Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi



Abstract
1. Introduction
2. Anticancer Activity in Similar Signaling Pathways
2.1. Induction of Apoptosis and Promotion of Autophagy
2.2. Cell Cycle Arrest
2.3. Suppression of Metastasis, Invasion, Proliferation and Mesenchymal Stem Cells
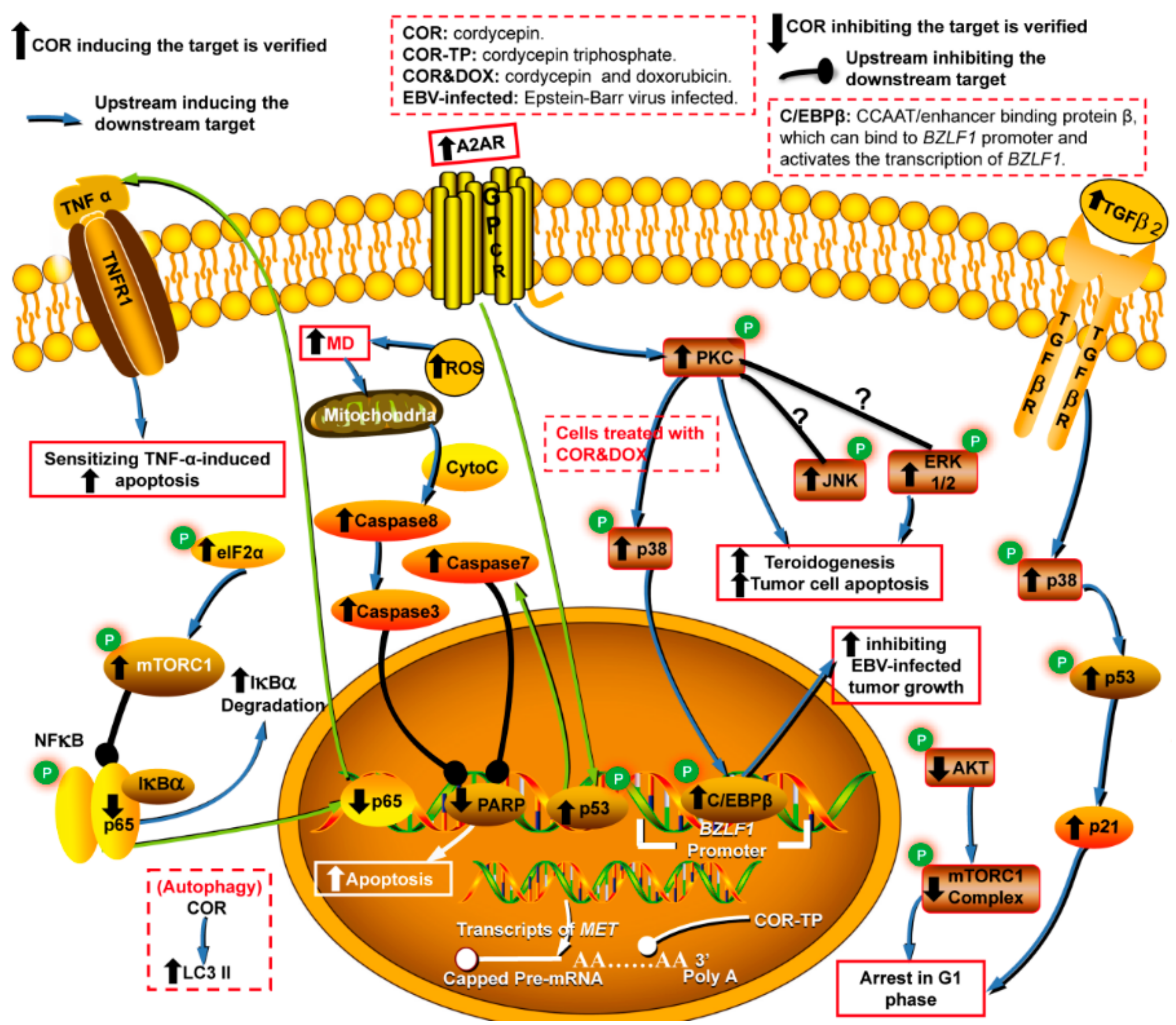
3. Antitumor Activity
3.1. Apoptosis Inhibition, mTOR Repression and Cell Cycle Arrest
3.2. Suppression of Tumor Growth via the GPCR/PKC Signaling Pathway
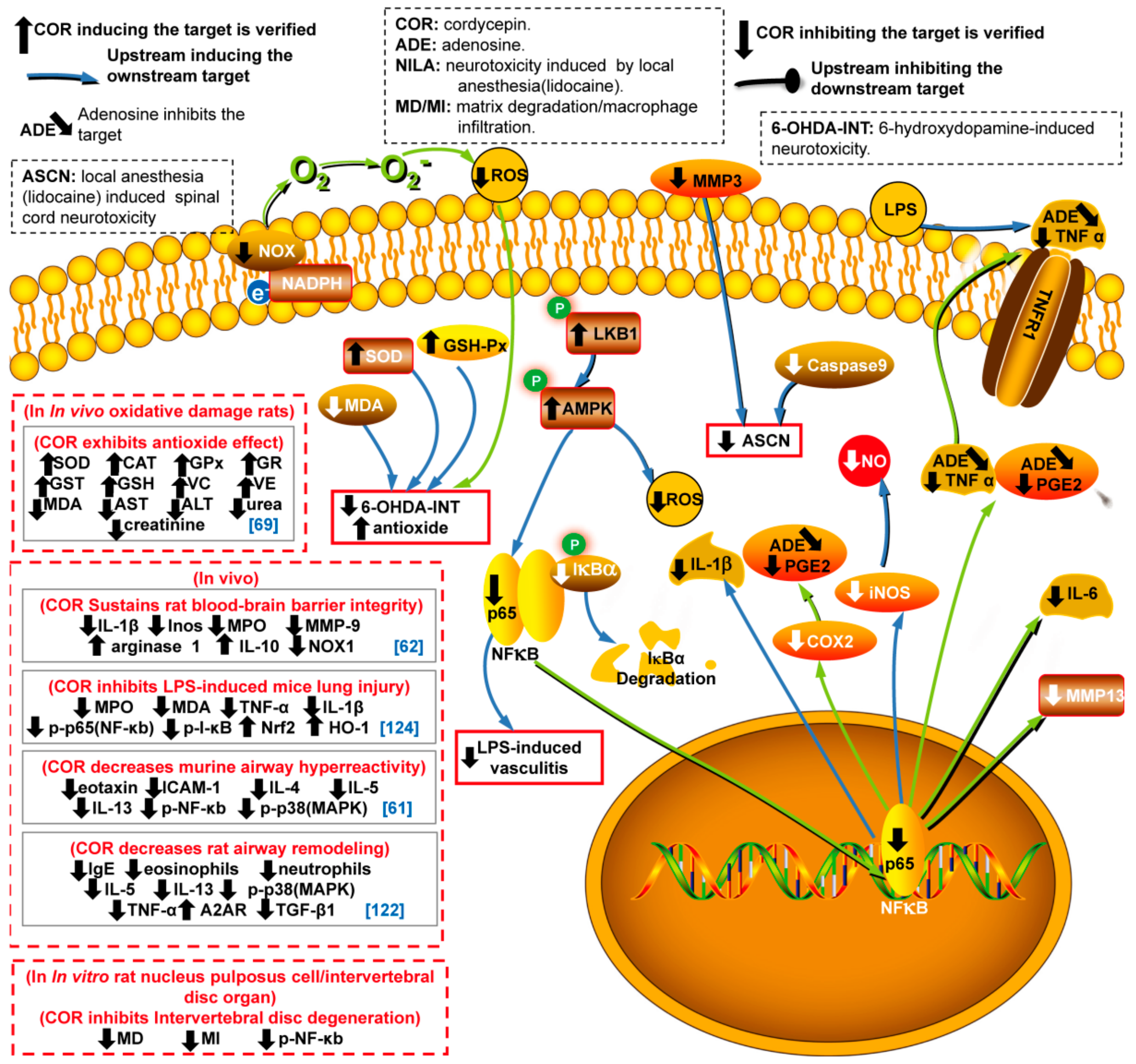
4. Anti-Inflammatory and Anti-Oxidant Activities in Different Cell Lines
4.1. Anti-Inflammatory Activity
4.2. Anti-Oxidant Activity
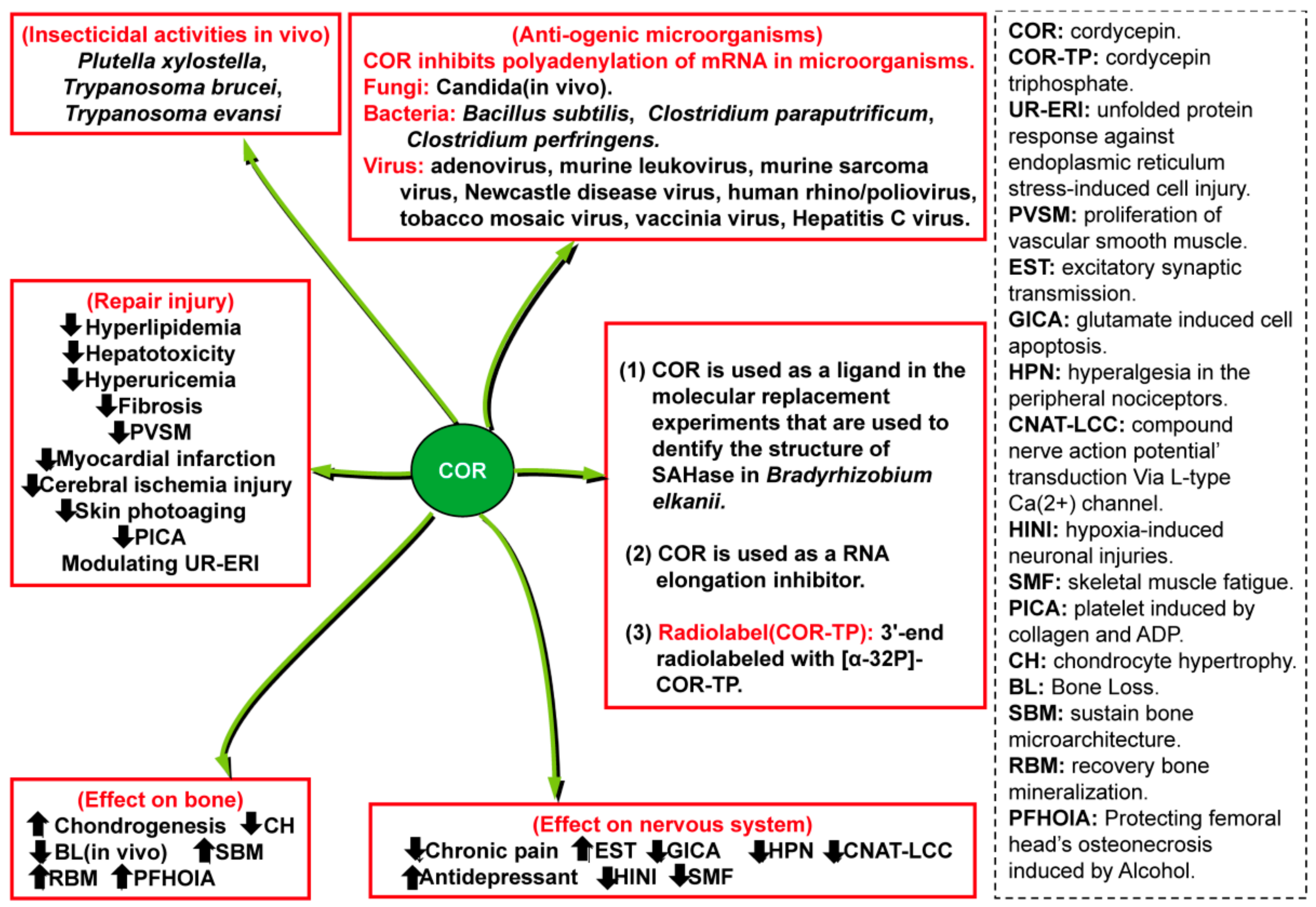
5. Other Medicinal Values and Biological Applications
5.1. Insecticidal Activity and Inhibition of the Growth of Pathogenic Microorganisms
5.2. Inhibition of External Factor-Induced Injury
5.3. Inhibition of Internal Factor-Induced Injury
5.4. Promotion of Chondrogenesis and Inhibition of Bone Loss
5.5. Biological Applications of COR and COR-TP
6. Pros and Cons of COR in Clinical Applications
6.1. Pros of COR in Clinical Applications
6.2. Cons of COR in Clinical Applications
- (a)
- (b)
- (c)
- Low solubility in water: The solubility of COR plays a vital role in drug storage and efficient therapeutic efficiency. The low solubility of COR in water causes low chemical stability, poor oral bioavailability and low cellular uptake. Previous work demonstrated that phosphate-buffered saline (PBS, pH 4.0) is a suitable solvent for COR in intravenous and oral treatments at low doses. Propylene glycol (PPG) is more applicable than PBS at pH 4.0 as a COR solvent for oral treatments [136].
