Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors

 ,
, 
 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Proximate Composition
2.2. Amino Acid Profile
2.3. Lipid Profile
2.4. Carbohydrates Composition
2.5. Pigment Profile
2.6. Vitamin Profile
2.7. Mineral Composition
2.8. Antioxidant Activity
2.9. Microbiological Evaluation
2.10. Toxicological Evaluation
3. Materials and Methods
3.1. Microalgae Growth
3.2. Proximate Composition
3.3. Amino Acid Profile
3.4. Fatty Acid Profile
3.5. Analysis of Carbohydrates
3.6. Determination of Pigments
3.7. Determination of Vitamins
3.8. Mineral Analysis
3.9. Evaluation of In Vitro Biological Activities
3.10. Microbiological Evaluation
3.11. Toxicological Evaluation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Austic, R.E.; Mustafa, A.; Jung, B.; Gatrell, S.; Lei, X.G. Potential and Limitation of a New Defatted Diatom Microalgal Biomass in Replacing Soybean Meal and Corn in Diets for Broiler Chickens. J. Agric. Food Chem. 2013, 61, 7341–7348. [Google Scholar] [CrossRef] [PubMed]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed]
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as healthy ingredients for functional food: a review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef] [PubMed]
- Caporgno, M.P.; Mathys, A. Trends in Microalgae Incorporation Into Innovative Food Products With Potential Health Benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Custódio, L.; Soares, F.; Pereira, H.; Barreira, L.; Vizetto-Duarte, C.; Rodrigues, M.J.; Rauter, A.P.; Alberício, F.; Varela, J. Fatty acid composition and biological activities of Isochrysis galbana T.-ISO, Tetraselmis sp. and Scenedesmus sp.: Possible application in the pharmaceutical and functional food industries. J. Appl. Phycol. 2014, 26, 151–161. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential Industrial Applications and Commercialization of Microalgae in the Functional Food and Feed Industries: A Short Review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M. High-value products from microalgae—Their development and commercialization. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Barreira, L.; Pereira, H.; Gangadhar, K.N.; Custódio, L.; Varela, J. Medicinal Effects of Microalgae-Derived Fatty Acids. In Handbook of Marine Microalgae; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 209–231. [Google Scholar]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 1405. [Google Scholar] [CrossRef]
- Lauritano, C.; Ianora, A. Marine Organisms with Anti-Diabetes Properties. Mar. Drugs 2016, 14, 220. [Google Scholar] [CrossRef]
- Levy-Ontman, O.; Huleihel, M.; Hamias, R.; Wolak, T.; Paran, E. An anti-inflammatory effect of red microalga polysaccharides in coronary artery endothelial cells. Atheroscler. 2017, 264, 11–18. [Google Scholar] [CrossRef] [PubMed]
- de Vera, C.; Díaz Crespín, G.; Hernández Daranas, A.; Montalvão Looga, S.; Lillsunde, K.E.; Tammela, P.; Perälä, M.; Hongisto, V.; Virtanen, J.; Rischer, H.; et al. Marine microalgae: Promising source for new bioactive compounds. Mar. Drugs 2018, 16, 317. [Google Scholar] [CrossRef] [PubMed]
- Andrade, K.A.M.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custodio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated Fatty Acids of Marine Macroalgae: Potential for Nutritional and Pharmaceutical Applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef]
