Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling

 , ,
, ,
Abstract
1. Introduction
2. Results
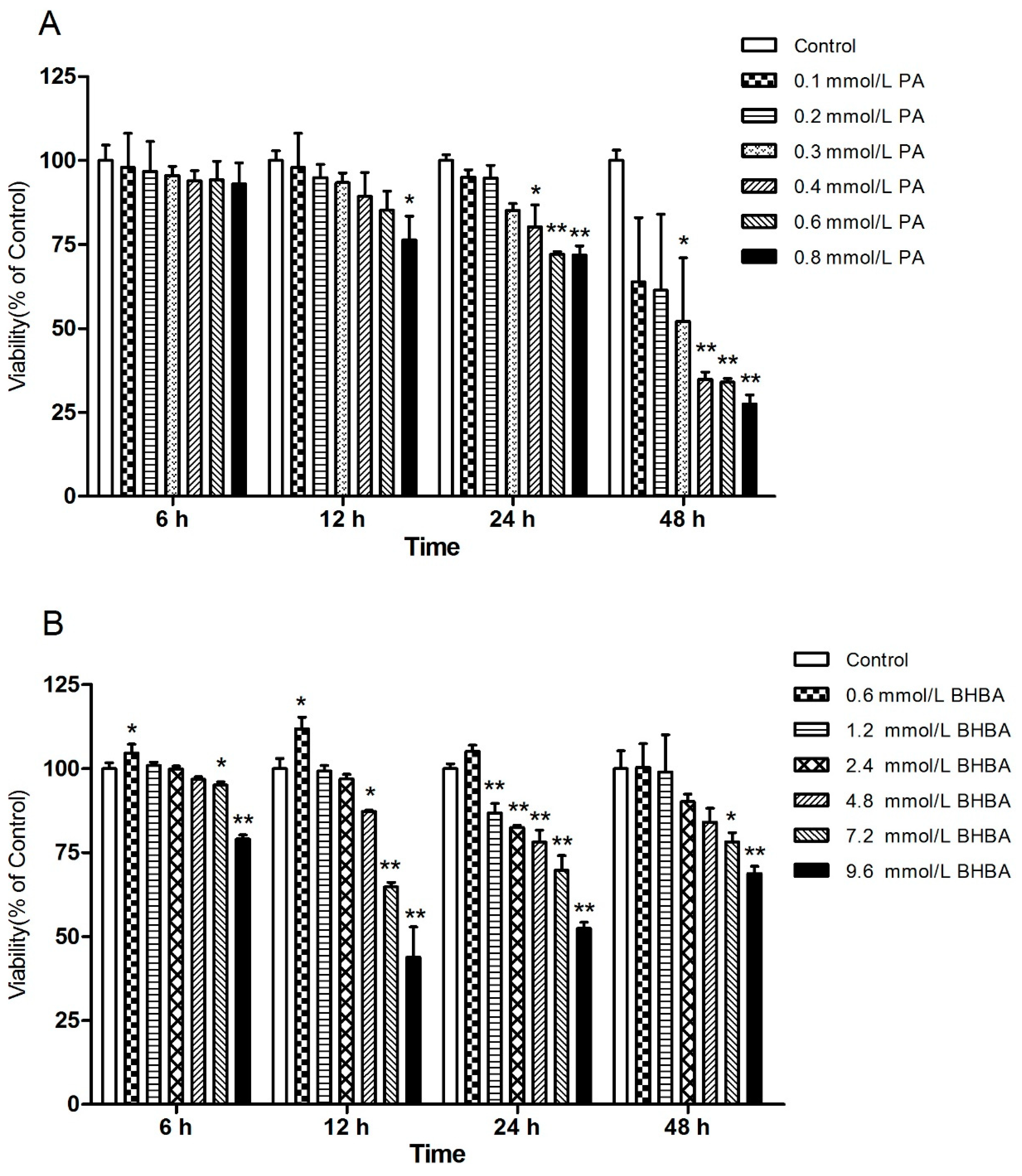
2.1. Effects of PA and BHBA on BEND Viability
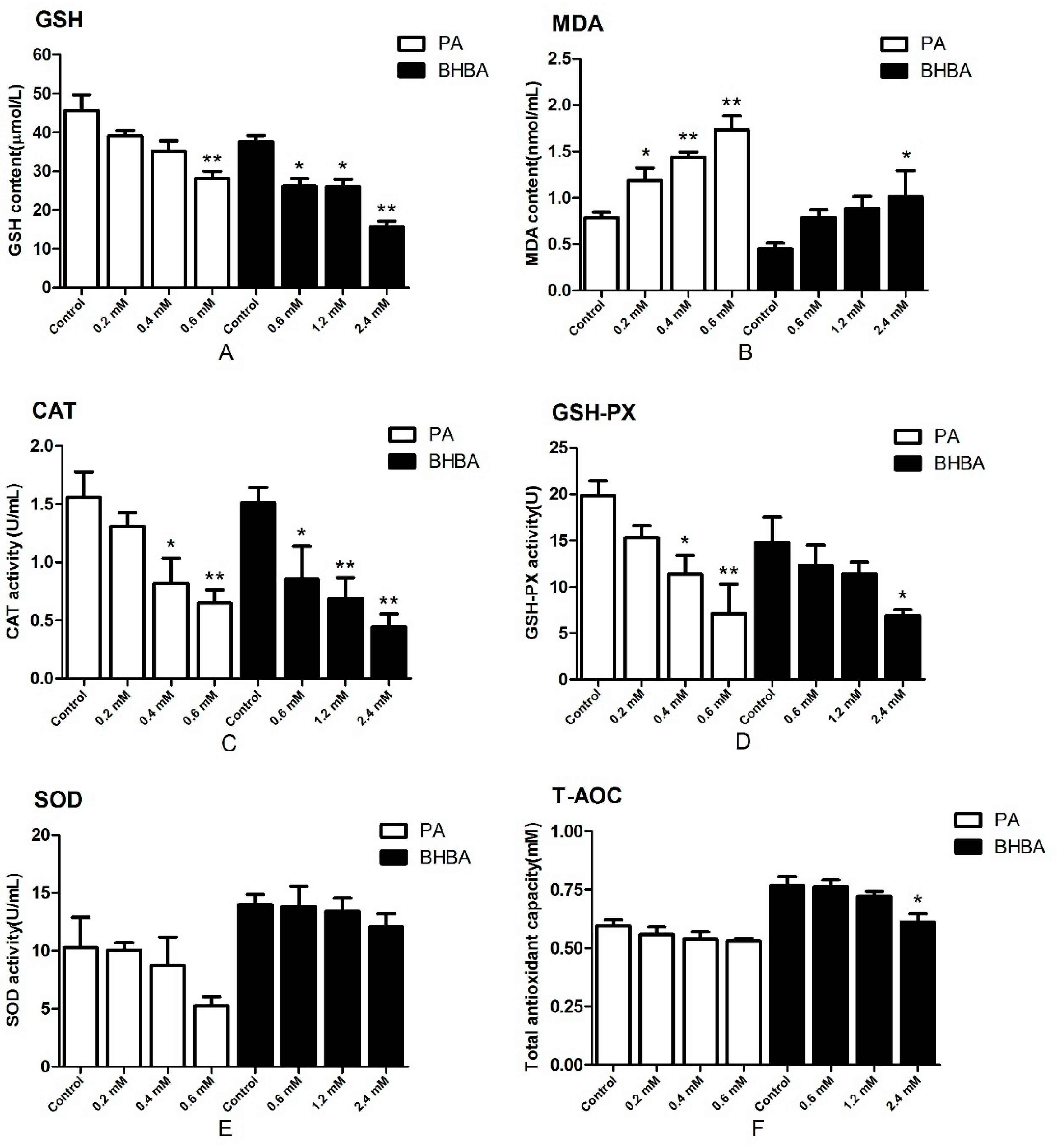
2.2. Oxidative Status
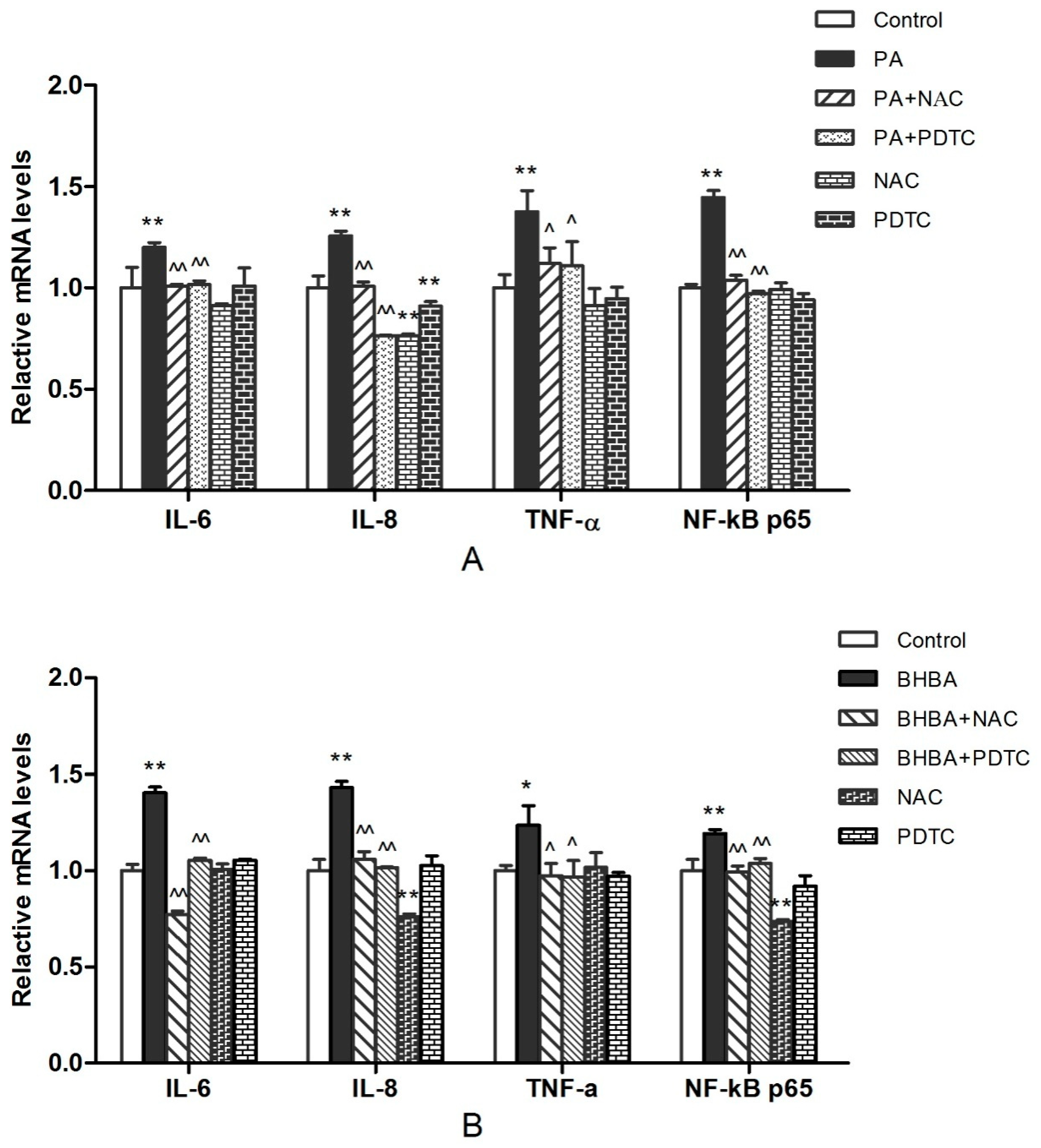
2.3. RT-PCR Analysis
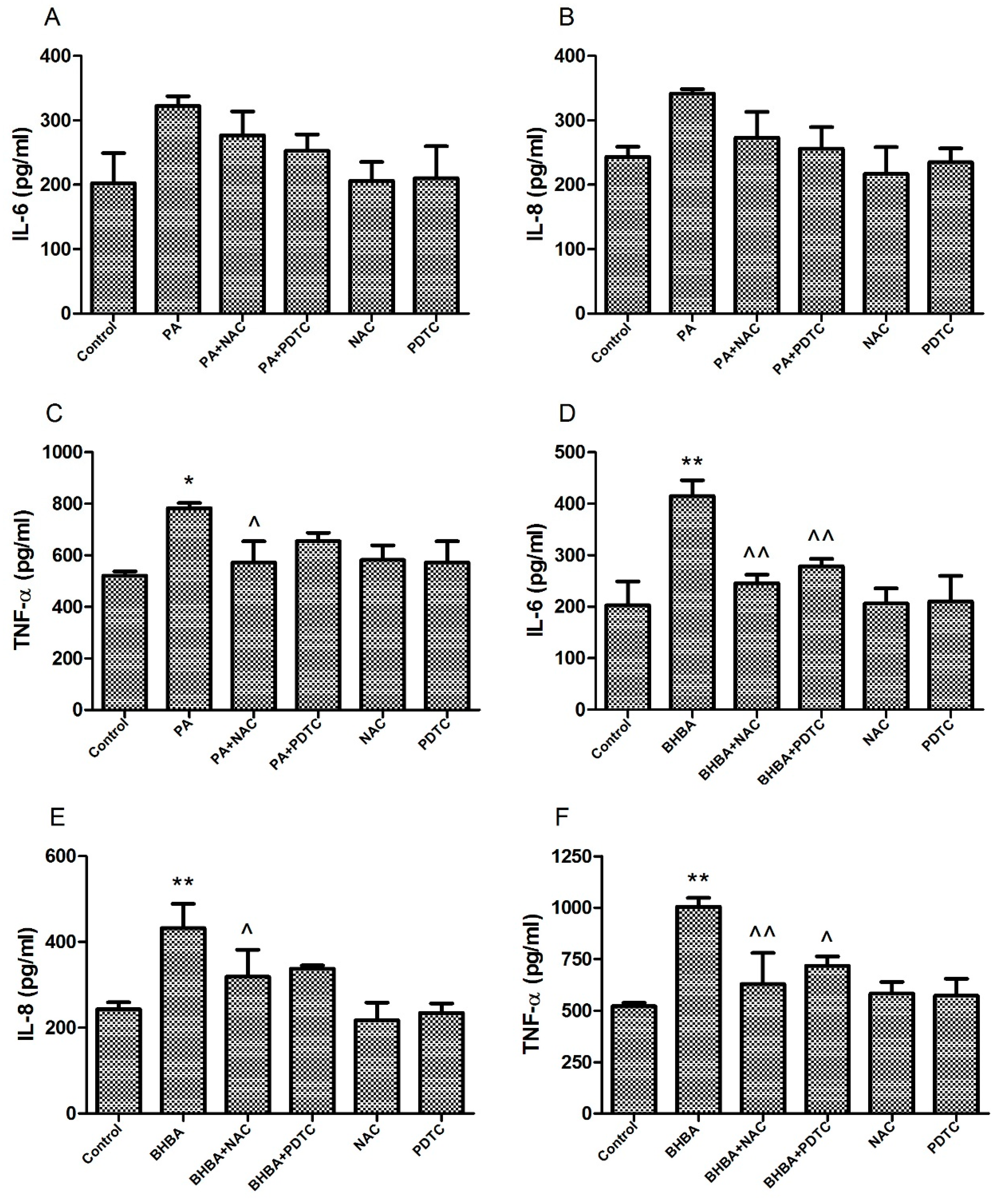
2.4. Levels of Pro-Inflammatory Factors in Cell Supernatants
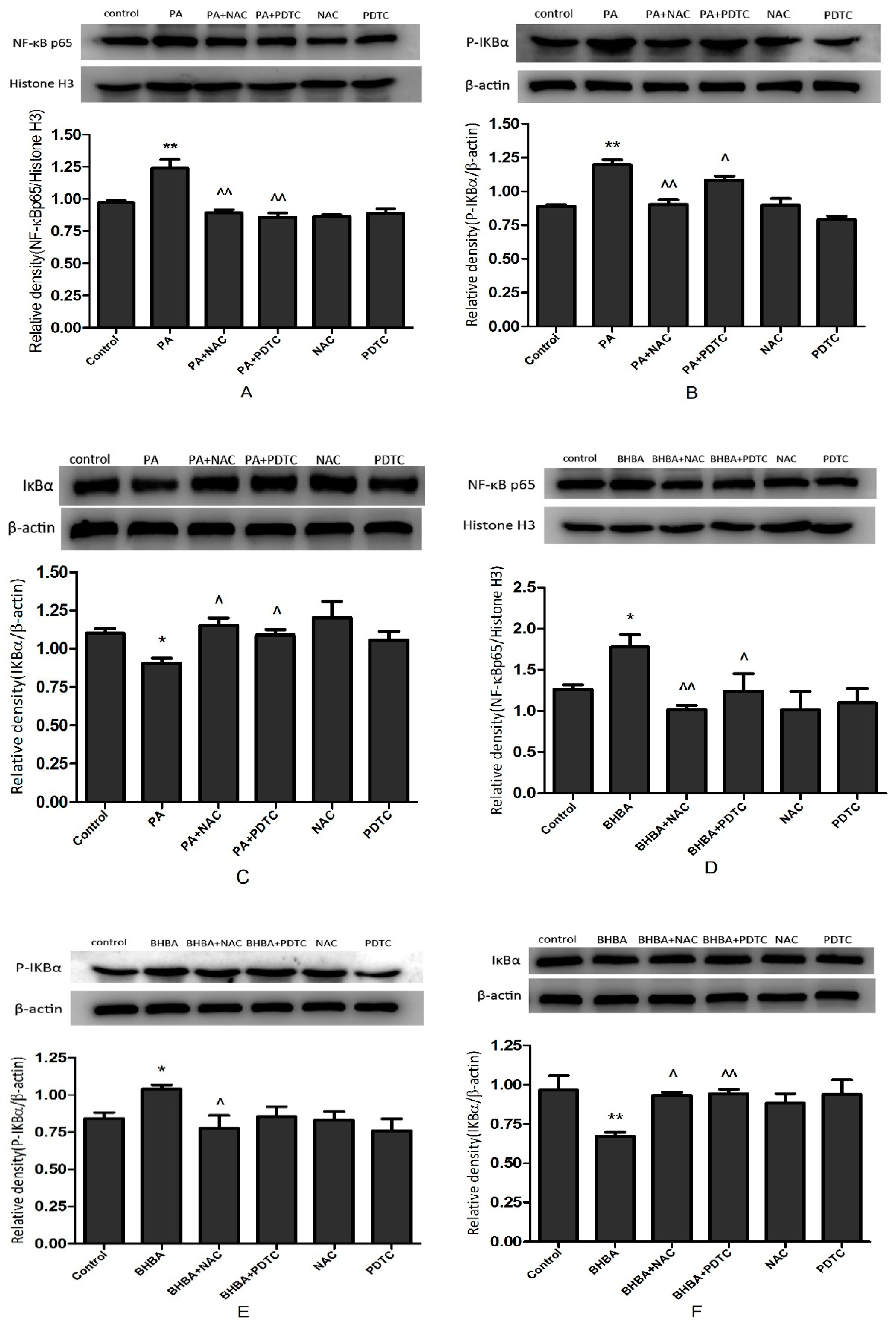
2.5. NF-κB Signal Protein Expression
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents and Antibodies
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Oxidative Stress Parameters Analysis
4.5. RNA Extraction and Quantitative Real-Time PCR
4.6. Enzyme-Linked Immunosorbent Assay
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vanholder, T.; Papen, J.; Bemers, R.; Vertenten, G.; Berge, A.C.B. Risk factors for subclinical and clinical ketosis and association with production parameters in dairy cows in the Netherlands. J. Dairy Sci. 2015, 98, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.; El-Ashker, M. Significance of insulin resistance and oxidative stress in dairy cattle with subclinical ketosis during the transition period. Trop. Anim. Health Pro. 2017, 49, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.A.; El-Khodery, S.A.; El-deeb, W.M.; Abou El-Amaiem, W.E. Ketosis in buffalo (Bubalus bubalis): clinical findings and the associated oxidative stress level. Trop. Anim. Health Pro. 2010, 42, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Xu, Q.; Chen, Y.; Yang, W.; Xia, C.; Yu, H.; Zhu, K.; Shen, T.; Zhang, Z. FGF-21: promising biomarker for detecting ketosis in dairy cows. Vet. Res. Commun. 