Effect of Chlorogenic Acid on the Physicochemical and Functional Properties of Coregonus Peled Myofibrillar Protein through Hydroxyl Radical Oxidation

Abstract
1. Introduction
2. Results and Discussion
2.1. Effect of Chlorogenic Acid on Oxidized MP Physicochemical Properties
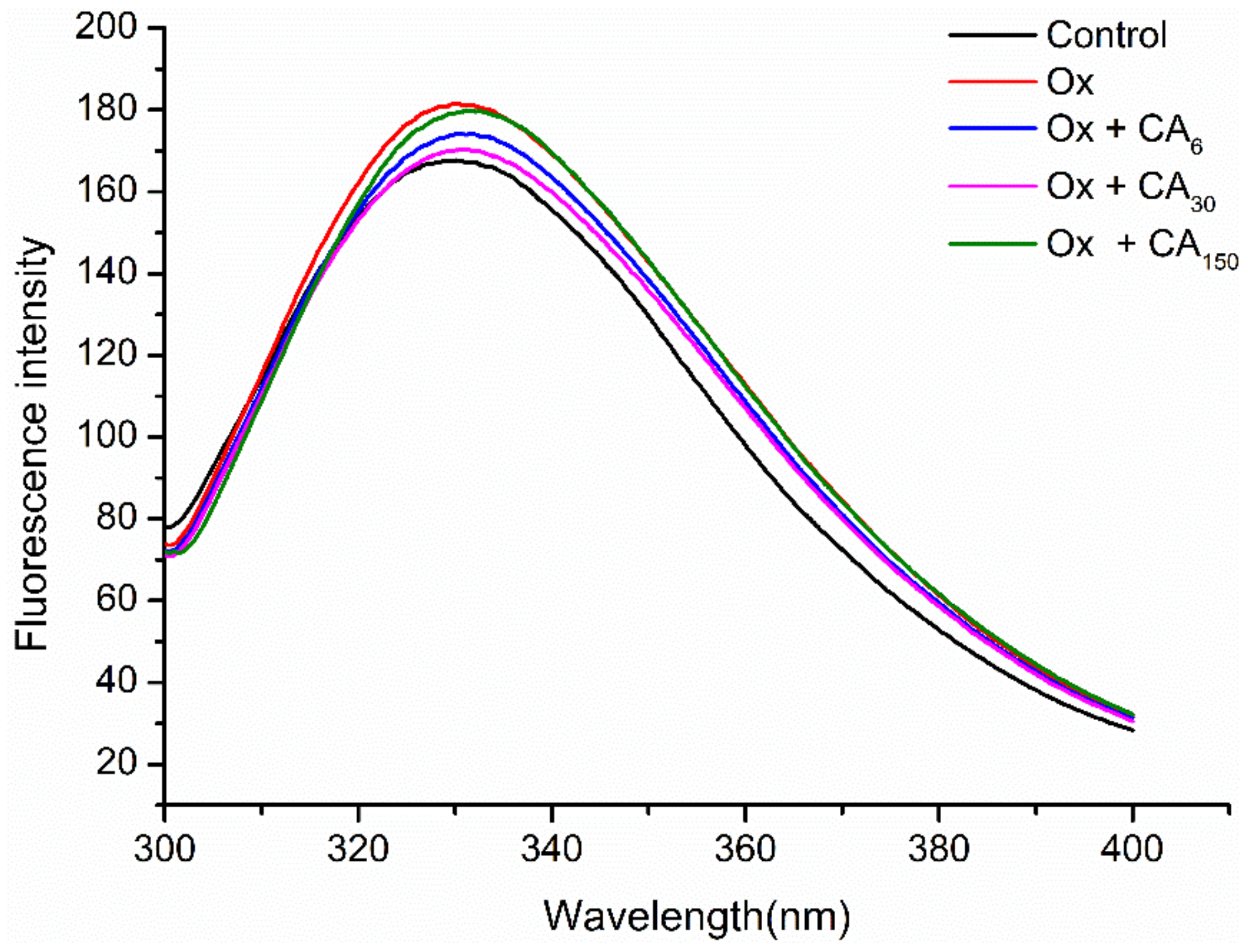
2.2. Changes in Endogenous Fluorescence Spectra
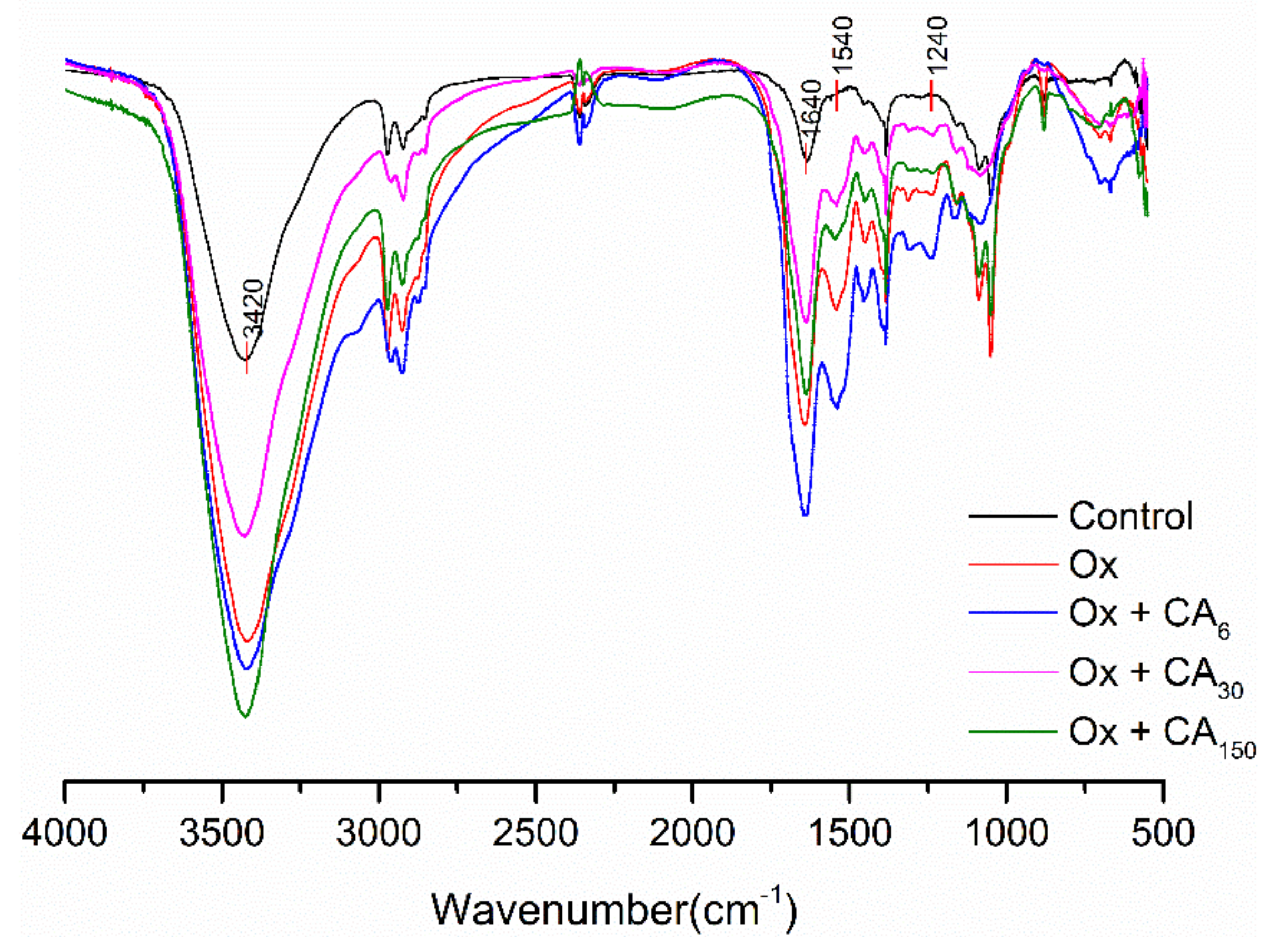
2.3. FT-IR
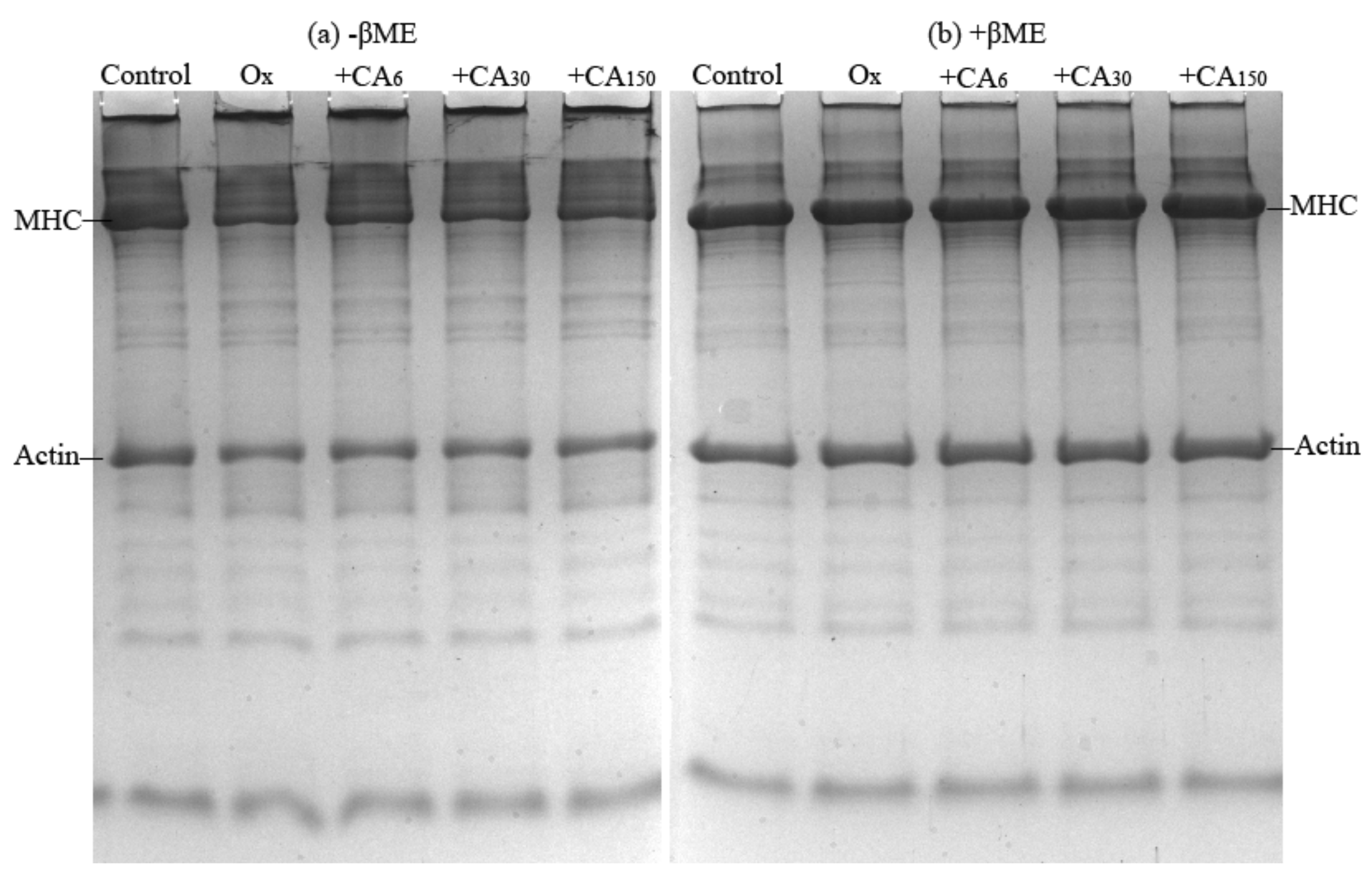
2.4. SDS- Polyacrylamide Gel Electrophoresis (SDS-PAGE)
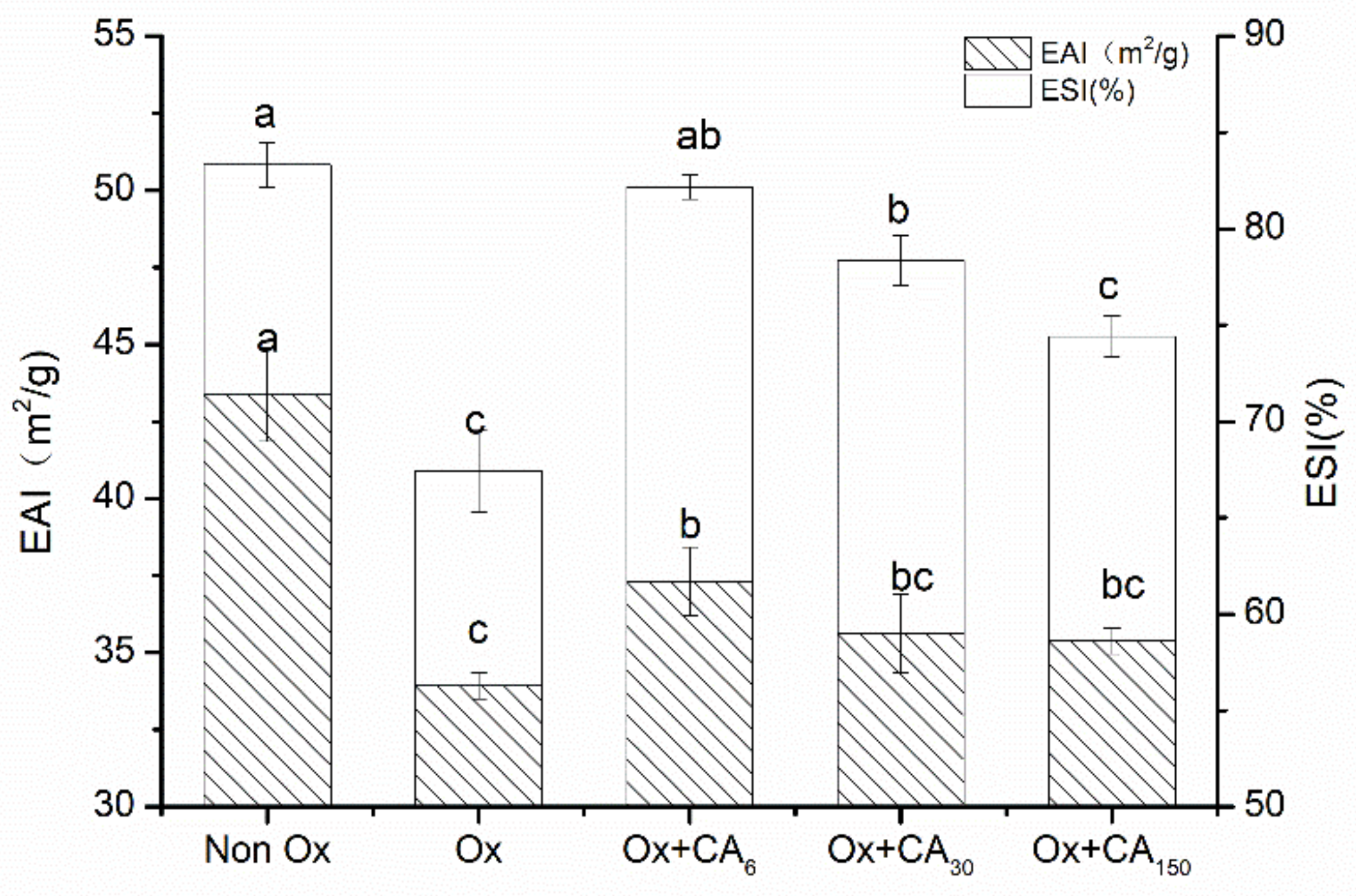
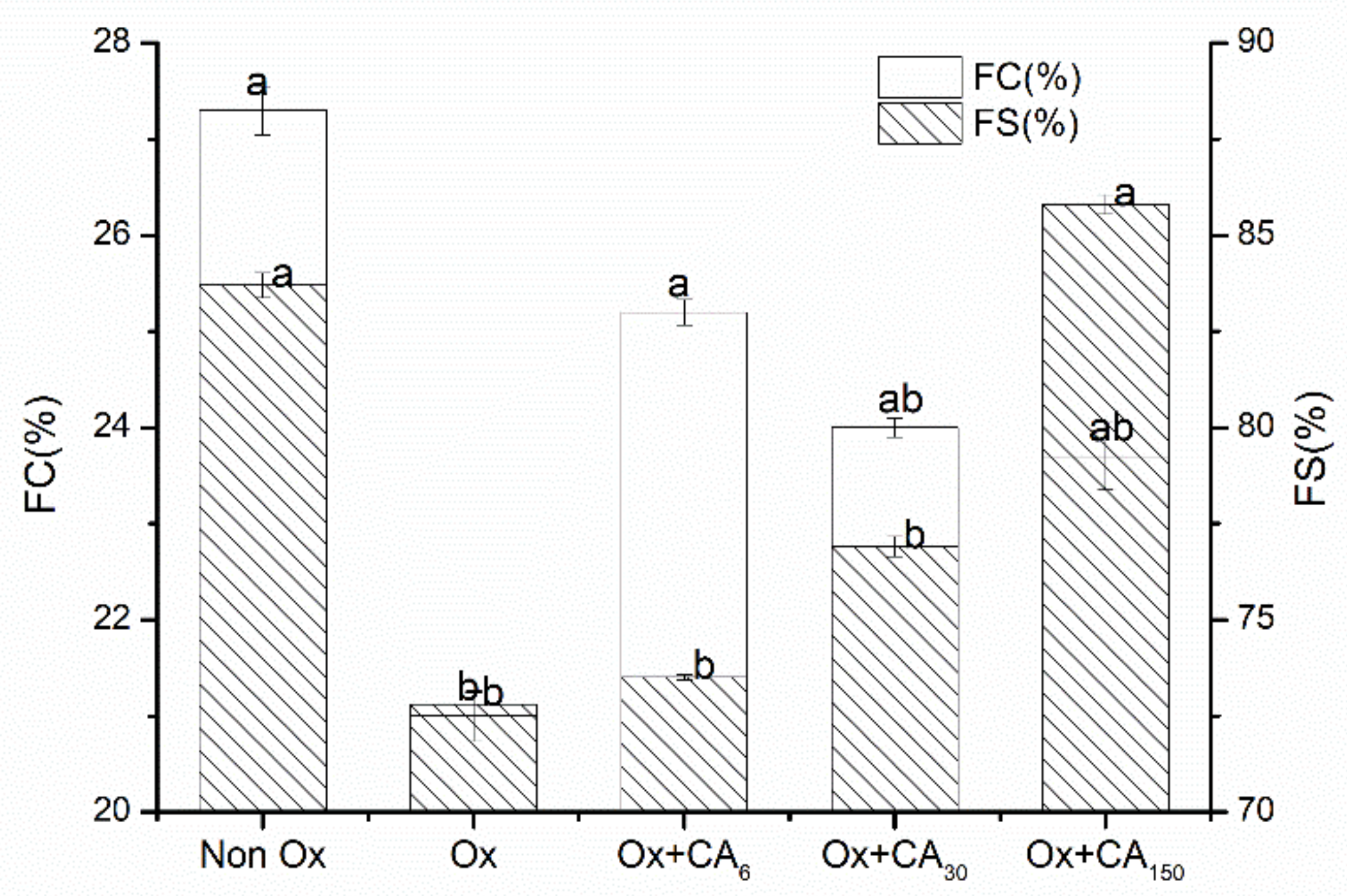
2.5. Effect of Chlorogenic Acid on Functional Properties of Oxidized MP
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Preparation of MP
3.3. Oxidative Treatment with Chlorogenic Acid (CA)
3.4. Measurement of Physicochemical Properties in MP
3.4.1. Determination of Carbonyl Content
3.4.2. Determination of Free Amino Group (–NH2) Content
3.4.3. Determination of Total Sulfhydryl Content (–SH)
3.4.4. Determination of Surface Hydrophobicity
3.4.5. Dityrosine Measurement
3.4.6. Determination of Endogenous Fuorescence Spectra
3.4.7. FT–IR
3.4.8. SDS–polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.5. Measurement of Functional Properties of MP
3.5.1. Protein Solubility
3.5.2. Emulsification Properties
3.5.3. Foaming Properties
3.5.4. Determination of Gel Strength and Cooking Loss
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estevez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [PubMed]