- Custódio, L.; Justo, T.; Silvestre, L.; Barradas, A.; Vizetto-Duarte, C.; Pereira, H.; Barreira, L.; Rauter, A.P.; Albericio, F.; Varela, J. Microalgae of different phyla display antioxidant, metal chelating and acetylcholinesterase inhibitory activities. Planta Medica 2012, 78, 134–140. [Google Scholar] [CrossRef]
- Parenti, F.; Righi, V.; Schenetti, L.; Mucci, A. Mycosporine-like Amino Acids and Other Phytochemicals Directly Detected by High-Resolution NMR on Klamath ( Aphanizomenon flos-aquae ) Blue-Green Algae. J. Agric. Food Chem. 2016, 64, 6708–6715. [Google Scholar]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The green microalga Tetraselmis suecica reduces oxidative stress and induces repairing mechanisms in human cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef]
- Ji, C.; Cao, X.; Yao, C.; Xue, S.; Xiu, Z. Protein–protein interaction network of the marine microalga Tetraselmis subcordiformis: prediction and application for starch metabolism analysis. J. Ind. Microbiol. Biotechnol. 2014, 41, 1287–1296. [Google Scholar] [CrossRef]
- Schwenzfeier, A.; Wierenga, P.A.; Gruppen, H. Isolation and characterization of soluble protein from the green microalgae Tetraselmis sp. Bioresour. Technol. 2011, 102, 9121–9127. [Google Scholar] [CrossRef]
- Pereira, H.; Gangadhar, K.N.; Schulze, P.S.C.; Santos, T.; De Sousa, C.B.; Schueler, L.M.; Custódio, L.; Malcata, F.X.; Gouveia, L.; Varela, J.C.S.; et al. Isolation of a euryhaline microalgal strain, Tetraselmis sp. CTP4, as a robust feedstock for biodiesel production. Sci. Rep. 2016, 6, 35663. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef] [PubMed]
- Schulze, P.S.C.; Carvalho, C.F.M.; Pereira, H.; Gangadhar, K.N.; Schüler, L.M.; Santos, T.; Varela, J.; Barreira, L. Urban wastewater treatment by Tetraselmis sp. CTP4 (Chlorophyta). Bioresour. Technol. 2017, 223, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Bernaerts, T.M.; Gheysen, L.; Kyomugasho, C.; Kermani, Z.J.; Vandionant, S.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Comparison of microalgal biomasses as functional food ingredients: Focus on the composition of cell wall related polysaccharides. Algal Res. 2018, 32, 150–161. [Google Scholar] [CrossRef]
- Tokuşoglu, Ö.; Ünal, M.K. Biomass nutrient profiles of three microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrisis galbana. J. Food Sci. 2006, 68, 1144–1148. [Google Scholar] [CrossRef]
- Molina, E.; Contreras, A.; Martinez, M.E.; Sanchez, S.; Garcia, F. Growth and biochemical composition with emphasis on the fatty acids of Tetraselmis sp. Appl. Microbiol. Biotechnol. 1991, 36, 21–25. [Google Scholar] [CrossRef]
- Tulli, F.; Zittelli, G.C.; Giorgi, G.; Poli, B.M.; Tibaldi, E.; Tredici, M.R. Effect of the Inclusion of Dried Tetraselmis suecica on Growth, Feed Utilization, and Fillet Composition of European Sea Bass Juveniles Fed Organic Diets. J. Aquat. Food Prod. Technol. 2012, 21, 188–197. [Google Scholar] [CrossRef]
- United States Department of Agriculture (2018) Basic Report: 11667, Seaweed, Spirulina, Dried. Available online: https://ndb.nal.usda.gov/ndb/foods/show/11667 (accessed on 5 July 2019).