2016, 40, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shu, S.; Xia, C.; Wang, P.; Sun, Y.; Xu, C.; Li, C. Mass spectral analysis of urine proteomic profiles of dairy cows suffering from clinical ketosis. Vet. Quart. 2015, 35, 133–141. [Google Scholar] [CrossRef]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy cows affected by ketosis show alterations in innate immunity and lipid and carbohydrate metabolism during the dry off period and postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- El-Deeb, W.M.; Younis, E.E. Clinical and biochemical studies on Theileria annulata in Egyptian buffaloes (Bubalus bubalis) with particular orientation to oxidative stress and ketosis relationship. Vet. Parasitol. 2009, 164, 301–305. [Google Scholar] [CrossRef]
- Hammon, D.S.; Evjen, I.M.; Dhiman, T.R.; Goff, J.P.; Walters, J.L. Neutrophil function and energy status in Holstein cows with uterine health disorders. Vet. Immunol. Immunop. 2006, 113, 21–29. [Google Scholar] [CrossRef]
- Contreras, G.A.; Raphael, W.; Mattmiller, S.A.; Gandy, J.; Sordillo, L.M. Nonesterified fatty acids modify inflammatory response and eicosanoid biosynthesis in bovine endothelial cells. J. Dairy Sci. 2012, 95, 5011–5023. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Osawa, T.; Dubuc, J. Reproductive tract defense and disease in postpartum dairy cows. Theriogenology 2011, 76, 1610–1618. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunop. 2009, 128, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Raboisson, D.; Mounie, M.; Khenifar, E.; Maigne, E. The economic impact of subclinical ketosis at the farm level: Tackling the challenge of over-estimation due to multiple interactions. Prev. Vet. Med. 2015, 122, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Lee, A.Y.; Park, S.; Kim, J.H.; Cho, M.H. Multiple pathways are involved in palmitic acid-induced toxicity. Food Chem. Toxicol. 