- Kanner, J. Oxidative processes in meat and meat products: Quality implications. Meat Sci. 1994, 36, 169–189. [Google Scholar] [CrossRef]
- Lee, D.S.; Woo, J.Y.; Ahn, C.B.; Je, J.Y. Chitosan-hydroxycinnamic acid conjugates: Preparation, antioxidant and antimicrobial activity. Food Chem. 2014, 148, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Wanasundara, P.K. Phenolic antioxidants. Crit. Rev. Food Sci. Nutr. 1992, 32, 67–103. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; True, A.D.; Chen, J.; Xiong, Y.L. Dual Role (Anti- and Pro-oxidant) of Gallic Acid in Mediating Myofibrillar Protein Gelation and Gel in Vitro Digestion. J. Agric. Food Chem. 2016, 64, 3054–3061. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Lei, F.; Liang, C.; Yuan, F.; Gao, Y. Covalent complexation and functional evaluation of (-)-epigallocatechin gallate and alpha-lactalbumin. Food Chem. 2014, 150, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Esposito, D.; Lila, M.A.; Foegeding, E.A. Formation of whey protein-polyphenol meso-structures as a natural means of creating functional particles. Food Funct. 2016, 7, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.G.; Sun, C.X.; Yang, W.; Yuan, F.; Gao, Y.X. Structural characterization and functional evaluation of lactoferrin-polyphenol conjugates formed by free-radical graft copolymerization. RSC Adv. 2015, 5, 15641–15651. [Google Scholar] [CrossRef]
