4′-Methoxyresveratrol Alleviated AGE-Induced Inflammation via RAGE-Mediated NF-κB and NLRP3 Inflammasome Pathway


Abstract
:1. Introduction
2. Results
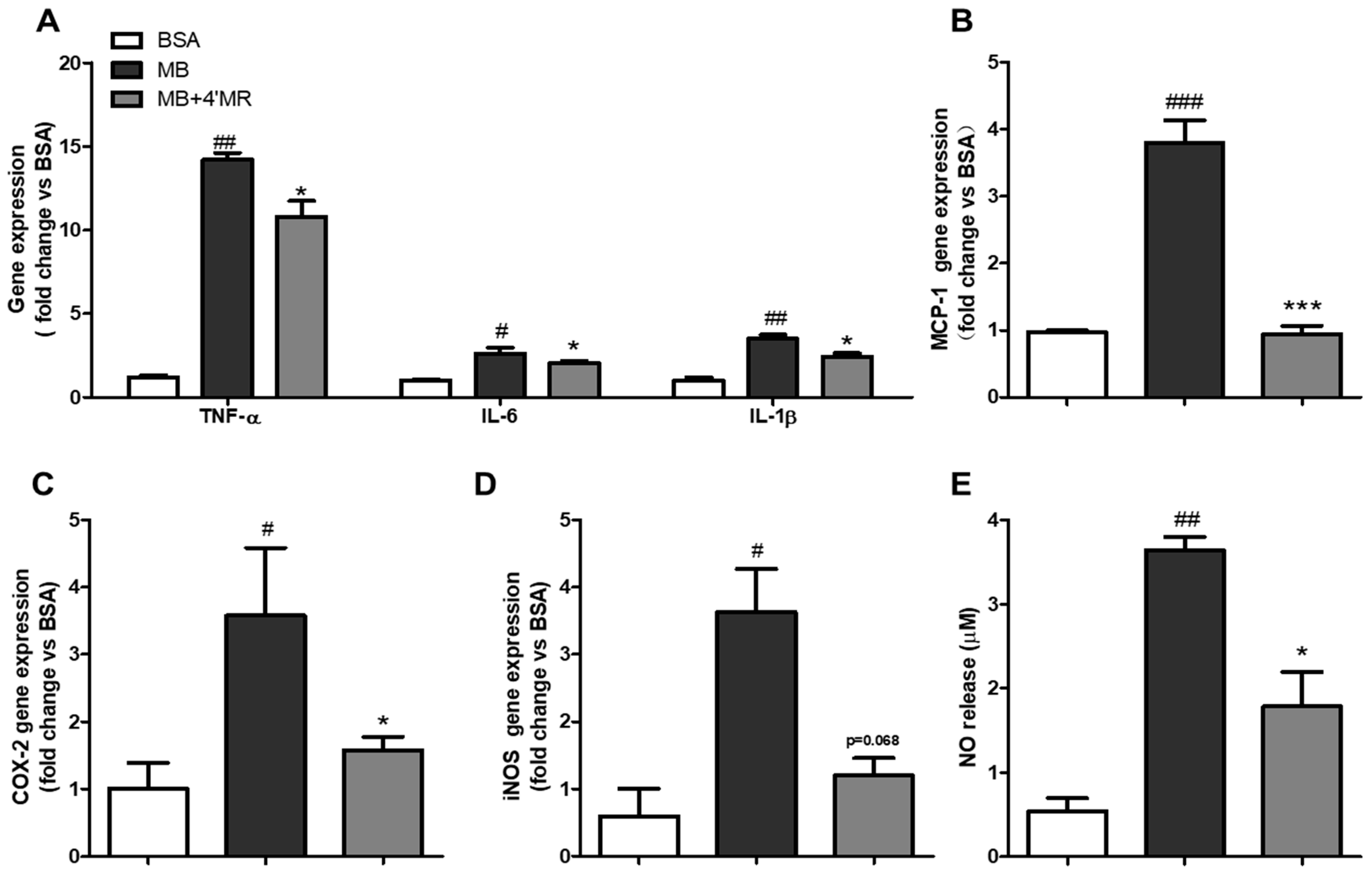
2.1. 4′-Methoxyresveratrol Modulated Pro-Inflammatory Markers Expression Induced by AGEs
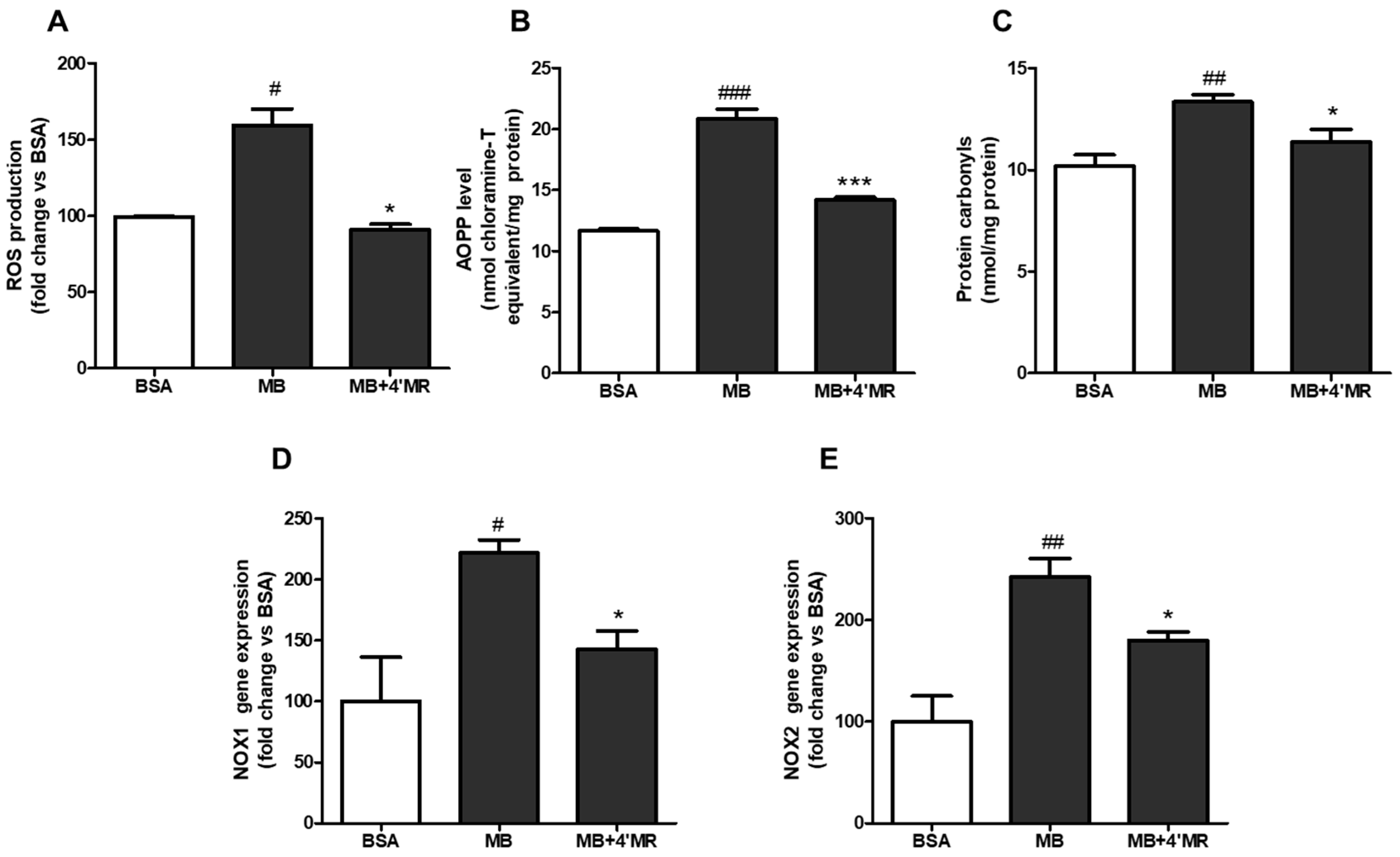
2.2. 4′-Methoxyresveratrol Alleviated Oxidative Stress Induced by AGEs
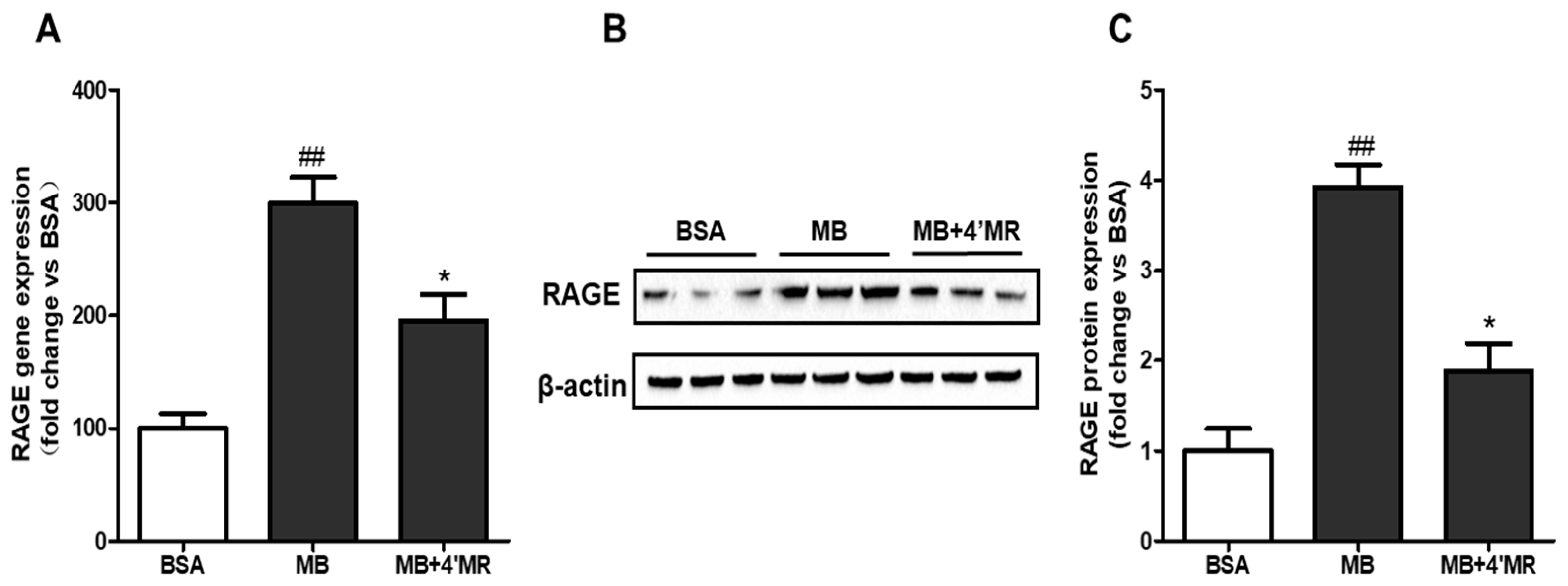
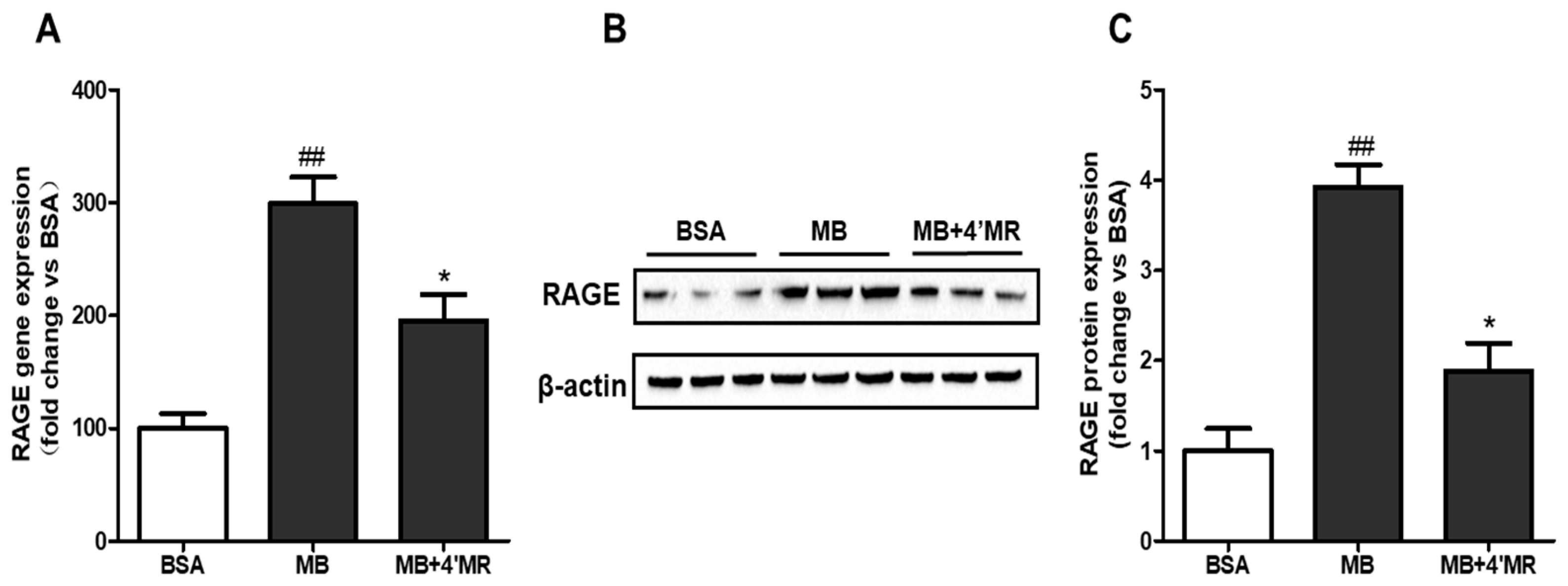
2.3. 4′-Methoxyresveratrol Suppressed mRNA and Protein Level of RAGE
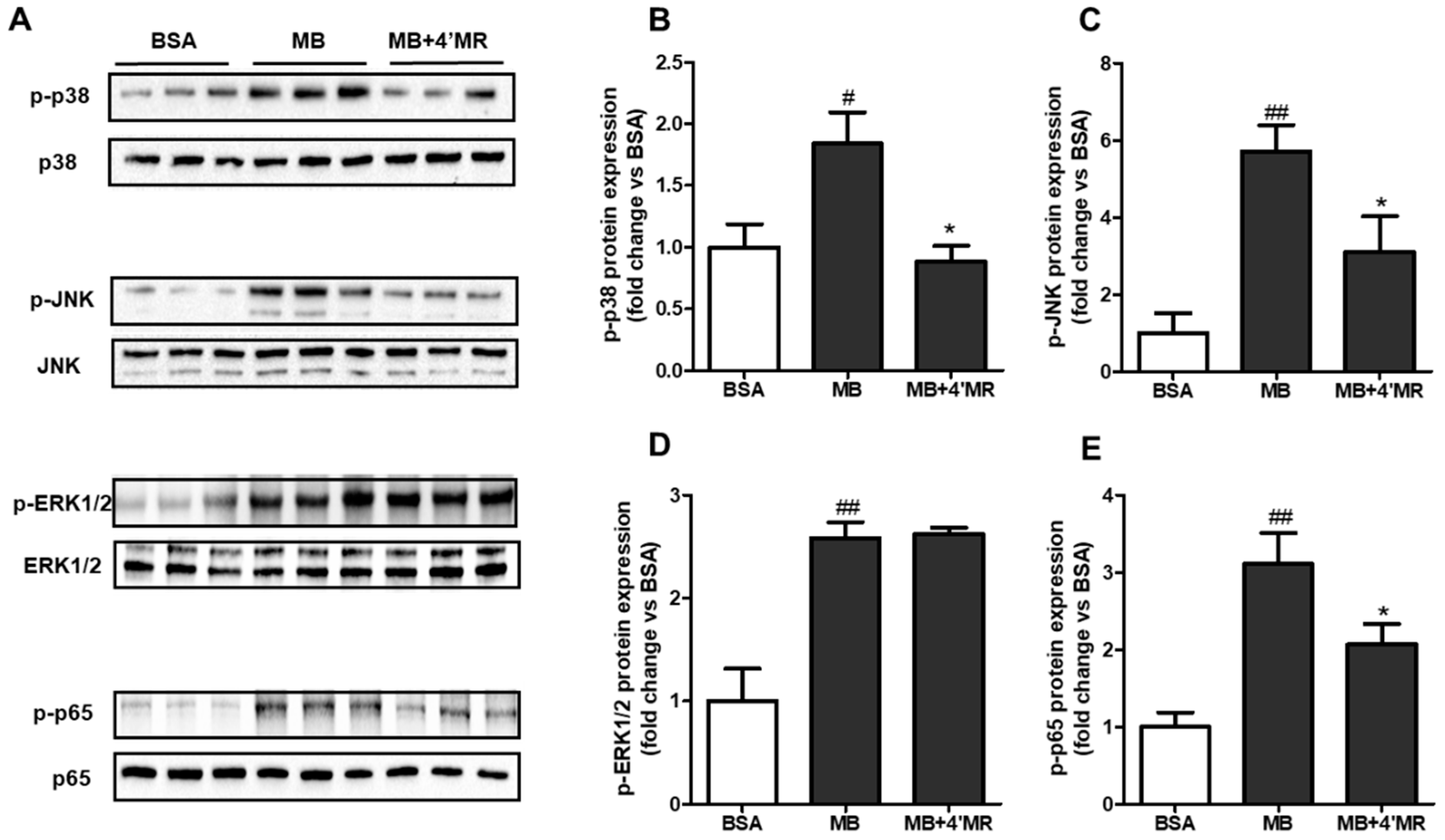
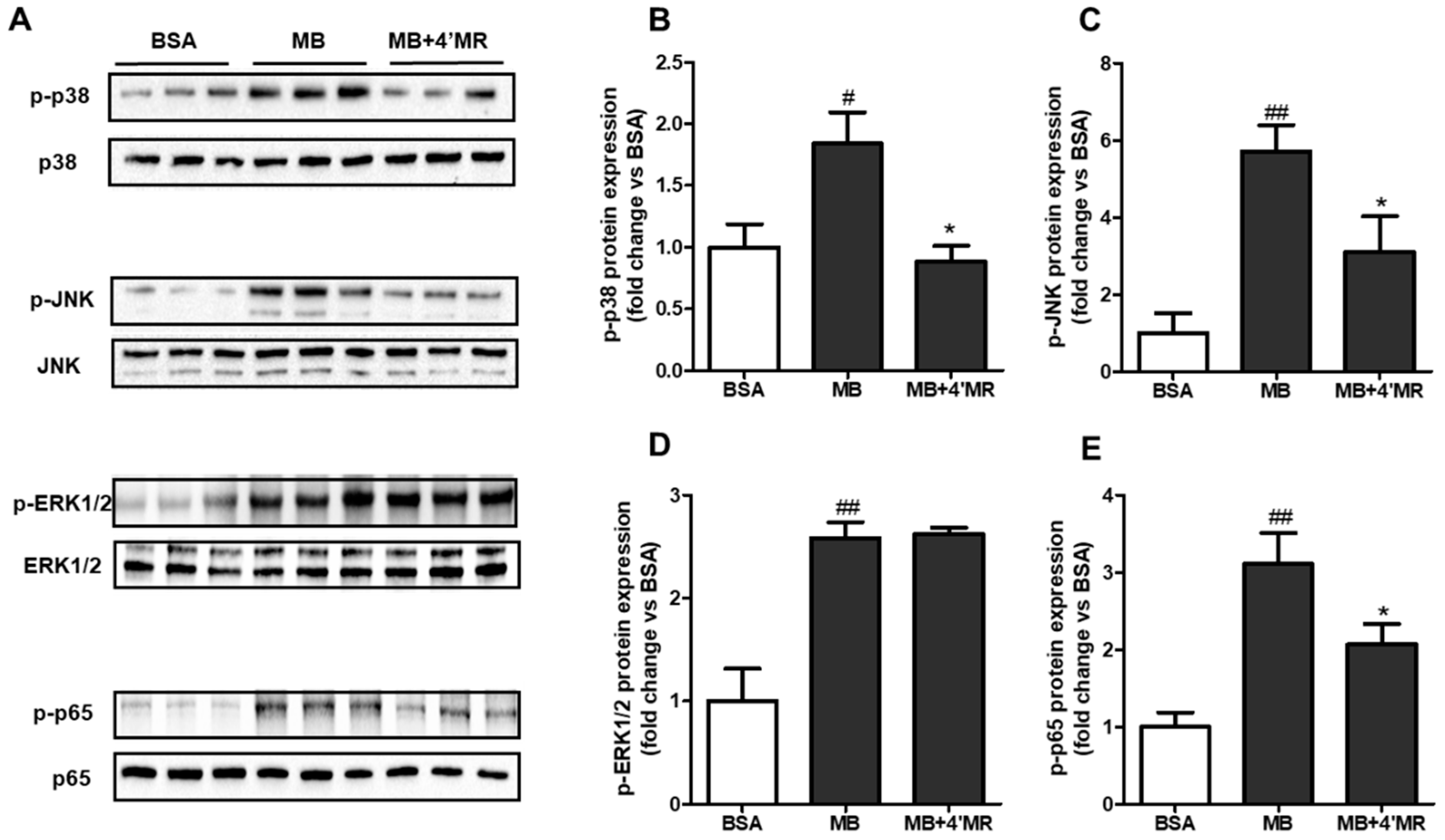
2.4. 4′-Methoxyresveratrol Modified MAPKs/NF-κB Pathway Induced by AGEs
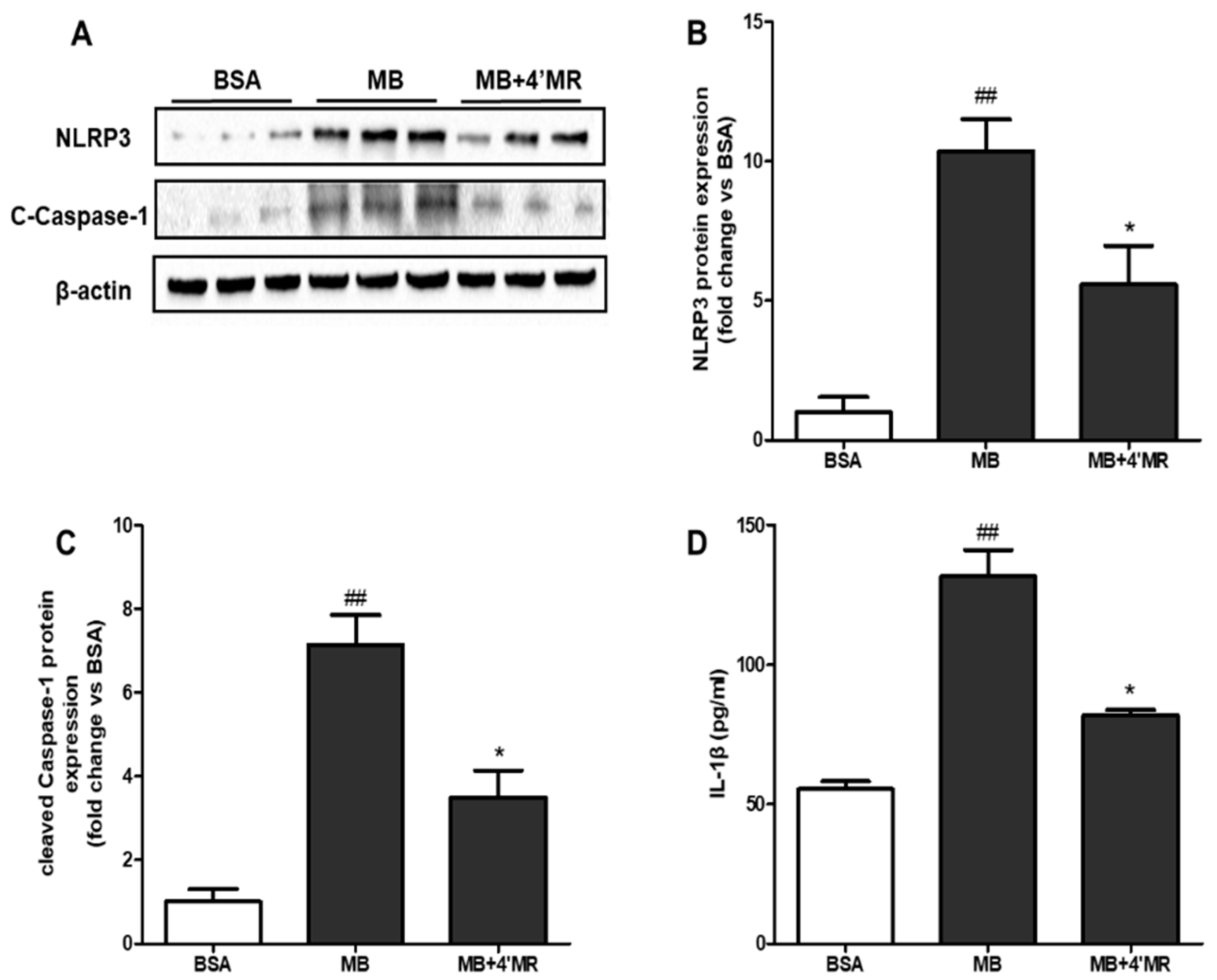
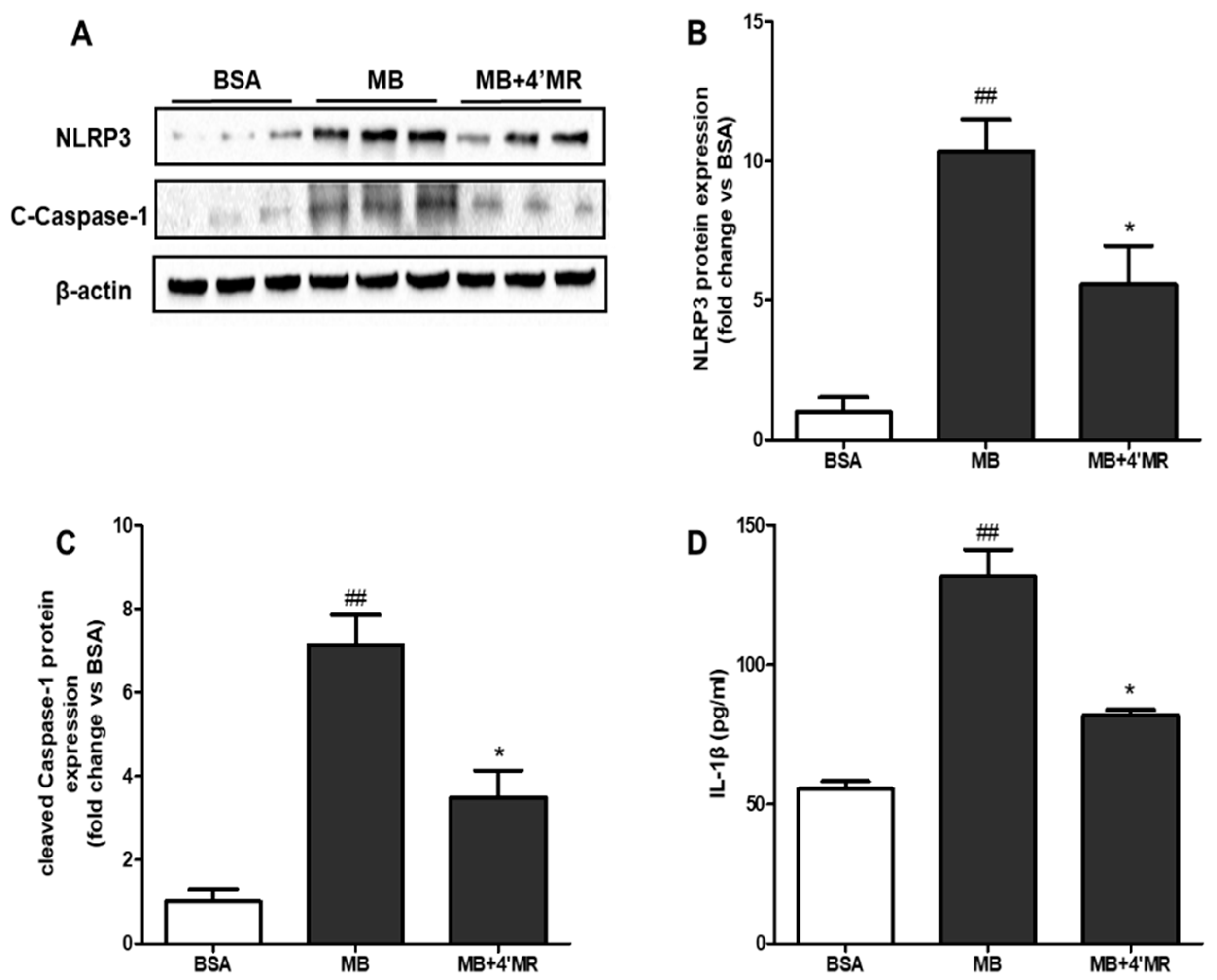
2.5. 4′-Methoxyresveratrol Counteracted the Activation of NLRP3 Inflammasome Induced by AGEs
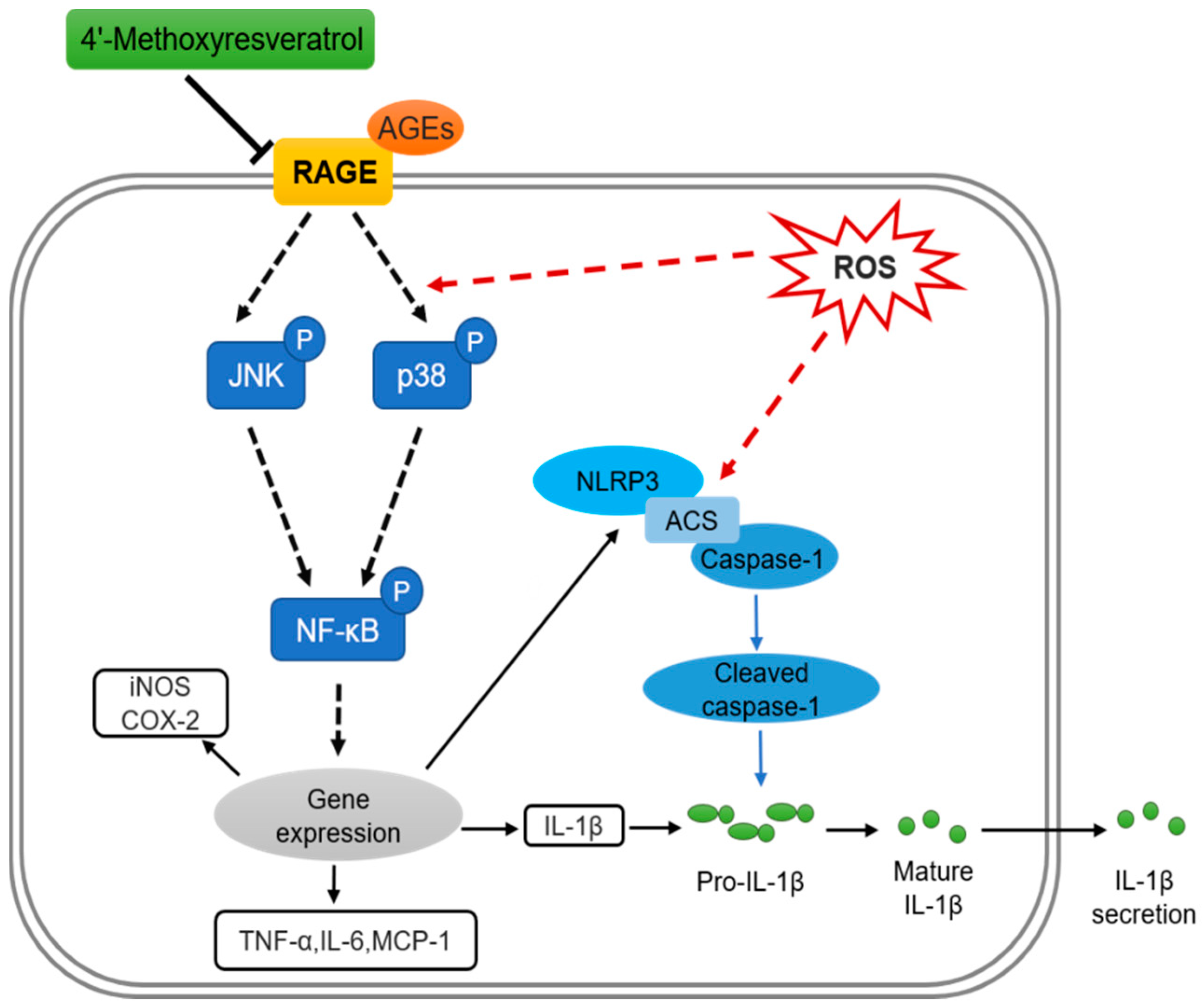
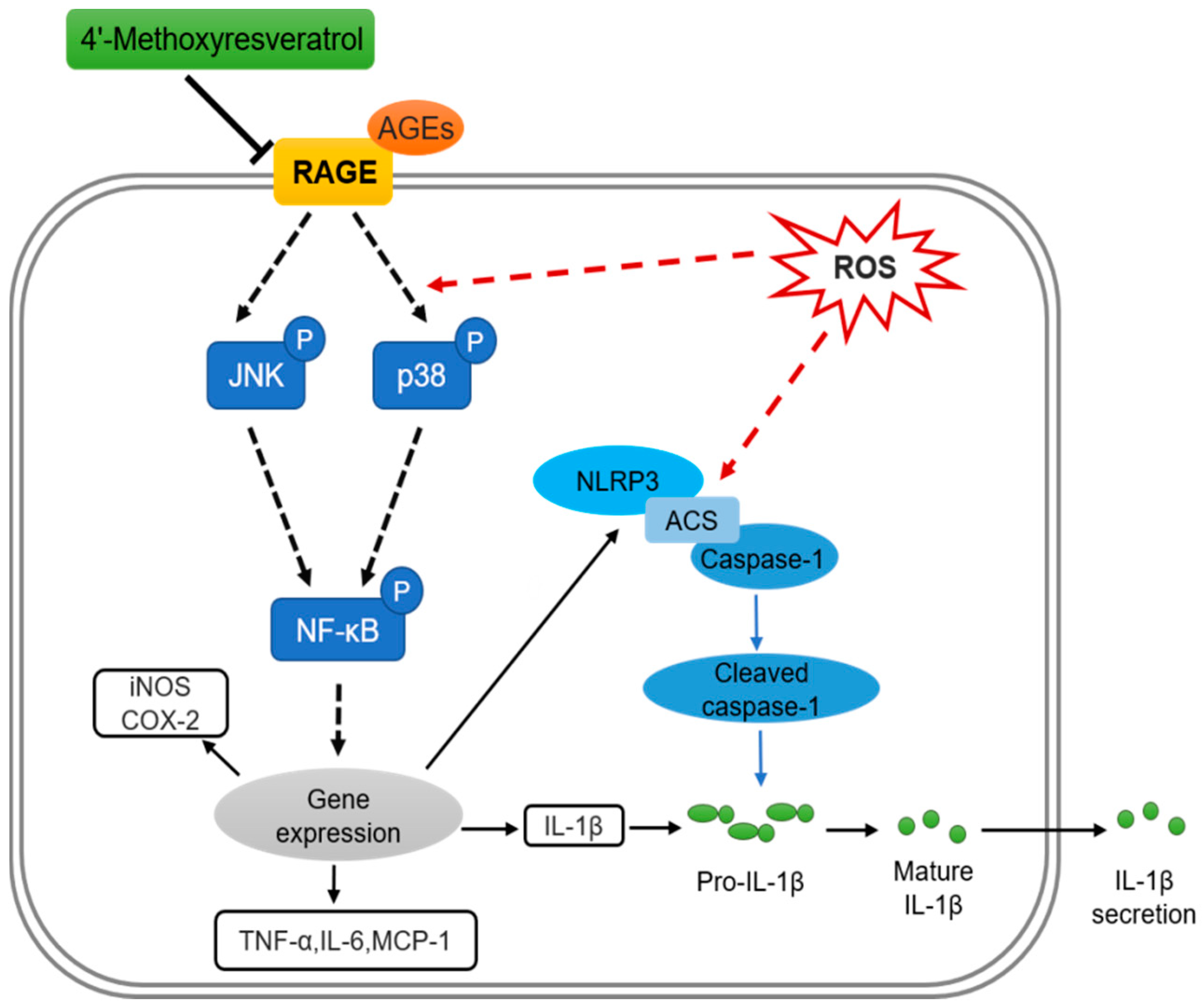
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture
4.3. Preparation of Methylglyoxal-Modified AGEs
4.4. Intracellular ROS Production Measurement
4.5. Protein Carbonyl and AOPP Levels Measurement
4.6. RNA Isolation and qPCR Analysis
4.7. Measurement of NO and IL-1β Levels
4.8. Protein Extraction and Western Blot Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AGEs | advanced glycation end products |
| AOPP | advanced oxidation protein product |
| ASC | apoptosis-associated speck-like protein containing CARD |
| BSA | bovine Serum Albumin |