2014, 67, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Lambertucci, R.H.; Leandro, C.G.; Vinolo, M.A.; Nachbar, R.T.; Dos Reis Silveira, L.; Hirabara, S.M.; Curi, R.; Pithon-Curi, T.C. The effects of palmitic acid on nitric oxide production by rat skeletal muscle: mechanism via superoxide and iNOS activation. Cell. Physiol. Biochem. 2012, 30, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhou, L.; Fan, Z.; Liu, S.; Fang, W. Palmitic acid, but not high-glucose, induced myocardial apoptosis is alleviated by N-acetylcysteine due to attenuated mitochondrial-derived ROS accumulation-induced endoplasmic reticulum stress. Cell Death Dis. 2018, 9, 568. [Google Scholar] [CrossRef]
- Jin, J.; Lu, Z.; Li, Y.; Cowart, L.A.; Lopes-Virella, M.F.; Huang, Y. Docosahexaenoic acid antagonizes the boosting effect of palmitic acid on LPS inflammatory signaling by inhibiting gene transcription and ceramide synthesis. PLoS ONE 2018, 13, e0193343. [Google Scholar] [CrossRef]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects on Health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef]
- Yang, C.-H.; Yen, T.-L.; Hsu, C.-Y.; Thomas, P.-A.; Sheu, J.-R.; Jayakumar, T. Multi-Targeting Andrographolide, a Novel NF-κB Inhibitor, as a Potential Therapeutic Agent for Stroke. Int. J. Mol. Sci. 2017, 18, 1638. [Google Scholar] [CrossRef]
- Pires, B.; Silva, R.; Ferreira, G.; Abdelhay, E. NF-kappaB: Two Sides of the Same Coin. Genes 2018, 9, 24. [Google Scholar] [CrossRef]
