Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
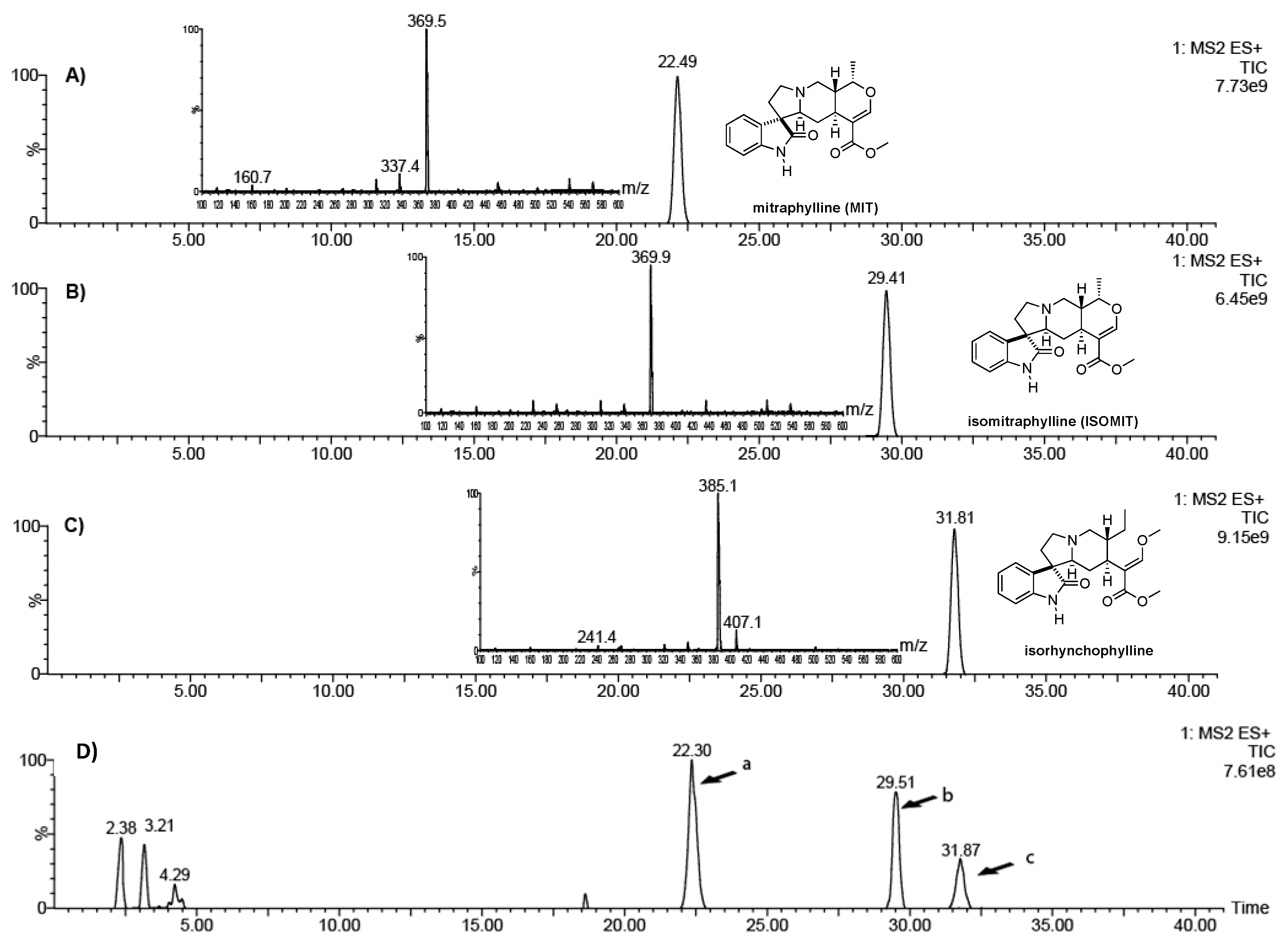
2.1. Chemical Characterization of a Leaf Extract from U. tomentosa
2.2. In Vitro Antioxidant Activity
2.3. In Vivo Antioxidant Activity of ALE, Mitraphylline, and Isomitraphylline in C. elegans
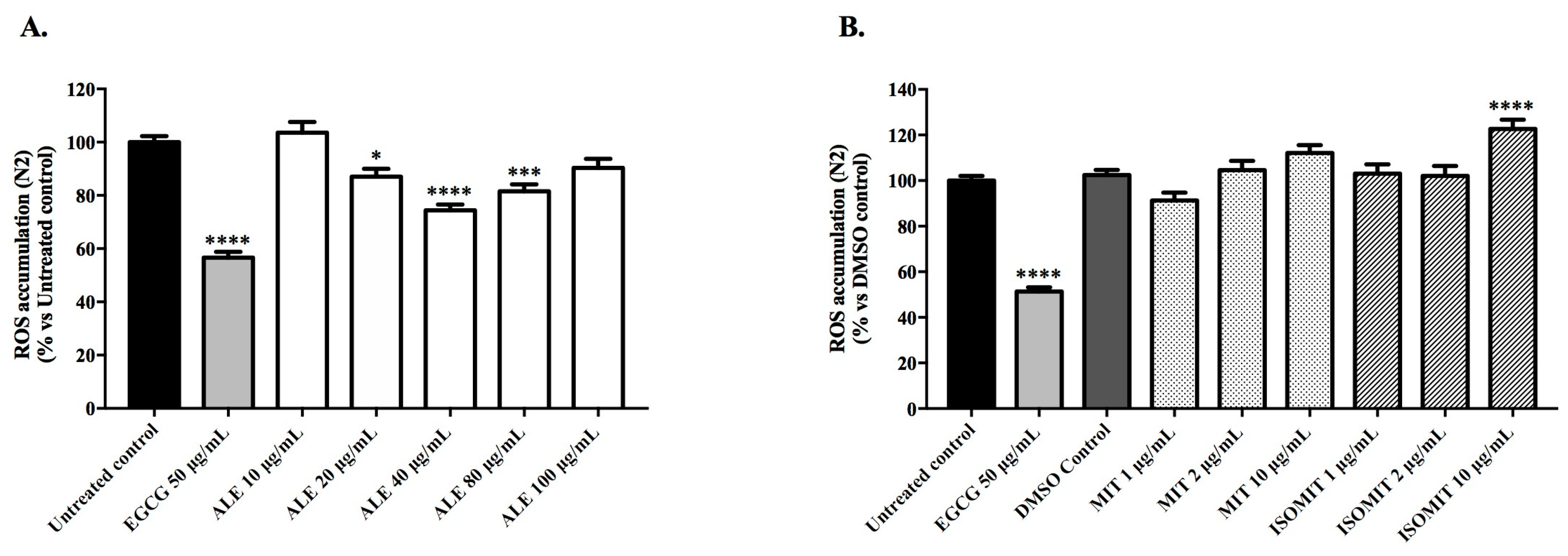
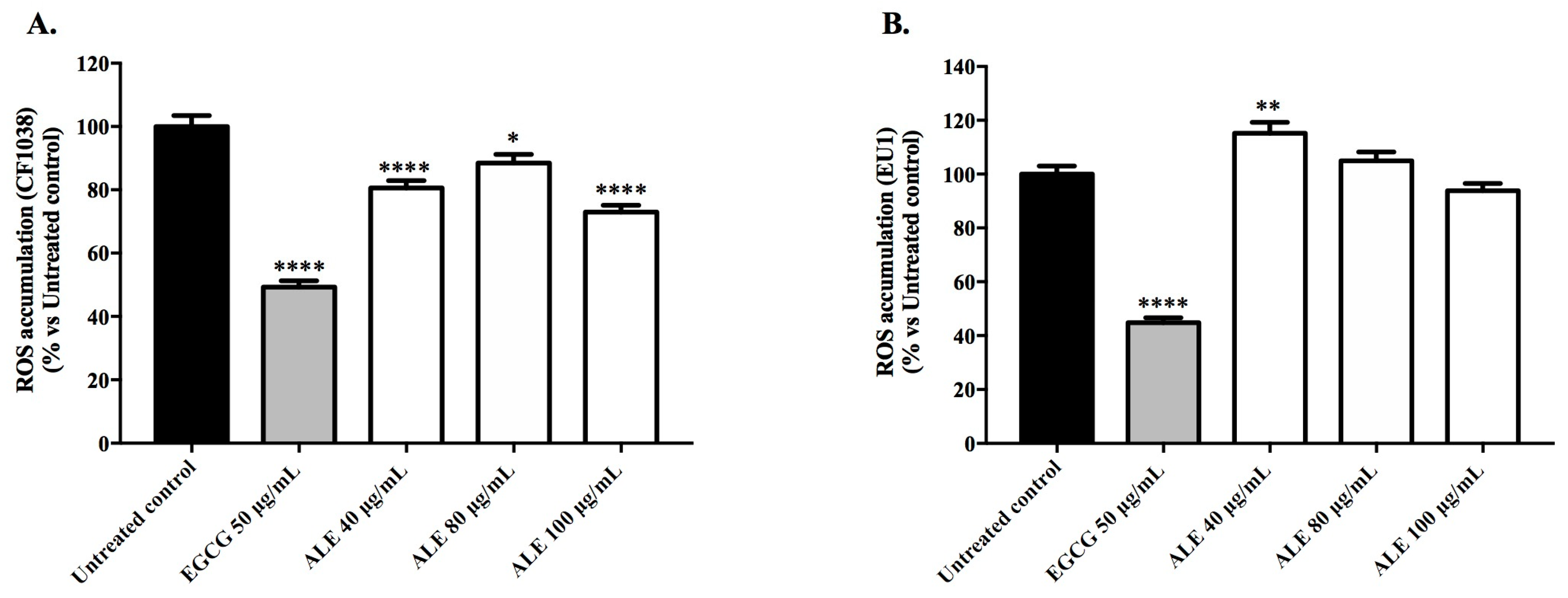
2.3.1. Intracellular Accumulation of ROS in Wild-Type Worms
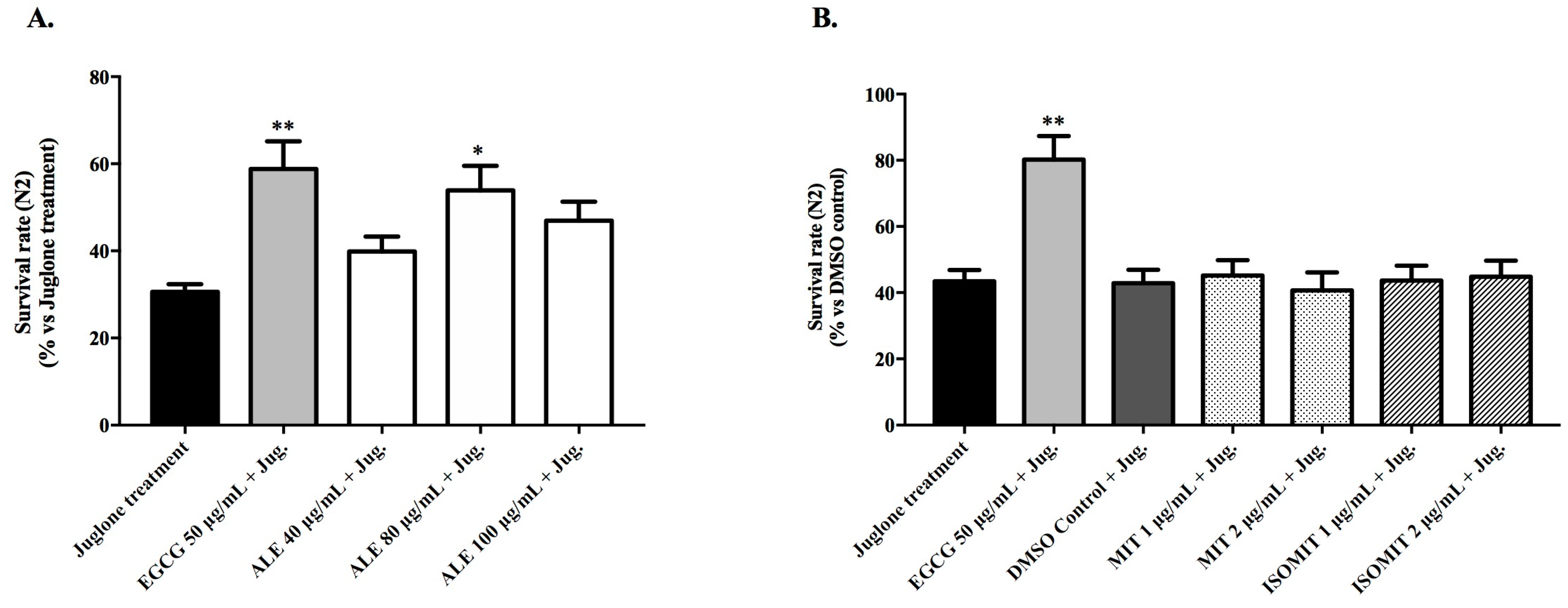
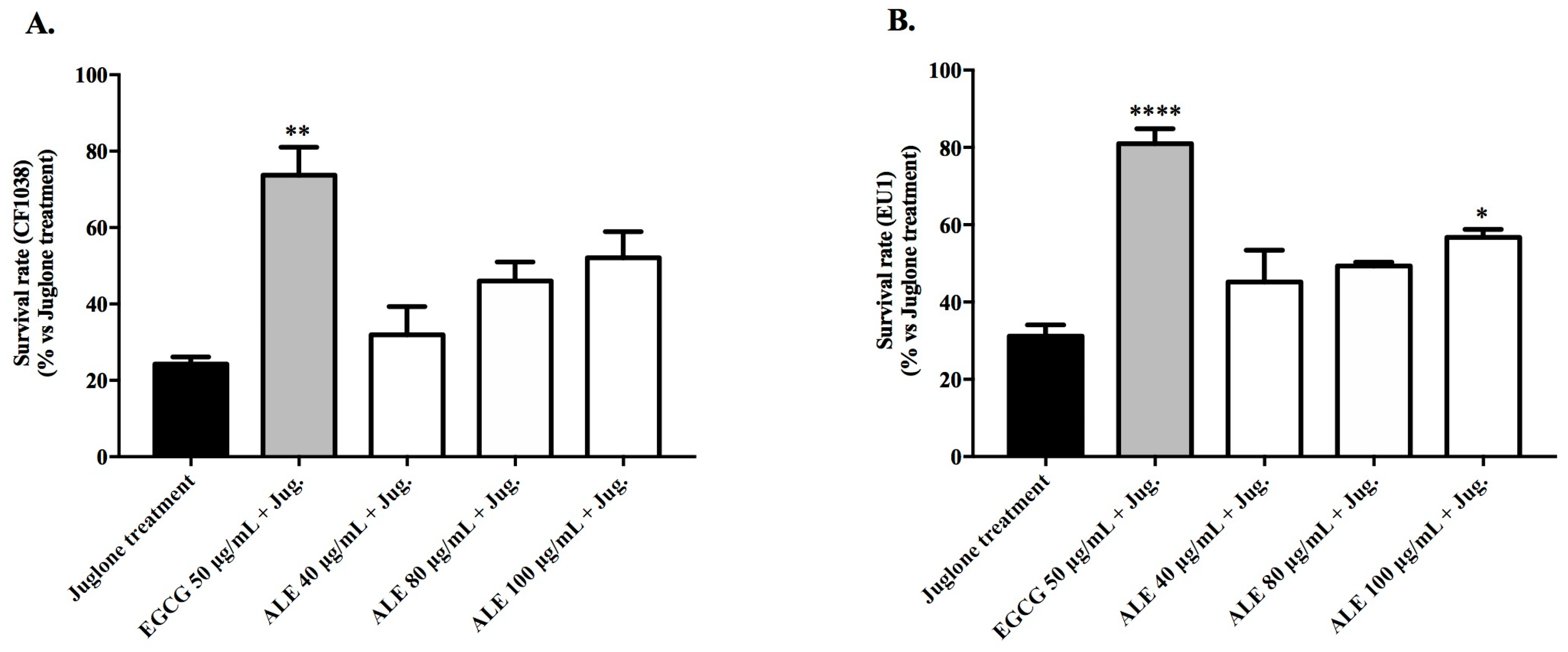
2.3.2. Survival Rate of Wild-Type Worms Exposed to a Lethal Dose of Juglone
2.4. Mechanism of the Antioxidant Activity of ALE in C. elegans
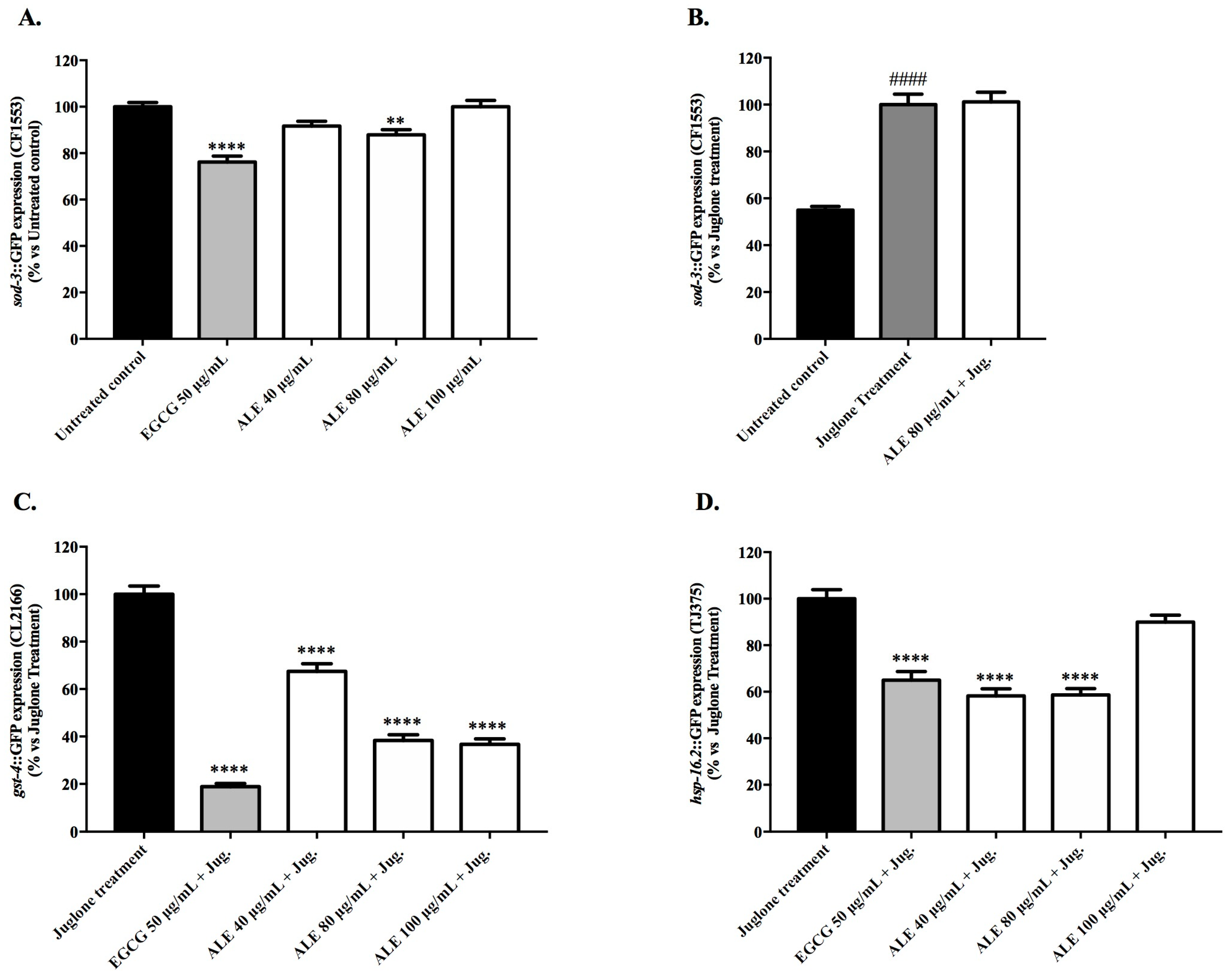
2.4.1. Effect of ALE on the Expression Levels of the Oxidative Stress Resistance Related Genes: sod-3, gst-4, and hsp-16.2
2.4.2. Involvement of DAF-16 and SKN-1 in the Antioxidant Activity of ALE
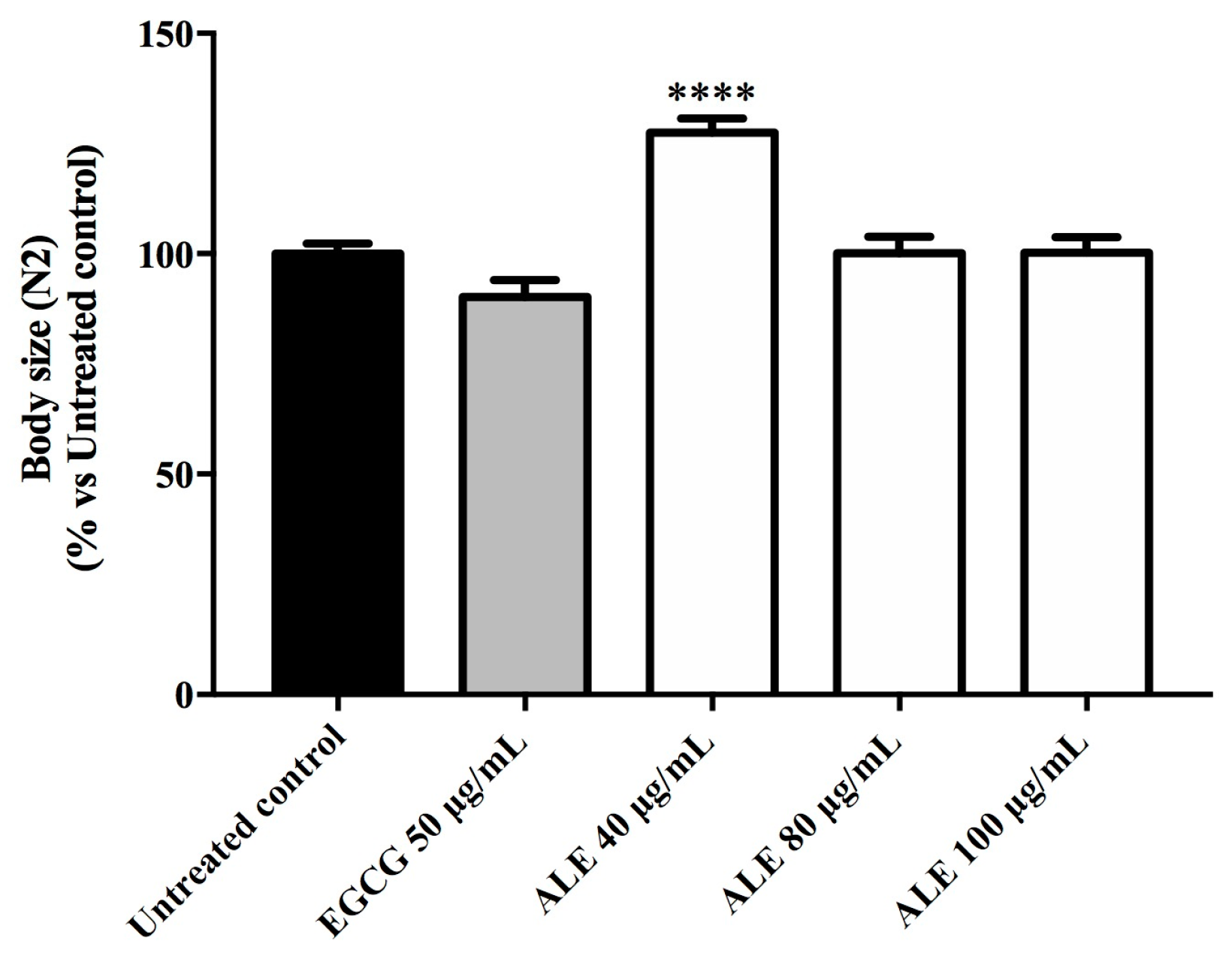
2.5. Effect of ALE on C. elegans Body Size and Escherichia coli OP50 Growth
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extracts
4.2. Analysis of Oxindole Alkaloids in the Leaf Extract
4.3. Isolation of Isorhynchophylline from Crude Extracts
4.4. In Vitro Antioxidant Activity and Total Phenolic Content of U. tomentosa Leaf Extract
4.5. Maintenance of C. elegans Strains
4.6. In Vivo Antioxidant Activity of ALE, Mitraphylline, and Isomitraphylline Using C. elegans as a Model Organism
4.6.1. Intracellular Accumulation of ROS in Wild-Type Worms
4.6.2. Survival Rate of Wild-Type Worms Exposed to a Lethal Dose of Juglone
4.7. Mechanism of the Antioxidant Activity of ALE in C. elegans