| Caspase-1 | cysteinyl aspartate specific proteinase 1 |
| C-Caspase-1 | cleaved cysteinyl aspartate specific proteinase 1 |
| COX-2 | cyclooxygenase 2 |
| DCFH-DA | 2’,7’-dichlorodihydrofluorescein diacetate |
| IL-6 | interleukin 6 |
| IL-1β | interleukin 1β |
| iNOS | inducible nitric oxide synthase |
| LPS | lipopolysaccharide |
| MAPK | mitogen activated protein kinase |
| MB | methylglyoxal-bovine serum albumin |
| MCP-1 | monocyte chemoattractant protein 1 |
| 4′MR | 4′-methoxyresveratrol; MGO, methylglyoxal |
| NF-κB | nuclear transcription factor kappa B |
| NLRP3 | nucleotide-binding oligomerization domain (NOD)-like receptor (NLR) pyrin domain containing 3 |
| NO | nitric oxide |
| NOX1/2 | nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 1/2 |
| RAGE | receptor for advanced glycation end products; ROS, reactive oxygen species |
| TNF-α | tumor necrosis factor alpha |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| COX2 | CTCACGAAGGAACTCAGCACT | TAGAATCCAGTCCGGGTACAGT |
| IL-1β | GCAACTGTTCCTGAACTCAACT | ATCTTTTGGGGTCCGTCAACT |
| IL-6 | AGCCAGAGTCCTTCAGAGAGAT | GCACTAGGTTTGCCGAGTAGAT |
| iNOS | GGCAGCCTGTGAGACCTTTG | GCATTGGAAGTGAAGCGTTTC |
| MCP-1 | AGCTCTTTCCTCCACCA | CTACAGCTTCTTTGGGACACCT |
| NOX1 | AATGCCCAGGATCGAGGT | GATGGAAGCAAAGGGAGTGA |
| NOX2 | CAGGAACCTCACTTTCCATAAGAT | AACGTTGAAGAGATGTGCAATTGT |
| RAGE | AACACAGCCCCCATCCAA | GCTCAACCAACAGCTGAATGC |
| TNF-α | CACCACGCTCTTCTGTCTACTG | CTTTGAGATCCATCGCGTTG |
| 18S | GTAACCCGTTGAACCCCATT | CCATCCAATCGGTAGTAGCG |