- Ssepuuya, G.; Mukisa, I.M.; Nakimbugwe, D. Nutritional composition, quality, and shelf stability of processed Ruspolia nitidula (edible grasshoppers). Food Sci. Nutr. 2016, 5, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Dunstan, G.A.; Volkman, J.K.; Jeffrey, S.W.; Barretta, S.M. Biochemical composition of microalgae from the green algal classes Chlorophyceae and Prasinophyceae. 2. Lipid classes and fatty acids. J. Exp. Mar. Bio. Ecol. 1992, 161, 115–134. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Boil. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Kermanshahi-Pour, A.; Sommer, T.J.; Anastas, P.T.; Zimmerman, J.B. Enzymatic and acid hydrolysis of Tetraselmis suecica for polysaccharide characterization. Bioresour. Technol. 2014, 173, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Becker, B.; Melkonian, M.; Kamerling, J.P. The cell wall (theca) of Tetraselmis striata (chlorophyta): macromolecular composition and structural elements of the complex polysaccharides. J. Phycol. 1998, 34, 779–787. [Google Scholar] [CrossRef]
- Shekharam, K.M.; Venkataraman, L.; Salimath, P. Carbohydrate composition and characterization of two unusual sugars from the blue green alga Spirulina platensis. Phytochem. 1987, 26, 2267–2269. [Google Scholar] [CrossRef]
- Baker, R.; Günther, C. The role of carotenoids in consumer choice and the likely benefits from their inclusion into products for human consumption. Trends Food Sci. Technol. 2004, 15, 79959283. [Google Scholar] [CrossRef]
- Balder, H.F.; Vogel, J.; Jansen, M.C.; Weijenberg, M.P.; Westenbrink, S.; Van Der Meer, R.; Goldbohm, R.A.; Brandt, P.A.V.D. Heme and Chlorophyll Intake and Risk of Colorectal Cancer in the Netherlands Cohort Study. Cancer Epidemiol. Biomark. Prev. 2006, 15, 717–725. [Google Scholar] [CrossRef]
- Cordero, B.F.; Obraztsova, I.; Couso, I.; León, R.; Vargas, M.A.; Rodríguez, H. Enhancement of Lutein Production in Chlorella sorokiniana (Chorophyta) by Improvement of Culture Conditions and Random Mutagenesis. Mar. Drugs 2011, 9, 1607–1624. [Google Scholar] [CrossRef]
- Ahmed, F.; Fanning, K.; Netzel, M.; Schenk, P.M. Induced carotenoid accumulation in Dunaliella salina and Tetraselmis suecica by plant hormones and UV-C radiation. Appl. Microbiol. Biotechnol. 2015, 99, 9407–9416. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, L.; Qiu, S.; Ge, S. Determination of Microalgal Lipid Content and Fatty Acid for Biofuel Production. BioMed Res. Int. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Fábregas, J.; Herrero, C. Vitamin content of four marine microalgae. Potential use as source of vitamins in nutrition. J. Ind. Microbiol. Biotechnol. 1990, 5, 259–263. [Google Scholar] [CrossRef]
