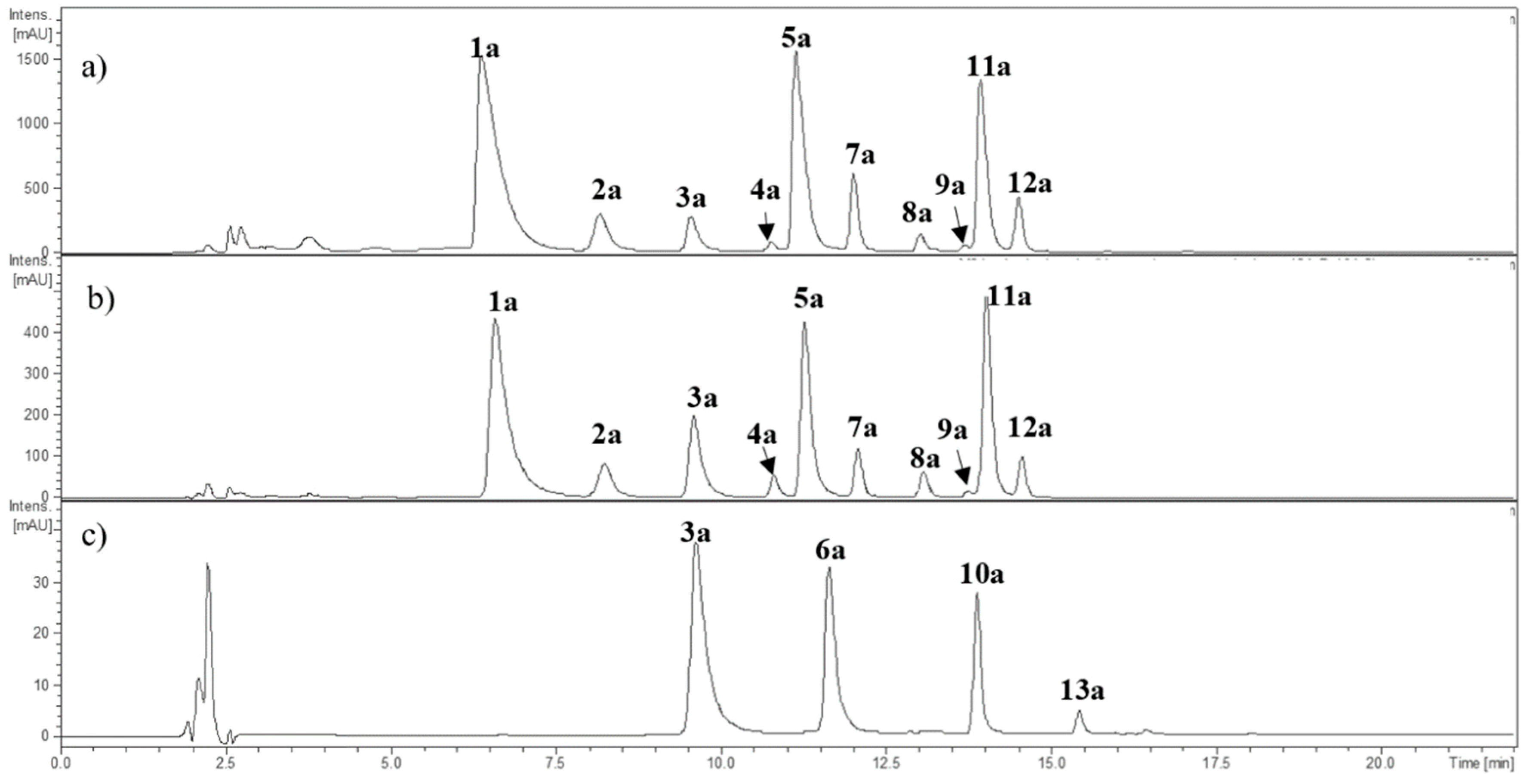
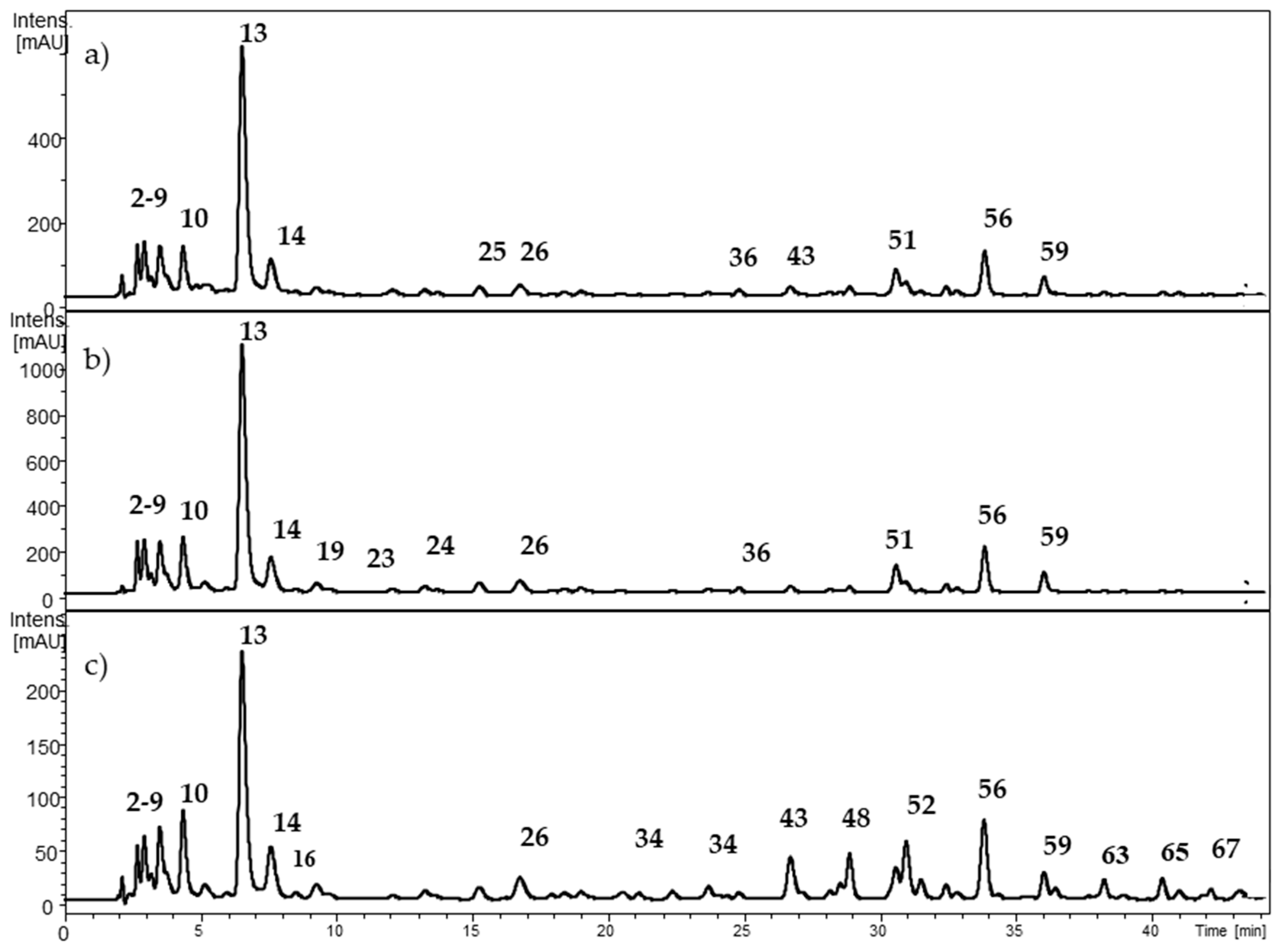
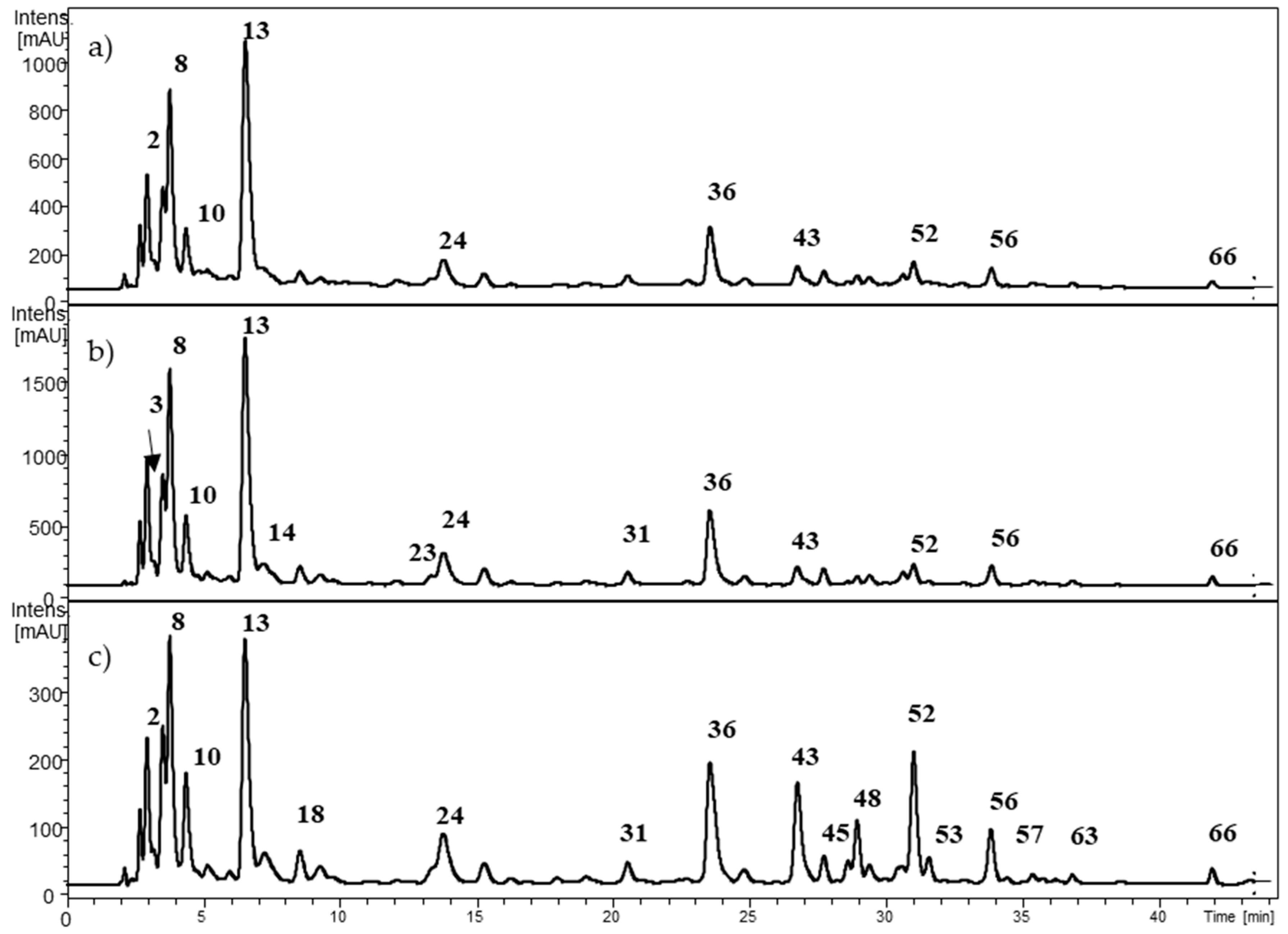
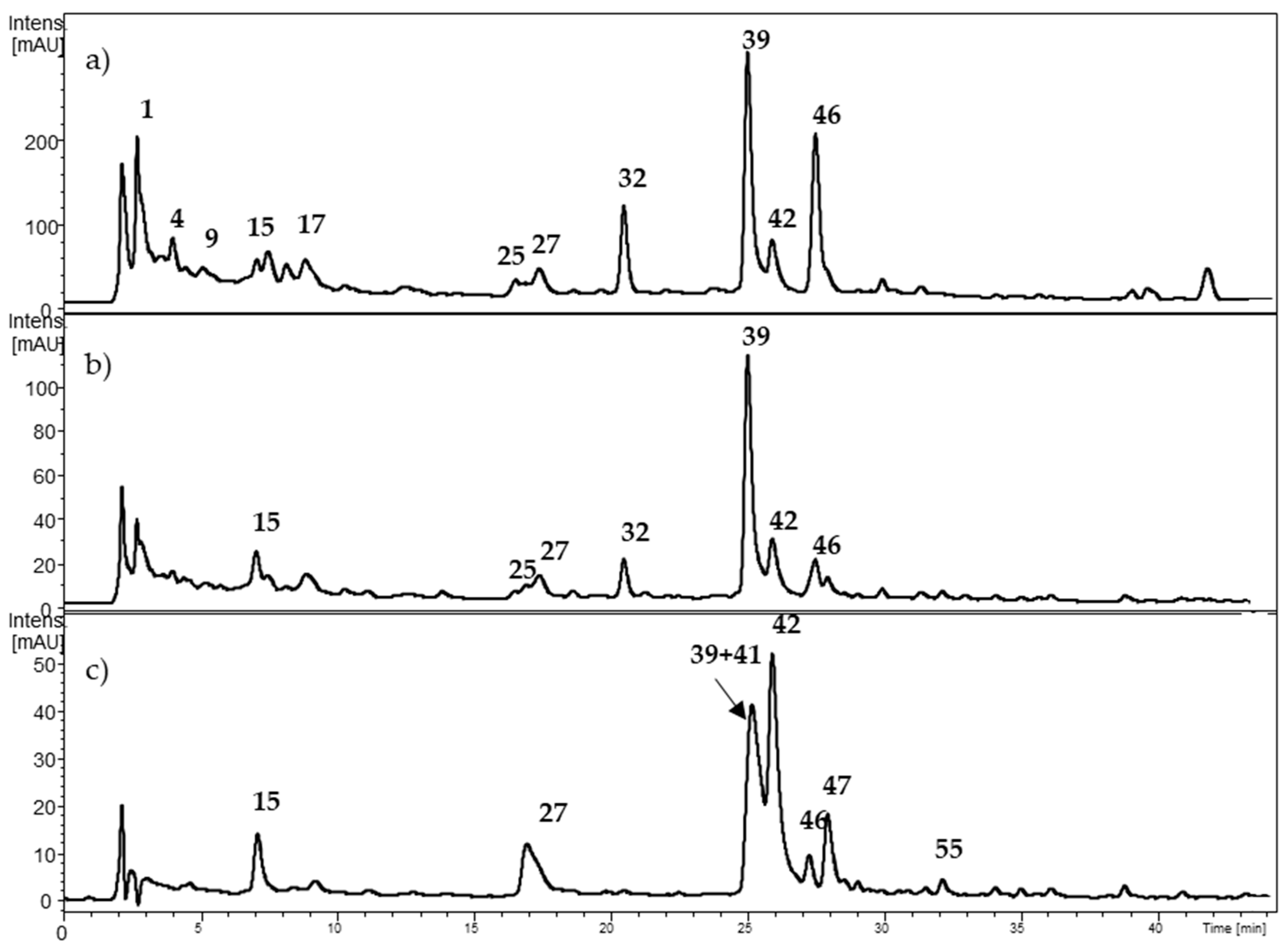
2.1. Characterization of Individual Components by High Performance Liquid Chromatography Coupled to Diode Array Detector and Electrospray Ionization Mass Spectrometry (HPLC-DAD-ESI-MSn)
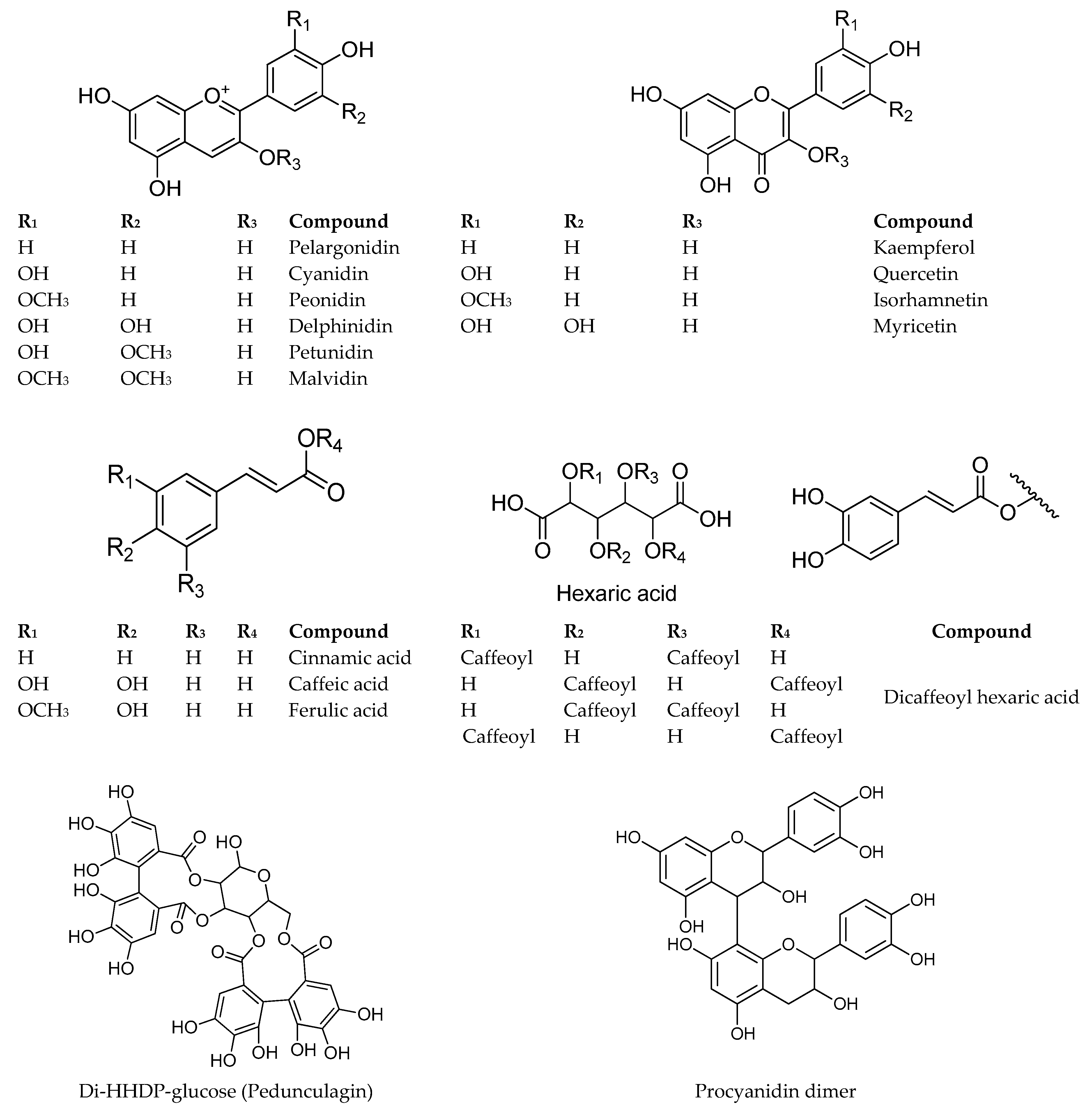
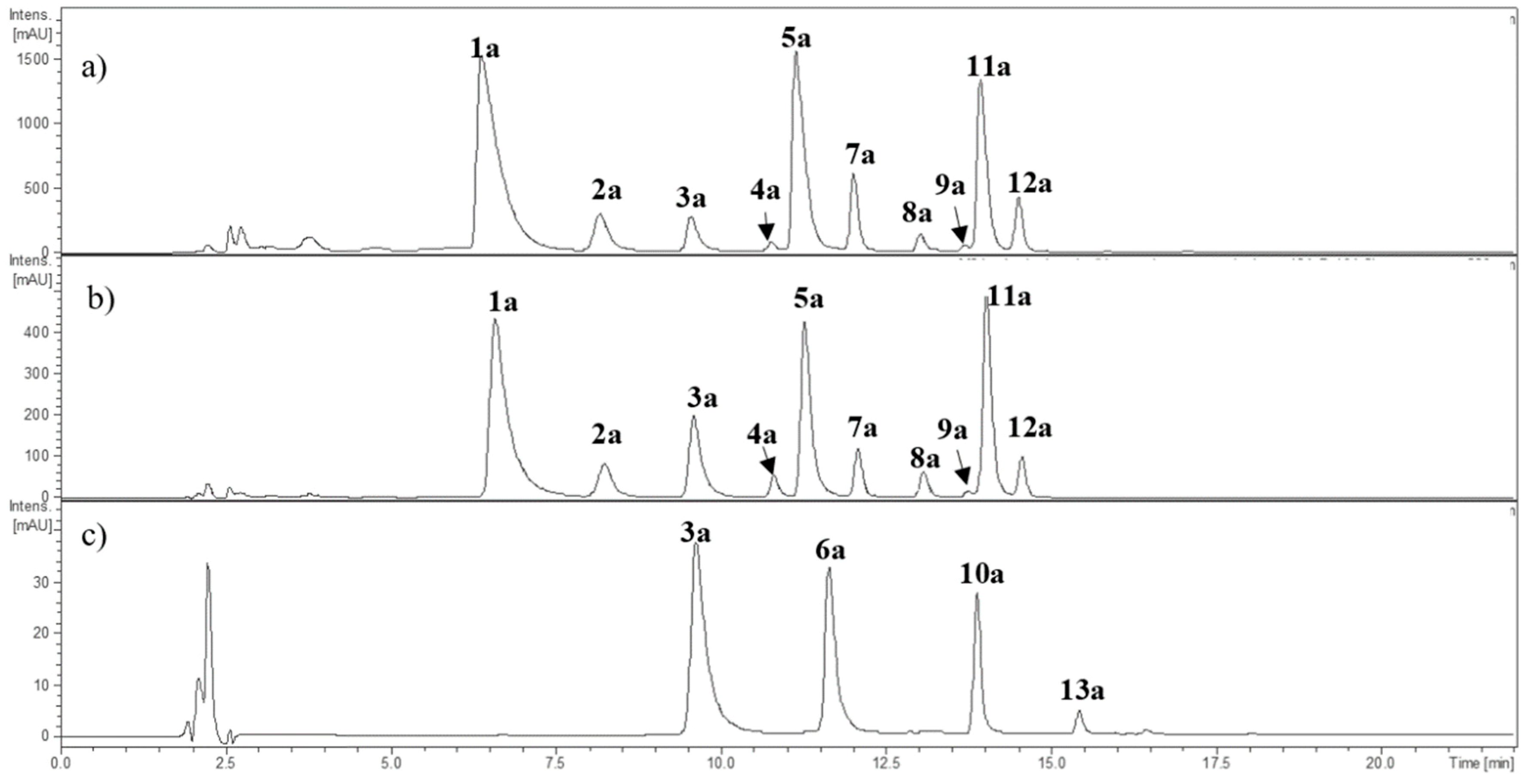
2.1.1. Anthocyanins
2.1.2. Hydroxycinnamic Acids (HCAs)
2.1.3. Ellagitannins
2.1.4. Flavan-3-ols and Proanthocyanidins
2.1.5. Flavonols
2.2. Quantification of Main Phenolics
2.3. Antioxidant Activity
2.4. Inhibition of Metabolic Syndrome-Associated Enzymes
3. Materials and Methods
3.1. Chemicals
3.2. Sample Collection
3.3. Anthocyanin and Non-Anthocyanin Polyphenol Fractionation
3.4. HPLC-DAD-ESI-MSn Analysis
3.5. Antioxidant Capacity Assays
3.6. Inhibition of Metabolic Syndrome-Associated Enzymes
3.7. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bvenura, C.; Sivakumar, D. The role of wild fruits and vegetables in delivering a balanced and healthy diet. Food Res. Int. 2017, 99, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Garcia, S.N.; Guevara-Gonzalez, R.G.; Miranda-López, R.; Feregrino-Perez, A.A.; Torres-Pacheco, I.; Vazquez-Cruz, M.A. Functional properties and quality characteristics of bioactive compounds in berries: Biochemistry, biotechnology, and genomics. Food Res. Int. 2013, 54, 1195–1207. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Jiménez-Aspee, F.; Theoduloz, C.; Ladio, A. Patagonian berries as native food and medicine. J. Ethnopharmacol. 2019, 241, 111979. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, M.F.; Ladio, A.H.; Molares, S. Patagonian Berries: An ethnobotanical approach to exploration of their nutraceutical potential. In Ethnobotany: Local Knowledge and Traditions, 1st ed.; Martínez, J., Muñoz-Acevedo, A., Rai, M., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 50–69. [Google Scholar]
- Speisky, H.; López-Alarcón, C.; Gómez, M.; Fuentes, J.; Sandoval-Acuña, C. First web-based database on total phenolics and oxygen radical absorbance capacity (ORAC) of fruits produced and consumed within the south Andes region of South America. J. Agric. Food Chem. 2012, 60, 8851–8859. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.E.; Lencinas, M.V.; Radice, S. Variability in floral traits and reproductive success among and within populations of Berberis microphylla G. Forst., an underutilized fruit species. Sci. Hortic. 2018, 241, 65–73. [Google Scholar] [CrossRef]
- Abbasi, A.M.; Shah, M.H.; Li, T.; Fu, X.; Guo, X.; Liu, R.H. Ethnomedicinal values, phenolic contents and antioxidant properties of wild culinary vegetables. J. Ethnopharmacol. 2015, 162, 333–345. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, M.; Misra, A.; Pandey, G.; Rawat, A. A review on biological and chemical diversity in Berberis (Berberidaceae). EXCLI J. 2015, 14, 247–267. [Google Scholar]
- Mokhber-Dezfuli, N.; Saeidnia, S.; Gohari, A.; Kurepaz-Mahmoodabadi, M. Phytochemistry and pharmacology of berberis species. Pharmacogn. Rev. 2014, 8, 8–15. [Google Scholar]
- Gundogdu, M. Determination of antioxidant capacities and biochemical compounds of Berberis vulgaris L. Fruits. Adv. Environ. Biol. 2013, 7, 344–348. [Google Scholar]
- Hassanpour, H.; Alizadeh, S. Evaluation of phenolic compound, antioxidant activities and antioxidant enzymes of barberry genotypes in Iran. Sci. Hortic. 2016, 200, 125–130. [Google Scholar] [CrossRef]