References
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bugel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef] [PubMed]
- Luevano-Contreras, C.; Chapman-Novakofski, K. Dietary advanced glycation end products and aging. Nutrients 2010, 2, 1247–1265. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-A.; Wu, C.-H.; Lu, C.-C.; Hsia, S.-M.; Yen, G.-C. Glycative stress from advanced glycation end products (AGEs) and dicarbonyls: An emerging biological factor in cancer onset and progression. Mol. Nutr. Food Res. 2016, 60, 1850–1864. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Iwata, D.; Kitaichi, N.; Takeuchi, M.; Sato, M.; Endo, N.; Iwabuchi, K.; Ando, R.; Fukuhara, J.; Kinoshita, S.; et al. Amelioration of experimental autoimmune uveoretinitis by inhibition of glyceraldehyde-derived advanced glycation end-product formation. J. Leukoc. Biol. 2014, 96, 1077–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wautier, M.P.; Guillausseau, P.J.; Wautier, J.L. Activation of the receptor for advanced glycation end products and consequences on health. Diabetes Metab. Syndr. 2017, 11, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Treuter, E.; Fan, R.; Huang, Z.; Jakobsson, T.; Venteclef, N. Transcriptional repression in macrophages-basic mechanisms and alterations in metabolic inflammatory diseases. FEBS Lett. 2017, 591, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zheng, Y.; Wang, Y.; Xiong, W.; Lin, L. Advanced glycation end products interfere with gastric smooth muscle contractile marker expression via the AGE/RAGE/NF-kappaB pathway. Exp. Mol. Pathol. 