4.7.1. Effect of ALE on the Expression Levels of the Oxidative Stress Resistance Related Genes: sod-3, gst-4, and hsp-16.2
4.7.2. Involvement of DAF-16 and SKN-1 in the Antioxidant Activity of ALE: ROS and Survival Assays Using DAF-16 and SKN-1 Mutants
4.8. Anti-Microbial Activity of ALE against E. coli OP50
4.9. Effect of ALE on C. elegans Body Size
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zevallos-Pollito, P.A.; Tomazello Filho, M. Levantamento e caracterização de duas espécies do gênero Uncaria schreb. (Rubiaceae) correntes no Estado do Acre, Brasil. Ecol. Apl. 2010, 9, 19–30. [Google Scholar] [CrossRef][Green Version]
- Laus, G.; Brössner, D.; Keplinger, K. Alkaloids of Peruvian Uncaria tomentosa. Phytochemistry 1997, 45, 855–860. [Google Scholar] [CrossRef]
- Montoro, P.; Carbone, V.; Quiroz, J.D.; De Simone, F.; Pizza, C. Identification and quantification of components in extracts of Uncaria tomentosa by HPLC-ES/MS. Phytochem. Anal. 2004, 15, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Pavei, C.; Kaiser, S.; Verza, S.G.; Borre, G.L.; Ortega, G.G. HPLC-PDA method for quinovic acid glycosides assay in Cat’s Claw (Uncaria tomentosa) associated with UPLC/QTOF– MS analysis. J. Pharm. Biomed. Anal. 2002, 62, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Akesson, C.; Holmgren, K.; Bryngelsson, C.; Giamapa, V.; Pero, R.W. An active ingredient of Cats´s Claw water extracts: Identification and efficacy of quinic acid. J. Ethnopharmacol. 2005, 96, 577–584. [Google Scholar] [CrossRef]