- Ersoy, N.; Kupe, M.; Sagbas, H.I.; Ercisli, S. Physicochemical diversity among barberry (Berberis vulgaris L.) fruits from Eastern Anatolia. Not. Bot. Hort. Agrobot. Cluj-Napoca. 2018, 46, 336–342. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosín-Guitérrez, I.; Mardones, C.; Vergara, C.; Herlitz, E.; Vega, M.; Dorau, C.; Winterhalter, P.; von Baer, D. Polyphenols and antioxidant activity of calafate (Berberis microphylla) fruits and other native berries from southern Chile. J. Agric. Food Chem. 2010, 58, 6081–6089. [Google Scholar] [CrossRef]
- Ramirez, J.E.; Zambrano, R.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC-HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Ladio, A.H.; Lozada, M. Patterns of use and knowledge of wild edible plants in distinct ecological environments: A case study of a Mapuche community from northwestern Patagonia. Biodivers. Conserv. 2004, 13, 1153–1173. [Google Scholar] [CrossRef]
- Cheel, J.; Theoduloz, C.; Rodríguez, J.A.; Caligari, P.D.S.; Schmeda-Hirschmann, G. Free radical scavenging activity and phenolic content in achenes and thalamus from Fragaria chiloensis ssp. chiloensis, F. vesca and F. x ananassa cv. Chandler. Food Chem. 2007, 102, 36–44. [Google Scholar] [CrossRef]
- Jiménez-Aspee, F.; Thomas-Valdés, S.; Schulz, A.; Ladio, A.; Theoduloz, C.; Schmeda-Hirschmann, G. Antioxidant activity and phenolic profiles of the wild currant Ribes magellanicum from Chilean and Argentinean Patagonia. Food Sci. Nutr. 2016, 4, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Aspee, F.; Theoduloz, C.; Ávila, F.; Thomas-Valdés, S.; Mardones, C.; von Baer, D.; Schmeda-Hirschmann, G. The Chilean wild raspberry (Rubus geoides Sm.) increases intracellular GSH content and protects against H2O2 and methylglyoxal-induced damage in AGS cells. Food Chem. 2016, 194, 908–919. [Google Scholar] [CrossRef]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin profiles in south Patagonian wild berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Ruiz, A.; Mardones, C.; Vergara, C.; Hermosín-Gutiérrez, I.; von Baer, D.; Hinrichsen, P.; Rodríguez, R.; Arribillaga, D.; Domínguez, E. Analysis of hydroxycinnamic acids derivatives in calafate (Berberis microphylla G. Forst) berries by liquid chromatography with photodiode array and mass spectrometry detection. J. Chromatogr. A 2013, 1281, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Bustamante, L.; Vergara, C.; von Baer, D.; Hermosín-Gutiérrez, I.; Obando, L.; Mardones, C. Hydroxycinnamic acids and flavonols in native edible berries of South Patagonia. Food Chem. 2015, 167, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Aspee, F.; Theoduloz, C.; Soriano, M.D.P.C.; Ugalde-Arbizu, M.; Alberto, M.R.; Zampini, I.C.; Isla, M.I.; Simirgiotis, M.J.; Schmeda-Hirschmann, G. The native fruit Geoffroea decorticans from arid northern Chile: Phenolic composition, antioxidant activity and in vitro inhibition of pro-inflammatory and metabolic syndrome-associated enzymes. Molecules 2017, 22, 1565. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Mardones, C.; Vergara, C.; von Baer, D.; Gómez-Alonso, S.; Gómez, M.V.; Hermosín-Gutiérrez, I. Isolation and structural elucidation of anthocyanidin 3,7-β-O-diglucosides and caffeoyl-glucaric acids from calafate berries. J. Agric. Food Chem. 2014, 62, 6918–6925. [Google Scholar] [CrossRef] [PubMed]