2017, 102, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Abderrazak, A.; Syrovets, T.; Couchie, D.; El Hadri, K.; Friguet, B.; Simmet, T.; Rouis, M. NLRP3 inflammasome: From a danger signal sensor to a regulatory node of oxidative stress and inflammatory diseases. Redox Biol. 2015, 4, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Huang, W.; Peng, J.; Zhu, T.T.; Sun, X.L.; Zhou, X.Y.; Yang, H.; Xiong, J.F.; He, H.Q.; Xu, Y.H.; et al. Irisin alleviates advanced glycation end products-induced inflammation and endothelial dysfunction via inhibiting ROS-NLRP3 inflammasome signaling. Inflammation 2018, 41, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, Y.; Zhang, Y.; Geng, W.; Liu, W.; Gao, Y.; Li, S.; Wang, K.; Wu, X.; Kang, L.; et al. Advanced glycation end products regulate anabolic and catabolic activities via NLRP3-inflammasome activation in human nucleus pulposus cells. J. Cell. Mol. Med. 2017, 21, 1373–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial action of resveratrol: How and why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, K.; Toyama, T.; Ito, T.; Shakui, T.; Usui, S.; Oyama, M.; Iinuma, M.; Hirano, K. Antiandrogenic activity of resveratrol analogs in prostate cancer LNCaP cells. J. Androl. 