- Colombo, F.; Zambrano, S.; Agresti, A. NF-kappaB, the Importance of Being Dynamic: Role and Insights in Cancer. Biomedicines 2018, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, W.; Gu, J.; Du, X.; Lei, L.; Yuan, X.; Sun, G.; Wang, Z.; Li, X.; Liu, G. SREBP-1c overactivates ROS-mediated hepatic NF-kappaB inflammatory pathway in dairy cows with fatty liver. Cell. Signal. 2015, 27, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti-inflammatory roles for NF-κB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Song, F.J.; Zeng, K.W.; Chen, J.F.; Li, Y.; Song, X.M.; Tu, P.F.; Wang, X.M. Extract of Fructus Schisandrae chinensis Inhibits Neuroinflammation Mediator Production from Microglia via NF-kappa B and MAPK Pathways. Chin. J. Integr. Med. 2019, 25, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Suthar, V.S.; Canelas-Raposo, J.; Deniz, A.; Heuwieser, W. Prevalence of subclinical ketosis and relationships with postpartum diseases in European dairy cows. J. Dairy Sci. 2013, 96, 2925–2938. [Google Scholar] [CrossRef]
- Li, C.; Dai, S.; Lu, J.; Zhao, B.; Wang, J.; Li, P.; Wu, Z.; Mu, Y.; Feng, C.; Dong, Q. Methylglyoxal: A newly detected and potentially harmful metabolite in the blood of ketotic dairy cows. J. Dairy Sci. 2018, 101, 8513–8523. [Google Scholar] [CrossRef]
- Ng, Y.W.; Say, Y.H. Palmitic acid induces neurotoxicity and gliatoxicity in SH-SY5Y human neuroblastoma and T98G human glioblastoma cells. PeerJ 2018, 6, e4696. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, Y.; Sun, X.; Zhao, H.; Yao, M.; Hou, L.; Jiang, L. Up-regulation of HO-1 by Nrf2 activation protects against palmitic acid-induced ROS increase in human neuroblastoma BE(2)-M17 cells. Nutr. Res. 2018, 52, 80–86. [Google Scholar] [CrossRef]