- Lopes da Silva, F.; Escribano-Bailón, M.T.; Pérez Alonso, J.J.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanin pigments in strawberry. LWT-Food Sci. Technol. 2007, 40, 374–382. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Theoduloz, C.; Caligari, P.D.S.; Schmeda-Hirschmann, G. Comparison of phenolic composition and antioxidant properties of two native Chilean and one domestic strawberry genotypes. Food Chem. 2009, 113, 377–385. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for the LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Cheel, J.; Theoduloz, C.; Rodríguez, J.A.; Saud, G.; Caligari, P.D.S.; Schmeda-Hirschmann, G. E-cinnamic acid derivatives and phenolics from Chilean strawberry fruits, Fragaria chiloensis ssp. chiloensis. J. Agric. Food Chem. 2005, 53, 8512–8518. [Google Scholar] [CrossRef]
- Schuster, B.; Herrmann, K. Hydroxybenzoic and hydroxycinnamic acid derivatives in soft fruits. Phytochemistry 1985, 24, 2761–2764. [Google Scholar] [CrossRef]
- Jiménez-Aspee, F.; Theoduloz, C.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I.; Reyes, M.; Schmeda-Hirschmann, G. Polyphenolic profile and antioxidant activity of meristem and leaves from “chagual” (Puya chilensis Mol.), a salad from central Chile. Food Res. Int. 2018, 114, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Quatrin, A.; Pauletto, R.; Maurer, L.H.; Minuzzi, N.; Nichelle, S.M.; Carvalho, J.F.C.; Maróstica Junior, M.R.; Rodrigues, E.; Bochi, V.C.; Emanuelli, T. Characterization and quantification of tannins, flavonols, anthocyains and matrix-bound polyphenols from jaboticaba fruit peel: A comparison between Myrciaria trunciflora and M. jaboticaba. J. Food Compos. Anal. 2019, 78, 59–74. [Google Scholar] [CrossRef]
- Thomas-Valdés, S.; Theoduloz, C.; Jiménez-Aspee, F.; Schmeda-Hirschmann, G. Effect of simulated gastrointestinal digestion on polyphenols and bioactivity of the native Chilean red strawberry (Fragaria chiloensis ssp. chiloensis f. patagonica). Food Res. Int. 2019, 23, 106–114. [Google Scholar]
- McDougall, G.J.; Kulkarni, N.N.; Stewart, D. Berry polyphenols inhibit pancreatic lipase activity in vitro. Food Chem. 2009, 115, 193–199. [Google Scholar] [CrossRef]
- Lin, L.Z.; Sun, J.; Chen, P.; Monagas, M.J.; Harnly, J.M. UHPLC-PDA-ESI/HRMSn profiling method to identify and quantify oligomeric proanthocyanidins in plant products. J. Agric. Food Chem. 2014, 62, 9387–9400. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Yu, D.K.; Lee, H.J. Changes in anthocyanidin and anthocyanin pigments in highbush blueberry (Vaccinium corymbosum cv. Bluecrop) fruits during ripening. Hort. Environ. Biotechnol. 2016, 57, 424–430. [Google Scholar] [CrossRef]
- Parra-Palma, C.; Fuentes, E.; Palomo, I.; Torres, C.A.; Moya-León, M.A.; Ramos, P. Linking the platelet anti-aggregation effect of different strawberries species with antioxidants: Metabolomic and transcript profiling of polyphenols. BLACPMA 2018, 17, 36–52. [Google Scholar]
- Xu, C.; Zhang, Y.; Zhu, L.; Huang, Y.; Lu, J. Influence of growing season on phenolic compounds and antioxidant properties of grape berries from vines grown in subtropical climate. J. Agric. Food Chem. 2011, 59, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.E. Estudio de algunos fenómenos morfofisiológicos y cambios bioquímicos en Berberis microphylla G. Forst (sinónimo B. buxifolia) asociados a la formación y maduración de frutos en Tierra del Fuego y su relación con la producción de metabolitos útiles. Ph.D. Thesis, Universidad Nacional del Sur, Bahía Blanca, Argentina, 2016. [Google Scholar]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Mykkänen, H.; Niskanen, L. Postprandial glucose, insulin, and free fatty acid responses to sucrose consumed with blackcurrants and lingonberries in healthy women. Am. J. Clin. Nutr. 2012, 96, 527–533. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Vasquez, K.; Fuentes, F.; Ovalle-Marin, A.; Parra-Ruiz, C.; Zamora, O.; Pino, M.T.; Quitral, V.; Jimenez, P.; Garcia, L.; et al. Extracts of Chilean native fruits inhibit oxidative stress, inflammation and insulin-resistance linked to the pathogenic interaction between adipocytes and macrophages. J. Funct. Foods 2016, 27, 69–83. [Google Scholar] [CrossRef]