- Balange, A.K.; Benjakul, S. Effect of oxidised phenolic compounds on the gel property of mackerel (Rastrelliger kanagurta) surimi. LWT Food Sci. Technol. 2009, 42, 1059–1064. [Google Scholar] [CrossRef]
- Dulmaa, A.; Slynko, Y.V.; Gordon, N.Y.; Stolbunova, V.V.; Politov, D.V. Coregonus peled (Gmelin) transplanted into Ulaagchny Khar lake (western mongolia) showed no evidence of hybridization with other introduced Coregonus species. Contemp. Probl. Ecol. 2016, 9, 172–176. [Google Scholar]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [PubMed]
- Liu, G.; Xiong, Y.L.; Butterfield, D.A. Chemical, physical, and gel-forming properties of oxidized myofibrils and whey- and soy-protein isolates. J. Food Sci. 2000, 65, 811–818. [Google Scholar]
- Jongberg, S.; Gislason, N.E.; Lund, M.N.; Skibsted, L.H.; Waterhouse, A.L. Thiol-quinone adduct formation in myofibrillar proteins detected by LC-MS. J. Agric. Food Chem. 2011, 59, 6900–6905. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Lee, J.H.; Won, M.; Song, K.B. Antioxidant activities of distiller dried grains with solubles as protein films containing tea extracts and their application in the packaging of pork meat. Food Chem. 2016, 196, 174–179. [Google Scholar] [PubMed]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid-protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Gillies, D.; Wagner, G.; Howell, N.K. ESR and NMR spectroscopy studies on protein oxidation and formation of dityrosine in emulsions containing oxidised methyl linoleate. Food Chem. Toxicol. 2006, 44, 1385–1392. [Google Scholar] [PubMed]