- Kapusta, A.; Kuczynska, B.; Puppel, K. Relationship between the degree of antioxidant protection and the level of malondialdehyde in high-performance Polish Holstein-Friesian cows in peak of lactation. PLoS ONE 2018, 13, e0193512. [Google Scholar] [CrossRef]
- Castillo, C.; Hernandez, J.; Valverde, I.; Pereira, V.; Sotillo, J.; Alonso, M.L.; Benedito, J.L. Plasma malonaldehyde (MDA) and total antioxidant status (TAS) during lactation in dairy cows. Res. Vet. Sci. 2006, 80, 133–139. [Google Scholar] [CrossRef]
- Xu, W.; Guo, Y.B.; Li, X.; He, M.R.; Liu, S.D. Palmitic acid induces hepatocellular oxidative stress and activation of inflammasomes. J. South. Med. Univ. 2016, 36, 655. [Google Scholar]
- Shen, X.; Yang, L.; Yan, S.; Wei, W.; Liang, L.; Zheng, H.; Cai, X. The effect of FFAR1 on pioglitazone-mediated attenuation of palmitic acid-induced oxidative stress and apoptosis in betaTC6 cells. Metab. Clin. Exp. 2014, 63, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Miao, Y.; Fan, R.; Song, C.; Wu, G.; Wan, Z.; Zhu, J.; Sun, G.; Zha, W.; Mu, X.; et al. Suppression of Grb2 expression improved hepatic steatosis, oxidative stress, and apoptosis induced by palmitic acid in vitro partly through insulin signaling alteration. In vitro Cell. Dev. Biol.-An. 2013, 49, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Manuel, C.R.; Charron, M.J.; Ashby, C.R.; Reznik, S.E. Saturated and unsaturated fatty acids differentially regulate in vitro and ex vivo placental antioxidant capacity. Am. J. Reprod. Immunol. 2018, 80, e12868. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, X.; Li, D.; Li, Y.; Song, Y.; Deng, Q.; Wang, J.; Zhang, Y.; Ding, H.; Yin, L.; et al. β-Hydroxybutyrate Activates the NF-κB Signaling Pathway to Promote the Expression of Pro-Inflammatory Factors in Calf Hepatocytes. Cell. Physiol. Biochem. 2014, 33, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yang, S.; Xu, C.; Li, L.; Zhang, Y.; Guo, Y.; Zhang, C.; Li, P.; Long, M.; He, J. Proanthocyanidins Protect against β-Hydroxybutyrate-Induced Oxidative Damage in Bovine Endometrial Cells. Molecules 2019, 24, 400. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, X.B.; Fu, S.X.; Wu, C.C.; Wang, X.X.; Yu, G.J.; Long, M.; Wang, Z.; Liu, G.W. Alterations of fatty acid beta-oxidation capability in the liver of ketotic cows. J. Dairy Sci. 2012, 95, 1759–1766. [Google Scholar] [CrossRef]
- Castillo, C.; Hernandez, J.; Bravo, A.; Lopez-Alonso, M.; Pereira, V.; Benedito, J.L. Oxidative status during late pregnancy and early lactation in dairy cows. Vet. J. 2005, 169, 286–292. [Google Scholar] [CrossRef]
- Colado-Velazquez, J., III; Mailloux-Salinas, P.; Medina-Contreras, J.; Cruz-Robles, D.; Bravo, G. Effect of Serenoa Repens on Oxidative Stress, Inflammatory and Growth Factors in Obese Wistar Rats with Benign Prostatic Hyperplasia. Phytother. Res. 2015, 29, 1525–1531. [Google Scholar] [CrossRef]
- Shi, X.; Li, D.; Deng, Q.; Li, Y.; Sun, G.; Yuan, X.; Song, Y.; Wang, Z.; Li, X.; Li, X.; et al. NEFAs activate the oxidative stress-mediated NF-kappaB signaling pathway to induce inflammatory response in calf hepatocytes. J. Steroid Biochem. Mol. Biol. 2015, 145, 103–112. [Google Scholar] [CrossRef]