- Thomas-Valdés, S.; Theoduloz, C.; Jiménez-Aspee, F.; Burgos-Edwards, A.; Schmeda-Hirschmann, G. Changes in polyphenol composition and bioactivity of the native Chilean white strawberry (Fragaria chiloensis spp. chiloensis f. chiloensis) after in vitro gastrointestinal digestion. Food Res. Int. 2018, 105, 10–18. [Google Scholar]
- Podsędek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Koziołkiewicz, M. In vitro inhibitory effect on digestive enzymes and antioxidant potential of commonly consumed fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Favre, G.; González-Neves, G.; Piccardo, D.; Gómez-Alonso, S.; Pérez-Navarro, J.; Hermosín-Gutierrez, I. New acylated flavonols identified in Vitis vinifera grapes and wines. Food Res. Int. 2018, 112, 98–107. [Google Scholar] [CrossRef] [PubMed]
- ICH. Validation of Analytical Procedures: Text and Methodology. 2005. Available online: https://www.ich.org/fileadmin/Public_Web_Site/ICH_Products/Guidelines/Quality/Q2_R1/Step4/Q2_R1__Guideline.pdf (accessed on 21 July 2019).
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH free radical method. LWT-Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Nenadis, N.; Wang, L.F.; Tsimidou, M.; Zhang, H.Y. Estimation of scavenging activity of phenolic compounds using the ABTS·+ assay. J. Agric. Food Chem. 2014, 5, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the polyphenol-enriched extracts are available from the authors at the Laboratorio de Productos Naturales, Instituto de Quimica de Recursos Naturales, Universidad de Talca, Chile. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Yield of Extraction (%w/w) | Moisture (%w/w) | DPPH (SC50, µg/mL) | FRAP (µmol TE/g PEE) | TEAC (µM TE/g PEE) | CUPRAC (µmol TE/g PEE) | ORAC (µmol TE/g PEE) | α-glucosidase (IC50, µg/mL) |
|---|---|---|---|---|---|---|---|---|
| Berberis microphylla | ||||||||
| Brazo Rincón | 9.2 | 72.1 | 16.0 ± 0.4 a | 1915.0 ± 61.0 a | Inactive | 2876.3 ± 78.7 a | 843.8 ± 43.0 a | 0.5 ± 0.0 a |
| Aeropuerto | 8.4 | 54.5 | 10.4 ± 0.3 b | 2656.6 ± 43.7 b | 1938.8 ± 58.8 a | 3355.0 ± 103.7 b | 3020.0 ± 99.3 b | 0.4 ± 0.0 d |
| Villa La Angostura | 9.9 | 58.4 | 10.4 ± 0.4 b | 2579.6 ± 45.5 b | 1565.8 ± 33.3 b | 3470.7 ± 19.0 b | 2800.8 ± 90.7 b | 0.3 ± 0.0 b |
| Cuyín Manzano | 7.3 | 53.0 | 10.9 ± 0.4 b | 2578.0 ± 36.8 b | 1760.2 ± 44.3 c | 3318.0 ± 73.2 b | 3299.8 ± 69.7 c | 0.1 ± 0.0 c |
| Llanquín | 8.7 | 57.5 | 15.3 ± 0.4 a | 2009.7 ± 46.3 a | 1748.3 ± 46.2 c | 3015.4 ± 96.4 a | 2928.0 ± 94.1 b | 0.4 ± 0.0 d |
| Berberis darwinii | ||||||||
| Brazo Rincón | 11.4 | 74.6 | 17.3 ± 0.2 | 2770.6 ± 76.8 | 2012.5 ± 39.8 | 4588.9 ± 157.4 | 3032.3 ± 60.3 | 0.5 ± 0.0 |
| Villa La Angostura | 12.3 | 72.8 | 8.8 ± 0.2 | 3014.6 ± 63.0 | 2280.0 ± 44.8 | 4937.6 ± 89.4 | 2160.3 ± 164.4 | 0.8 ± 0.0 |
| Fragaria chiloensis | ||||||||
| Arroyo Llodcondo | 6.1 | 83.1 | 10.1 ± 0.3 | 1064.8 ± 15.4 | 1484.8 ± 29.1 | 2292.8 ± 27.3 | 772.5 ± 0.9 | 0.3 ± 0.0 |
| Frey | 5.3 | 82.0 | 8.4 ± 0.1 | 1722.4 ± 36.8 | 1234.8 ± 40.1 | 2744.4 ± 59.7 | 811.0 ± 2.6 | 1.2 ± 0.2 |
| Reference compounds | ||||||||
| Quercetin | - | - | 7.8 ± 0.3 | 1077.2 ± 16.4 | 8157.9 ± 22.1 | 27,526.1 ± 97.5 | 22,561.6 ± 808.8 | - |
| Acarbose | - | - | - | - | - | - | - | 137.7 ± 1.3 |
| Orlistat | - | - | - | - | - | - | - | - |
