Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro




Abstract
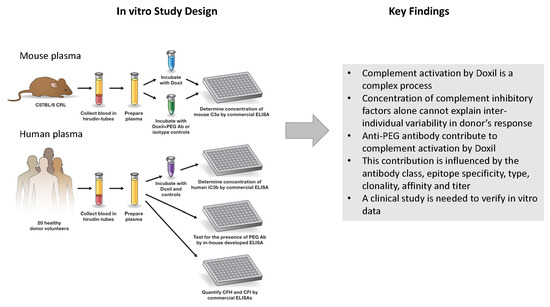
1. Introduction
2. Results and Discussion
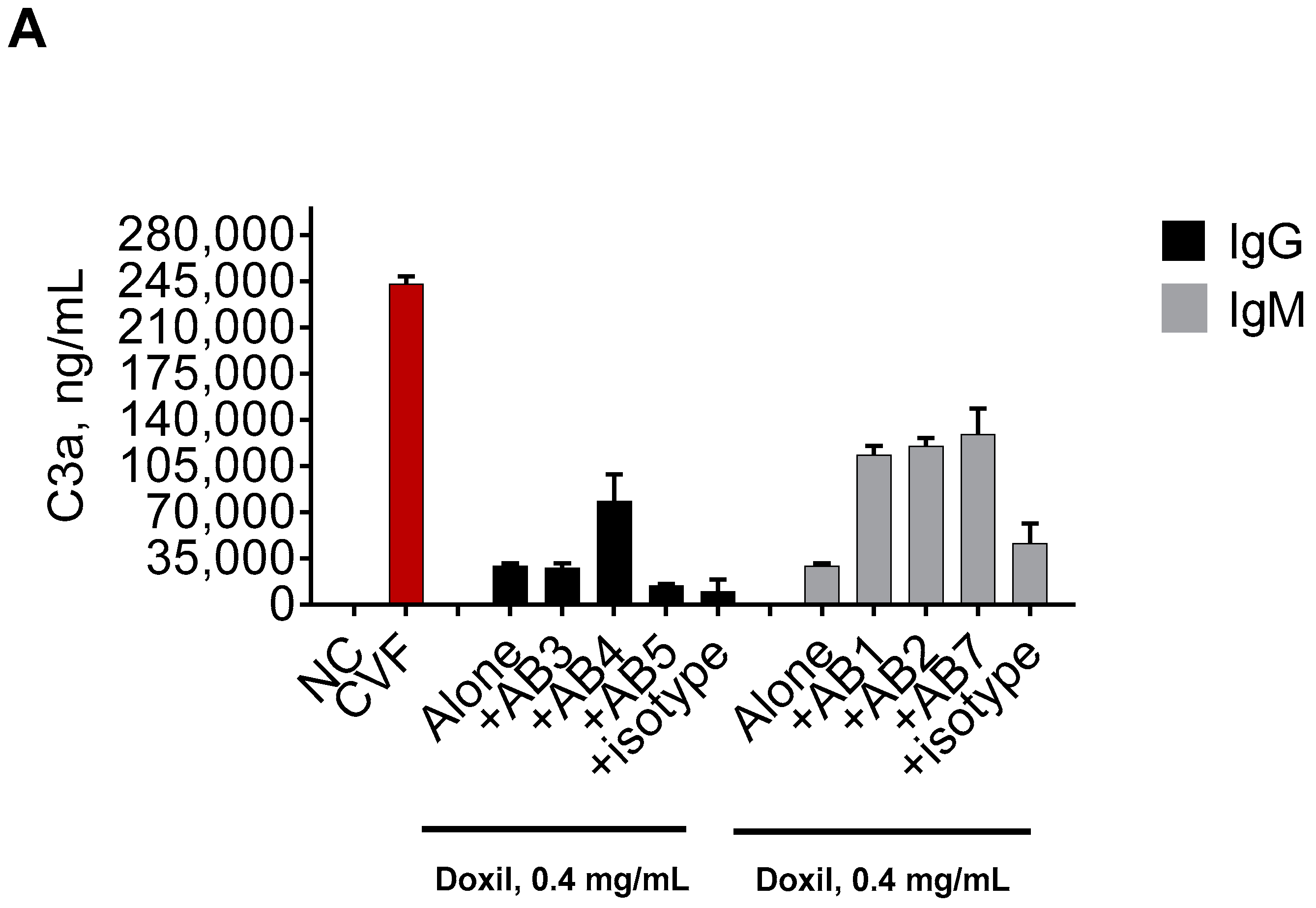
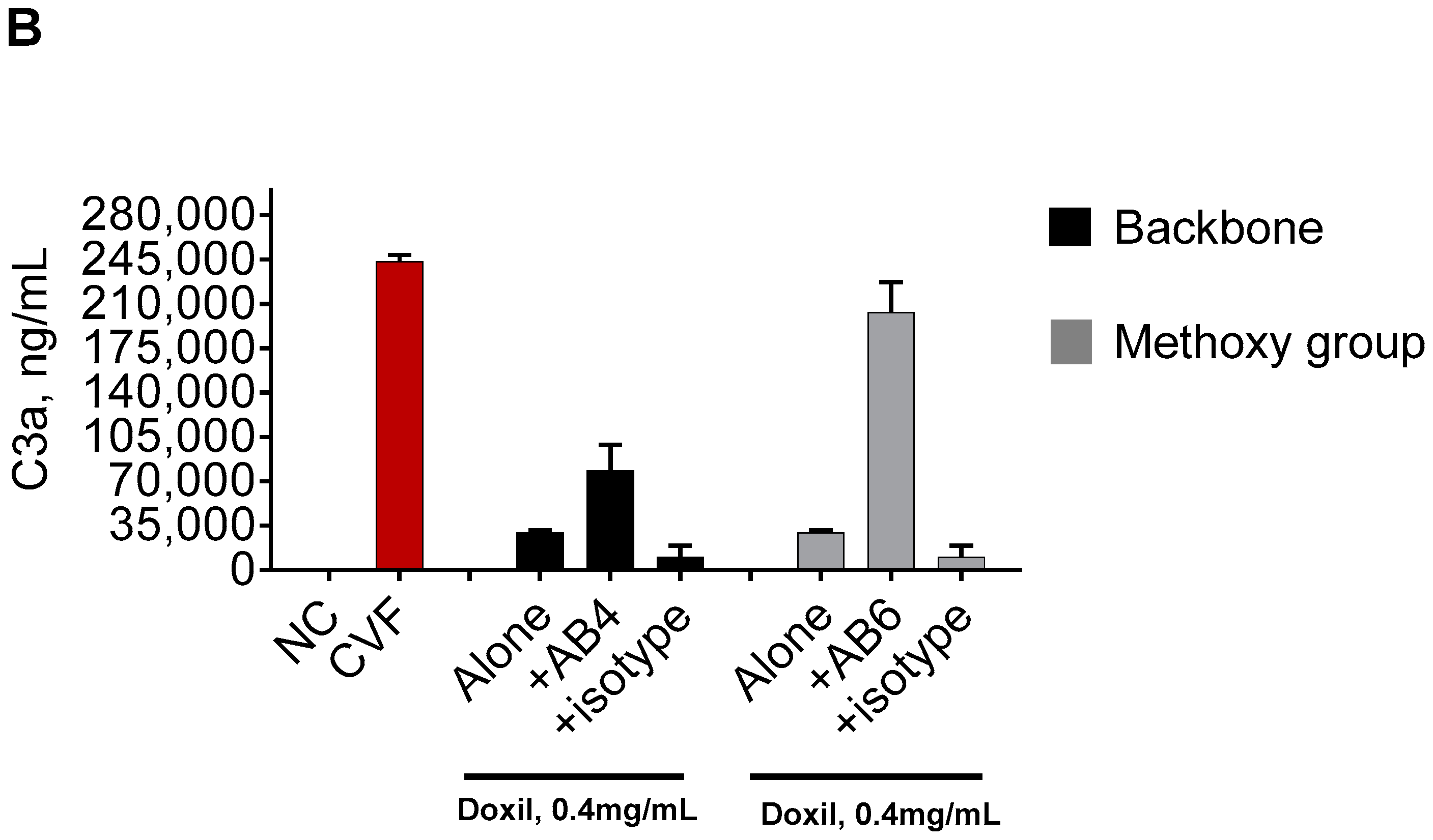
2.1. Mouse Monoclonal Anti-PEG Antibody May Contribute to the Complement Activation by Doxil
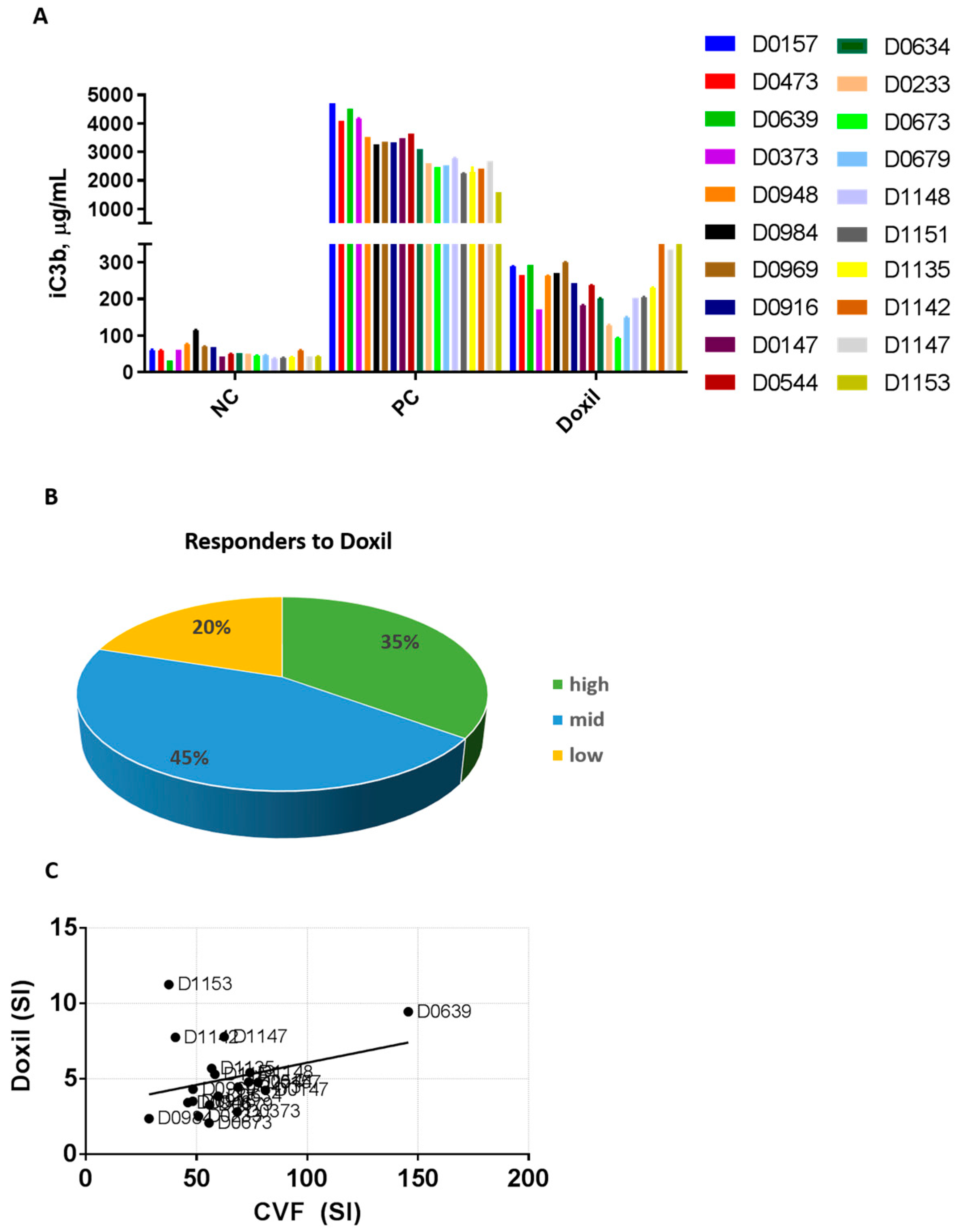
2.2. Plasma from Human Donors Shows Variability in the Complement Activation by Doxil and Cobra Venom Factor
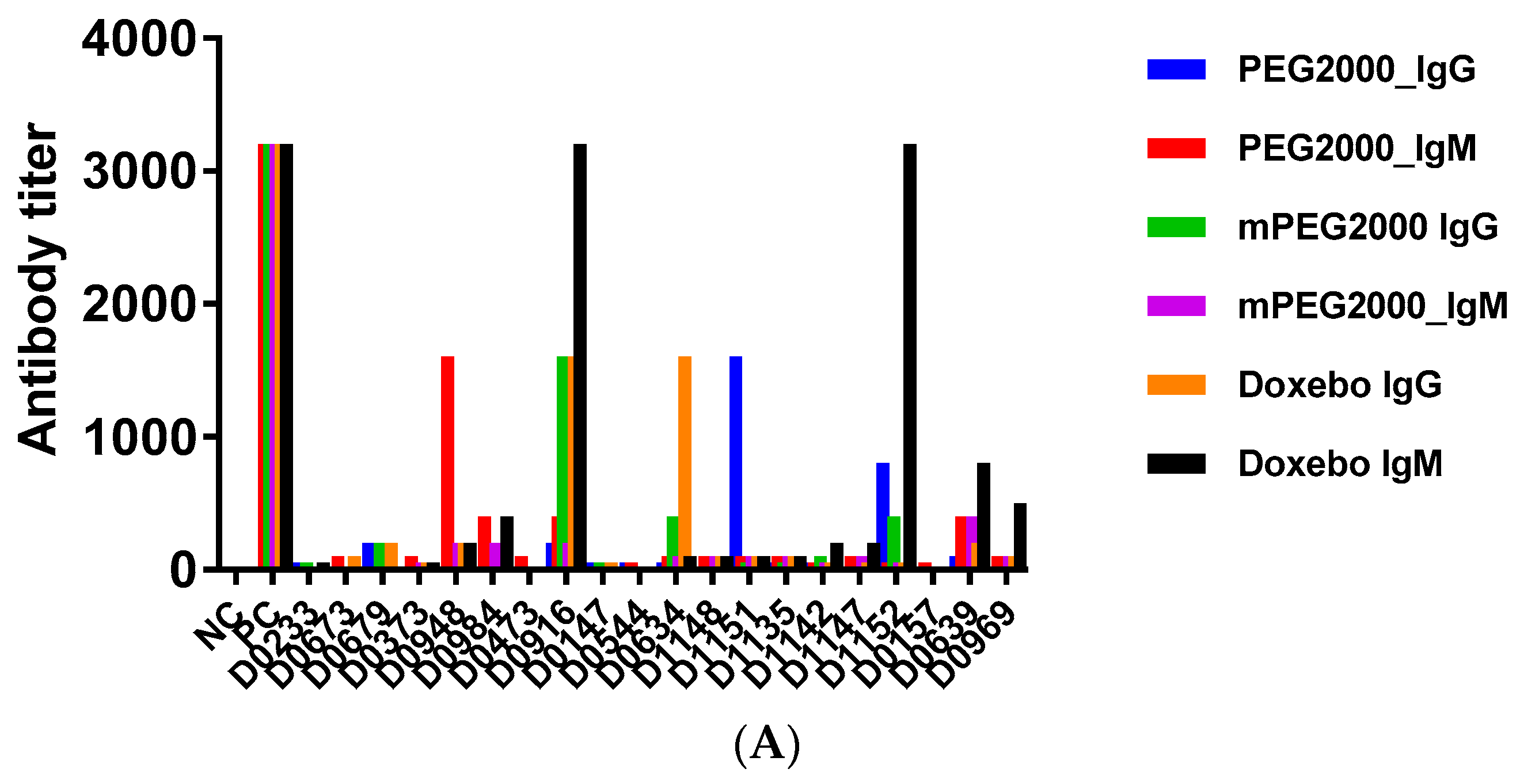
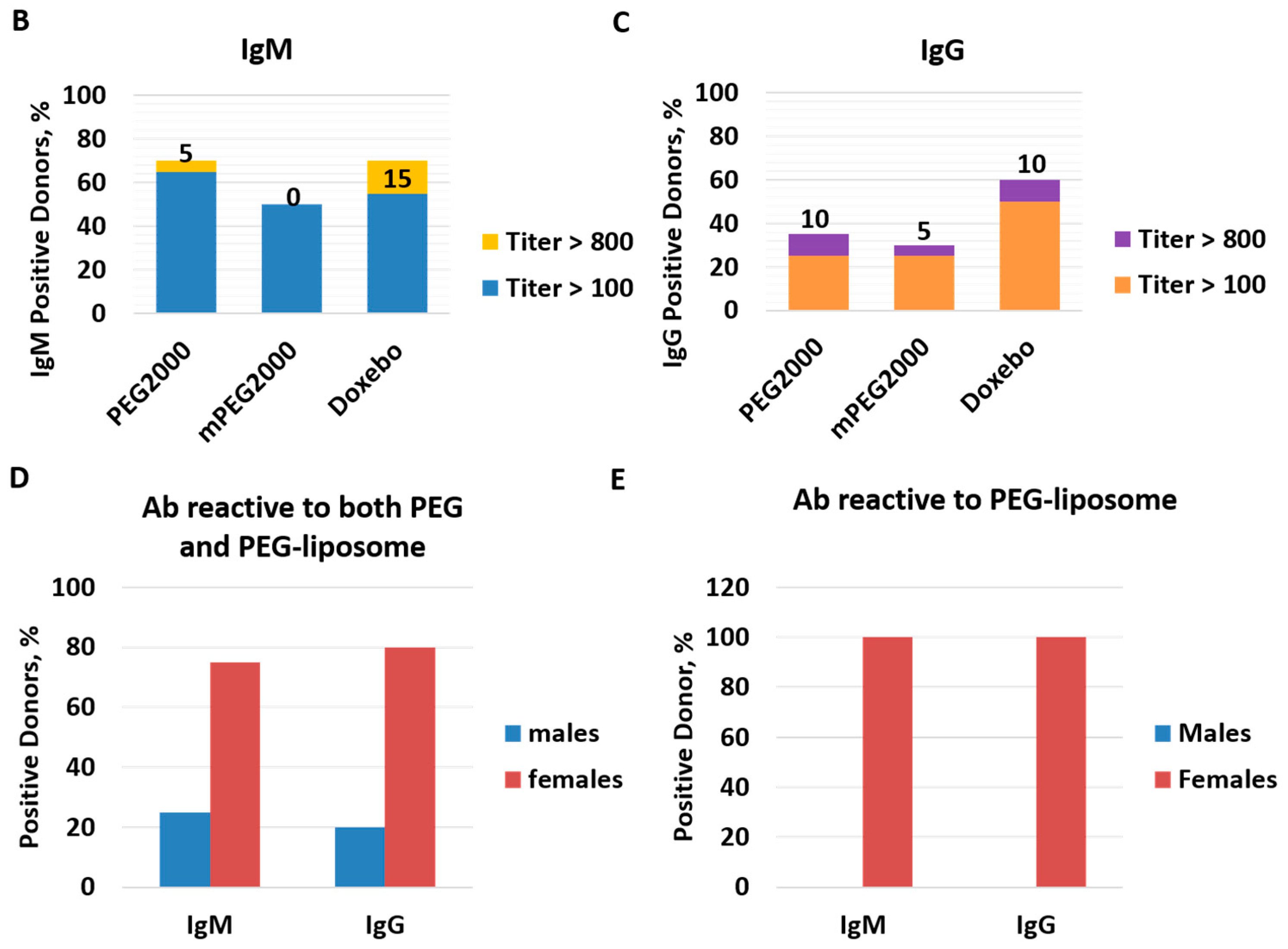
2.3. Pre-Existing PEG-Reactive Antibodies are Present in Plasma of Healthy Donor Volunteers
2.4. Complement Activation by Doxil in Healthy Human Plasma Does Not Correlate with the Titer of Pre-Existing PEG-Reactive Antibodies
2.5. Plasma Levels of the Complement Inhibitory Factors I and H Do Not Correlate with Complement Activation
3. Materials and Methods
3.1. Reagents
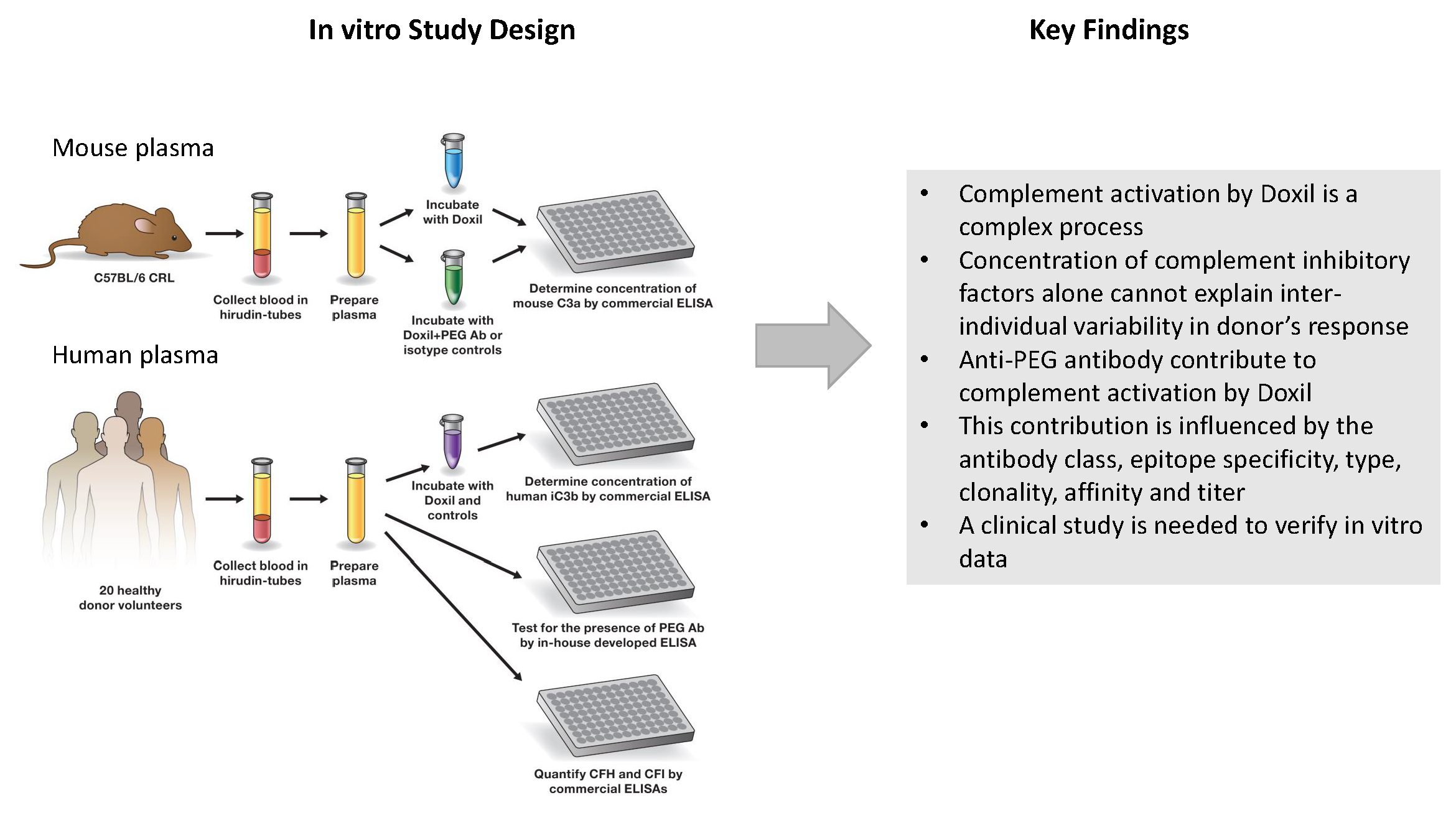
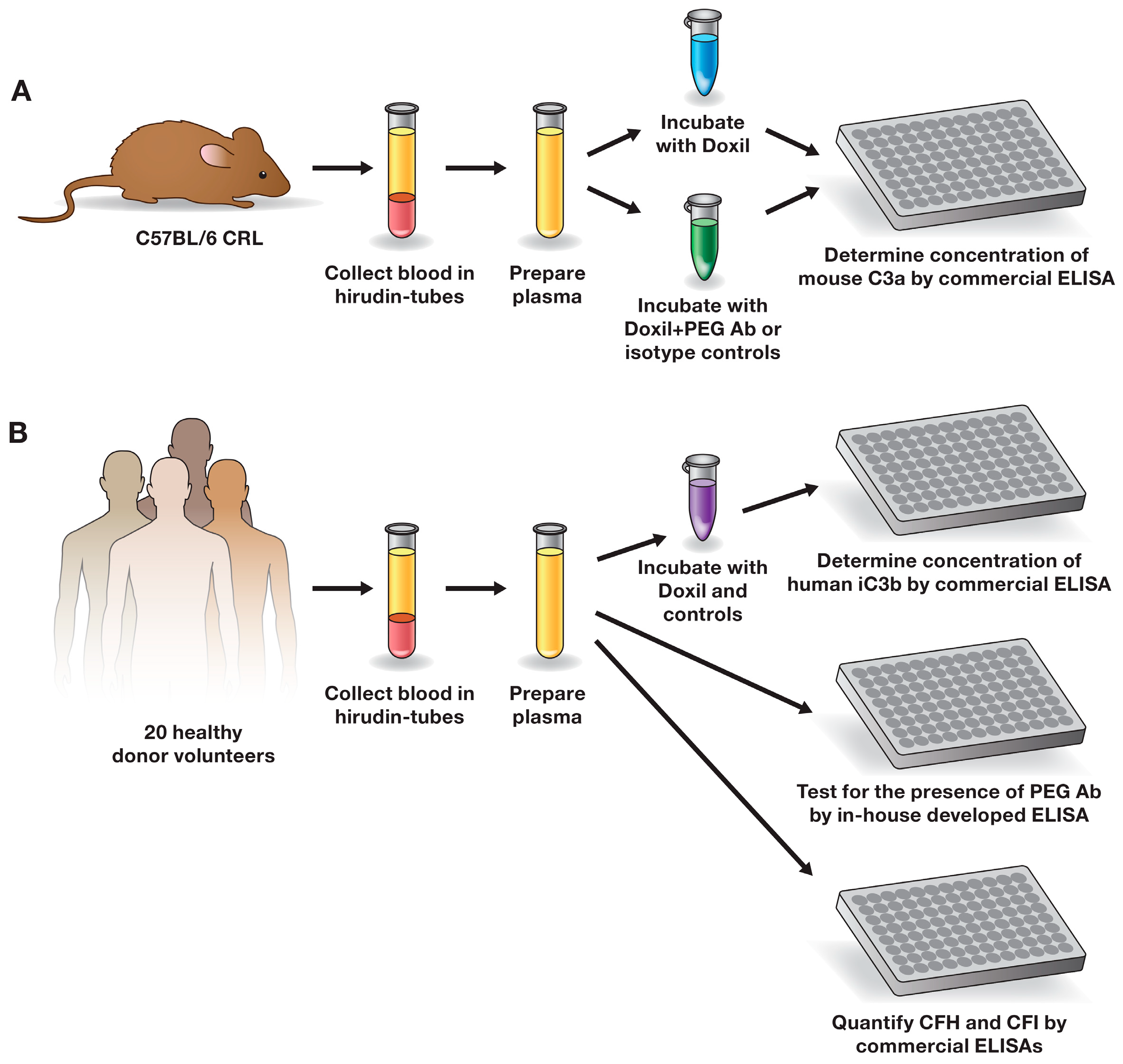
3.2. Research Blood
3.3. Complement Activation
3.4. Detection of PEG-Reactive Antibodies
3.5. Determination of CFH and CFI Plasma Levels
3.6. Statistical and Data Analysis
4. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Terminology
References
- Corominas, M.; Gastaminza, G.; Lobera, T. Hypersensitivity reactions to biological drugs. J. Investig. Allergol. Clin. Immunol. 2014, 24, 212–225. [Google Scholar] [PubMed]
- Joerger, M. Prevention and handling of acute allergic and infusion reactions in oncology. Ann. Oncol. 2012, 23 (Suppl. 10), x313–x319. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.G.O.; Bieber, T.; Dahl, R.; Friedmann, P.S.; Lanier, B.Q.; Lockey, R.F.; Motala, C.; Martell, J.A.O.; Platts-Mills, T.A.; Ring, J.; et al. Revised nomenclature for allergy for global use: Report of the Nomenclature Review Committee of the World Allergy Organization, October 2003. J. Allergy Clin. Immunol. 2004, 113, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Johansson, S.G.O.; Hourihane, J.B.; Bousquet, J.; Bruijnzeel-Koomen, C.; Dreborg, S.; Haahtela, T.; Kowalski, M.L.; Mygind, N.; Ring, J.; van Cauwenberge, P.; et al. A revised nomenclature for allergy: An EAACI position statement from the EAACI nomenclature task force. Allergy 2001, 56, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Agache, I.; Bonini, S.; Burks, A.W.; Calderon, M.; Canonica, W.; Cox, L.; Demoly, P.; Frew, A.J.; O’Hehir, R.; et al. International consensus on allergy immunotherapy. J. Allergy Clin. Immunol. 