Identification of 2′,4′-Dihydroxychalcone as an Antivirulence Agent Targeting HlyU, a Master Virulence Regulator in Vibrio vulnificus



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
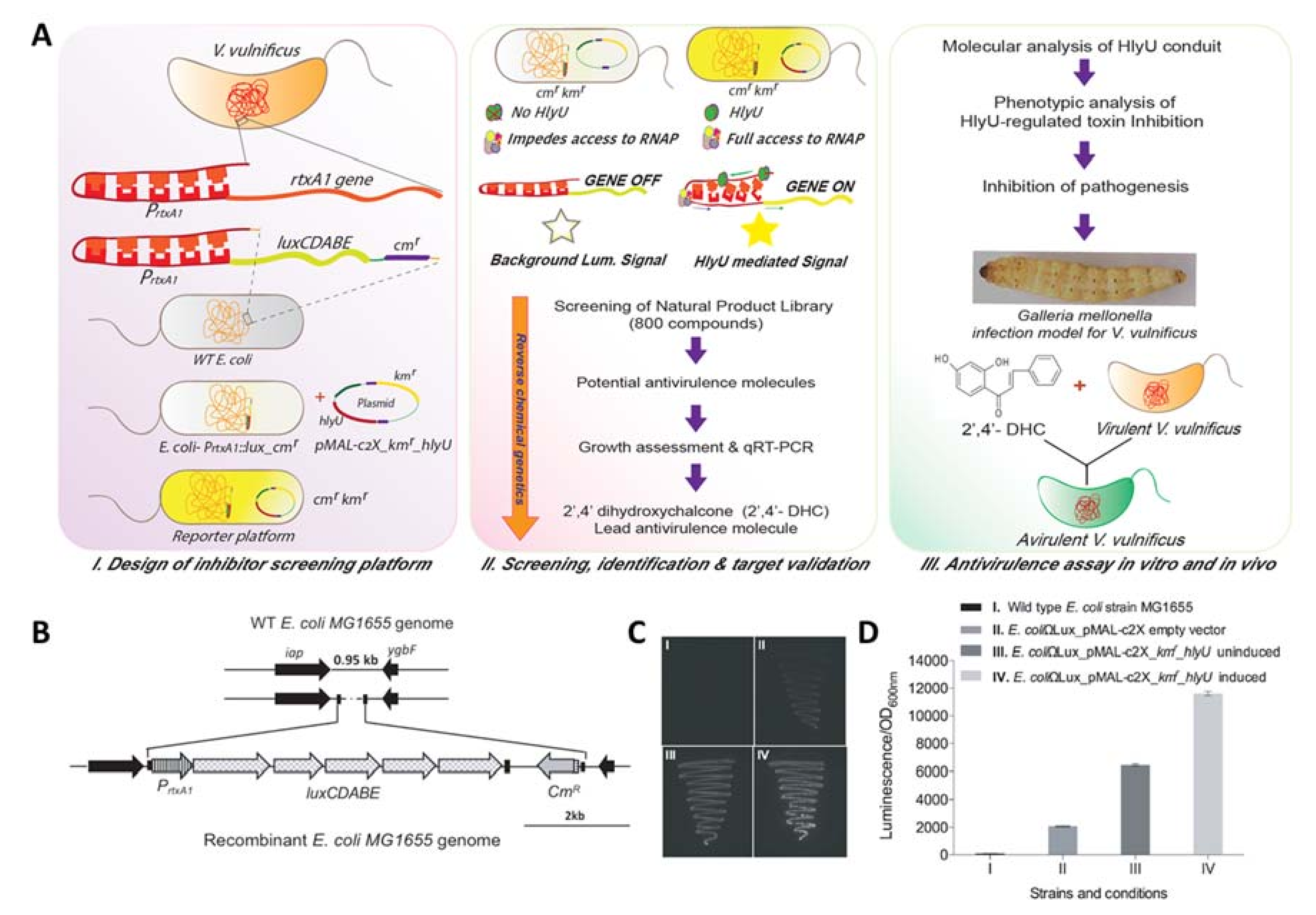
2.1. Development of Orthogonal Reporter Platform
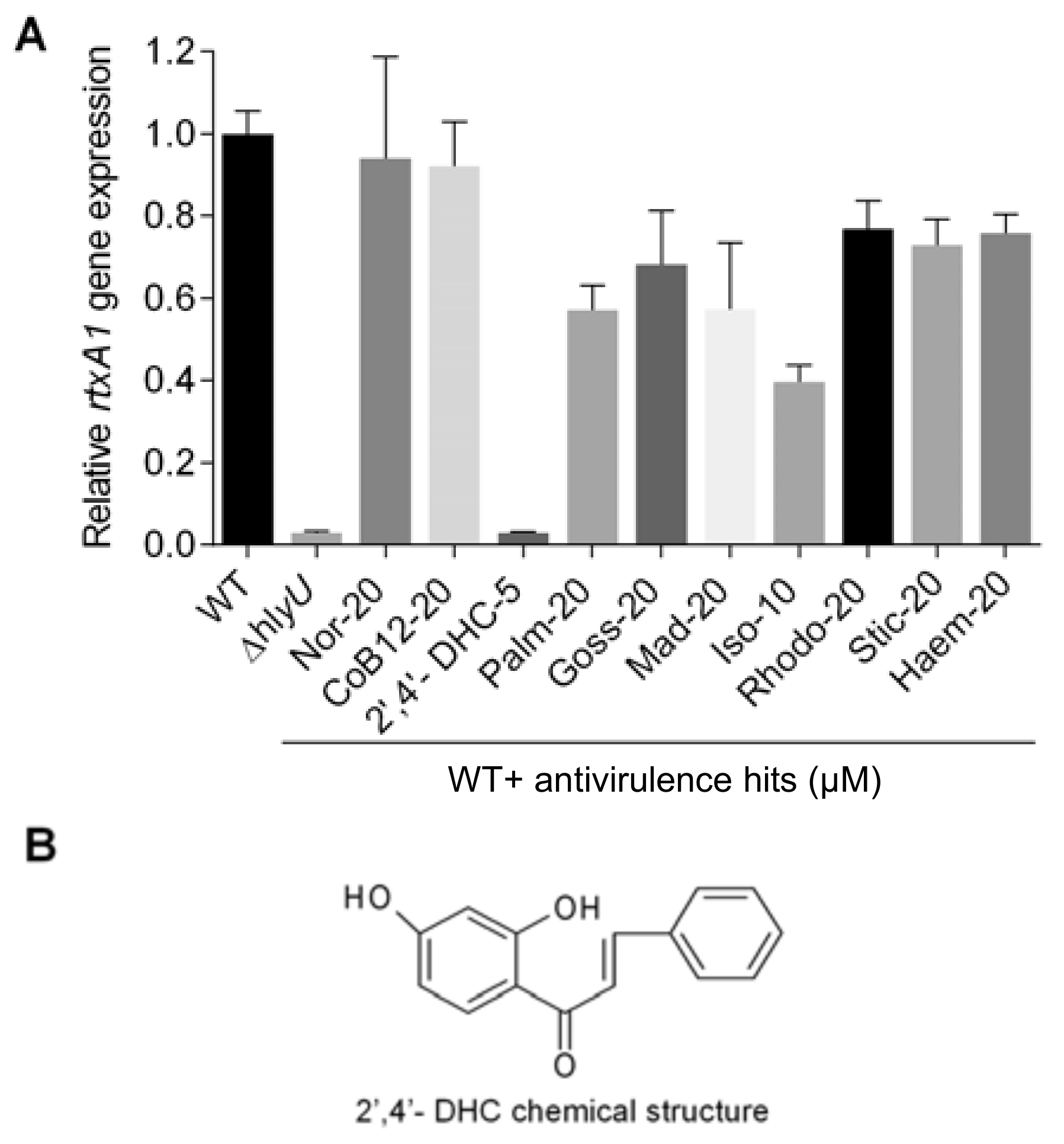
2.2. Screening, Identification, and Assessment of Potent Antivirulence Inhibitors from the Natural Product Library
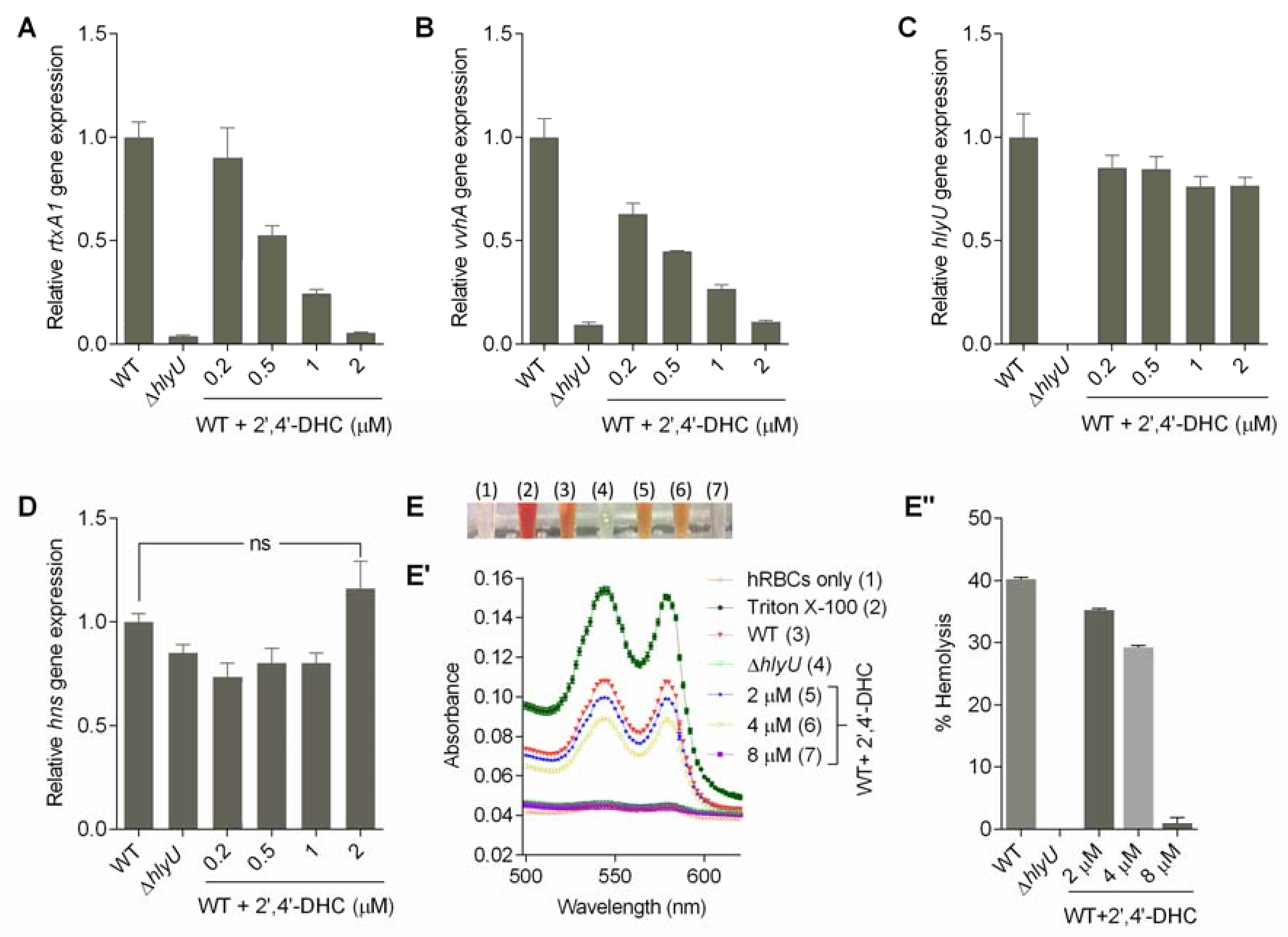
2.3. Toxicity of 2′,4′-DHC on the Mammalian Host Cell and V. vulnificus Growth
2.4. Assessment of Antivirulence Activity Using qRT-PCR and Phenotype
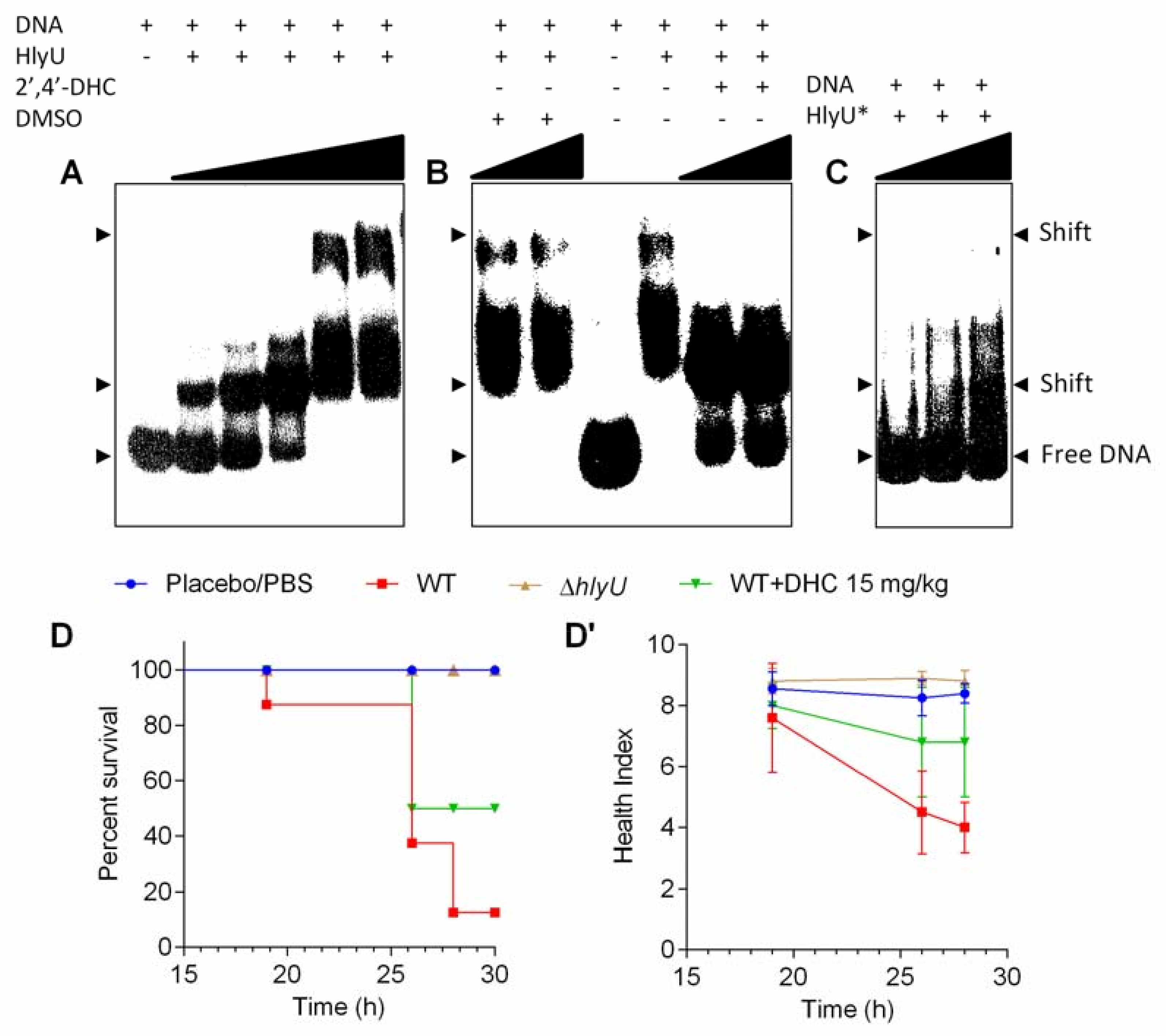
2.5. Mechanism of Action of 2′,4′-DHC and Its In Vivo Validation
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Development of Orthogonal Reporter Platform
4.3. Screening of the Natural Product Library and Antivirulence Lead Compound Identification
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. Host and Pathogen Cytotoxicity by Small Molecule 2′,4′-DHC
4.6. Hemolysis Assay
4.7. Cloning, Expression, and Purification of WT and Dimerization Mutant (L91A/L17A) of V. vulnificus HlyU
4.8. Electrophoretic Mobility Shift Assay (EMSA)