- Vilches, L.E.O. Género Uncaria-Estudios Botanicos, Químicos Y Farmacologicos De Uncaria Tomentosa Y Uncaria Guianensis, 3rd ed.; Instituto de Fitoterapia Americano: Lima, Peru, 1996; pp. 54–55. [Google Scholar]
- Snow, A.D.; Castillo, G.M.; Nguyen, B.P.; Choi, P.Y.; Cummings, J.A.; Cam, J.; Hu, Q.; Lake, T.; Pan, W.; Kastin, A.J.; et al. The Amazon rain forest plant Uncaria tomentosa (cat’s claw) and its specific proanthocyanidin constituents are potent inhibitors and reducers of both brain plaques and tangles. Sci. Rep. 2019, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, B.C.; Morel, L.J.F.; Carmona, F.; Cunha, T.M.; Contini, S.H.T.; Delprete, P.G.; Ramalho, F.S.; Crevelin, E.; Bertoni, B.W.; França, S.C.; et al. Aqueous extracts from Uncaria tomentosa (Willd. ex Schult.) DC. reduce bronchial hyperresponsiveness and inflammation in a murine model of asthma. J. Ethnopharmacol. 2018, 218, 76–89. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, J.J.; Xu, J.; Feng, F.; Qu, W. Medicinal uses, phytochemistry and pharmacology of the genus Uncaria. J. Ethnopharmacol. 2015, 173, 48–80. [Google Scholar] [CrossRef]
- Sroka, Z.; Fecka, I.; Cisowski, W.; Łuczkiewicz, M. Investigation into the anti-radical activity of extracts obtained from Uncaria tomentosa Willd DC. Herb. Pol. 2001, 47, 218–224. [Google Scholar]
- Sandoval, M.; Charbonnet, R.M.; Okuhama, N.N.; Roberts, J.; Krenova, Z.; Trentacosti, A.M.; Miller, M.J.S. Cat’s claw inhibits TNFα production and scavenges free radicals: Role in cytoprotection. Free Radic. Biol. Med. 2000, 29, 71–78. [Google Scholar] [CrossRef]