- (d)
- Complex mechanisms of action: COR inhibits and/or induces multiple medicinal targets in a dose-dependent, condition-dependent and nonspecific manner: Different concentrations of COR exhibit diverse effects on MA-10 mouse Leydig tumor cells (MLTCs). A low concentration of COR activates the caspase-3/caspase-6/caspase-7/caspase-8/PARP pathway in MMLTCs, while a high dose of COR markedly increases the levels of p-AKT and p-mTOR and stimulates only caspase-3 rather than caspase-6/caspase-7/caspase-8 [59]. In addition, since COR is less reactive to PARP than to a specific inhibitor of PARP, COR is considered to exhibit PARP-inhibitory activity rather than specifically inhibit PARP [57]. In addition, COR exerts bidirectional regulatory activity under disparate stress-induced conditions [61,62] and in different cell types [59,61].
- (e)
- Drug resistance: Multidrug resistance, one of the major obstacles, markedly decreases the curative potency of anticancer agents and the treatment of other diseases.
- (f)
- Clinical safety and potency: The safety and efficacy of COR as a TdT-positive leukemia treatment is currently being evaluated in phase Ⅱ clinical trials [137]. Currently, the clinical application of COR in the treatment of TdT-positive leukemia is not permitted.
6.3. Medicinal Strategies for Promoting COR Efficiency and Safety
- (a)
- Inhibition of deamination by ADA. Three strategies have been used to approach this problem. (i) combined use of COR and ADA inhibitor; (ii) natural and designed ADA-resistant derivatives of COR; and (iii) nanocarrier for ADA-resistance. In strategy i, the combination of COR and an ADA inhibitor markedly improves the stability of COR. Efficient ADA inhibitors, such as actinomycin D [116], erythro-9-(2-hydroxy-3-nonyl)-adenine [135] and PTN [24], significantly enhance the bioavailability of COR. In strategy ii, multiple natural derivatives of COR in mushrooms [29] and their therapeutic value [138] have been extensively reviewed. In addition, designed ADA-resistant derivatives of COR can overcome treatment failure, such as the high bioavailability pro-cordycepin(N-acyloctanoylcordycepin), which is 4(time of maximum concentration)/30(maximum concentration)/68(area under concentration) times higher than that of COR [139], and ADA-resistant 2-fluoro-3′-deoxyadenosine has similar cytotoxicity (IC50) values of 2.44 ± 0.69 μM (this compound alone) and 2.13 ± 0.87 μM (this compound co-incubated with ADA inhibitor PTN) while COR has markedly different IC50 values of 0.10 ± 0.03 μM(COR co-existing with PTN) and over 100 μM (COR alone) on MOLT4 cells in vitro [140]. Interestingly, an efficient and eco-friendly biotransformation system for generating 5′-O-acetylcordycepin at the 25-g scale and a 96.2% isolated yield in solvent 2-methyltetrahydrofuran was developed, and Novozym 435 (an immobilized Candida antarctica lipase B) used in this system retains 63% of its original activity after 7 recycling batches [141]. In strategy iii, a nanocarrier composed of layered double hydroxides was developed to prevent COR from deamination by ADA, such as [Mg–Al–cordycepin] nanohybrids. At the same concentration, this nanohybrid inhibit the growth of U937 cells at a rate that is 3.185 times higher than that of COR [142].
- (b)
- (c)
- Decreasein toxicity. Several potential strategies are available to decrease the toxicity of COR: (i) low dose of treatment; (ii) natural and designed derivates; (iii) COR carriers for region-targeted treatment and the specific accumulation of COR. In strategy i, an appropriate dose of COR should be selected. In strategy ii, few studies have discovered and designed novel derivatives of COR, such as designed N-octanoylcordycepin which exhibits lower metabolic velocity and higher bioavailability than COR [139]. In strategy iii, appropriate region-targeted drug carriers, such as transferrin-conjugated liposomes for COR delivery to liver cancer cells [143] and gelatin type A nanoparticle for COR delivery to lung cancer cells [134], increase the specific accumulation of COR at the desired region, decrease the total dose of COR and reduce unintended extravasation into healthy regions.
- (d)
- Overcoming drug resistance. Drug resistance presents a serious challenge when diseased cells develop resistance to COR over time through various mechanisms that markedly reduce the curative potency of COR. Previous work uncovered several mechanisms of novel drug resistance, such as extracellular vesicles that mediate drug resistance due to direct exportation [144], cytotoxic drugs sequestration [145], and decreased effective concentration of the drug. COR resistance can be promoted through extracellular vesicle-mediated pathways. In addition, the combination of COR and other agents can efficiently overcome drug resistance. Water or ethanol extract rich in COR of fruiting bodies and/or mycelium of C. militaris (ER-COR) is also a potential strategy to conquer drug resistance.
- (e)
- The low cost and high potency of ER-COR. The proliferation of renal carcinoma cells is more efficiently suppressed by the ER-COR from FB-CM than the ER-COR from mycelia [119]; moreover, ER-COR stimulated apoptosis more effectively than COR alone in vitro in human leukemia cells [53]. These effects may also be due to other components of FB-CM, such as PTN [24] and adenosine. In addition, ER-COR can strengthen immunity by increasing the level of cytokines (IL-2/IFN-γ/TNF-α) in vitro in splenocytes and cytokines (IL-2/IFN-γ/TNF-α/IL-10) in vivo in immunosuppressed mice [146]. Nevertheless, the bioactive components of ER-COR are distinct due to their recognition of different targets. In vitro studies of human hepatocellular carcinoma cells revealed that the inhibitory effects of COR on ERp57 are more efficient than those of zhankuic acid A and adenosine, while the inhibition of PGK-1 mediated by COR is less pronounced than that mediated by zhankuic acid A and adenosine [124].
7. Metabolic Mechanisms of COR in COR-Producing Fungi
7.1. Scientific Name of the Caterpillar Fungus Called DongChongXiaCao in Chinese
7.2. COR-Producing Fungi
7.3. COR Metabolic Mechanisms in C. militaris under Hypoxia, Light and Heat Stress
7.4. Metabolic Mechanisms of COR in C. militaris and O. sinensis
7.5. Comparative Analysis of the COR-Associated Mechanisms Involved in the Fruiting Body of C. militaris and O. sinensis
8. Summary and Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Latest Global Cancer Data in 2018. Available online: http://www.iarc.fr/en/media-centre/pr/2018/pdfs/pr263_E.pdf (accessed on 12 September 2018).