- Brown, M.; Mular, M.; Miller, I.; Farmer, C.; Trenerry, C. The vitamin content of microalgae used in aquaculture. Environ. Boil. Fishes 1999, 11, 247–255. [Google Scholar]
- Carballo-Cárdenas, E.C.; Tuan, P.M.; Janssen, M.; Wijffels, R.H. Vitamin E (α-tocopherol) production by the marine microalgae Dunaliella tertiolecta and Tetraselmis suecica in batch cultivation. Biomol. Eng. 2003, 20, 139–147. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional value and use of microalgae in aquaculture. In Avances En Nutrición Acuícola, proceeding of Memorias Del VI Simposium Internacional de Nutrición Acuícola; Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Gaxiola-Cortés, M.G., Simoes, N., Eds.; Universidad Autónoma de Nuevo León: Monterrey, México, 2002; pp. 281–292. [Google Scholar]
- Lešková, E.; Kubíková, J.; Kováčiková, E.; Košická, M.; Porubská, J.; Holčíková, K. Vitamin losses: Retention during heat treatment and continual changes expressed by mathematical models. J. Food Compost. Anal. 2006, 19, 252–276. [Google Scholar] [CrossRef]
- Gojkovic, Ž.; Vilchez, C.; Torronteras, R.; Vigara, J.; Gómez-Jacinto, V.; Janzer, N.; Gómez-Ariza, J.-L.; Márova, I.; Garbayo, I. Effect of Selenate on Viability and Selenomethionine Accumulation ofChlorella sorokinianaGrown in Batch Culture. Sci. World J. 2014, 2014, 1–13. [Google Scholar] [CrossRef]
- Viña, J. Molecular bases of the treatment of Alzheimer’s disease with antioxidants: prevention of oxidative stress. Mol. Asp. Med. 2004, 25, 117–123. [Google Scholar] [CrossRef]
- Lanigan, R.S.; A Yamarik, T. Final Report on the Safety Assessment of BHT. Int. J. Toxicol. 2002, 21, 19–94. [Google Scholar]
- Megías, C.; Pastor-Cavada, E.; Torres-Fuentes, C.; Girón-Calle, J.; Alaiz, M.; Juan, R.; Pastor, J.; Vioque, J. Chelating, antioxidant and antiproliferative activity of Vicia sativa polyphenol extracts. Eur. Food Res. Technol. 2009, 230, 353–359. [Google Scholar] [CrossRef]
- Nunes, C.; Silva, L.; Fernandes, A.P.; Guiné, R.P.; Domingues, M.R.M.; Coimbra, M.A.; Domingues, M.R. Occurrence of cellobiose residues directly linked to galacturonic acid in pectic polysaccharides. Carbohydr. Polym. 2012, 87, 620–626. [Google Scholar] [CrossRef]