2012, 33, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Houille, B.; Papon, N.; Boudesocque, L.; Bourdeaud, E.; Besseau, S.; Courdavault, V.; Enguehard-Gueiffier, C.; Delanoue, G.; Guerin, L.; Bouchara, J.P.; et al. Antifungal activity of resveratrol derivatives against Candida species. J. Nat. Prod. 2014, 77, 1658–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csont, T.; Viappiani, S.; Sawicka, J.; Slee, S.; Altarejos, J.Y.; Batinić-Haberle, I.; Schulz, R. The involvement of superoxide and iNOS-derived NO in cardiac dysfunction induced by pro-inflammatory cytokines. J. Mol. Cell. Cardiol. 2005, 39, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanaja, S.K.; Rathinam, V.A.; Fitzgerald, K.A. Mechanisms of inflammasome activation: Recent advances and novel insights. Trends Cell Biol. 2015, 25, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Passini, F.S.; Silvan, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Schoenhuber, H.; Redaelli, A.; Berli, M.; et al. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol. 2017, 59, 95–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byun, K.; Yoo, Y.; Son, M.; Lee, J.; Jeong, G.B.; Park, Y.M.; Salekdeh, G.H.; Lee, B. Advanced glycation end-products produced systemically and by macrophages: A common contributor to inflammation and degenerative diseases. Pharmacol. Ther. 2017, 177, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Hu, X.; Wang, M. Pterostilbene inhibited advanced glycation end products (AGEs)-induced oxidative stress and inflammation by regulation of RAGE/MAPK/NF-κB in RAW264.7 cells. J. Funct. Foods 2018, 40, 272–279. [Google Scholar] [CrossRef]