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. Ca Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, S.; Malling, H.; Klenow, H. Isolation of 3′-deoxyadenosine (cordycepin) from the liquid medium of Cordyceps militaris (L. ex Fr.) Link. Biochim. Et Biophys. Acta (Bba)-Nucleic Acids Protein Synth. 1965, 95, 189–193. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.Y.; Moon, A.; Duffin, R.; Barthet-Barateig, A.; Meijer, H.A.; Clemens, M.J.; de Moor, C.H. Cordycepin inhibits protein synthesis and cell adhesion through effects on signal transduction. J. Biol. Chem. 2010, 285, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Holbein, S.; Wengi, A.; Decourty, L.; Freimoser, F.M.; Jacquier, A.; Dichtl, B. Cordycepin interferes with 3’ end formation in yeast independently of its potential to terminate RNA chain elongation. RNA 2009, 15, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Levenson, R.; Kernen, J.; Housman, D. Synchronization of MEL cell commitment with cordycepin. Cell 1979, 18, 1073–1078. [Google Scholar] [CrossRef]
- Holbein, S.; Freimoser, F.M.; Werner, T.P.; Wengi, A.; Dichtl, B. Cordycepin-hypersensitive growth links elevated polyphosphate levels to inhibition of poly(A) polymerase in Saccharomyces cerevisiae. Nucleic Acids Res. 2008, 36, 353–363. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, X.; Liang, Y.N.; Wang, L.; Song, Z.X.; Liu, J.L.; Tang, Z.S. Cordycepin Induces Apoptosis and Inhibits Proliferation of Human Lung Cancer Cell Line H1975 via Inhibiting the Phosphorylation of EGFR. Molecules 2016, 21, 1267. [Google Scholar] [CrossRef]
- Hueng, D.Y.; Hsieh, C.H.; Cheng, Y.C.; Tsai, W.C.; Chen, Y. Cordycepin inhibits migration of human glioblastoma cells by affecting lysosomal degradation and protein phosphatase activation. J. Nutr. Biochem. 2017, 41, 109–116. [Google Scholar] [CrossRef]
- Wu, W.D.; Hu, Z.M.; Shang, M.J.; Zhao, D.J.; Zhang, C.W.; Hong, D.F.; Huang, D.S. Cordycepin down-regulates multiple drug resistant (MDR)/HIF-1α through regulating AMPK/mTORC1 signaling in GBC-SD gallbladder cancer cells. Int. J. Mol. Sci. 2014, 15, 12778–12790. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of Cordyceps with special reference to Cordycepin. 3 Biotech. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cheutin, T.; O’Donohue, M.; Beorchia, A.; Vandelaer, M.; Kaplan, H.; Defever, B.; Ploton, D.; Thiry, M. Three-dimensional organization of active rRNA genes within the nucleolus. J. Cell Sci. 2002, 115, 3297–3307. [Google Scholar] [PubMed]
- Sakaguchi, S.; Fukuda, T.; Takano, H.; Ono, K.; Takio, S. Photosynthetic electron transport differentially regulates the expression of superoxide dismutase genes in liverwort, Marchantia paleacea var. diptera. Plant. Cell Physiol. 2004, 45, 318–324. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Eberhardt, W.; Huwiler, A.; Beck, K.F.; Walpen, S.; Pfeilschifter, J. Amplification of IL-1-Induced Matrix Metalloproteinase-9 Expression by Superoxide in Rat Glomerular Mesangial Cells Is Mediated by Increased Activities of NF-B and Activating Protein-1 and Involves Activation of the Mitogen-Activated Protein Kinase Pathways. J. Immunol. 2000, 165, 5788–5797. [Google Scholar] [PubMed]
- Bommareddy, A.; Hahm, E.R.; Xiao, D.; Powolny, A.A.; Fisher, A.L.; Jiang, Y.; Singh, S.V. Atg5 regulates phenethyl isothiocyanate-induced autophagic and apoptotic cell death in human prostate cancer cells. Cancer Res. 2009, 69, 3704–3712. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Yu, J.; Du, L.; Tang, J.; Feng, W.H. Cordycepin enhances Epstein-Barr virus lytic infection and Epstein-Barr virus-positive tumor treatment efficacy by doxorubicin. Cancer Lett. 2016, 376, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.H.; Huang, J.N.; Li, C.H.; Li, H.H.; Yan, W.W.; Cai, Z.L.; Liu, W.X.; Xiao, P. Cordycepin suppresses excitatory synaptic transmission in rat hippocampal slices via a presynaptic mechanism. Cns Neurosci. 2013, 19, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Oh, J.I.; Lee, S.J.; Park, G. The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells. Neurotoxicology 2014, 41, 102–111. [Google Scholar] [CrossRef]
- Kwon, H.W.; Shin, J.H.; Lim, D.H.; Ok, W.J.; Nam, G.S.; Kim, M.J.; Kwon, H.K.; Noh, J.H.; Lee, J.Y.; Kim, H.H.; et al. Antiplatelet and antithrombotic effects of cordycepin-enriched WIB-801CE from Cordyceps militaris ex vivo, in vivo, and in vitro. Bmc Complement. Altern. Med. 2016, 16, 508. [Google Scholar] [CrossRef]
- Oztas, O.; Selby, C.P.; Sancar, A.; Adebali, O. Genome-wide excision repair in Arabidopsis is coupled to transcription and reflects circadian gene expression patterns. Nat. Commun. 2018, 9, 1503. [Google Scholar] [CrossRef]
- Masuda, M.; Das, S.K.; Hatashita, M.; Fujihara, S.; Sakurai, A. Efficient production of cordycepin by the Cordyceps militaris mutant G81-3 for practical use. Process. Biochem. 2014, 49, 181–187. [Google Scholar] [CrossRef]
- Lin, Q.; Long, L.; Wu, L.; Zhang, F.; Wu, S.; Zhang, W.; Sun, X. Evaluation of different agricultural wastes for the production of fruiting bodies and bioactive compounds by medicinal mushroom Cordyceps militaris. J. Sci. Food Agric. 2017, 97, 3476–3480. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Kodama, K.; Kusakabe, H.; Machida, H.; Midorikawa, Y.; Shibuya, S.; Kuninaka, A.; Yoshino, H. Isolation of 2′-Deoxycoformycin and Cordycepin from Wheat Bran Culture of Aspergillus nidulans Y176–2. Agric. Biol. Chem. 1979, 43, 2375–2377. [Google Scholar]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef]
- Tian, X.; Li, Y.; Shen, Y.; Li, Q.; Wang, Q.; Feng, L. Apoptosis and inhibition of proliferation of cancer cells induced by cordycepin. Oncol. Lett. 2015, 10, 595–599. [Google Scholar] [CrossRef]
- Paterson, R.R. Cordyceps: A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef]
- Phan, C.W.; Wang, J.K.; Cheah, S.C.; Naidu, M.; David, P.; Sabaratnam, V. A review on the nucleic acid constituents in mushrooms: Nucleobases, nucleosides and nucleotides. Crit. Rev. Biotechnol. 2018, 38, 762–777. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, Y.H.; Pan, B.S.; Chang, M.M.; Huang, B.M. Functional study of Cordyceps sinensis and cordycepin in male reproduction: A review. J. Food Drug Anal. 2017, 25, 197–205. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.C.; Lin, Y.T.; Huang, S.H.; Wang, S.M. Cordycepin induces apoptosis of CGTH W-2 thyroid carcinoma cells through the calcium-calpain-caspase 7-PARP pathway. J. Agric. Food Chem. 2010, 58, 11645–11652. [Google Scholar] [CrossRef]
- Choi, S.; Lim, M.H.; Kim, K.M.; Jeon, B.H.; Song, W.O.; Kim, T.W. Cordycepin-induced apoptosis and autophagy in breast cancer cells are independent of the estrogen receptor. Toxicol. Appl. Pharm. 2011, 257, 165–173. [Google Scholar] [CrossRef]
- Jang, K.J.; Kwon, G.S.; Jeong, J.W.; Kim, C.H.; Yoon, H.M.; Kim, G.Y.; Shim, J.H.; Moon, S.K.; Kim, W.J.; Choi, Y.H. Cordycepin induces apoptosis through repressing hTERT expression and inducing extranuclear export of hTERT. J. Biosci. Bioeng. 2015, 119, 351–357. [Google Scholar] [CrossRef]
- Lee, S.Y.; Debnath, T.; Kim, S.K.; Lim, B.O. Anti-cancer effect and apoptosis induction of cordycepin through DR3 pathway in the human colonic cancer cell HT-29. Food Chem. Toxicol. 2013, 60, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Ling, J.; Zhang, G.; Liu, F.; Tao, S.; Han, Z.; Chen, S.; Chen, Z.; Le, H. Cordycepin induces cell cycle arrest and apoptosis by inducing DNA damage and up-regulation of p53 in Leukemia cells. Cell Cycle 2015, 14, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Chaicharoenaudomrung, N.; Jaroonwitchawan, T.; Noisa, P. Cordycepin induces apoptotic cell death of human brain cancer through the modulation of autophagy. Toxicol. Vitr. 2018, 46, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Lian, S.; Li, D.; Lin, X.; Chen, B.; Wei, H.; Yang, T. Anti-hepatocarcinoma effect of cordycepin against NDEA-induced hepatocellular carcinomas via the PI3K/Akt/mTOR and Nrf2/HO-1/NF-κB pathway in mice. Biomed. Pharm. 2017, 95, 1868–1875. [Google Scholar] [CrossRef]
- Jeong, J.W.; Jin, C.Y.; Park, C.; Hong, S.H.; Kim, G.Y.; Jeong, Y.K.; Lee, J.D.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by cordycepin via reactive oxygen species generation in human leukemia cells. Toxicol. Vitr. 2011, 25, 817–824. [Google Scholar] [CrossRef]