- Oliveira, C.; Ferreira, A.S.; Novoa-Carballal, R.; Nunes, C.C.; Pashkuleva, I.; Neves, N.M.; Coimbra, M.A.; Reis, R.L.; Martins, A.; Silva, T.H. The key role of sulfation and branching on fucoidan antitumor activity. Macromol. Biosci. 2017, 17, 1600340. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.; Jeffrey, S.; Mantoura, R.; Llewellyn, C.; Bjornland, T.; Repeta, D.; Welschmeyer, N. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Soszynski, A.; Martins, A.; Rauter, A.P.; Neng, N.R.; Nogueira, J.M.; Varela, J.; Barreira, L.; Custódio, L. Unravelling the antioxidant potential and the phenolic composition of different anatomical organs of the marine halophyte Limonium algarvense. Ind. Crop. Prod. 2015, 77, 315–322. [Google Scholar] [CrossRef]
Sample Availability: Samples of industrially produced microalgal biomass of Tetraselmis sp. CTP4 are available from the authors. |


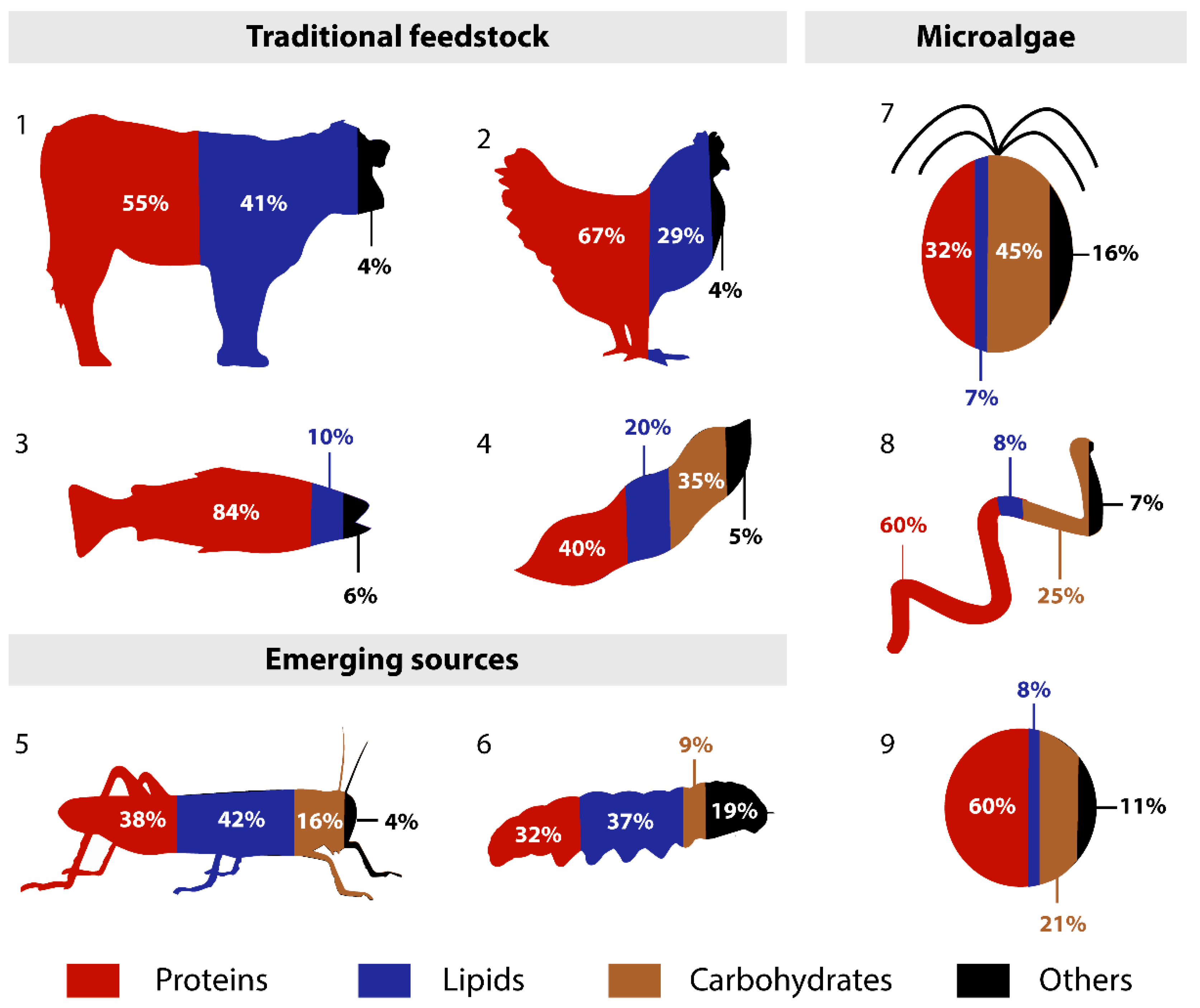{kind=link}
| Contents (g/100 g) | Tetraselmis sp. CTP4 | Tetraselmis chui1 | Tetraselmis suecica2 | Chlorellavulgaris3 | Arthrospira sp. 4 |
|---|---|---|---|---|---|
| Moisture | 3.88 ± 0.35 | <7 | n.r. | 4.9 | 4.7 |
| Protein | 31.20 ± 0.48 | 35–40 | 48.7 | 56.9 | 57.5 |
| Lipids | 7.04 ± 0.42 | 5–8 | 8.0 | 7.5 | 7.7 |
| Digestible carbohydrates | 18.08 ± 4.18 | 30–32 | 22.4 | 19.2 | 20.3 |
| Dietary fibres | 24.60 ± 3.85 | 2–3 | 3.4 | 0.5 | 3.6 |
| Ash | 15.20 ± 0.80 | 14–16 | 17.5 | 10.9 | 6.2 |
| Energy (kJ/100 g) | 1241 ± 49 | n.r. | n.r. | n.r. | 1213 |
| Energy (kcal/100 g) | 297 ± 12 | n.r. | n.r. | n.r. | 290 |
| Amino acid | Tetraselmis sp. CTP4 | Tetraselmischui1 | Chlorella sp. 1 | Arthrospira sp. 2 |
|---|---|---|---|---|
| Indispensable amino acids (IAA) | ||||
| Leucine | 2.28 ± 0.02 (8.83) | 7.5 | 9.9 | 8.5 |
| Isoleucine | 1.12 ± 0.02 (4.34) | 3.5 | 4.8 | 5.5 |
| Valine | 1.55 ± 0.02 (6.01) | 4.9 | 6.8 | 6.0 |
| Lysine | 1.70 ± 0.09 (6.59) | 5.7 | 6.6 | 5.2 |
| Threonine | 1.27 ± 0.05 (4.92) | 4.1 | 5.1 | 5.1 |
| Tryptophan | 0.37 ± 0.03 (1.43) | 2.4 | 1.0 | 1.6 |
| Methionine | 0.61 ± 0.03 (2.36) | 2.5 | 2.8 | 2.0 |
| Phenylalanine | 1.44 ± 0.08 (5.58) | 4.8 | 6.0 | 4.8 |
| Histidine | 0.04 ± 0.01 (0.15) | 1.6 | 2.3 | 1.9 |
| Cystine + Cysteine | 0.28 ± 0.01 (1.08) | 2.9 | n.r. | 1.1 |
| Total IAA | 10.7 (41.3) | 39.8 | 45.4 | 41.7 |
| Non-indispensable amino acids (NIAA) | ||||
| Alanine | 2.04 ± 0.06 (7.90) | 6.1 | 9.2 | 7.1 |