- Navarro-Hoyos, M.; Alvarado-Corella, D.; Moreira-Gonzalez, I.; Arnaez-Serrano, E.; Monagas-Juan, M. Polyphenolic composition and antioxidant activity of aqueous and ethanolic extracts from Uncaria tomentosa bark and leaves. Antioxidants 2018, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Amaral, S.; Mira, L.; Nogueira, J.M.F.; da Silva, A.P.; Florêncio, M.H. Plant extracts with anti-inflammatory properties-A new approach for characterization of their bioactive compounds and establishment of structure–antioxidant activity relationships. Bioorganic Med. Chem. 2009, 17, 1876–1883. [Google Scholar] [CrossRef] [PubMed]
- Pilarski, R.; Zieliński, H.; Ciesiołka, D.; Gulewicz, K. Antioxidant activity of ethanolic and aqueous extracts of Uncaria tomentosa (Willd.) DC. J. Ethnopharmacol. 2006, 104, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, C.; Dinis, T.; Batista, M.T. Antioxidant properties of proanthocyanidins of Uncaria tomentosa bark decoction: A mechanism for anti-inflammatory activity. Phytochemistry 2005, 66, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Bors, M.; Bukowska, B.; Pilarski, R.; Gulewicz, K.; Oszmiański, J.; Michałowicz, J.; Koter-Michalak, M. Protective activity of the Uncaria tomentosa extracts on human erythrocytes in oxidative stress induced by 2, 4-dichlorophenol (2, 4-DCP) and catechol. Food Chem. Toxicol. 2011, 49, 2202–2211. [Google Scholar] [CrossRef]
- Dreifuss, A.A.; Bastos-Pereira, A.L.; Ávila, T.V.; Soley, B.S.; Rivero, A.J.; Aguilar, J.L.; Acco, A. Antitumoral and antioxidant effects of a hydroalcoholic extract of cat’s claw (Uncaria tomentosa) (Willd. Ex Roem. & Schult) in an in vivo carcinosarcoma model. J. Ethnopharmacol. 2010, 130, 127–133. [Google Scholar] [PubMed]