- Zhang, J.L.; Xu, Y.; Shen, J. Cordycepin inhibits lipopolysaccharide(LPS)-induced tumor necrosis factor (TNF)-α production via activating amp-activated protein kinase (AMPK) signaling. Int. J. Mol. Sci. 2014, 15, 12119–12134. [Google Scholar] [CrossRef]
- Hwang, J.H.; Park, S.J.; Ko, W.G.; Kang, S.M.; Lee, D.B.; Bang, J.; Park, B.J.; Wee, C.B.; Kim, D.J.; Jang, I.S.; et al. Cordycepin induces human lung cancer cell apoptosis by inhibiting nitric oxide mediated ERK/Slug signaling pathway. Am. J. Cancer Res. 2017, 7, 417–432. [Google Scholar]
- Ko, B.S.; Lu, Y.J.; Yao, W.L.; Liu, T.A.; Tzean, S.S.; Shen, T.L.; Liou, J.Y. Cordycepin Regulates GSK-3b/b-Catenin Signaling in Human Leukemia Cells. PLoS ONE 2013, 8, e76320. [Google Scholar] [CrossRef]
- Liu, F.C.; Lai, M.T.; Chen, Y.Y.; Lin, W.H.; Chang, S.J.; Sheu, M.J.; Wu, C.H. Elucidating the inhibitory mechanisms of the ethanolic extract of the fruiting body of the mushroom Antrodia cinnamomea on the proliferation and migration of murine leukemia WEHI-3 cells and their tumorigenicity in a BALB/c allograft tumor model. Phytomedicine 2013, 20, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Seong da, B.; Hong, S.; Muthusami, S.; Kim, W.D.; Yu, J.R.; Park, W.Y. Cordycepin increases radiosensitivity in cervical cancer cells by overriding or prolonging radiation-induced G2/M arrest. Eur. J. Pharm. 2016, 771, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Yoshioka, Y.; Yamamoto, Y.; Ochiya, T. How cancer cells dictate their microenvironment: Present roles of extracellular vesicles. Cell. Mol. Life Sci. 2017, 74, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Maresh, G.; Zhang, X.; Salomon, C.; Hooper, J.; Margolin, D.; Li, L. The Emerging Roles of Extracellular Vesicles As Communication Vehicles within the Tumor Microenvironment and Beyond. Front. Endocrinol. 2017, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.M.; Lu, Y.J.; Ko, B.S.; Jan, Y.J.; Shyue, S.K.; Yet, S.F.; Liou, J.Y. Cordycepin disrupts leukemia association with mesenchymal stromal cells and eliminates leukemia stem cell activity. Sci. Rep. 2017, 7, 43930. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Bottazzi, B.; Colotta, F.; Sozzani, S.; Ruco, L. The origin and function of tumor-associated macrophages. Immunol. Today 1992, 13, 265–270. [Google Scholar] [CrossRef]
- Xing, F. ancer associated fibroblasts (CAFs) in tumor microenvironment. Front. Biosci. 2010, 15, 166. [Google Scholar]
- Jeong, J.W.; Jin, C.Y.; Park, C.; Han, M.H.; Kim, G.Y.; Moon, S.K.; Kim, C.G.; Jeong, Y.K.; Kim, W.J.; Lee, J.D.; et al. Inhibition of migration and invasion of LNCaP human prostate carcinoma cells by cordycepin through inactivation of Akt. Int. J. Oncol. 2012, 40, 1697–1704. [Google Scholar]
- Chen, Y.Y.; Chou, P.Y.; Chien, Y.C.; Wu, C.H.; Wu, T.S.; Sheu, M.J. Ethanol extracts of fruiting bodies of Antrodia cinnamomea exhibit anti-migration action in human adenocarcinoma CL1-0 cells through the MAPK and PI3K/AKT signaling pathways. Phytomedicine 2012, 19, 768–878. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.M.; Youn, H.J.; Jung, S.H.; Han, J.H.; Jeong, Y.J.; Chung, E.Y.; Jung, J.Y.; Kim, B.S.; Lee, S.H.; Lee, Y.R.; et al. Cordycepin inhibits TPA-induced matrix metalloproteinase-9 expression by suppressing the MAPK/AP-1 pathway in MCF-7 human breast cancer cells. Int. J. Mol. Med. 2010, 25, 255–260. [Google Scholar]
- Su, N.W.; Wu, S.H.; Chi, C.W.; Liu, C.J.; Tsai, T.H.; Chen, Y.J. Metronomic Cordycepin Therapy Prolongs Survival of Oral Cancer-Bearing Mice and Inhibits Epithelial-Mesenchymal Transition. Molecules 2017, 22, 629. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Shichiri, H.; Uda, A.; Yamashita, K.; Nishioka, T.; Kume, M.; Makimoto, H.; Nakagawa, T.; Hirano, T.; Hirai, M. Apoptotic Effects of the Extracts of Cordyceps militaris via Erk Phosphorylation in a Renal Cell Carcinoma Cell Line. Phytother. Res. 2015, 29, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Aramwit, P.; Bang, N.; Ratanavaraporn, J.; Nakpheng, T.; Srichana, T. An Anti-Cancer Cordycepin Produced by Cordyceps militaris Growing on the Dead Larva of Bombyx mori Silkworm. J. Agric. Sci. 2014, 6, 41–53. [Google Scholar] [CrossRef]
- Glosse, P.; Foller, M. AMP-Activated Protein Kinase (AMPK)-Dependent Regulation of Renal Transport. Int. J. Mol. Sci. 2018, 19, 3481. [Google Scholar] [CrossRef] [PubMed]
- Niida, A.; Hiroko, T.; Kasai, M.; Furukawa, Y.; Nakamura, Y.; Suzuki, Y.; Sugano, S.; Akiyama, T. DKK1, a negative regulator of Wnt signaling, is a target of the beta-catenin/TCF pathway. Oncogene 2004, 23, 8520–8526. [Google Scholar] [CrossRef]
- Lee, H.J.; Burger, P.; Vogel, M.; Friese, K.; Bruning, A. The nucleoside antagonist cordycepin causes DNA double strand breaks in breast cancer cells. Investig. New Drugs 2012, 30, 1917–1925. [Google Scholar] [CrossRef]
- Chen, L.S.; Stellrecht, C.M.; Gandhi, V. RNA-directed agent, cordycepin, induces cell death in multiple myeloma cells. Br. J. Haematol. 2008, 140, 682–691. [Google Scholar] [CrossRef]
- Pan, B.S.; Wang, Y.K.; Lai, M.S.; Mu, Y.F.; Huang, B.M. Cordycepin induced MA-10 mouse Leydig tumor cell apoptosis by regulating p38 MAPKs and PI3K/AKT signaling pathways. Sci Rep. 2015, 5, 13372. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, S.H.; Hueng, D.Y.; Syu, J.P.; Liao, C.C.; Wu, Y.C. Cordycepin induces apoptosis of C6 glioma cells through the adenosine 2A receptor-p53-caspase-7-PARP pathway. Chem. Biol. Interact. 2014, 216, 17–25. [Google Scholar] [CrossRef]
- Kadomatsu, M.; Nakajima, S.; Kato, H.; Gu, L.; Chi, Y.; Yao, J.; Kitamura, M. Cordycepin as a sensitizer to tumour necrosis factor (TNF)-α-induced apoptosis through eukaryotic translation initiation factor 2α (eIF2α)- and mammalian target of rapamycin complex 1 (mTORC1)-mediated inhibition of nuclear factor (NF)-κB. Clin. Exp. Immunol. 2012, 168, 325–332. [Google Scholar] [CrossRef]
- Kitamura, M.; Kato, H.; Saito, Y.; Nakajima, S.; Takahashi, S.; Johno, H.; Gu, L.; Katoh, R. Aberrant, differential and bidirectional regulation of the unfolded protein response towards cell survival by 3′-deoxyadenosine. Cell Death Differ. 2011, 18, 1876–1888. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G protein regulation of MAPK networks. Oncogene 2007, 26, 3122–3142. [Google Scholar] [CrossRef] [PubMed]
- Pao, H.Y.; Pan, B.S.; Leu, S.F.; Huang, B.M. Cordycepin stimulated steroidogenesis in MA-10 mouse Leydig tumor cells through the protein kinase C Pathway. J. Agric. Food Chem. 2012, 60, 4905–4913. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.Y.; Chen, C.C.; Lee, L.Y.; Lin, T.W.; Kuo, C.F. N(6)-(2-Hydroxyethyl)adenosine in the Medicinal Mushroom Cordyceps cicadae Attenuates Lipopolysaccharide-Stimulated Pro-inflammatory Responses by Suppressing TLR4-Mediated NF-κB Signaling Pathways. J. Nat. Prod. 2015, 78, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Peng, L.; Chen, H.; Shen, Y.; Yu, K.; Cheng, S. Cordycepin prevented IL-β-induced expression of inflammatory mediators in human osteoarthritis chondrocytes. Int. Orthop. 2014, 38, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.; He, Y.; Li, T.; Wang, W.; Zhang, J.; Wei, J.; Deng, Y.; Lin, R. Cordycepin alleviates airway hyperreactivity in a murine model of asthma by attenuating the inflammatory process. Int. Immunopharmacol. 2015, 26, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Wang, A.; He, Y.; Si, Z.; Xu, S.; Zhang, S.; Wang, K.; Wang, D.; Liu, Y. Cordycepin attenuates traumatic brain injury-induced impairments of blood-brain barrier integrity in rats. Brain Res. Bull. 2016, 127, 171–176. [Google Scholar] [CrossRef]
- Yoou, M.S.; Yoon, K.W.; Choi, Y.; Kim, H.M.; Jeong, H.J. Cordycepin diminishes thymic stromal lymphopoietin-induced interleukin-13 production. Eur J. Pharm. 2017, 802, 1–6. [Google Scholar] [CrossRef]
- Kan, H.; Wang, Y.; Wang, D.; Sun, H.; Zhou, S.; Wang, H.; Guan, J.; Li, M. Cordycepin rescues lidocaine-induced neurotoxicity in dorsal root ganglion by interacting with inflammatory signaling pathway MMP3. Eur J. Pharm. 2018, 827, 88–93. [Google Scholar] [CrossRef]