- Komath, S.S.; Swamy, M.J. Fluorescence quenching, time-resolved fluorescence and chemical modification studies on the tryptophan residues of snake gourd (Trichosanthes anguina) seed lectin. J. Photochem. Photobiol. B Biol. 1999, 50, 108–118. [Google Scholar]
- Von Staszewski, M.; Jara, F.L.; Ruiz, A.L.T.G.; Jagus, R.J.; Carvalho, J.E.; Pilosof, A.M.R. Nanocomplex formation between β-lactoglobulin or caseinomacropeptide and green tea polyphenols: Impact on protein gelation and polyphenols antiproliferative activity. J. Funct. Foods 2012, 4, 800–809. [Google Scholar] [CrossRef]
- Lefevre, F.; Fauconneau, B.; Thompson, J.W.; Gill, T.A. Thermal denaturation and aggregation properties of Atlantic salmon myofibrils and myosin from white and red muscles. J. Agric. Food Chem. 2007, 55, 4761–4770. [Google Scholar]
- Li, H.; Liu, B.L.; Gao, L.Z.; Chen, H.L. Studies on bullfrog skin collagen. Food Chem. 2004, 84, 65–69. [Google Scholar]
- Subirade, M.; Kelly, I.; Gueguen, J.; Pezolet, M. Molecular basis of film formation from a soybean protein: Comparison between the conformation of glycinin in aqueous solution and in films. Int. J. Biol. Macromol. 1998, 23, 241–249. [Google Scholar] [CrossRef]
- Estevez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Chen, L.; Xu, X.; Zhou, G.; Li, Z.; Feng, X. Dose-dependent effects of rosmarinic acid on formation of oxidatively stressed myofibrillar protein emulsion gel at different NaCl concentrations. Food