| Arginine | 1.70 ± 0.04 (6.59) | 9.6 | 7.0 | 10.0 |
| Aspartic acid (Asx) | 2.89 ± 0.02 (11.2) | 14.4 | 9.9 | 14.4 |
| Glutamic acid (Glx) | 3.64 ± 0.02 (14.1) | 12.3 | 12.4 | 5.3 |
| Glycine | 1.58 ± 0.06 (6.12) | 6.7 | 6.6 | 4.1 |
| Proline | 1.26 ± 0.03 (4.88) | 3.7 | 5.4 | 5.2 |
| Serine | 1.19 ± 0.08 (4.61) | 4.3 | 4.2 | 4.4 |
| Tyrosine | 0.85 ± 0.01 (3.29) | 3.1 | n.r. | 8.5 |
| Contents | Tetraselmis sp. CTP4 | Tetraselmis chui1 | Tetraselmis suecica1 | Arthrospira sp. 2 |
|---|---|---|---|---|
| Arabinose | 0.18 ± 0.01 (1.0) | 0.41 | 0.90 | n.r. |
| Xylose | 0.10 ± 0.01 (0.6) | n.d. | n.d. | 7.0 |
| Mannose | 1.33 ± 0.02 (6.5) | 1.8 | 3.0 | 9.3 |
| Galactose | 4.98 ± 0.03 (24.5) | 11.3 | 15.7 | 2.6 |
| Glucose | 13.68 ± 0.07 (67.3) | 84.7 | 74.8 | 54.4 |
| Rhamnose | v. | 0.04 | 0.97 | 22.3 |
| Ribose | v. | 1.8 | 4.5 | n.r. |
| Others | - | n.r. | n.r. | 4.3 |
| Pigments (mg/100 g) | Tetraselmis sp. CTP4 | Tetraselmis chui1 | Tetraselmis sp. M8 1 | Chlorella vulgaris2 |
|---|---|---|---|---|
| Chlorophyll a and b | 3531.2 ± 152.1 | n.r. | n.r. | 2600 |
| Violaxanthin | 130.8 ± 5.7 | 54.6 | 22.9 | n.r. |
| Antheraxanthin | n.d. | 20.1 | 12.6 | n.r. |
| Neoxanthin | 236.4 ± 11.9 | n.d. | n.d. | n.r. |
| Zeaxanthin | 10.8 ± 1.3 | n.d. | n.d. | 626 |
| Lutein | 225.6 ± 8.5 | 62.4 | 66.5 | 1011 |
| α-carotene | n.d. | 17.4 | 3.0 | 6.92 |
| β-carotene | 8.4 ± 0.7 | 94.1 | 105.7 | 8.26 |
| Vitamins | Tetraselmis sp. CTP4 | Tetraselmis sp. CS-362 1 | Chlorella vulgaris2 | Arthrospira sp.3 |
|---|---|---|---|---|
| A—Retinol (µg/100 g) | <4 | 220 | <20 | 29 |
| B1—Thiamin (mg/100 g) | 0.18 | 10.9 | 0.03 | 2.38 |
| B2—Riboflavin (mg/100 g) | 0.53 | 2.6 | 0.05 | 3.67 |
| B3—Niacin (mg/100 g) | 7.98 | n.r. | 0.10 | 12.8 |
| B5—Pantothenic Acid (mg/100 g) | 0.65 | n.r. | 0.08 | 3.48 |
| B6—Pyridoxal phosphate (mg/100 g) | 6.9 | 0.6 | 0.08 | 0.36 |
| B7—Biotin (mg/100 g) | n.d. | 0.13 | n.r. | n.r. |
| B9—Folic acid (µg/100 g) | 0.02 | 2000 | 30.6 | 94 |
| B12—Cianocobalamin (µg/100 g) | 7.8 | 195 | 0.10 | - |
| C—Ascorbic acid (mg/100 g) | 79.2 | 300 | <100 | 10.1 |
| E—Tocopherol (mg/100 g) | 20.28 | 7 | 6.57 | 5 |
| Sample | DPPH | ABTS | FRAP | CCA | ICA |
|---|---|---|---|---|---|
| Hexane | >10 | >10 | 1.1 ± 0.1 | >10 | >10 |
| Ethyl acetate | 2.6 ± 0.2 | 6.9 ± 0.4 | 0.5 ± 0.0 | >10 | >10 |
| Acetone | 4.9 ± 0.3 | 8.7 ± 0.3 | 0.3 ± 0.0 | >10 | 6.1 ± 0.2 |
| Ethanol | >10 | >10 | 1.1 ± 0.1 | >10 | >10 |