| Peak | Rt (min) | [M + H]+ | MS2 | λmax (nm) | Tentative Identification | Detected in | ||
|---|---|---|---|---|---|---|---|---|
| B. microphylla | B. darwinii | F. chiloensis | ||||||
| 1a | 6.7 | 465.3 | 302.9 (100) | 523 | Delphinidin-3-O-glucoside * | x | x | |
| 2a | 8.2 | 611.4 | 302.9 (100) | 525 | Delphinidin rutinoside | x | x | |
| 3a | 9.7 | 449.1 | 286.9 (100) | 516 | Cyanidin-3-O-glucoside * | x | x | x |
| 4a | 10.9 | 595.4 | 286.9 (100) | 521 | Cyanidin-3-O-rutinoside * | x | x | |
| 5a | 11.3 | 479.3 | 316.8 (100) | 525 | Petunidin hexoside | x | x | |
| 6a | 11.6 | 433.3 | 271.1 (100) | 502 | Pelargonidin hexoside | x | ||
| 7a | 12.1 | 625.5 | 316.9 (100) | 527 | Petunidin rutinoside | x | x | |
| 8a | 13.1 | 463.3 | 300.9 (100) | 518 | Peonidin hexoside | x | x | |
| 9a | 13.8 | 609.6 | 300.9 (100) | 526 | Peonidin rutinoside | x | x | |
| 10a | 13.9 | 535.1 | 286.9 (100) | 517 | Cyanidin manlonyl hexoside | x | ||
| 11a | 14.1 | 493.3 | 331.0 (100) | 527 | Malvidin-3-O-glucoside * | x | x | |
| 12a | 14.6 | 639.4 | 331.0 (100) | 530 | Malvidin rutinoside | x | x | |
| 13a | 15.5 | 519.1 | 271.1 (100) | 505 | Pelargonidin-malonyl hexoside | x | ||
| Peak | Rt (min) | [M − H]− | MS2 | λmax (nm) | Tentative Identification | Detected in | ||
|---|---|---|---|---|---|---|---|---|
| B. microphylla | B. darwinii | F. chiloensis | ||||||
| 1 | 2.8 | 783.3 | 480.8 (41), 300.7 (100) | 280 | Di-HHDP-glucose (Pedunculagin) | x | ||
| 2 | 3.0 | 371.0 | 208.6 (100), 190.7 (100) | 326 | Caffeoylhexaric acid isomer1 | x | x | |
| 3 | 3.6 | 353.3 | 190.6 (100), 178.9 (26), 134.9 (7) | 328 | 5-Caffeoylquinic acid * | x | x | |
| 4 | 3.7 | 577.3 | 558.7 (18), 450.7 (43) 424.8 (100), 289.1 (16) | 274 | Procyanidin B-type dimer 1 | x | ||
| 5 | 3.8 | 370.9 | 208.6 (100), 190.7 (26) | 327 | Caffeoylhexaric acid isomer 2 | x | x | |
| 6 | 4.0 | 633.9 | 450.7 (6), 300.7 (100) | 270 | HHDP-galloyl-hexose 1 | x | ||
| 7 | 4.2 | 577.4 | 450.6 (41), 424.7 (100), 288.7 (36) | 280 | Procyanidin B-type dimer 2 | x | ||
| 8 | 4.5 | 353.2 | 190.6 (100), 178.8 (5), 128.7 (3) | 329 | 3-Caffeoylquinic acid * | x | x | |
| 9 | 5.1 | 289.3 | 244.7 (100), 204.7 (41), 124.7 (7) | 282 | (epi)-catechin | x | x | |
| 10 | 5.3 | 353.3 | 190.6 (100) | 320 | Caffeoylquinic acid isomer 1 | x | x | |
| 11 | 5.7 | 337.3 | 162.6 (100), 118.8 (5) | 330 | 3-p-Coumaroylquinic acid | x | ||
| 12 | 6.1 | 577.2 | 558.8 (14), 450.7 (35), 424.9 (100), 407.4 (58), 289.2 (11) | 281 | Procyanidin B-type dimer 3 | x | x | |
| 13 | 6.5–6.9 | 352.9 | 190.6 (100) | 324 | Caffeoylquinic acid isomer 2 | x | x | |
| 14 | 7.6 | 533.1 | 370.6 (100), 208.7 (12) | 324 | Dicaffeoylhexaric acid isomer 1 | x | ||
| 15 | 7.7 | 935.0 | 632.9 (100), 300.8 (25) | 270 | Casuarictin/Potentillin isomer 1 | x | ||
| 16 | 8.6 | 353.3 | 190.7 (100) | 330 | Caffeoylquinic acid isomer 3 | x | x | |
| 17 | 9.0 | 934.9 | 632.7 (100), 450.9 (7), 300.8 (25) | 270 | Casuarictin/Potentillin isomer 2 | x | ||
| 18 | 9.3 | 367.3 | 179.7 (100) | 328 | Feruloylquinic acid isomer 1 | x | x | |
| 19 | 9.4 | 533.5 | 370.7 (100), 208.7 (18) | 326 | Dicaffeoylhexaric acid isomer 2 | x | x | |
| 20 | 10.5 | 577.0 | 558.7 (100), 450.7 (35) 424.7 (100), 406.9 (62) 288.7 (21) | 280 | Procyanidin B-type dimer 4 | x | ||
| 21 | 11.2 | 367.2 | 192.6 (22), 178.7 (100) 134.9 (32) | 330 | Feruloylquinic acid isomer 2 | x | ||
| 22 | 12.0 | 337.4 | 190.7 (100) | 330 | 5-p-Coumaroylquinic acid | x | ||
| 23 | 12.2 | 353.1 | 190.7 (100) | 320 | Caffeoylquinic acid isomer 4 | x | ||
| 24 | 13.9 | 533.1 | 370.6 (100), 208.7 (100) | 326 | Dicaffeoylhexaric acid isomer 3 | x | x | |