2015, 136, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.A. Hypersensitivity and immunologic reactions to biologics: Opportunities for the allergist. Ann. Allergy Asthma Immunol. 2016, 117, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.J. Management and preparedness for infusion and hypersensitivity reactions. Oncologist 2007, 12, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Abalde, S.; Gonzalez-Fernandez, A. Nanostructures and Allergy. In Handbook of Immunological Properties of Engineered Nanomaterials; Dobrovolskaia, M.A., McNeil, S.E., Eds.; World Scientific Publishing Ltd.: Singapore, 2013; pp. 517–546. [Google Scholar]
- Picard, M.; Galvao, V.R. Current Knowledge and Management of Hypersensitivity Reactions to Monoclonal Antibodies. J. Allergy Clin. Immunol. Pract. 2017, 5, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Rosello, S.; Blasco, I.; Fabregat, L.G.; Cervantes, A.; Jordan, K. Management of infusion reactions to systemic anticancer therapy: ESMO Clinical Practice Guidelines. Ann. Oncol. 2017, 28, iv100–iv118. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Munoz-Furlong, A.; Campbell, R.L.; Adkinson, N.F., Jr.; Bock, S.A.; Branum, A.; Brown, S.G.; Camargo, C.A., Jr.; Cydulka, R.; Galli, S.J.; et al. Second symposium on the definition and management of anaphylaxis: Summary report—Second National Institute of Allergy and Infectious Disease/Food Allergy and Anaphylaxis Network symposium. J. Allergy Clin. Immunol. 2006, 117, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Landesman-Milo, D.; Peer, D. Altering the immune response with lipid-based nanoparticles. J. Control. Release 2012, 161, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Markman, M.; Kennedy, A.; Webster, K.; Kulp, B.; Peterson, G.; Belinson, J. Paclitaxel-associated hypersensitivity reactions: Experience of the gynecologic oncology program of the Cleveland Clinic Cancer Center. J. Clin. Oncol. 