- Santo, G.D.; Grotto, A.; Boligon, A.A.; Costa, B.D.; Rambo, C.L.; Fantini, E.A.; Sauer, E.; Lazzarotto, L.M.V.; Bertoncello, K.T.; Júnior, O.T.; et al. Protective effect of Uncaria tomentosa extract against oxidative stress and genotoxicity induced by glyphosate-Roundup® using zebrafish (Danio rerio) as a model. Environ. Sci. Pollut. Res. 2018, 25, 11703–11715. [Google Scholar] [CrossRef]
- O’Reilly, L.P.; Luke, C.J.; Perlmutter, D.H.; Silverman, G.A.; Pak, S.C.C. Elegans in high-throughput drug discovery. Adv. Drug Deliv. Rev. 2014, 69, 247–253. [Google Scholar] [CrossRef]
- Culetto, E.; Sattelle, D.B. A role for Caenorhabditis elegans in understanding the function and interactions of human disease genes. Hum. Mol. Genet. 2000, 9, 869–877. [Google Scholar] [CrossRef]
- Koch, K.; Havermann, S.; Büchter, C.; Wätjen, W. Caenorhabditis elegans as model system in pharmacology and toxicology: Effects of flavonoids on redox-sensitive signalling pathways and ageing. Sci. World J. 2014, 2014, 920398. [Google Scholar] [CrossRef]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef]
- Rojas-Duran, R.; González-Aspajo, G.; Ruiz-Martel, C.; Bourdy, G.; Doroteo-Ortega, V.H.; Alban-Castillo, J.; Robert, G.; Auberger, P.; Deharo, E. Anti-inflammatory activity of mitraphylline isolated from Uncaria tomentosa bark. J. Ethnopharmacol. 2012, 143, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-de la Paz, S.; Fernandez-Arche, A.; de la Puerta, R.; Quilez, A.M.; Muriana, F.J.G.; Garcia-Gimenez, M.D.; Bermudez, B. Mitraphylline inhibits lipopolysaccharide-mediated activation of primary human neutrophils. Phytomedicine 2016, 23, 141–148. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxid. Med. Cell Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed]