- Yu, H.M.; Wang, B.S.; Huang, S.C.; Duh, P.D. Comparison of protective effects between cultured Cordyceps militaris and natural Cordyceps sinensis against oxidative damage. J. Agric. Food Chem. 2006, 54, 3132–3138. [Google Scholar] [CrossRef]
- Jeong, M.H.; Park, Y.S.; Jeong, D.H.; Lee, C.G.; Kim, J.S.; Oh, S.J.; Jeong, S.K.; Yang, K.; Jo, W.S. In vitro evaluation of Cordyceps militaris as a potential radioprotective agent. Int. J. Mol. Med. 2014, 34, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Ge, Y.; Sun, L.; Xu, X.; Xie, P.; Zhan, M.; Wang, M.; Dong, Z.; Li, J.; Duan, S.; et al. Cordycepin inhibits albumin-induced epithelial-mesenchymal transition of renal tubular epithelial cells by reducing reactive oxygen species production. Free Radic Res. 2012, 46, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, O.J.; Feng, Y.; Olatunji, O.O.; Tang, J.; Ouyang, Z.; Su, Z. Cordycepin protects PC12 cells against 6-hydroxydopamine induced neurotoxicity via its antioxidant properties. Biomed. Pharm. 2016, 81, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, T.; Yoo, S.K.; Kim, S.W.; Hwang, S.Y.; Sohn, S.H.; Kim, I.W.; Kim, S.K. Cordycepin (3′-deoxyadenosine) attenuates age-related oxidative stress and ameliorates antioxidant capacity in rats. Exp. Gerontol. 2012, 47, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Rottman, F.; Guarino, A.J. The inhibition of purine biosynthesis De novo in Bacillus subtilis by cordycepin. Biochim. Et Biophys. Acta 1964, 80, 640–647. [Google Scholar] [CrossRef]
- Philipson, L.; Wall, R.; Glickman, G.; Darnell, J.E. Addition of polyadenylate sequences to virus-specific RNA during adenovirus replication. Proc. Natl. Acad. Sci. USA 1971, 68, 2806–2809. [Google Scholar] [CrossRef]
- Aspegren, A.; Bridge, E. Release of snRNP and RNA from transcription sites in adenovirus-infected cells. Exp. Cell Res. 2002, 276, 273–283. [Google Scholar] [CrossRef]
- Wu, A.M.; Ting, R.C.; Paran, M.; Gallo, R.C. Cordycepin inhibits induction of murine leukovirus production by 5-iodo-2’-deoxyuridine. Proc. Natl. Acad. Sci. USA 1972, 69, 3820–3824. [Google Scholar] [CrossRef]
- Richardson, L.S.; Ting, R.C.; Gallo, R.C.; Wu, A.M. Effect of cordycepin on the replication of type-c RNA tumor viruses. Int. J. Cancer 1975, 15, 451–456. [Google Scholar] [CrossRef]
- Weiss, S.R.; Bratt, M.A. Effect of cordycepin (3′-deoxyadenosine) on virus-specific RNA species synthesized in Newcastle disease virus-infected cells. J. Virol. 1975, 16, 1575–1583. [Google Scholar]
- Nair, C.N.; Panicali, D.L. Polyadenylate sequences of human rhinovirus and poliovirus RNA and cordycepin sensitivity of virus replication. J. Virol. 1976, 20, 170–176. [Google Scholar] [PubMed]
- Leinwand, L.; Ruddle, F.H. Stimulation of in vitro translation of messenger RNA by actinomycin D and cordycepin. Science 1977, 197, 381–383. [Google Scholar] [CrossRef] [PubMed]
- Person, A.; Ben-Hamida, F.; Beaud, G. Inhibition of 40S-Met-tRNAfMet ribosomal initiation complex formation by vaccinia virus. Nature 1980, 287, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Hamatake, R.K.; Mathis, D.M.; Racela, J.; Rigat, K.L.; Lemm, J.; Colonno, R.J. De novo initiation of RNA synthesis by the RNA-dependent RNA polymerase (NS5B) of hepatitis C virus. J. Virol. 2000, 74, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Park, S.J.; Lee, S.G.; Shin, S.C.; Choi, D.H. Cordycepin: Selective growth inhibitor derived from liquid culture of Cordyceps militaris against Clostridium spp. J. Agric. Food Chem. 2000, 48, 2744–2748. [Google Scholar] [CrossRef] [PubMed]
- Sugar, A.M.; McCaffrey, R.P. Antifungal activity of 3′-deoxyadenosine (cordycepin). Antimicrob. Agents Chemother. 1998, 42, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Tomaka, F.L.; Wegmann, F.; Stieh, D.J.; Alter, G.; Robb, M.L.; Michael, N.L.; Peter, L.; Nkolola, J.P.; Borducchi, E.N.; et al. Evaluation of a mosaic HIV-1 vaccine in a multicentre, randomised, double-blind, placebo-controlled, phase 1/2a clinical trial (APPROACH) and in rhesus monkeys (NHP 13-19). Lancet 2018, 392, 232–243. [Google Scholar] [CrossRef]
- Kim, J.R.; Yeon, S.H.; Kim, H.S.; Ahn, Y.J. Larvicidal activity against Plutella xylostella of cordycepin from the fruiting body of Cordyceps militaris. Pest. Manag. Sci. 2002, 58, 713–717. [Google Scholar] [CrossRef]
- Vodnala, S.K.; Ferella, M.; Lunden-Miguel, H.; Betha, E.; van Reet, N.; Amin, D.N.; Oberg, B.; Andersson, B.; Kristensson, K.; Wigzell, H.; et al. Preclinical assessment of the treatment of second-stage African trypanosomiasis with cordycepin and deoxycoformycin. PLoS Negl. Trop. Dis. 2009, 3, e495. [Google Scholar] [CrossRef]
- Dalla Rosa, L.; da Silva, A.S.; Gressler, L.T.; Oliveira, C.B.; Dambros, M.G.; Miletti, L.C.; Franca, R.T.; Lopes, S.T.; Samara, Y.N.; da Veiga, M.L.; et al. Cordycepin(3′-deoxyadenosine) pentostatin (deoxycoformycin) combination treatment of mice experimentally infected with Trypanosoma evansi. Parasitology 2013, 140, 663–671. [Google Scholar] [CrossRef]
- Dalla Rosa, L.; Da Silva, A.S.; Oliveira, C.B.; Gressler, L.T.; Arnold, C.B.; Baldissera, M.D.; Sagrillo, M.; Sangoi, M.; Moresco, R.; Mendes, R.E.; et al. Dose finding of 3’deoxyadenosine and deoxycoformycin for the treatment of Trypanosoma evansi infection: An effective and nontoxic dose. Microb. Pathog. 2015, 85, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Yeo, S.G. Aging. Korean J. Audiol. 2013, 17, 39–44. [Google Scholar] [CrossRef] [PubMed][Green Version]
- He, Y.T.; Zhang, X.L.; Xie, Y.M.; Xu, Y.X.; Li, J.R. Extraction and Antioxidant Property In Vitro of Cordycepin in Artificially Cultivated Cordyceps Militaris. Adv. Mater. Res. 2013, 750–752, 1593–1596. [Google Scholar] [CrossRef]
- Lee, Y.R.; Noh, E.M.; Jeong, E.Y.; Yun, S.K.; Jeong, Y.J.; Kim, J.H.; Kwon, K.B.; Kim, B.S.; Lee, S.H.; Park, C.S.; et al. Cordycepin inhibits UVB-induced matrix metalloproteinase expression by suppressing the NF-kappaB pathway in human dermal fibroblasts. Exp. Mol. Med. 2009, 41, 548–854. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cheung, F.W.; Chan, M.H.; Hui, P.K.; Ip, S.P.; Ling, Y.H.; Che, C.T.; Liu, W.K. Protective roles of Cordyceps on lung fibrosis in cellular and rat models. J. Ethnopharmacol. 2012, 143, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Johno, H.; Nakajima, S.; Kato, H.; Takahashi, S.; Katoh, R.; Kitamura, M. Blockade of Smad signaling by 3’-deoxyadenosine: A mechanism for its anti-fibrotic potential. Lab. Investig. 2013, 93, 450–461. [Google Scholar] [CrossRef]
- Park, E.S.; Kang, D.H.; Yang, M.K.; Kang, J.C.; Jang, Y.C.; Park, J.S.; Kim, S.K.; Shin, H.S. Cordycepin, 3′-Deoxyadenosine, Prevents Rat Hearts from Ischemia/Reperfusion Injury Via Activation of Akt/GSK-3 beta/p70S6K Signaling Pathway and HO-1 Expression. Cardiovasc. Toxicol. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Cheng, Z.; He, W.; Zhou, X.; Lv, Q.; Xu, X.; Yang, S.; Zhao, C.; Guo, L. Cordycepin protects against cerebral ischemia/reperfusion injury in vivo and in vitro. Eur J. Pharm. 2011, 664, 20–28. [Google Scholar] [CrossRef]
- Jung, S.M.; Park, S.S.; Kim, W.J.; Moon, S.K. Ras/ERK1 pathway regulation of p27KIP1-mediated G1-phase cell-cycle arrest in cordycepin-induced inhibition of the proliferation of vascular smooth muscle cells. Eur J. Pharm. 2012, 681, 15–22. [Google Scholar] [CrossRef]
- Yao, L.H.; Yu, H.M.; Xiong, Q.P.; Sun, W.; Xu, Y.L.; Meng, W.; Li, Y.P.; Liu, X.P.; Yuan, C.H. Cordycepin Decreases Compound Action Potential Conduction of Frog Sciatic Nerve In Vitro Involving Ca(2+) -Dependent Mechanisms. Neural Plast. 2015, 2015, 927817. [Google Scholar] [CrossRef]
- Ferrari, L.F.; Araldi, D.; Levine, J.D. Distinct terminal and cell body mechanisms in the nociceptor mediate hyperalgesic priming. J. Neurosci 2015, 35, 6107–6116. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, L.F.; Bogen, O.; Chu, C.; Levine, J.D. Peripheral administration of translation inhibitors reverses increased hyperalgesia in a model of chronic pain in the rat. J. Pain 2013, 14, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.H.; Meng, W.; Song, R.F.; Xiong, Q.P.; Sun, W.; Luo, Z.Q.; Yan, W.W.; Li, Y.P.; Li, X.P.; Li, H.H.; et al. Modulation effects of cordycepin on the skeletal muscle contraction of toad gastrocnemius muscle. Eur J. Pharm. 2014, 726, 9–15. [Google Scholar] [CrossRef]