2000, 18, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J. Complement activation-related pseudoallergy: A stress reaction in blood triggered by nanomedicines and biologicals. Mol. Immunol. 2014, 61, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J. Mechanism of nanoparticle-induced hypersensitivity in pigs: Complement or not complement? Drug Discov. Today 2018, 23, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Alving, C.R.; Rosivall, L.; Bunger, R.; Baranyi, L.; Bedocs, P.; Toth, M.; Barenholz, Y. Animal models of complement-mediated hypersensitivity reactions to liposomes and other lipid-based nanoparticles. J. Liposome Res. 2007, 17, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Bedocs, P.; Urbanics, R.; Bunger, R.; Rosivall, L.; Toth, M.; Barenholz, Y. Prevention of infusion reactions to PEGylated liposomal doxorubicin via tachyphylaxis induction by placebo vesicles: A porcine model. J. Control. Release 2012, 160, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Fontana, J.L.; Wassef, N.M.; Mongan, P.D.; Morse, D.S.; Dobbins, D.E.; Stahl, G.L.; Bunger, R.; Alving, C.R. Hemodynamic changes induced by liposomes and liposome-encapsulated hemoglobin in pigs: A model for pseudoallergic cardiopulmonary reactions to liposomes. Role of complement and inhibition by soluble CR1 and anti-C5a antibody. Circulation 1999, 99, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Muggia, F.; Gabizon, A.; Barenholz, Y. Activation of complement by therapeutic liposomes and other lipid excipient-based therapeutic products: Prediction and prevention. Adv. Drug Deliv. Rev. 2011, 63, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Storm, G. Complement activation as a bioequivalence issue relevant to the development of generic liposomes and other nanoparticulate drugs. Biochem. Biophys. Res. Commun. 2015, 468, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Wassef, N.M.; Rudolph, A.S.; Alving, C.R. Complement activation in human serum by liposome-encapsulated hemoglobin: The role of natural anti-phospholipid antibodies. Biochim. Biophys. Acta 1996, 1285, 127–130. [Google Scholar] [CrossRef]