Chem. 2018, 243, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.L.; Agyare, K.K.; Addo, K. Hydrolyzed wheat gluten suppresses transglutaminase-mediated gelation but improves emulsification of pork myofibrillar protein. Meat Sci. 2008, 80, 535–544. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Decker, E.A. Lipid oxidation in oil-in-water emulsions: Impact of molecular environment on chemical reactions in heterogeneous food systems. J. Food Sci. 2000, 65, 1270–1282. [Google Scholar]
- Erlejman, A.G.; Verstraeten, S.V.; Fraga, C.G.; Oteiza, P.I. The interaction of flavonoids with membranes: Potential determinant of flavonoid antioxidant effects. Free Radic. Res. 2004, 38, 1311–1320. [Google Scholar]
- Yuliana, M.; Truong, C.T.; Huynh, L.H.; Ho, Q.P.; Ju, Y.-H. Isolation and characterization of protein isolated from defatted cashew nut shell: Influence of pH and NaCl on solubility and functional properties. LWT Food Sci. Technol. 2014, 55, 621–626. [Google Scholar] [CrossRef]
- Davis, J.P.; Foegeding, E.A. Foaming and interfacial properties of polymerized whey protein isolate. J. Food Sci. 2004, 69, C404–C410. [Google Scholar] [CrossRef]
- Sarker, D.K.; Wilde, P.J.; Clark, D.C. Control of surfactant-induced destabilization of foams through polyphenol-mediated protein-protein interactions. J. Agric. Food Chem. 1995, 43, 295–300. [Google Scholar] [CrossRef]
- Balange, A.; Benjakul, S. Enhancement of gel strength of bigeye snapper (Priacanthus tayenus) surimi using oxidised phenolic compounds. Food Chem. 2009, 113, 61–70. [Google Scholar] [CrossRef]