- Nedic, O.; Rattan, S.I.; Grune, T.; Trougakos, I.P. Molecular effects of advanced glycation end products on cell signalling pathways, ageing and pathophysiology. Free Radic. Res. 2013, 47, 28–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaens, K.; Stehouwer, C.D.A.; Schalkwijk, C.G. The Nε-(carboxymethyl)lysine–RAGE axis: Putative implications for the pathogenesis of obesity-related complications. Expert Rev. Endocrinol. Metab. 2010, 5, 839–854. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.; Zhang, H.; Li, H.; Liu, J.; Zhang, F.; Jiang, T.; Jiang, S. Curcumin prevents osteoarthritis by inhibiting the activation of inflammasome NLRP3. J. Interferon Cytokine Res. 2017, 37, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, A.; Shibutani, Y.; Seno, K.; Iwata, H.; Kuwayama, T.; Shirasuna, K. Advanced glycation end products and lipopolysaccharides stimulate interleukin-6 secretion via the RAGE/TLR4-NF-kappaB-ROS pathways and resveratrol attenuates these inflammatory responses in mouse macrophages. Exp. Ther. Med. 2017, 14, 4363–4370. [Google Scholar] [PubMed]
- Fukuhara, K.; Nakanishi, I.; Matsuoka, A.; Matsumura, T.; Honda, S.; Hayashi, M.; Ozawa, T.; Miyata, N.; Saito, S.; Ikota, N.; et al. Effect of methyl substitution on the antioxidative property and genotoxicity of resveratrol. Chem. Res. Toxicol. 2008, 21, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, K.I.; Guha, S.N.; Rao, M.N. Physico-chemical properties and antioxidant activities of methoxy phenols. Free Radic. Biol. Med. 1998, 24, 933–941. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Kreiner, L.H.; Johnson, N.M.; Brown, L.A.; Helms, M.N. Receptor for advanced glycation end-products regulates lung fluid balance via protein kinase C-gp91(phox) signaling to epithelial sodium channels. Am. J. Respir. Cell Mol. Biol. 2015, 52, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Mendez, J.D.; Mendez-Valenzuela, V.; Aguilar-Hernandez, M.M. Cellular signalling of the receptor for advanced glycation end products (RAGE). Cell Signal. 2013, 25, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.H.; Sturgis, L.; Haidacher, J.; Zhang, X.N.; Sherwood, S.J.; Bjercke, R.J.; Juhasz, O.; Crow, M.T.; Tilton, R.G.; Denner, L. Requirement for p38 and p44/p42 mitogen-activated protein kinases in RAGE-mediated nuclear factor-kappaB transcriptional activation and cytokine secretion. Diabetes 2001, 50, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.-C.; Martinez, K.; Xie, G.; Kennedy, A.; Bumrungpert, A.; Overman, A.; Jia, W.; McIntosh, M.K. Quercetin is equally or more effective than resveratrol in attenuating tumor necrosis factor-{alpha}-mediated inflammation and insulin resistance in primary human adipocytes. Am. J. Clin. Nutr. 2010, 92, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Xu, W.; Zhang, H.; Hua, X.; Zheng, D.; Guo, R.; Shen, N.; Hu, F.; Feng, J.; Liu, D. Inhibition of ROS-activated p38MAPK pathway is involved in the protective effect of H2S against chemical hypoxia-induced inflammation in PC12 cells. Neurochem. Res. 2013, 38, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Kamata, H.; Honda, S.