- Noris, M.; Remuzzi, G. Overview of complement activation and regulation. Semin. Nephrol. 2013, 33, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Varela, J.C.; Tomlinson, S. Complement: An overview for the clinician. Hematol. Oncol. Clin. N. Am. 2015, 29, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Szebeni, J. Stealth liposomes and long circulating nanoparticles: Critical issues in pharmacokinetics, opsonization and protein-binding properties. Prog. Lipid Res. 2003, 42, 463–478. [Google Scholar] [CrossRef]
- Unterweger, H.; Dezsi, L.; Matuszak, J.; Janko, C.; Poettler, M.; Jordan, J.; Bauerle, T.; Szebeni, J.; Fey, T.; Boccaccini, A.R.; et al. Dextran-coated superparamagnetic iron oxide nanoparticles for magnetic resonance imaging: Evaluation of size-dependent imaging properties, storage stability and safety. Int. J. Nanomed. 2018, 13, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Baranyi, L.; Savay, S.; Gotze, O.; Alving, C.R.; Bunger, R.; Mongan, P.D. Complement activation during hemorrhagic shock and resuscitation in swine. Shock 2003, 20, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Wassef, N.M.; Rudolph, A.S.; Alving, C.R. Complement activation by liposome-encapsulated hemoglobin in vitro: The role of endotoxin contamination. Artif. Cells Blood Substit. Immobil. Biotechnol. 1995, 23, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.K.; Hempel, G.; Koling, S.; Chan, L.S.; Fisher, T.; Meiselman, H.J.; Garratty, G. Antibody against poly(ethylene glycol) adversely affects PEG-asparaginase therapy in acute lymphoblastic leukemia patients. Cancer 2007, 110, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Garay, R.P.; El-Gewely, R.; Armstrong, J.K.; Garratty, G.; Richette, P. Antibodies against polyethylene glycol in healthy subjects and in patients treated with PEG-conjugated agents. Exp. Opin. Drug Deliv. 2012, 9, 1319–1323. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.W.; Akerblom, E. Polyethylene glycol reactive antibodies in man: Titer distribution in allergic patients treated with monomethoxy polyethylene glycol modified allergens or placebo, and in healthy blood donors. Int. Arch. Allergy Appl. Immunol. 