- Takahashi, S.; Tamai, M.; Nakajima, S.; Kato, H.; Johno, H.; Nakamura, T.; Kitamura, M. Blockade of adipocyte differentiation by cordycepin. Br. J. Pharm. 2012, 167, 561–575. [Google Scholar] [CrossRef]
- Cha, J.Y.; Ahn, H.Y.; Cho, Y.S.; Je, J.Y. Protective effect of cordycepin-enriched Cordyceps militaris on alcoholic hepatotoxicity in Sprague-Dawley rats. Food Chem. Toxicol. 2013, 60, 52–57. [Google Scholar] [CrossRef]
- Guo, P.; Kai, Q.; Gao, J.; Lian, Z.Q.; Wu, C.M.; Wu, C.A.; Zhu, H.B. Cordycepin prevents hyperlipidemia in hamsters fed a high-fat diet via activation of AMP-activated protein kinase. J. Pharm. Sci. 2010, 113, 395–403. [Google Scholar] [CrossRef]
- Wu, C.; Guo, Y.; Su, Y.; Zhang, X.; Luan, H.; Zhang, X.; Zhu, H.; He, H.; Wang, X.; Sun, G.; et al. Cordycepin activates AMP-activated protein kinase (AMPK) via interaction with the gamma1 subunit. J. Cell Mol. Med. 2014, 18, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hou, Y.; Zhu, M.; Bao, H.; Nie, J.; Zhang, G.Y.; Shan, L.; Yao, Y.; Du, K.; Yang, H.; et al. 3′-Deoxyadenosine (Cordycepin) Produces a Rapid and Robust Antidepressant Effect via Enhancing Prefrontal AMPA Receptor Signaling Pathway. Int. J. Neuropsychopharmacol. 2016, 19, pyv112. [Google Scholar] [CrossRef]
- Dou, C.; Cao, Z.; Ding, N.; Hou, T.; Luo, F.; Kang, F.; Yang, X.; Jiang, H.; Xie, Z.; Hu, M.; et al. Cordycepin Prevents Bone Loss through Inhibiting Osteoclastogenesis by Scavenging ROS Generation. Nutrients 2016, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Zhu, D.Y.; Xu, Z.L.; Yin, J.H.; Yu, X.W.; Mei, J.; Gao, Y.S.; Zhang, C.Q. The Protective Effect of Cordycepin On Alcohol-Induced Osteonecrosis of the Femoral Head. Cell Physiol. Biochem. 2017, 42, 2391–2403. [Google Scholar] [CrossRef]
- Cao, Z.; Dou, C.; Li, J.; Tang, X.; Xiang, J.; Zhao, C.; Zhu, L.; Bai, Y.; Xiang, Q.; Dong, S. Cordycepin inhibits chondrocyte hypertrophy of mesenchymal stem cells through PI3K/Bapx1 and Notch signaling pathway. Bmb Rep. 2016, 49, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Manszewski, T.; Szpotkowski, K.; Jaskolski, M. Crystallographic and SAXS studies of S-adenosyl-l-homocysteine hydrolase from Bradyrhizobium elkanii. IUCrJ 2017, 4, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Nausch, L.W.; Martin, G.; Keller, W.; Doublie, S. Crystal structure of human poly(A) polymerase gamma reveals a conserved catalytic core for canonical poly(A) polymerases. J. Mol. Biol. 2014, 426, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.C.; Wong, J.W.; Suen, Y.K.; Kwok, T.T.; Fung, K.P.; Kong, S.K. Cordycepin induced eryptosis in mouse erythrocytes through a Ca2+-dependent pathway without caspase-3 activation. Arch. Toxicol. 2007, 81, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Beauchemin, M.; Dagenais-Bellefeuille, S.; Letourneau, L.; Cappadocia, M.; Morse, D. The Lingulodinium circadian system lacks rhythmic changes in transcript abundance. BMC Biol. 2014, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.C.; Nakaminami, K.; Matsui, A.; Kobayashi, S.; Kurihara, Y.; Toyooka, K.; Tanaka, M.; Seki, M. Oligouridylate Binding Protein 1b Plays an Integral Role in Plant Heat Stress Tolerance. Front. Plant. Sci. 2016, 7, 853. [Google Scholar] [CrossRef] [PubMed]
- Koc, Y.; Urbano, A.G.; Sweeney, E.B.; McCaffrey, R. Induction of apoptosis by cordycepin in ADA-inhibited TdT-positive leukemia cells. Leukemia 1996, 10, 1019–1024. [Google Scholar] [PubMed]
- Chou, S.M.; Lai, W.J.; Hong, T.W.; Lai, J.Y.; Tsai, S.H.; Chen, Y.H.; Yu, S.H.; Kao, C.H.; Chu, R.; Ding, S.T.; et al. Synergistic property of cordycepin in cultivated Cordyceps militaris-mediated apoptosis in human leukemia cells. Phytomedicine 2014, 21, 1516–1524. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Liu, J.; Xu, P.; Zhang, X.M.; Tian, Y.Y.; Xue, Y.M.; Gao, X.Y.; Liu, Y.; Wang, J.H. Synergistic effect of HMGB1 knockdown and cordycepin in the K562 human chronic myeloid leukemia cell line. Mol. Med. Rep. 2015, 12, 4462–4468. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Naura, A.S.; Errami, Y.; Ju, J.; Boulares, A.H. Cordycepin blocks lung injury-associated inflammation and promotes BRCA1-deficient breast cancer cell killing by effectively inhibiting PARP. Mol. Med. 2011, 17, 893–900. [Google Scholar] [CrossRef]
- Lee, H.H.; Park, C.; Jeong, J.W.; Kim, M.J.; Seo, M.J.; Kang, B.W.; Park, J.U.; Kim, G.Y.; Choi, B.T.; Choi, Y.H.; et al. Apoptosis induction of human prostate carcinoma cells by cordycepin through reactive oxygen species mediated mitochondrial death pathway. Int. J. Oncol. 2013, 42, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Jeong, J.W.; Lee, J.H.; Kim, G.Y.; Cheong, J.; Jeong, Y.K.; Yoo, Y.H.; Choi, Y.H. Cordycepin increases sensitivity of Hep3B human hepatocellular carcinoma cells to TRAIL-mediated apoptosis by inactivating the JNK signaling pathway. Oncol. Rep. 2013, 30, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Liu, F.C.; Wu, T.S.; Sheu, M.J. Antrodia cinnamomea Inhibits Migration in Human Hepatocellular Carcinoma Cells: The Role of ERp57 and PGK-1. Am. J. Chin. Med. 2015, 43, 1671–1696. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.A.; Xiang, S.S.; Li, H.F.; Wu, X.S.; Li, M.L.; Shu, Y.J.; Zhang, F.; Cao, Y.; Ye, Y.Y.; Bao, R.F.; et al. Cordycepin induces S phase arrest and apoptosis in human gallbladder cancer cells. Molecules 2014, 19, 11350–11365. [Google Scholar] [CrossRef] [PubMed]
- Qing, R.; Huang, Z.; Tang, Y.; Xiang, Q.; Yang, F. Cordycepin negatively modulates lipopolysaccharide-induced cytokine production by up-regulation of heme oxygenase-1. Int. Immunopharmacol. 2017, 47, 20–27. [Google Scholar] [CrossRef]
- Li, Y.; Li, K.; Mao, L.; Han, X.; Zhang, K.; Zhao, C.; Zhao, J. Cordycepin inhibits LPS-induced inflammatory and matrix degradation in the intervertebral disc. PeerJ 2016, 4, e1992. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Zhang, X.; Zhang, G.Q.; Bao, W.P.; Zhang, Y.Y.; Zhang, M.; Zhou, X. Cordycepin inhibits airway remodeling in a rat model of chronic asthma. Biomed. Pharm. 2017, 88, 335–341. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choi, H.Y.; Baik, H.H.; Ju, B.G.; Kim, W.K.; Yune, T.Y. Cordycepin-enriched WIB-801C from Cordyceps militaris improves functional recovery by attenuating blood-spinal cord barrier disruption after spinal cord injury. J. Ethnopharmacol. 2017, 203, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Zhang, T.; Feng, Q.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur J. Pharm. 2018, 818, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Araldi, D.; Ferrari, L.F.; Levine, J.D. Gi-protein-coupled 5-HT1B/D receptor agonist sumatriptan induces type I hyperalgesic priming. Pain 2016, 157, 1773–1782. [Google Scholar] [CrossRef]
- Liu, Z.B.; Liu, C.; Zeng, B.; Huang, L.P.; Yao, L.H. Modulation Effects of Cordycepin on Voltage-Gated Sodium Channels in Rat Hippocampal CA1 Pyramidal Neurons in the Presence/Absence of Oxygen. Neural. Plast. 2017, 2017, 2459053. [Google Scholar] [CrossRef]
- Yong, T.; Chen, S.; Xie, Y.; Chen, D.; Su, J.; Shuai, O.; Jiao, C.; Zuo, D. Cordycepin, a Characteristic Bioactive Constituent in Cordyceps militaris,Ameliorates Hyperuricemia through URAT1 in Hyperuricemic Mice. Front. Microbiol. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Aramwit, P.; Porasuphatana, S.; Srichana, T.; Nakpheng, T. Toxicity evaluation of cordycepin and its delivery system for sustained in vitro anti-lung cancer activity. Nanoscale Res. Lett. 2015, 10, 152. [Google Scholar] [CrossRef]
- Tsai, Y.J.; Lin, L.C.; Tsai, T.H. Pharmacokinetics of adenosine and cordycepin, a bioactive constituent of Cordyceps sinensis in rat. J. Agric. Food Chem. 2010, 58, 4638–4643. [Google Scholar] [CrossRef]
- Lee, J.B.; Adrower, C.; Qin, C.; Fischer, P.M.; de Moor, C.H.; Gershkovich, P. Development of Cordycepin Formulations for Preclinical and Clinical Studies. Aaps Pharmscitech. 2017, 18, 3219–3226. [Google Scholar] [CrossRef] [PubMed]
- OncoVista, I.; AAIPharma, I. Study of Cordycepin Plus Pentostatin in Patients With Refractory TdT-Positive Leukemia. Available online: https://clinicaltrials.gov/show/NCT00709215 (accessed on 9 January 2009).