| 25 | 16.7 | 935.3 | 632.9 (45), 300.6 (100) | 270 | Casuarictin/Potentillin isomer 3 | x | ||
| 26 | 16.9 | 515.1 | 353.0 (100), 190.6 (41) | 325 | Dicaffeoylquinic acid isomer 1 | x | ||
| 27 | 17.2 | 463.0 | 300.5 (100) | 350 | Ellagic acid hexoside | x | ||
| 28 | 18.1 | 367.6 | 178.6 (100), 134.8 (31) | 330 | Feruloylquinic acid isomer 3 | x | x | |
| 29 | 18.6 | 533.4 | 370.6 (100), 208.6 (19) | 328 | Dicaffeoylhexaric acid isomer 4 | x | ||
| 30 | 18.8 | 479.3 | 316.7 (100) | 360 | Myricetin hexoside isomer 1 | x | ||
| 31 | 20.6 | 479.5 | 316.7 (100) | 365 | Myricetin hexoside isomer 2 | x | ||
| 32 | 20.6 | 934.5 | 915.0 (68), 632.9 (35), 300.7 (42) | 280 | Casuarictin/Potentillin isomer 4 | x | ||
| 33 | 20.7 | 533.1 | 370.6 (100), 208.7 (17) | 330 | Dicaffeoylhexaric acid isomer 5 | x | x | |
| 34 | 21.2 | 625.3 | 316.5 (100) | 360 | Myricetin rutinoside | x | x | |
| 35 | 23.3 | 367.2 | 192.6 (22), 178.7 (100) 134.9 (32) | 320 | Feruloylquinic acid isomer 4 | x | ||
| 36 | 23.7 | 533.1 | 370.6 (100), 208.7 (42) | 332 | Dicaffeoylhexaric acid isomer 6 | x | x | |
| 37 | 24.8 | 515.1 | 353.0 (100), 190.6 (42) | 330 | Dicaffeoylquinic acid isomer 2 | x | ||
| 38 | 25.0 | 367.1 | 178.6 (100), 134.7 (31) | 326 | Feruloylquinic acid isomer 5 | x | x | |
| 39 | 25.1 | 435.0 | 301.3 (100) | 350 | Ellagic acid pentoside | x | ||
| 40 | 25.2 | 577.0 | 424.7 (100), 406.9 (62) 288.7 (21) | 280 | Procyanidin B-type dimer 3 | x | ||
| 41 | 25.4 | 435.0 | 301.0 (100) | 367 | Quercetin pentoside isomer 1 | x | ||
| 42 | 26.0 | 435.1 | 301.3 (100) | 361 | Quercetin pentoside isomer 2 | x | ||
| 43 | 26.8 | 463.4 | 301.2 (100) | 351 | Quercetin hexoside isomer 1 | x | x | |
| 44 | 27.1 | 533.0 | 370.5 (100) | 340 | Dicaffeoylhexaric acid isomer 7 | x | ||
| 45 | 27.4 | 477.3 | 300.6 (100) | 350 | Quercetin glucuronide | x | x | |
| 46 | 28.0 | 447.1 | 300.6 (100) | 367 | Ellagic acid rhamnoside | x | ||
| 47 | 28.5 | 463.0 | 300.6 (100) | 354 | Quercetin hexoside isomer 2 | x | x | x |
| 48 | 29.0 | 609.3 | 300.6 (100) | 352 | Quercetin rutinoside | x | x | |
| 49 | 30.0 | 533.3 | 370.6 (100), 208.6 (43) | 330 | Dicaffeoylhexaric acid isomer 8 | x | ||
| 50 | 30.4 | 505.6 | 300.6 (100) | 354 | Quercetin acetylhexoside isomer 1 | x | ||
| 51 | 30.6 | 515.4 | 352.7 (100), 190.7 (22) | 327 | Dicaffeoylquinic acid isomer 3 | x | x | |
| 52 | 31.1 | 505.3 | 300.6 (100) | 353 | Quercetin acetylhexoside isomer 2 | x | x | |
| 53 | 31.7 | 505.3 | 300.6 (100) | 352 | Quercetin acetylhexoside isomer 3 | x | x | |
| 54 | 32.2 | 447.4 | 284.8 (100) | 340 | Kampferol hexoside | x | ||
| 55 | 32.2 | 433.4 | 300.6 (100) | 350 | Quercetin pentoside isomer 3 | x | ||
| 56 | 33.8 | 447.3 | 300.6 (100) | 356 | Quercetin rhamnoside | x | x | |
| 57 | 35.4 | 593.4 | 284.6 (100) | 340 | Kaempferol rutinoside | x | x | |
| 58 | 35.7 | 785.5 | 314.6 (100) | 354 | Isorhamnetin rutinoside hexoside | x | x | |
| 59 | 36.1 | 515.3 | 352.7 (100) | 326 | Dicaffeoylquinic acid isomer 4 | x | x | |
| 60 | 36.3 | 489.4 | 284.6 (100) | 340 | Kaempferol acetylhexoside | x | ||
| 61 | 36.5 | 507.1 | 344.6 (100) | 320 | Siringetin hexoside | x | ||
| 62 | 36.6 | 477.3 | 315.7 (100) | 345 | Isorhamnetin hexoside | x | ||
| 63 | 38.3 | 623.4 | 315.7 (100) | 355 | Isorhamnetin rutinoside | x | x | |
| 64 | 39.5 | 447.5 | 284.6 (100) | 340 | Kaempferol hexoside | x | ||
| 65 | 40.5 | 519.3 | 315.6 (100) | 355 | Isorhamnetin acetylhexoside isomer 1 | x | ||
| 66 | 41.0 | 431.4 | 284.7 (100) | 340 | Kaempferol rhamnoside | x | x | |
| 67 | 42.3 | 519.6 | 315.6 (100) | 340 | Isorhamnetin acetylhexoside isomer 2 | x | ||