- Jongberg, S.; Terkelsen, L.S.; Miklos, R.; Lund, M.N. Green tea extract impairs meat emulsion properties by disturbing protein disulfide cross-linking. Meat Sci 2015, 100, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Liu, X.C.; Zhang, Y.M.; Wang, H.; Luo, Y.K. Effects of Chilling and Partial Freezing on Rigor Mortis Changes of Bighead Carp (Aristichthys nobilis) Fillets: Cathepsin Activity, Protein Degradation and Microstructure of Myofibrils. J. Food Sci. 2015, 80, C2725–C2731. [Google Scholar] [PubMed]
- Oliver, C.N.; Ahn, B.W.; Moerman, E.J.; Goldstein, S.; Stadtman, E.R. Age-related changes in oxidized proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [PubMed]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar]
- Lv, Y.; Chen, L.; Wu, H.; Xu, X.; Zhou, G.; Zhu, B.; Feng, X. (-)-Epigallocatechin-3-gallate-mediated formation of myofibrillar protein emulsion gels under malondialdehyde-induced oxidative stress. Food Chem. 2019, 285, 139–146. [Google Scholar] [CrossRef] [PubMed]
- LI-Chan, E.; Nakai, S.; Wood, D.F. Relationship Between Functional (Fat Binding, Emulsifying) and Physicochemical Properties of Muscle Proteins. Effects of Heating, Freezing, pH and Species. Food Sci. 1985, 50, 1034–1040. [Google Scholar] [CrossRef]