- De Clercq, E. Curious (Old and New) Antiviral Nucleoside Analogues with Intriguing Therapeutic Potential. Curr. Med. Chem 2015, 22, 3866–3880. [Google Scholar] [CrossRef]
- Wei, H.P.; Ye, X.L.; Chen, Z.; Zhong, Y.J.; Li, P.M.; Pu, S.C.; Li, X.G. Synthesis and pharmacokinetic evaluation of novel N-acyl-cordycepin derivatives with a normal alkyl chain. Eur. J. Med. Chem. 2009, 44, 665–669. [Google Scholar] [CrossRef]
- Vodnala, S.K.; Lundback, T.; Yeheskieli, E.; Sjoberg, B.; Gustavsson, A.L.; Svensson, R.; Olivera, G.C.; Eze, A.A.; de Koning, H.P.; Hammarstrom, L.G.; et al. Structure-activity relationships of synthetic cordycepin analogues as experimental therapeutics for African trypanosomiasis. J. Med. Chem. 2013, 56, 9861–9873. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.G.; Zhang, D.N.; Cao, L.; Han, Y.B. Highly efficient and regioselective acylation of pharmacologically interesting cordycepin catalyzed by lipase in the eco-friendly solvent 2-methyltetrahydrofuran. Bioresour. Technol. 2013, 133, 82–86. [Google Scholar] [CrossRef]
- Yang, Q.Z.; Yang, J.; Zhang, C.K. Synthesis and properties of cordycepin intercalates of Mg-Al-nitrate layered double hydroxides. Int J. Pharm. 2006, 326, 148–152. [Google Scholar] [CrossRef]
- Bi, Y.E.; Zhou, Y.; Wang, M.; Li, L.; Lee, R.J.; Xie, J.; Teng, L. Targeted Delivery of Cordycepin to Liver Cancer Cells Using Transferrin-conjugated Liposomes. Anticancer Res. 2017, 37, 5207–5214. [Google Scholar] [PubMed]
- Federici, C.; Petrucci, F.; Caimi, S.; Cesolini, A.; Logozzi, M.; Borghi, M.; D’Ilio, S.; Lugini, L.; Violante, N.; Azzarito, T.; et al. Exosome release and low pH belong to a framework of resistance of human melanoma cells to cisplatin. PLoS ONE 2014, 9, e88193. [Google Scholar] [CrossRef] [PubMed]
- Chapuy, B.; Koch, R.; Radunski, U.; Corsham, S.; Cheong, N.; Inagaki, N.; Ban, N.; Wenzel, D.; Reinhardt, D.; Zapf, A.; et al. Intracellular ABC transporter A3 confers multidrug resistance in leukemia cells by lysosomal drug sequestration. Leukemia 2008, 22, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Chung, S.H.; Lee, W.S.; Lee, J.Y.; Kim, J.L.; Lee, K.T. Immunostimulatory effects of cordycepin-enriched WIB-801CE from Cordyceps militaris in splenocytes and cyclophosphamide-induced immunosuppressed mice. Phytother. Res. 2018, 32, 132–139. [Google Scholar] [CrossRef]
- Taylor, J.W. One Fungus = One Name: DNA and fungal nomenclature twenty years after PCR. Ima Fungus 2011, 2, 113–120. [Google Scholar] [CrossRef]
- Berkeley, M.J. On some entomogenous Sphaeriae. Lond. J. Bot. 1843, 2, 205–211. [Google Scholar] [CrossRef]
- Saccardo, P.A. Enumeratio pyrenomycetum hypocreaceorum huscusque congnitorum systemate xarpologico dispositorum. Michelia 1878, 1, 277–325. [Google Scholar]
- Hu, T.; Jiang, C.; Huang, Q.; Sun, F. A comb-like branched β-d-glucan produced by a Cordyceps sinensis fungus and its protective effect against cyclophosphamide-induced immunosuppression in mice. Carbohydr. Polym. 2016, 142, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef]
- Wei, J.C.; Wei, X.L.; Zheng, W.F.; Guo, W.; Liu, R. Species identification and component detection of Ophiocordyceps sinensis cultivated by modern industry. Mycosystema 2016, 35, 404–410. [Google Scholar]
- Jiangm, Y.; Yao, Y.J. Names related to Cordyceps sinensis anamorph. Mycotaxon 2002, 84, 245–254. [Google Scholar]
- Liu, Z.-Y.; Yao, Y.-J.; Qi Liang, Z.; Liu, A.-Y.; Pegler, D.N.; Chase, M.W. Molecular evidence for the anamorph—teleomorph connection in Cordyceps sinensis. Mycol. Res. 2001, 105, 827–832. [Google Scholar] [CrossRef]
- Jiapeng, T.; Yiting, L.; Li, Z. Optimization of fermentation conditions and purification of cordycepin from Cordyceps militaris. Prep. Biochem. Biotechnol. 2014, 44, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical composition and nutritional and medicinal value of fruit bodies and submerged cultured mycelia of culinary-medicinal higher Basidiomycetes mushrooms. Int J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Li, Y.; Zhu, Y.; Luo, H.; Li, C.; Xu, X.; Sun, C.; Song, J.; Shi, L.; He, L.; et al. Transcriptome analysis of the Ophiocordyceps sinensis fruiting body reveals putative genes involved in fruiting body development and cordycepin biosynthesis. Genomics 2014, 103, 154–159. [Google Scholar] [CrossRef]
- Jin, J.; Kang, W.; Zhong, C.; Qin, Y.; Zhou, R.; Liu, H.; Xie, J.; Chen, L.; Qin, Y.; Zhang, S. The pharmacological properties of Ophiocordyceps xuefengensis revealed by transcriptome analysis. J. Ethnopharmacol. 2018, 219, 195–201. [Google Scholar] [CrossRef]
- Hu, H.; Xiao, L.; Zheng, B.; Wei, X.; Ellis, A.; Liu, Y.M. Identification of chemical markers in Cordyceps sinensis by HPLC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 8059–8066. [Google Scholar] [CrossRef]
- Jin, J.; Zhong, C.; Qin, Y.; Cai, Y.; Zhen, L.; Shen, B.; Chen, L.; Wan, D.; Qin, Y.; Zhang, S. A new cordycepin-producing caterpillar fungus Ophiocordyceps xuefengensis with artificial infection to the host, cultivation of mycelia and stromata. FEMS Microbiol. Lett. 2017, 364, fnx181. [Google Scholar] [CrossRef]
- Masuda, M.; Urabe, E.; Honda, H.; Sakurai, A.; Sakakibara, M. Enhanced production of cordycepin by surface culture using the medicinal mushroom Cordyceps militaris. Enzym. Microb. Technol. 2007, 40, 1199–1205. [Google Scholar] [CrossRef]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Effects of additives on Cordycepin production using a Cordyceps militaris mutant induced by ion beam irradiation. Afr. J. Biotechnol. 2009, 8, 3041–3047. [Google Scholar]
- Yang, F.Q.; Ge, L.; Yong, J.W.; Tan, S.N.; Li, S.P. Determination of nucleosides and nucleobases in different species of Cordyceps by capillary electrophoresis-mass spectrometry. J. Pharm. Biomed. Anal. 2009, 50, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Das, S.K.; Fujihara, S.; Hatashita, M.; Sakurai, A. Production of cordycepin by a repeated batch culture of a Cordyceps militaris mutant obtained by proton beam irradiation. J. Biosci. Bioeng. 2011, 111, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Y.; Zhang, L.; Wu, J. Characterizations of a new Cordyceps cicadae isolate and production of adenosine and cordycepin. Braz J. Microbiol. 2012, 43, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Pan, M.C.; Chang, C.K.; Chang, S.W.; Hsieh, C.W. Optimization of ultrasonic-assisted extraction of cordycepin from Cordyceps militaris using orthogonal experimental design. Molecules 2014, 19, 20808–20820. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.P.; Wang, J.J.; Wei, B.L.; Lee, C.L. Effect of the salts of deep ocean water on the production of cordycepin and adenosine of Cordyceps militaris-fermented product. Amb Express 2015, 5, 140. [Google Scholar] [CrossRef] [PubMed]
- Sari, N.; Suparmin, A.; Kato, T.; Park, E.Y. Improved cordycepin production in a liquid surface culture of Cordyceps militaris isolated from wild strain. Biotechnol. Bioprocess. Eng. 2016, 21, 595–600. [Google Scholar] [CrossRef][Green Version]
- Kang, N.; Lee, H.H.; Park, I.; Seo, Y.S. Development of High Cordycepin-Producing Cordyceps militaris Strains. Mycobiology 2017, 45, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Suparmin, A.; Kato, T.; Dohra, H.; Park, E.Y. Insight into cordycepin biosynthesis of Cordyceps militaris: Comparison between a liquid surface culture and a submerged culture through transcriptomic analysis. PLoS ONE 2017, 12, e0187052. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Zhao, X.; Ling, J. Genome shuffling improved the nucleosides production in Cordyceps kyushuensis. J. Biotechnol. 2017, 260, 42–47. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Liu, K.; Liu, Q.; Yang, Y.; Dong, C. Heat and light stresses affect metabolite production in the fruit body of the medicinal mushroom Cordyceps militaris. Appl. Microbiol. Biotechnol. 2018, 102, 4523–4533. [Google Scholar]
- Lin, L.T.; Lai, Y.J.; Wu, S.C.; Hsu, W.H.; Tai, C.J. Optimal conditions for cordycepin production in surface liquid-cultured Cordyceps militaris treated with porcine liver extracts for suppression of oral cancer. J. Food Drug Anal. 2018, 26, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Wen, T.; Kang, J.; Lei, B.; Hyde, K. Cordyceps pruinosa produces cordycepin and N6-(2-hydroxyethyl)-adenosine in culture. Arch. Biol. Sci. 2014, 66, 1411–1421. [Google Scholar] [CrossRef]
- Pang, F.; Wang, L.; Jin, Y.; Guo, L.; Song, L.; Liu, G.; Feng, C. Transcriptome analysis of Paecilomyces hepiali at different growth stages and culture additives to reveal putative genes in cordycepin biosynthesis. Genomics 2018, 110, 162–170. [Google Scholar] [CrossRef]
- Xia, E.H.; Yang, D.R.; Jiang, J.J.; Zhang, Q.J.; Liu, Y.; Liu, Y.L.; Zhang, Y.; Zhang, H.B.; Shi, C.; Tong, Y.; et al. The caterpillar fungus, Ophiocordyceps sinensis, genome provides insights into highland adaptation of fungal pathogenicity. Sci. Rep. 2017, 7, 1806. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Dong, C. Photo morphogenesis and photo response of the blue-light receptor gene Cmwc-1 in different strains of Cordyceps militaris. Fems Microbiol. Lett. 2014, 352, 190–197. [Google Scholar] [CrossRef][Green Version]
- Wang, F.; Song, X.; Dong, X.; Zhang, J.; Dong, C. DASH-type cryptochromes regulate fruiting body development and secondary metabolism differently than CmWC-1 in the fungus Cordyceps militaris. Appl. Microbiol. Biotechnol. 2017, 101, 4645–4657. [Google Scholar] [CrossRef] [PubMed]
- Kredich, N.M.; Guarino, A.J. Studies on the biosynthesis of cordycepin. Biochim. Et Biophys. Acta 1961, 47, 529–534. [Google Scholar] [CrossRef]
- Chassy, B.M.; Suhadolnik, R.J. Nucleoside antibiotics. IV. Metabolic fate of adenosine and cordycepin by Cordyceps militaris during cordycepin biosynthesis. Biochim. Et Biophys. Acta 1969, 182, 307–315. [Google Scholar] [CrossRef]