| 68 | 44.2 | 489.7 | 284.8 (100) | 340 | Kaempferol acetylhexoside | x | ||
| Compounds | Fragaria chiloensis | |
|---|---|---|
| Arroyo Llodcondo | Frey | |
| Anthocyanins | ||
| Cyanidin-3-glucoside (3a) | 7.1 ± 0.2 | 0.7 ± 0.1 |
| Pelargonidin hexoside (6a) | 5.8 ± 0.1 | 1.7 ± 0.0 |
| Cyanidin-malonyl hexoside (10a) | 0.8 ± 0.3 | BQL |
| Flavonols | ||
| Quercetin pentoside 2 (42) | 6.2 ± 0.2 | 7.1 ± 0.1 |
| Quercetin glucuronide (45) | 2.2 ± 0.3 | 0.9 ± 0.0 |
| Quercetin pentoside 3 (55) | 2.8 ± 0.0 | 2.4 ± 0.0 |
| Compounds | Berberis microphylla | Berberis darwinii | |||||
|---|---|---|---|---|---|---|---|
| Brazo Rincón | Aeropuerto | Villa La Angostura | Cuyín Manzano | Llanquín | Brazo Rincón | Villa La Angostura | |
| Anthocyanins | |||||||
| Delphinidin-3-glucoside (1a) | 78.6 ± 0.9 a | 621.7 ± 9.1 b | 459.8 ± 5.8 c | 397.1 ± 0.9 d | 301.7 ± 1.6 e | 115.3 ± 2.7 | 163.3 ± 4.4 |
| Delphinidin rutinoside (2a) | 1.8 ± 0.3 a | 66.4 ± 3.5 b | 35.5 ± 1.7 c | 29.1 ± 1.0 d | 27.1 ± 0.3 d | 11.0 ± 0.1 | 10.9 ± 0.7 |
| Cyanidin-3-glucoside (3a) | 1.2 ± 0.0 a | 51.3 ± 3.4 b | 40.0 ± 1.7 c | 52.3 ± 1.1 b | 47.8 ± 2.6 b | 30.4 ± 0.0 | 68.2 ± 2.0 |
| Cyanidin-3-rutinoside (4a) | BQL | BQL | BQL | BQL | BQL | BQL | 2.3 ± 0.0 |
| Petunidin hexoside (5a) | 35.7 ± 1.0 a | 363.6 ± 6.0 b | 271.2 ± 1.4 c | 229.8 ± 0.8 d | 185.7 ± 9.7 e | 61.9 ± 0.1 | 83.7 ± 2.8 |
| Petunidin rutinoside (7a) | 1.0 ± 0.0 a | 78.5 ± 3.9 b | 37.2 ± 0.1 c | 30.3 ± 0.6 d | 33.5 ± 0.0 c,d | 7.3 ± 0.1 | 8.7 ± 0.9 |
| Peonidin hexoside (8a) | BQL | 11.2 ± 0.9 a | 6.4 ± 0.8 b | 15.3 ± 0.2 c | 16.5 ± 0.8 c | 1.2 ± 0.0 | 5.3 ± 0.7 |
| Peonidin rutinoside (9a) | BQL | BQL | BQL | BQL | 1.4 ± 0.1 | BQL | BQL |
| Malvidin-3-glucoside (11a) | 11.5 ± 0.4 a | 247.0 ± 3.6 b | 172.4 ± 0.2 c | 163.4 ± 0.3 d | 181.7 ± 4.8 e | 59.3 ± 0.8 | 66.8 ± 1.7 |
| Malvidin rutinoside (12a) | BQL | 54.5 ± 4.8 a | 21.9 ± 1.1 b | 17.4 ± 0.4 b | 35.6 ± 1.2 c | 4.3 ± 0.8 | 3.8 ± 0.7 |
| Hydroxycinnamic acids | |||||||
| Caffeoylglucaric acid 1 (2) | 20.8 ± 0.6 a | 46.2 ± 1.6 b | 48.4 ± 0.3 c | 21.5 ± 0.4 a | 15.1 ± 0.3 d | 22.4 ± 0.4 | 96.3 ± 2.0 |
| Caffeoylglucaric acid 2 (5) | 9.2 ± 0.2 a | 23.3 ± 0.3 b | 27.4 ± 0.3 c | 46.3 ± 1.4 d | 22.3 ± 0.3 b | 59.2 ± 0.2 | 217.6 ± 1.4 |
| Caffeoylquinic acid 3 (13) | 163.7 ± 0.7 a | 74.1 ± 0.6 b | 163.4 ± 1.1 a | 35.2 ± 0.2 c | 31.6 ± 1.1 d | 100.0 ± 0.3 | 328.3 ± 2.2 |
| Dicaffeoyl glucaric acid 2 (14) | 28.2 ± 0.3 a | 20.0 ± 0.3 b | 56.3 ± 0.5 c | 25.2 ± 0.2 d | 17.6 ± 0.1 e | 7.1 ± 0.7 | 67.4 ± 0.1 |
| Caffeoylquinic acid 3 (16) | 3.2 ± 0.1 a | 4.9 ± 0.3 b | 6.2 ± 0.4 c | 6.8 ± 0.2 c | 4.2 ± 0.2 d | 7.6 ± 0.1 | 28.2 ± 0.2 |
| Feruloylquinic acid (18) | 6.3 ± 0.0 a | 17.3 ± 0.9 b | 16.2 ± 0.9 b | 23.3 ± 0.8 c | 12.6 ± 0.2 d | 6.8 ± 0.3 | 21.6 ± 0.5 |
| Dicaffeoyl glucaric acid 3 (24) | 2.1 ± 0.3 a | 1.8 ± 0.2 a | 2.8 ± 1.2 a | 5.4 ± 0.0 b | 7.1 ± 0.0 c | 27.2 ± 1.2 | 93.2 ± 0.1 |
| Dicaffeoyl glucaric acid 6 (36) | 2.8 ± 0.2 a | 3.3 ± 0.1 b | 6.4 ± 0.3 c | 5.2 ± 0.0 d | 9.5 ± 0.0 e | 24.1 ± 1.5 | 123.6 ± 3.4 |
| Dicaffeoylquinic acid 7 (51) | 16.5 ± 0.3 a | 11.6 ± 0.3 b | 20.3 ± 0.2 c | 9.6 ± 0.1 d | 4.5 ± 0.8 e | 10.4 ± 0.8 | 21.1 ± 0.5 |
| Flavonols | |||||||
| Quercetin hexoside (43) | 10.5 ± 0.0 a | 41.5 ± 0.5 b | 24.6 ± 1.3 c | 31.2 ± 1.0 d | 26.2 ± 0.9 c | 29.8 ± 0.1 | 56.6 ± 2.5 |
| Quercetin glucuronide (45) | ND | ND | 2.8 ± 0.1 | ND | ND | 3.0 ± 0.3 | 13.6 ± 0.4 |
| Quercetin rutinoside (48) | 14.1 ± 0.0 a | 53.5 ± 2.7 b | 34.6 ± 0.5 c | 40.0 ± 0.9 d | 18.5 ± 1.2 e | 25.0 ± 0.7 | 57.3 ± 5.8 |
| Quercetin acetylhexoside 1 (52) | 12.2 ± 0.4 a | BQL | 20.7 ± 0.1 b | 7.1 ± 0.1 c | 5.4 ± 0.2 d | 25.1 ± 0.5 | 59.8 ± 1.7 |