4.9. Galleria mellonella Infection Model for V. vulnificus
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Daniels, N.A. Vibrio vulnificus oysters: Pearls and perils. Clin. Infect. Dis. 2011, 52, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Heng, S.P.; Letchumanan, V.; Deng, C.Y.; Ab Mutalib, N.S.; Khan, T.M.; Chuah, L.H.; Chan, K.G.; Goh, B.H.; Pusparajah, P.; Lee, L.H. Vibrio vulnificus: An environmental and clinical burden. Front. Microbiol. 2017, 8, 997. [Google Scholar] [CrossRef] [PubMed]
- Horseman, M.A.; Surani, S. A comprehensive review of Vibrio vulnificus: An important cause of severe sepsis and skin and soft-tissue infection. Int. J. Infect. Dis. 2011, 15, e157–e166. [Google Scholar] [CrossRef] [PubMed]
- Bross, M.H.; Soch, K.; Morales, R.; Mitchell, R.B. Vibrio vulnificus infection: Diagnosis and treatment. Am. Fam. Phys. 2007, 76, 539–544. [Google Scholar]
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.R.; Chuang, Y.C. Vibrio vulnificus infection: Clinical manifestations, pathogenesis, and antimicrobial therapy. J. Microbiol. Immunol. Infect. 2003, 36, 81–88. [Google Scholar] [PubMed]