1984, 74, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.M.; Su, Y.C.; Chang, C.J.; Burnouf, P.A.; Chuang, K.H.; Chen, C.H.; Cheng, T.L.; Chen, Y.T.; Wu, J.Y.; Roffler, S.R. Measurement of Pre-Existing IgG and IgM Antibodies against Polyethylene Glycol in Healthy Individuals. Anal. Chem. 2016, 88, 10661–10666. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Lai, S.K. Anti-PEG immunity: Emergence, characteristics, and unaddressed questions. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 655–677. [Google Scholar] [CrossRef] [PubMed]
- Povsic, T.J.; Lawrence, M.G.; Lincoff, A.M.; Mehran, R.; Rusconi, C.P.; Zelenkofske, S.L.; Huang, Z.; Sailstad, J.; Armstrong, P.W.; Steg, P.G.; et al. Pre-existing anti-PEG antibodies are associated with severe immediate allergic reactions to pegnivacogin, a PEGylated aptamer. J. Allergy Clin. Immunol. 2016, 138, 1712–1715. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A. Pre-clinical immunotoxicity studies of nanotechnology-formulated drugs: Challenges, considerations and strategy. J. Control. Release 2015, 220, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Bedocs, P.; Csukas, D.; Rosivall, L.; Bunger, R.; Urbanics, R. A porcine model of complement-mediated infusion reactions to drug carrier nanosystems and other medicines. Adv. Drug Deliv. Rev. 2012, 64, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H.; Brembilla, N.C. Immunogenicity of biologic therapies: Causes and consequences. Exp. Rev. Clin. Immunol. 2018, 14, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Pedras-Vasconcelos, J.A. The Immunogenicity of Therapeutic Proteins-What you Don’t Know Can Hurt YOU and the Patient; CBER, Ed.; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2014. [Google Scholar]
- Spiegelberg, H.L. Biological role of different antibody classes. Int. Arch. Allergy Appl. Immunol. 1989, 90 (Suppl. 1), 22–27. [Google Scholar] [CrossRef] [PubMed]
- Gadjeva, M.G.; Rouseva, M.M.; Zlatarova, A.S.; Reid, K.B.; Kishore, U.; Kojouharova, M.S. Interaction of human C1q with IgG and IgM: Revisited. Biochemistry 2008, 47, 13093–13102. [Google Scholar] [CrossRef] [PubMed]