- Davies, K.J.; Delsignore, M.E.; Lin, S.W. Protein damage and degradation by oxygen radicals. II. Modification of amino acids. J. Biol. Chem. 1987, 262, 9902–9907. [Google Scholar] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Joo, S.T.; Kauffman, R.G.; Kim, B.C.; Park, G.B. The relationship of sarcoplasmic and myofibrillar protein solubility to colour and water-holding capacity in porcine longissimus muscle. Meat Sci. 1999, 52, 291–297. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar]
- Poole, S.; West, S.I.; Walters, C.L. Protein–protein interactions: Their importance in the foaming of heterogeneous protein systems. J. Sci. Food Agric. 1984, 35, 701–711. [Google Scholar] [CrossRef]
Sample Availability: Samples of the raw material of Coregonus peled are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Free Amino (nM/mg protein) | Carbonyl (nM/mg protein) | Thiol Groups (nM/mg protein) | Surface Hydrophobicity | Dimeric Tyrosine (A.U.) |
|---|---|---|---|---|---|
| Control | 50.74 ± 0.19 a | 0.24 ± 0.035 c | 90.05 ± 1.93 a | 138.6 ± 2.99 b | 123.58 ± 6.52 c |
| Oxidized | 46.25 ± 0.16 d | 1.01 ± 0.008 a | 81.36 ± 0.65 b | 148.09 ± 5.06 a | 143.69 ± 3.62 a |
| Ox + CA6 | 49.65 ± 0.15 b | 0.46 ± 0.057 bc | 82.39 ± 1.38 b | 111.66 ± 2.7 d | 126.92 ± 5.01 c |
| Ox + CA30 | 48.35 ± 0.19 c | 0.49 ± 0.086 b | 81.49 ± 0.42 b | 125.59 ± 8.1 c | 134.17 ± 4.21 b |
| Ox + CA150 | 43.43 ± 0.25 e | 0.52 ± 0.054 bc | 78.7 ± 1.05 c | 135.11 ± 1.77 b | 144.37 ± 7.8 a |
| Samples | Solubility (%) | Gel Strength (N) | Cooking Loss (%) |
|---|---|---|---|
| Control | 67.8 ± 0.4 a | 0.38 ± 0.12 ab | 41.54 ± 0.34 c |