- Wu, D.; Liu, J.; Pang, X.; Wang, S.; Zhao, J.; Zhang, X.; Feng, L. Palmitic acid exerts pro-inflammatory effects on vascular smooth muscle cells by inducing the expression of C-reactive protein, inducible nitric oxide synthase and tumor necrosis factor-alpha. Int. J. Mol. Med. 2014, 34, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Punetha, M.; Roy, A.K.; Ajithakumar, H.M.; Para, I.A.; Gupta, D.; Singh, M.; Bharati, J. Immunomodulatory effects of probiotics and prilled fat supplementation on immune genes expression and lymphocyte proliferation of transition stage Karan Fries cows. Vet. World 2018, 11, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Berk, M.; Malhi, G.S.; Gray, L.J.; Dean, O.M. The promise of N-acetylcysteine in neuropsychiatry. Trends Pharmacol. Sci. 2013, 34, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, X.; Li, J.J. The role of NF-κB in the regulation of cell stress responses. Int. Immunopharmacol. 2002, 2, 1509–1520. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Fattori, V.; Zarpelon, A.C.; Borghi, S.M.; Staurengo-Ferrari, L.; Carvalho, T.T.; Alves-Filho, J.C.; Cunha, F.Q.; Cunha, T.M.; Casagrande, R.; et al. Pyrrolidine dithiocarbamate inhibits superoxide anion-induced pain and inflammation in the paw skin and spinal cord by targeting NF-kappaB and oxidative stress. Inflammopharmacology 2016, 24, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kim, J.H.; Lee, J.; Ahn, Y.S. Zinc-induced NF-κB inhibition can be modulated by changes in the intracellular metallothionein level. Toxicol. Appl. Pharm. 2003, 190, 189–196. [Google Scholar] [CrossRef]