- Chanan-Khan, A.; Szebeni, J.; Savay, S.; Liebes, L.; Rafique, N.M.; Alving, C.R.; Muggia, F.M. Complement activation following first exposure to pegylated liposomal doxorubicin (Doxil): Possible role in hypersensitivity reactions. Ann. Oncol. 2003, 14, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Baranyi, L.; Savay, S.; Milosevits, J.; Bunger, R.; Laverman, P.; Metselaar, J.M.; Storm, G.; Chanan-Khan, A.; Liebes, L.; et al. Role of complement activation in hypersensitivity reactions to doxil and hynic PEG liposomes: Experimental and clinical studies. J. Liposome Res. 2002, 12, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R. Antibodies to lipids and liposomes: Immunology and safety. J. Liposome Res. 2006, 16, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R.; Wassef, N.M. Naturally occurring antibodies to cholesterol: A new theory of LDL cholesterol metabolism. Immunol. Today 1999, 20, 362–366. [Google Scholar] [CrossRef]
- Alving, C.R.; Wassef, N.M.; Potter, M. Antibodies to cholesterol: Biological implications of antibodies to lipids. Curr. Top. Microbiol. Immunol. 1996, 210, 181–186. [Google Scholar] [PubMed]
- Reis, E.S.; Falcao, D.A.; Isaac, L. Clinical aspects and molecular basis of primary deficiencies of complement component C3 and its regulatory proteins factor I and factor H. Scand. J. Immunol. 2006, 63, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.U.; Jelezarova, E. Complement amplification revisited. Mol. Immunol. 2006, 43, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, P.F.; Jokiranta, T.S.; Hellwage, J.; Koistinen, V.; Meri, S. The factor H protein family. Immunopharmacology 1999, 42, 53–60. [Google Scholar] [CrossRef]
- Shen, L.; Frazer-Abel, A.; Reynolds, P.R.; Giclas, P.C.; Chappell, A.; Pangburn, M.K.; Younis, H.; Henry, S.P. Mechanistic understanding for the greater sensitivity of monkeys to antisense oligonucleotide-mediated complement activation compared with humans. J. Pharmacol. Exp. Ther. 2014, 351, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Sofat, R.; Casas, J.P.; Kumari, M.; Talmud, P.J.; Ireland, H.; Kivimaki, M.; Marmot, M.; Hughes, A.D.; Thom, S.; Ebrahim, S.; et al. Genetic variation in complement factor H and risk of coronary heart disease: Eight new studies and a meta-analysis of around 48,000 individuals. Atherosclerosis 2010, 213, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.F.; Wang, J.F.; Wang, Y.; Zhu, L.G.; Lei, L. Complement factor H Y402H gene polymorphism and coronary heart disease susceptibility: A meta-analysis. Mol. Biol. Rep. 2011, 38, 2933–2938. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; Hancox, L.S.; Taiber, A.J.; Hardisty, L.I.; Hageman, J.L.; Stockman, H.A.; Borchardt, J.D.; Gehrs, K.M.; et al. A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 7227–7232. [Google Scholar] [CrossRef] [PubMed]
- Maller, J.; George, S.; Purcell, S.; Fagerness, J.; Altshuler, D.; Daly, M.J.; Seddon, J.M. Common variation in three genes, including a noncoding variant in CFH, strongly influences risk of age-related macular degeneration. Nat. Genet. 2006, 38, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.X.; Zhang, L.; Zhang, M.; Du, H.; Zhao, L.; Lee, C.; Grob, S.; Lim, S.L.; Hughes, G.; Lee, J.; et al. Complement factor H genotypes impact risk of age-related macular degeneration by interaction with oxidized phospholipids. Proc. Natl. Acad. Sci. USA 2012, 109, 13757–13762. [Google Scholar] [CrossRef] [PubMed]
- Bonyadi, M.; Norouzi, N.; Babaei, E.; Bonyadi, M.H.J.; Javadzadeh, A.; Yaseri, M.; Soheilian, M. Association of polymorphisms of complement factor I rs141853578 (G119R) with age-related macular degeneration in Iranian population. Int. Ophthalmol. 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.L.; Huang, X.F.; Wang, Q.F.; Cai, W.J.; Jin, Z.B.; Wang, Y. CFI-rs7356506 polymorphisms associated with Vogt-Koyanagi-Harada syndrome. Mol. Vision 2016, 22, 9–17. [Google Scholar]
- Jlajla, H.; Dehman, F.; Jallouli, M.; Khedher, R.; Ayadi, I.; Zerzeri, Y.; Laadhar, L.; Sfar, I.; Mahfoudh, A.; Gorgi, Y.; et al. Molecular basis of complement factor I deficiency in Tunisian atypical hemolytic and uremic syndrome Patients. Nephrology 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bi, Y.; Lin, J.; Cao, L.; He, B.; Zhang, Z.; Cui, Y.; Zhang, X. Tag SNPs of CFI contributed to the susceptibility for non-small cell lung cancer in Chinese population. Tumour Biol. 2015, 36, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.F.; Yang, M.M.; Cai, W.J.; Zheng, M.Q.; Mao, G.; Pang, C.P.; Jin, Z.B. CFI-rs7356506 is a genetic protective factor for acute anterior uveitis in Chinese patients. Br. J. Ophthalmol. 2014, 98, 1592–1596. [Google Scholar] [CrossRef] [PubMed]