- Castellani, P.; Balza, E.; Rubartelli, A. Inflammation, DAMPs, tumor development, and progression: A vicious circle orchestrated by redox signaling. Antioxid. Redox Signal. 2014, 20, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Wink, M. Modes of action of herbal medicines and plant secondary metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, M.; Okuhama, N.N.; Zhang, X.J.; Condezo, L.A.; Lao, J.; Angeles, F.M.; Musah, R.A.; Bobrowski, P.; Miller, M.J. Anti-inflammatory and antioxidant activities of cat’s claw (Uncaria tomentosa and Uncaria guianensis) are independent of their alkaloid content. Phytomedicine 2002, 9, 325–337. [Google Scholar] [CrossRef]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein Hsp16-2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J. 2003, 17, 2305–2307. [Google Scholar] [CrossRef]
- Masella, R.; Benedetto, R.D.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Cheng, Q.; Peng, Q.; Yu, X.; Yin, X.; Liang, M.; Ma, C.W.; Huang, Z.; Jia, W. Antioxidant peptides derived from the hydrolyzate of purple sea urchin (Strongylocentrotus nudus) gonad alleviate oxidative stress in Caenorhabditis elegans. J. Funct. Foods. 2018, 48, 594–604. [Google Scholar] [CrossRef]
- Shi, Y.C.; Yu, C.W.; Liao, V.H.C.; Pan, T.M. Monascus-fermented dioscorea enhances oxidative stress resistance via DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE 2012, 7, e39515. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.P.; Abate, J.P.; Dilks, K.; Landis, J.; Ashraf, J.; Murphy, C.T.; Blackwell, T.K. Condition-adapted stress and longevity gene regulation by Caenorhabditis elegans SKN-1/Nrf. Aging Cell 2009, 8, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 6, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Senchuk, M.M.; Dues, D.J.; Schaar, C.E.; Johnson, B.K.; Madaj, Z.B.; Bowman, M.J.; Winn, M.E.; Van Raamsdonk, J.M. Activation of DAF-16/FOXO by reactive oxygen species contributes to longevity in long-lived mitochondrial mutants in Caenorhabditis elegans. PLoS Genet. 2018, 3, e1007268. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Pielarski, T.; Rohrig, R.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. The Ginkgo biloba extract EGb761 reduces stress sensitivity, ROS accumulation and expression of catalase and glutathione S-transferase 4 in Caenorhabditis elegans. Pharmacol. Res. 2007, 55, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Röhrig, T.; Richling, E.; Wang, X.; Wink, M. Anti-aging and antioxidant potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans indicate a new utilization for roasted seeds of guarana. Medicines 2017, 4, 61. [Google Scholar] [PubMed]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. Elegans. In C. elegans: A Practical Approach; Hope, I.A., Ed.; Oxford University Press: Oxford, UK, 1999; pp. 51–67. [Google Scholar]
- Ashour, M.L.; El-Readi, M.Z.; Hamoud, R.; Eid, S.Y.; El Ahmady, S.H.; Nibret, E.; Herrmann, F.; Youns, M.; Tahrani, A.; Kaufmann, D.; et al. Anti-infective and cytotoxic properties of Bupleurum marginatum. Chin. Med. 2014, 9, 4. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | ABTS mmol Trolox/mg AA and Extract | DPPH IC50 µg/mL | FRAP mM FeSO4/mg AA and Extract |