- Lennon, M.B.; Suhadolnik, R.J. Biosynthesis of 3′-deoxyadenosine by Cordyceps militaris. Mechanism of reduction. Biochim. Et Biophys. Acta 1976, 425, 532–536. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.-d.; Wang, W.; Zhong, J.-J. Enhancement of cordycepin production in submerged cultures of Cordyceps militaris by addition of ferrous sulfate. Biochem. Eng. J. 2012, 60, 30–35. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y. The strategies for increasing cordycepin production of Cordyceps militaris by liquid fermentation. Fungal Genom. Biol. 2016, 6, 134. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Huang, I.C.; Chen, T.-Y. Cordyceps s.L. (Ascomycetes) species used as medicinal mushrooms are closely related with higher ability to produce cordycepin. Int. J. Med. Mushrooms 2016, 17, 1077–1085. [Google Scholar] [CrossRef]
- Vongsangnak, W.; Raethong, N.; Mujchariyakul, W.; Nguyen, N.N.; Leong, H.W.; Laoteng, K. Genome-scale metabolic network of Cordyceps militaris useful for comparative analysis of entomopathogenic fungi. Gene 2017, 626, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Ahmad, S.; Park, E.Y. Functional Analysis of Ribonucleotide Reductase from Cordyceps militaris Expressed in Escherichia coli. Appl. Biochem. Biotechnol. 2017, 182, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wan, D.; Xu, G.; Wang, G.; Ma, H.; Wang, T.; Gao, Y.; Qi, J.; Chen, X.; Zhu, J.; et al. An unusual protector-prote´ ge´ strategy for the biosynthesis of purine nucleoside antibiotics. Cell Chem. Biol. 2017, 24, 171–181. [Google Scholar] [CrossRef]
- Li, G.; Nakagome, I.; Hirono, S.; Itoh, T.; Fujiwara, R. Inhibition of adenosine deaminase (ADA)-mediated metabolism of cordycepin by natural substances. Pharm. Res. Perspect 2015, 3, e00121. [Google Scholar] [CrossRef]
- Yin, Y.; Yu, G.; Chen, Y.; Jiang, S.; Wang, M.; Jin, Y.; Lan, X.; Liang, Y.; Sun, H. Genome-wide transcriptome and proteome analysis on different developmental stages of Cordyceps militaris. PLoS ONE 2012, 7, e51853. [Google Scholar] [CrossRef]
- Tudzynski, P.; Heller, J.; Siegmund, U. Reactive oxygen species generation in fungal development and pathogenesis. Curr. Opin. Microbiol. 2012, 15, 653–659. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Z.; Li, Q. Research progress of host insect of Cordyceps and its raising technique. World Chin. Med. 2017, 12, 3142–3150. [Google Scholar]
- Zhang, Y.; Zhang, S.; Li, Y.; Ma, S.; Wang, C.; Xiang, M.; Liu, X.; An, Z.; Xu, J.; Liu, X. Phylogeography and evolution of a fungal-insect association on the Tibetan Plateau. Mol. Ecol. 2014, 23, 5337–5355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ding, Y.F.; Yang, L.; Yu, J.H.; Liu, G.M.; Wang, X.M.; Zhang, S.Y.; Yu, D.; Song, L.; Zhang, H.X.; et al. Integrated omics study delineates the dynamics of lipid droplets in Rhodococcus opacus PD630. Nucleic Acids Res. 2014, 42, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Covington, B.C.; McLean, J.A.; Bachmann, B.O. Comparative mass spectrometry-based metabolomics strategies for the investigation of microbial secondary metabolites. Nat. Prod. Rep. 2017, 34, 6–24. [Google Scholar] [CrossRef]
- Gesmundo, N.J.; Sauvagnat, B.; Curran, P.J.; Richards, M.P.; Andrews, C.L.; Dandliker, P.J.; Cernak, T. Nanoscale synthesis and affinity ranking. Nature 2018, 557, 228–232. [Google Scholar] [CrossRef]
- Baslan, T.; Hicks, J. Unravelling biology and shifting paradigms in cancer with single-cell sequencing. Nat. Rev. Cancer 2017, 17, 557–569. [Google Scholar] [CrossRef]
- Packer, J.; Trapnell, C. Single-Cell Multi-omics: An Engine for New Quantitative Models of Gene Regulation. Trends Genet. 2018, 34, 653–665. [Google Scholar] [CrossRef]
- Rosenberg, A.B.; Roco, C.M.; Muscat, R.A.; Kuchina, A.; Sample, P.; Yao, Z.; Graybuck, L.T.; Peeler, D.J.; Mukherjee, S.; Chen, W.; et al. Single-cell profiling of the developing mouse brain and spinal cord with split-pool barcoding. Science 2018, 360, 176–182. [Google Scholar] [CrossRef]
- Miller, M.A.; Weissleder, R. Imaging of anticancer drug action in single cells. Nat. Rev. Cancer 2017, 17, 399–414. [Google Scholar] [CrossRef]
- Jing, M.; Zhang, P.; Wang, G.; Feng, J.; Mesik, L.; Zeng, J.; Jiang, H.; Wang, S.; Looby, J.C.; Guagliardo, N.A.; et al. A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies. Nat. Biotechnol. 2018, 36, 726–737. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential Applications | COR Actions |
|---|---|
| Specific inhibitor of mRNA polyadenylation [7,117] (+++) | Inhibition of PolyA formation of mRNA. |
| Anticancer activity(+++) | Inhibition of leukemia [7,33,35,38,41,42,46,118,119,120], thyroid cancer [31], breast cancer [32,51,57], lung cancer [9,40,50,54,121], prostate cancer [49,122], hepatocellular cancer [37,123,124], colonic cancer [34], gallbladder cancer [11,125], renal cancer [53], cervical cancer [43], glioblastoma cancer [10], oral cancer [52], brain cancer [36] and glioma [60]. |
| Antitumor activity(++) | Inhibition of multiple myeloma [58], renal tumor [61], leydig tumor [59,64] and EBV-infected tumor growth-infected tumor growth [17]. |
| Anti-inflammatory activity(+++) | Inhibition of human osteoarthritis [66], Kawasaki disease [39], lipopolysaccharide (LPS)-induced inflammation [65,126], asthma [67], intervertebral disc degeneration-induced inflammation [127], traumatic brain injury-induced inflammation [68], airway inflammation [128], spinal cord injury-induced inflammation [129], atopic dermatitis [69], local anesthesia (lidocaine) induced spinal cord neurotoxicity [70] and acute lung injury-induced inflammation [130]. |
| Anti-oxidant activity(+) | Inhibition of radical-induced oxidative damage [75], tubulointerstitial fibrosis [73] and Parkinson’s disease [74]. |
| Pathogen growth inhibition | Inhibition of Bacillus subtilis(+++) [76], adenovirus(++) [77,78], murine leukovirus(++) [79], murine sarcoma virus(+++) [80], Newcastle disease virus(+++) [81], human poliovirus (+++) [82], tobacco mosaic virus(+++) [83], vaccinia virus(+++) [84], Hepatitis C virus by terminating PolyA formation of D(+) RNA(+++) [85], Clostridium paraputrificum(+++) [86], Clostridium perfringens(+++) [86] and Candida(++) [87]. |
| Other medicinal potential | Induction of cell death of Plutella xylostella(++) [89,90,91,92], chondrogenesis(++) [112] and insulin sensitivity(+++) [107]. |
| Inhibition of skin photoaging(+++) [95], hyperlipidemia(++) [107,108], endoplasmic reticulum-induced injury(+++) [62], fibrosis(++) [96,97], proliferation of vascular smooth muscle(++) [100], adipogenesis-induced hepatotoxicity(+++) [105], pain(+++) [101,102,103,131], depressant(+++) [109], hypoxia-induced neuronal injuries(++) [132], myocardial infarction(+++) [98], cerebral ischemia injury(+++) [99], skeletal muscle fatigue(+++) [104], bone loss(+++) [110], hyperuricemia(+++) [133]. | |
| Simple structure, but high potency | Derivatives of COR are easy to design as a result of the simple structure of COR. Previous work demonstrated that COR exhibits higher potency than zhankuic acid A, adenosine [124] and N(6)-(2-hydroxyethyl)adenosine [65]. |
| Ligand | COR and COR-TP can be used as ligands in the molecular replacement experiments that solve the molecular structures of SAHase in B. elkanii [113] and human Poly(A) polymeraseγ [114], respectively. |
| RNA elongation inhibitor | COR was used as an RNA elongation inhibitor and bromine in BrUTP was used to elucidate the structure of active rRNA genes in the nucleolus [13]. |
| Multiple targets recognized by COR | COR can recognize many medicinal targets. In addition, COR can inhibit PolyA formation, activates protein kinases [11] and stimulates protein phosphatases [10]. |
| Strain | Strain ID | Yield | Mesurement | Strain Source | References | |
|---|---|---|---|---|---|---|
| CM | (W)NBRC 9787 | 2500 | AL | D | NITE, Japan | [161] |
| CM | (M)G81-3 | 8570 | AL | D | UF, Japan | [162] |
| CM | (W)- | 2.276 | BS | E | UM-SAR, China | [163] |
| CM | (M)G81-3 | 8600 | AL | D | UF, Japan | [164] |
| CC | (W)- | 1.398 | BS | E | SNJM-HB, China | [165] |
| CM | (W)- | 1.743 | BS | E | FNS and IE-UHF | [156] |
| CM | (W)14014 | 7350 | AL | D | CCICC, China | [155] |
| CM | (M)G81-3 | 14300 | AL | D | UF, Japan | [22] |
| CM | (W)- | 7.04 | BS | E | MDU, Taiwan | [166] |
| CS | (W)- | 0.0068–0.029 | BS | E | QH, HB and AH, china | [159] |
| CM | (W)BCRC 32219 | 1.7 | AL | D | BCRC, Hsinchu, Taiwan | [167] |
| CM | (W)NBRC 10352-3 | 6200 | AL | D | SU, Japan | [168] |
| OX | (W)HACM 001 | 0.0371 | BS | E | XFM-HN, China | [160] |
| CM | (Md)KACC44455+SPNU1006 | 6.63 | BS | E | KACC and SPNU, Korea | [169] |
| CM | (W)NBRC 103752 | 4920 | AL | D | BRC, NITE, Tokyo, Japan | [170] |
| CK | (GS)Y9 | 0.7135 | AS | C | SDU, China | [171] |
| CM | (W)CGMCC 3.16321 | 5.56 | BS | E | Beijing, China | [172] |
| CM | (W)BCRC34380 | 3483 | AL | C | Hsinchu, Taiwan | [173] |
| CP | (W)GZUCC 8552 | 5.311 | AS | C | BM, Guizhou, China | [174] |
| CM | (W)CGMCC 3.16321 | 5.56 | BS | E | Beijing, China | [172] |
| CM | (W)No.20130508 | 9.45 | BS | E | Nanjing, China | [23] |
| PH | (W)Isolated strain | 0.0346 | AL | D | Qinghai, China | [175] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, P.; Li, X.; Yang, H.; Wang, Z.-Y.; Lu, D. Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules 2019, 24, 2231. https://doi.org/10.3390/molecules24122231
Qin P, Li X, Yang H, Wang Z-Y, Lu D. Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules. 2019; 24(12):2231. https://doi.org/10.3390/molecules24122231
Chicago/Turabian StyleQin, Peng, XiangKai Li, Hui Yang, Zhi-Ye Wang, and DengXue Lu. 2019. "Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi" Molecules 24, no. 12: 2231. https://doi.org/10.3390/molecules24122231
APA StyleQin, P., Li, X., Yang, H., Wang, Z.-Y., & Lu, D. (2019). Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules, 24(12), 2231. https://doi.org/10.3390/molecules24122231