| Oxidized | 51.8 ± 0.8 d | 0.34 ± 0.14 ab | 42.31 ± 7.04 b |
| Ox + CA6 | 59.8 ± 2 b | 0.51 ± 0.20 a | 40.04 ± 0.84 d |
| Ox + CA30 | 58.9 ± 0.2 b | 0.47 ± 0.06 ab | 40.38 ± 5.97 d |
| Ox + CA150 | 57.5 ± 0.4 c | 0.23 ± 0.06 b | 50.55 ± 5.50 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Qiu, H.; Deng, X.; Mao, X.; Guo, X.; Xu, C.; Zhang, J. Effect of Chlorogenic Acid on the Physicochemical and Functional Properties of Coregonus Peled Myofibrillar Protein through Hydroxyl Radical Oxidation. Molecules 2019, 24, 3205. https://doi.org/10.3390/molecules24173205
Guo X, Qiu H, Deng X, Mao X, Guo X, Xu C, Zhang J. Effect of Chlorogenic Acid on the Physicochemical and Functional Properties of Coregonus Peled Myofibrillar Protein through Hydroxyl Radical Oxidation. Molecules. 2019; 24(17):3205. https://doi.org/10.3390/molecules24173205
Chicago/Turabian StyleGuo, Xin, Hengheng Qiu, Xiaorong Deng, Xiaoying Mao, Xiaobing Guo, Chengjian Xu, and Jian Zhang. 2019. "Effect of Chlorogenic Acid on the Physicochemical and Functional Properties of Coregonus Peled Myofibrillar Protein through Hydroxyl Radical Oxidation" Molecules 24, no. 17: 3205. https://doi.org/10.3390/molecules24173205
APA StyleGuo, X., Qiu, H., Deng, X., Mao, X., Guo, X., Xu, C., & Zhang, J. (2019). Effect of Chlorogenic Acid on the Physicochemical and Functional Properties of Coregonus Peled Myofibrillar Protein through Hydroxyl Radical Oxidation. Molecules, 24(17), 3205. https://doi.org/10.3390/molecules24173205