| Water | >10 | >10 | >10 | >10 | >10 |
| BHT | 0.14 ± 0.01 | 0.11 ± 0.01 | - | - | - |
| EDTA | - | - | - | 0.08 ± 0.01 | 0.03 ± 0.00 |
| Tetraselmis sp. CTP4 | |
|---|---|
| Aerobic plate total counts (30 °C) | 3.6 × 102 CFU/g |
| Enterobacteria | <1.0 × 101 CFU/g |
| Staphylococcus aureus | <1.0 × 101 CFU/g |
| Listeria monocytogenes | <1.0 × 101 CFU/g |
| Escherichia coli | <1.0 × 101 CFU/g |
| Salmonella spp. | Negative (25 g) |
| Pseudomonas spp. | <1.0 × 101 CFU/g |
| Vibrio spp. | Negative (25 g) |
| Yeasts (25 °C) | 1.0 × 102 CFU/g |
| Moulds (25 °C) | <1.0 × 101 CFU/g |
| Toxic Substances | Tetraselmis sp. CTP4 |
|---|---|
| Toxic metals | |
| Lead | <0.10 µg/g |
| Cadmium | 0.2 ± 0.0 µg/g |
| Mercury | <0.10 µg/g |
| Arsenic | <0.80 µg/g |
| Tin | <2.50 µg/g |
| Cyanotoxins | |
| Microcystin LR | n.d. |
| Microcystin RR | n.d. |
| Microcystin LA | n.d. |
| Cylindrospermopsin | n.d. |
| Mycotoxins | |
| Aflatoxins B1 and B2 | <0.5 ng/g |
| Aflatoxins G1 and G2 | <0.5 ng/g |
| Dioxins | |
| Benzo[a]pyrene | <0.5 ng/g |
| Benzo[a]anthracene | <0.5 ng/g |
| Benzo[b]fluoranthene | <0.5 ng/g |
| Chrysene | <0.5 ng/g |
| Other polycyclic aromatic hydrocarbons 1 | <0.5 ng/g |
| Pesticides | |
| Organochlorine pesticides 2 | <0.01 µg/g |
| Screening of >200 residues 3 | <0.01 µg/g |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, H.; Silva, J.; Santos, T.; Gangadhar, K.N.; Raposo, A.; Nunes, C.; Coimbra, M.A.; Gouveia, L.; Barreira, L.; Varela, J. Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors. Molecules 2019, 24, 3192. https://doi.org/10.3390/molecules24173192
Pereira H, Silva J, Santos T, Gangadhar KN, Raposo A, Nunes C, Coimbra MA, Gouveia L, Barreira L, Varela J. Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors. Molecules. 2019; 24(17):3192. https://doi.org/10.3390/molecules24173192
Chicago/Turabian StylePereira, Hugo, Joana Silva, Tamára Santos, Katkam N. Gangadhar, Ana Raposo, Cláudia Nunes, Manuel A. Coimbra, Luísa Gouveia, Luísa Barreira, and João Varela. 2019. "Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors" Molecules 24, no. 17: 3192. https://doi.org/10.3390/molecules24173192
APA StylePereira, H., Silva, J., Santos, T., Gangadhar, K. N., Raposo, A., Nunes, C., Coimbra, M. A., Gouveia, L., Barreira, L., & Varela, J. (2019). Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors. Molecules, 24(17), 3192. https://doi.org/10.3390/molecules24173192