- Chen, S.C.; Lee, Y.T.; Tsai, S.J.; Chan, K.S.; Chao, W.N.; Wang, P.H.; Lin, D.B.; Chen, C.C.; Lee, M.C. Antibiotic therapy for necrotizing fasciitis caused by Vibrio vulnificus: Retrospective analysis of an 8 year period. J. Antimicrob. Chemother. 2012, 67, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.C.; Brown, A.M.; Luscombe, G.M.; Wong, S.J.; Mendis, K. Antibiotic use for Vibrio infections: Important insights from surveillance data. BMC Infect. Dis. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; McArthur, J.V.; Lindell, A.H.; Wright, M.S.; Tuckfield, R.C.; Gooch, J.; Warner, L.; Oliver, J.; Stepanauskas, R. Multi-site analysis reveals widespread antibiotic resistance in the marine pathogen Vibrio vulnificus. Microb. Ecol. 2009, 57, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Imdad, S.; Chaurasia, A.K.; Kim, K.K. Identification and validation of an antivirulence agent targeting HlyU-regulated virulence in Vibrio vulnificus. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.G.; Satchell, K.J. Additive function of Vibrio vulnificus MATRX(Vv) and VvhA cytolysins promotes rapid growth and epithelial tissue necrosis during intestinal infection. PLoS Pathog. 2012, 8, e1002581. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.R.; Lin, J.H.; Chen, Y.H.; Chen, C.L.; Shao, C.P.; Lai, Y.C.; Hor, L.I. Rtx toxin enhances the survival of Vibrio vulnificus during infection by protecting the organism from phagocytosis. J. Infect. Dis. 2011, 203, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, M.W.; Kim, B.S.; Kim, S.M.; Lee, B.C.; Kim, T.S.; Choi, S.H. Identification and characterization of the Vibrio vulnificus RtxA essential for cytotoxicity in vitro and virulence in mice. J. Microbiol. 2007, 45, 146–152. [Google Scholar] [PubMed]
- Liu, M.; Crosa, J.H. The regulator HlyU, the repeat-in-toxin gene rtxA1, and their roles in the pathogenesis of Vibrio vulnificus infections. Microbiologyopen 2012, 1, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Naka, H.; Crosa, J.H. HlyU acts as an H-NS antirepressor in the regulation of the RTX toxin gene essential for the virulence of the human pathogen Vibrio vulnificus CMCP6. Mol. Microbiol. 2009, 72, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Lee, S.E.; Kook, H.; Yeom, J.A.; Na, H.S.; Kim, S.Y.; Chung, S.S.; Choy, H.E.; Rhee, J.H. Vibrio vulnificus RTX toxin kills host cells only after contact of the bacteria with host cells. Cell. Microbiol. 2008, 10, 848–862. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Cha, M.H.; Oh, D.R.; Oh, W.K.; Rhee, J.H.; Kim, Y.R. Resveratrol modulates RTX toxin-induced cytotoxicity through interference in adhesion and toxin production. Eur. J. Pharmacol. 2010, 642, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Na, H.S.; Cha, M.H.; Oh, D.R.; Cho, C.W.; Rhee, J.H.; Kim, Y.R. Protective mechanism of curcumin against Vibrio vulnificus infection. FEMS Immunol. Med. Microbiol. 2011, 63, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Joray, M.B.; Trucco, L.D.; Gonzalez, M.L.; Napal, G.N.; Palacios, S.M.; Bocco, J.L.; Carpinella, M.C. Antibacterial and cytotoxic activity of compounds isolated from Flourensia oolepis. Evid. Based Complement. Alternat. Med. 2015, 2015, 912484. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Rose, M.; Crosa, J.H. Homodimerization and binding of specific domains to the target DNA are essential requirements for hlyU to regulate expression of the virulence gene rtxA1, encoding the repeat-in-toxin protein in the human pathogen Vibrio vulnificus. J. Bacteriol. 2011, 193, 6895–6901. [Google Scholar] [CrossRef] [PubMed]
- Food Safety. 2017. Available online: http://www.who.int/en/news-room/fact-sheets/detail/food-safety (accessed on 9 May 2018).