- Denk, A.; Wirth, T.; Baumann, B. NF-kB transcription factors: critical regulators of hematopoiesis and neuronal survival. Cytokine Growth Factor Rev. 2000, 11, 303–320. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M.; Spurlock, M.E. Palmitate Activates the NF-κB Transcription Factor and Induces IL-6 and TNFα Expression in 3T3-L1 Adipocytes. J. Nutr. 2005, 135, 1841–1846. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′→3′) | Length of Amplified Fragment (bp) |
|---|---|---|
| IL-8 | For: TTAGGCAGACCTCGTTTCCAT Rev: ATGACTTCCAAGCTGGCTGTT | 235 |
| TNF-α | For: GGTCAACATCCTGTCTGCCA Rev: ACTGAGGCGATCTCCCTTCT | 130 |
| NF-κB p65 | For: AGGACCAACCAGACCG Rev: TGTCACCAGGCGAGTTAT | 240 |
| IL-6 | For: AACGAGTGGGTAAAGAACGC Rev: CTGACCAGAGGAGGGAATGC | 144 |
| β-actin | For: GCCCTGAGGCTCTCTTCCA Rev: GCGGATGTCGACGTCACA | 101 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Li, L.; Zhang, C.; Cheng, X.; Zhang, Y.; Guo, Y.; Long, M.; Yang, S.; He, J. Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling. Molecules 2019, 24, 2421. https://doi.org/10.3390/molecules24132421
Li P, Li L, Zhang C, Cheng X, Zhang Y, Guo Y, Long M, Yang S, He J. Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling. Molecules. 2019; 24(13):2421. https://doi.org/10.3390/molecules24132421
Chicago/Turabian StyleLi, Peng, Lanzhi Li, Cai Zhang, Xi Cheng, Yi Zhang, Yang Guo, Miao Long, Shuhua Yang, and Jianbin He. 2019. "Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling" Molecules 24, no. 13: 2421. https://doi.org/10.3390/molecules24132421
APA StyleLi, P., Li, L., Zhang, C., Cheng, X., Zhang, Y., Guo, Y., Long, M., Yang, S., & He, J. (2019). Palmitic Acid and β-Hydroxybutyrate Induce Inflammatory Responses in Bovine Endometrial Cells by Activating Oxidative Stress-Mediated NF-κB Signaling. Molecules, 24(13), 2421. https://doi.org/10.3390/molecules24132421