-I.; Maeda, S.; Chang, L.; Hirata, H.; Karin, M. Reactive oxygen species promote TNFalpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 2005, 120, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Kong, H.; Pan, Y.B.; Jiang, L.; Pan, X.X.; Hu, L.; Qian, Y.N.; Jiang, C.Y.; Liu, W.T. Procyanidins alleviates morphine tolerance by inhibiting activation of NLRP3 inflammasome in microglia. J. Neuroinflamm. 2016, 13. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, W.; Tao, M.; Zhao, Y.; Hu, X.; Wang, M. 4′-Methoxyresveratrol Alleviated AGE-Induced Inflammation via RAGE-Mediated NF-κB and NLRP3 Inflammasome Pathway. Molecules 2018, 23, 1447. https://doi.org/10.3390/molecules23061447
Yu W, Tao M, Zhao Y, Hu X, Wang M. 4′-Methoxyresveratrol Alleviated AGE-Induced Inflammation via RAGE-Mediated NF-κB and NLRP3 Inflammasome Pathway. Molecules. 2018; 23(6):1447. https://doi.org/10.3390/molecules23061447
Chicago/Turabian StyleYu, Wenzhe, Mengru Tao, Yueliang Zhao, Xiaoqian Hu, and Mingfu Wang. 2018. "4′-Methoxyresveratrol Alleviated AGE-Induced Inflammation via RAGE-Mediated NF-κB and NLRP3 Inflammasome Pathway" Molecules 23, no. 6: 1447. https://doi.org/10.3390/molecules23061447
APA StyleYu, W., Tao, M., Zhao, Y., Hu, X., & Wang, M. (2018). 4′-Methoxyresveratrol Alleviated AGE-Induced Inflammation via RAGE-Mediated NF-κB and NLRP3 Inflammasome Pathway. Molecules, 23(6), 1447. https://doi.org/10.3390/molecules23061447