|---|---|---|---|
| Ascorbic acid (AA) | 4.15 ± 0.15 | 2.05 ± 0.19 | 9.54 ± 0.54 |
| ALE | 2.15 ± 0.29 | 10.68 ± 0.64 | 2.9 ± 0.15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
C. Azevedo, B.; Roxo, M.; C. Borges, M.; Peixoto, H.; Crevelin, E.J.; W. Bertoni, B.; H. T. Contini, S.; Lopes, A.A.; C. França, S.; S. Pereira, A.M.; et al. Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans. Molecules 2019, 24, 3299. https://doi.org/10.3390/molecules24183299
C. Azevedo B, Roxo M, C. Borges M, Peixoto H, Crevelin EJ, W. Bertoni B, H. T. Contini S, Lopes AA, C. França S, S. Pereira AM, et al. Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans. Molecules. 2019; 24(18):3299. https://doi.org/10.3390/molecules24183299
Chicago/Turabian StyleC. Azevedo, Bruna, Mariana Roxo, Marcos C. Borges, Herbenya Peixoto, Eduardo J. Crevelin, Bianca W. Bertoni, Silvia H. T. Contini, Adriana A. Lopes, Suzelei C. França, Ana M. S. Pereira, and et al. 2019. "Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans" Molecules 24, no. 18: 3299. https://doi.org/10.3390/molecules24183299
APA StyleC. Azevedo, B., Roxo, M., C. Borges, M., Peixoto, H., Crevelin, E. J., W. Bertoni, B., H. T. Contini, S., Lopes, A. A., C. França, S., S. Pereira, A. M., & Wink, M. (2019). Antioxidant Activity of an Aqueous Leaf Extract from Uncaria tomentosa and Its Major Alkaloids Mitraphylline and Isomitraphylline in Caenorhabditis elegans. Molecules, 24(18), 3299. https://doi.org/10.3390/molecules24183299