- Song, E.J.; Lee, S.J.; Lim, H.S.; Kim, J.S.; Jang, K.K.; Choi, S.H.; Han, H.J. Vibrio vulnificus VvhA induces autophagy-related cell death through the lipid raft-dependent c-Src/Nox signaling pathway. Sci. Rep. 2016, 6, 27080. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Jung, Y.H.; Oh, S.Y.; Song, E.J.; Choi, S.H.; Han, H.J. Vibrio vulnificus vvhA induces NF-ĸB-dependent mitochondrial cell death via lipid raft-mediated ROS production in intestinal epithelial cells. Cell Death Dis. 2015, 6, 1655. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.C.; Simpson, L.M.; Oliver, J.D.; Morris, J.G., Jr. Phenotypic evaluation of acapsular transposon mutants of Vibrio vulnificus. Infect. Immun. 1990, 58, 1769–1773. [Google Scholar] [PubMed]
- Jang, K.K.; Lee, Z.W.; Kim, B.; Jung, Y.H.; Han, H.J.; Kim, M.H.; Kim, B.S.; Choi, S.H. Identification and characterization of Vibrio vulnificus PlpA encoding a phospholipase A2 essential for pathogenesis. J. Biol. Chem. 2017, 292, 17129–17143. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, A.K.; Thorat, N.D.; Tandon, A.; Kim, J.H.; Park, S.H.; Kim, K.K. Coupling of radiofrequency with magnetic nanoparticles treatment as an alternative physical antibacterial strategy against multiple drug resistant bacteria. Sci. Rep. 2016, 6, 33662. [Google Scholar] [CrossRef] [PubMed]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Walker, I.H.; Hsieh, P.C.; Riggs, P.D. Mutations in maltose-binding protein that alter affinity and solubility properties. Appl. Microbiol. Biotechnol. 2010, 88, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(t) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, J.A.; Lee, M.A.; Park, S.J.; Lee, K.H. Regulation of haemolysin (vvhA) production by ferric uptake regulator (Fur) in Vibrio vulnificus: Repression of vvhA transcription by Fur and proteolysis of vvhA by Fur-repressive exoproteases. Mol. Microbiol. 2013, 88, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Carter, P. Site-directed mutagenesis. Biochem. J. 1986, 237, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nishi, K.; Lee, H.J.; Park, S.Y.; Bae, S.J.; Lee, S.E.; Adams, P.D.; Rhee, J.H.; Kim, J.S. Crystal structure of the transcriptional activator HlyU from Vibrio vulnificus CMCP6. FEBS Lett. 2010, 584, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Coote, P.J. Wax moth larva (Galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother. 2011, 66, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.; Veli, N.; Coote, P.J. Evaluation of Galleria mellonella larvae for measuring the efficacy and pharmacokinetics of antibiotic therapies against Pseudomonas aeruginosa infection. Int. J. Antimicrob. Agents 2014, 43, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Loh, J.M.; Adenwalla, N.; Wiles, S.; Proft, T. Galleria mellonella larvae as an infection model for group A Streptococcus. Virulence 2013, 4, 419–428. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds ...... are available from the authors. |




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imdad, S.; Batool, N.; Pradhan, S.; Chaurasia, A.K.; Kim, K.K. Identification of 2′,4′-Dihydroxychalcone as an Antivirulence Agent Targeting HlyU, a Master Virulence Regulator in Vibrio vulnificus. Molecules 2018, 23, 1492. https://doi.org/10.3390/molecules23061492
Imdad S, Batool N, Pradhan S, Chaurasia AK, Kim KK. Identification of 2′,4′-Dihydroxychalcone as an Antivirulence Agent Targeting HlyU, a Master Virulence Regulator in Vibrio vulnificus. Molecules. 2018; 23(6):1492. https://doi.org/10.3390/molecules23061492
Chicago/Turabian StyleImdad, Saba, Nayab Batool, Subhra Pradhan, Akhilesh Kumar Chaurasia, and Kyeong Kyu Kim. 2018. "Identification of 2′,4′-Dihydroxychalcone as an Antivirulence Agent Targeting HlyU, a Master Virulence Regulator in Vibrio vulnificus" Molecules 23, no. 6: 1492. https://doi.org/10.3390/molecules23061492
APA StyleImdad, S., Batool, N., Pradhan, S., Chaurasia, A. K., & Kim, K. K. (2018). Identification of 2′,4′-Dihydroxychalcone as an Antivirulence Agent Targeting HlyU, a Master Virulence Regulator in Vibrio vulnificus. Molecules, 23(6), 1492. https://doi.org/10.3390/molecules23061492