- Cedrone, E.; Neun, B.W.; Rodriguez, J.; Vermilya, A.; Clogston, J.D.; McNeil, S.E.; Barenholz, Y.; Szebeni, J.; Dobrovolskaia, M.A. Anticoagulants Influence the Performance of In Vitro Assays Intended for Characterization of Nanotechnology-Based Formulations. Molecules 2017, 23, 12. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Assay Development and Validation for Immunogenicity Testing of Therapeutic Protein Products; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2016.
- Benasutti, H.; Wang, G.; Vu, V.P.; Scheinman, R.; Groman, E.; Saba, L.; Simberg, D. Variability of Complement Response toward Preclinical and Clinical Nanocarriers in the General Population. Bioconj. Chem. 2017, 28, 2747–2755. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the model nanoparticles Doxil and Doxebo as well as mouse mouse anti-PEG antibodies are available from a commercial supplier. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Antibody | Host | Type | Epitope Specificity |
|---|---|---|---|---|
| AB1 | AGP3 | Mouse | IgM | Backbone |
| AB2 | AGP4 | Mouse | IgM | Backbone |
| AB3 | E11 | Mouse | IgG1 | Backbone |
| AB4 | 3.3 | Mouse | IgG | Backbone |
| AB5 | 6.3 | Mouse | IgG1 | Backbone |
| AB6 | 15-2b | Mouse | IgG1 | -OCH3 |
| AB7 | ANPEG-1 | Mouse | IgM | Backbone |
| AB8 | PEG-B-47 | Rabbit | IgG | -OCH3 |
| AB9 | AGP6 | Rat | IgM | Backbone |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neun, B.W.; Barenholz, Y.; Szebeni, J.; Dobrovolskaia, M.A. Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro. Molecules 2018, 23, 1700. https://doi.org/10.3390/molecules23071700
Neun BW, Barenholz Y, Szebeni J, Dobrovolskaia MA. Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro. Molecules. 2018; 23(7):1700. https://doi.org/10.3390/molecules23071700
Chicago/Turabian StyleNeun, Barry W., Yechezkel Barenholz, Janos Szebeni, and Marina A. Dobrovolskaia. 2018. "Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro" Molecules 23, no. 7: 1700. https://doi.org/10.3390/molecules23071700
APA StyleNeun, B. W., Barenholz, Y., Szebeni, J., & Dobrovolskaia, M. A. (2018). Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro. Molecules, 23(7), 1700. https://doi.org/10.3390/molecules23071700