| Quercetin rhamnoside (56) | 18.8 ± 0.6 a | 0.2 ± 0.3 b | ND | 4.2 ± 0.2 c | 1.5 ± 0.3 d | 28.0 ± 0.3 | 26.6 ± 1.1 |
| Isorhamnetin rutinoside (63) | 4.2 ± 0.1 a | 55.7 ± 2.6 b | 9.6 ± 0.9 a | 26.6 ± 3.1 c | 22.1 ± 2.2 c | BQL | BQL |
| Isorhamnetin acetylhexoside 1 (65) | 3.9 ± 0.0 a | 8.6 ± 0.5 b | 6.8 ± 0.0 c | 6.7 ± 0.2 c | 9.9 ± 0.2 d | ND | ND |
| Isorhamnetin acetylhexoside 2 (67) | 2.1 ± 0. 2a | 6.8 ± 0.6 b | 6.5 ± 0.0 b | 6.5 ± 0.4 b | 3.2 ± 0.1 c | ND | 5.3 ± 0.4 |
| Compound | DPPH | FRAP | TEAC | CUPRAC | ORAC | α-glucosidase |
|---|---|---|---|---|---|---|
| Anthocyanins | ||||||
| 1a | −0.866 ** | 0.884 ** | 0.851 ** | 0.832 ** | 0.796 ** | −0.518 * |
| 2a | −0.736 ** | 0.773 ** | 0.798 ** | 0.702 ** | 0.708 ** | −0.311 |
| 3a | −0.630 * | 0.675 ** | 0.991 ** | 0.664 ** | 0.992 ** | −0.653 ** |
| 5a | −0.848 ** | 0.869 ** | 0.873 ** | 0.830 ** | 0.816 ** | −0.512 |
| 7a | −0.674 ** | 0.709 ** | 0.784 ** | 0.633 * | 0.682 ** | −0.240 |
| 8a | 0.675 * | −0.568 | 0.446 | −0.717 ** | 0.607 * | 0.104 |
| 10a | −0.678 ** | 0.706 ** | 0.962 ** | 0.687 ** | 0.900 ** | −0.456 |
| 11a | 0.078 | 0.002 | 0.774 ** | −0.127 | −0.150 | 0.765 ** |
| Hydroxycinnamic acids | ||||||
| 2 | −0.755 ** | 0.734 ** | 0.331 | 0.759 ** | 0.259 | −0.212 |
| 5 | −0.642 ** | 0.666 ** | 0.665 ** | 0.617 * | 0.786 ** | −0.957 ** |
| 13 | 0.074 | −0.133 | −0.664 ** | −0.030 | −0.682 ** | 0.436 |
| 14 | −0.380 | 0.311 | −0.091 | 0.489 | −0.052 | −0.285 |
| 16 | −0.799** | 0.825 ** | 0.649 ** | 0.842 ** | 0.755 ** | −0.952 ** |
| 18 | −0.795 ** | 0.837 ** | 0.790 ** | 0.765 ** | 0.870 ** | −0.918 ** |
| 24 | 0.287 | −0.236 | 0.385 | −0.167 | 0.454 | −0.242 |
| 36 | 0.176 | −0.178 | 0.471 | 0.012 | 0.482 | −0.150 |
| 51 | −0.265 | 0.216 | −0.439 | 0.298 | −0.428 | 0.016 |
| Flavonols | ||||||
| 43 | −0.712 ** | 0.768 ** | 0.887 ** | 0.659 ** | 0.839 ** | −0.476 |
| 48 | −0.901 ** | 0.938 ** | 0.689 ** | 0.816 ** | 0.665 ** | −0.564 * |
| 52 | −0.441 | 0.383 | −0.186 | 0.498 | −0.200 | −0.045 |
| 56 | 0.614 * | −0.620 * | −0.987 ** | −0.677 * | −0.944 ** | 0.498 |
| 64 | −0.496 | 0.571 * | 0.668 ** | 0.408 | 0.586 * | −0.167 |
| 66 | −0.191 | 0.235 | 0.842 ** | 0.284 | 0.766 ** | −0.138 |
| 69 | −0.975 ** | 0.992 ** | 0.738 | 0.948 ** | 0.759 ** | −0.779 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamorro, M.F.; Reiner, G.; Theoduloz, C.; Ladio, A.; Schmeda-Hirschmann, G.; Gómez-Alonso, S.; Jiménez-Aspee, F. Polyphenol Composition and (Bio)Activity of Berberis Species and Wild Strawberry from the Argentinean Patagonia. Molecules 2019, 24, 3331. https://doi.org/10.3390/molecules24183331
Chamorro MF, Reiner G, Theoduloz C, Ladio A, Schmeda-Hirschmann G, Gómez-Alonso S, Jiménez-Aspee F. Polyphenol Composition and (Bio)Activity of Berberis Species and Wild Strawberry from the Argentinean Patagonia. Molecules. 2019; 24(18):3331. https://doi.org/10.3390/molecules24183331
Chicago/Turabian StyleChamorro, Melina F., Gabriela Reiner, Cristina Theoduloz, Ana Ladio, Guillermo Schmeda-Hirschmann, Sergio Gómez-Alonso, and Felipe Jiménez-Aspee. 2019. "Polyphenol Composition and (Bio)Activity of Berberis Species and Wild Strawberry from the Argentinean Patagonia" Molecules 24, no. 18: 3331. https://doi.org/10.3390/molecules24183331
APA StyleChamorro, M. F., Reiner, G., Theoduloz, C., Ladio, A., Schmeda-Hirschmann, G., Gómez-Alonso, S., & Jiménez-Aspee, F. (2019). Polyphenol Composition and (Bio)Activity of Berberis Species and Wild Strawberry from the Argentinean Patagonia. Molecules, 24(18), 3331. https://doi.org/10.3390/molecules24183331