
Screening of Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from Caulerpa lentillifera



Abstract
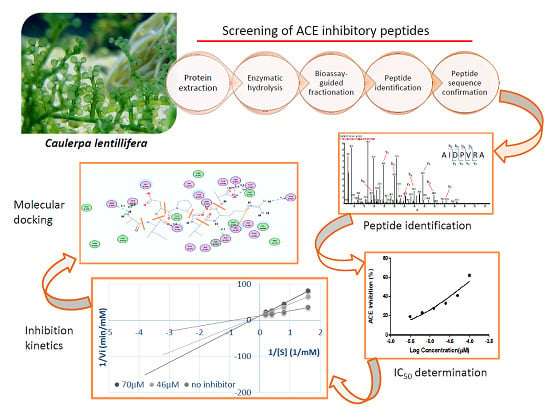
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
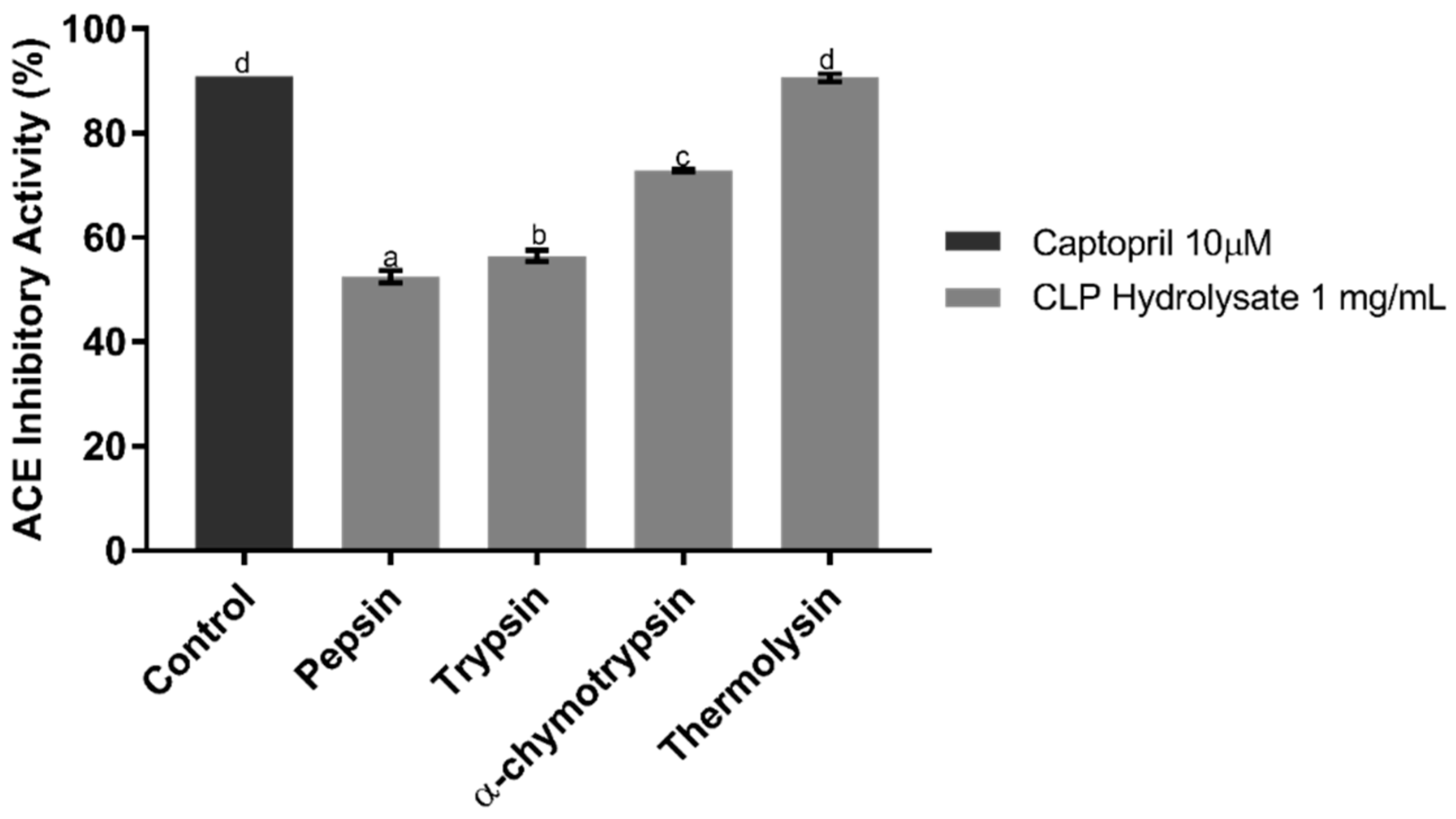
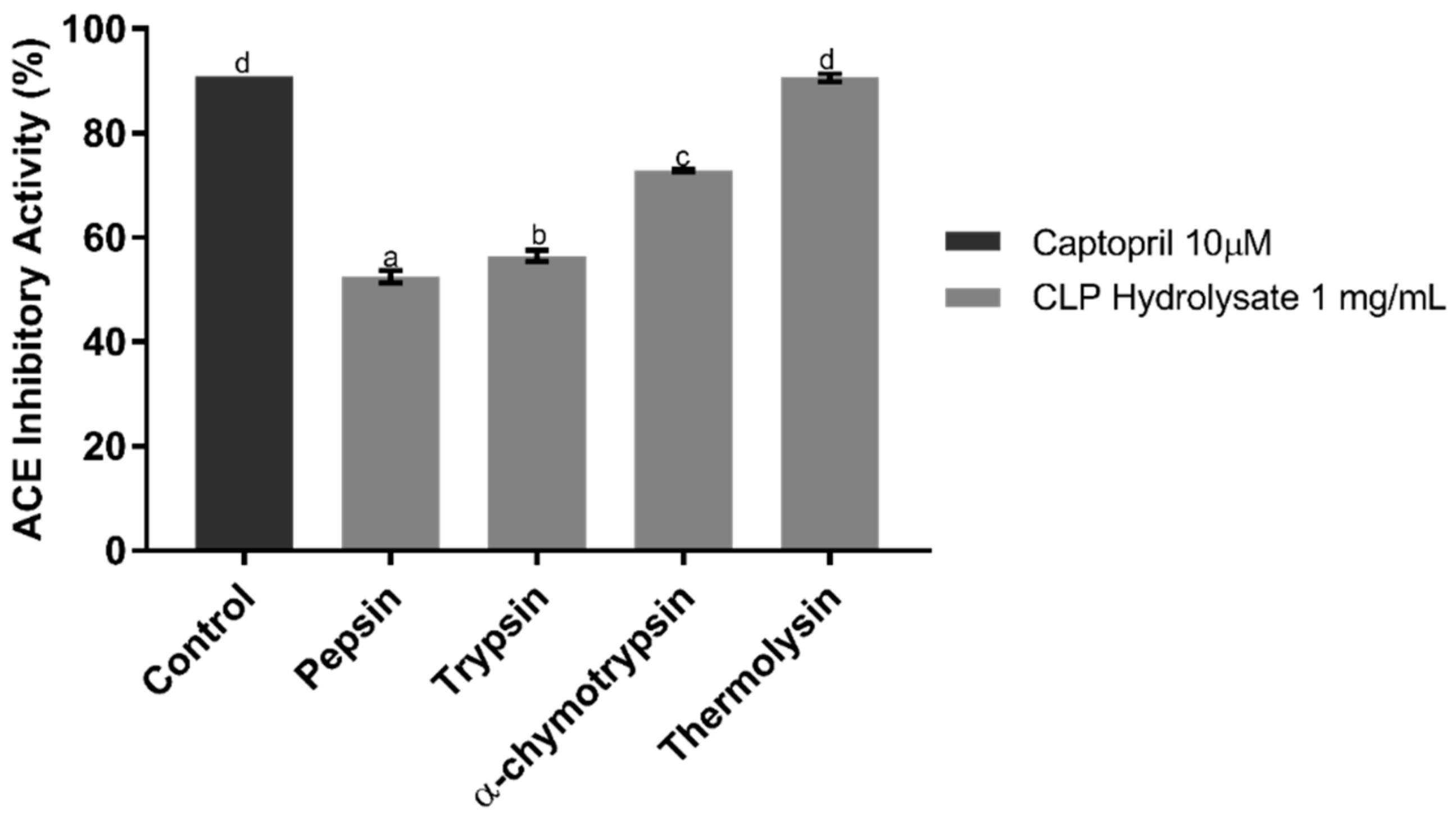
2.1. ACE Inhibitory Assay of CLP Hydrolysates
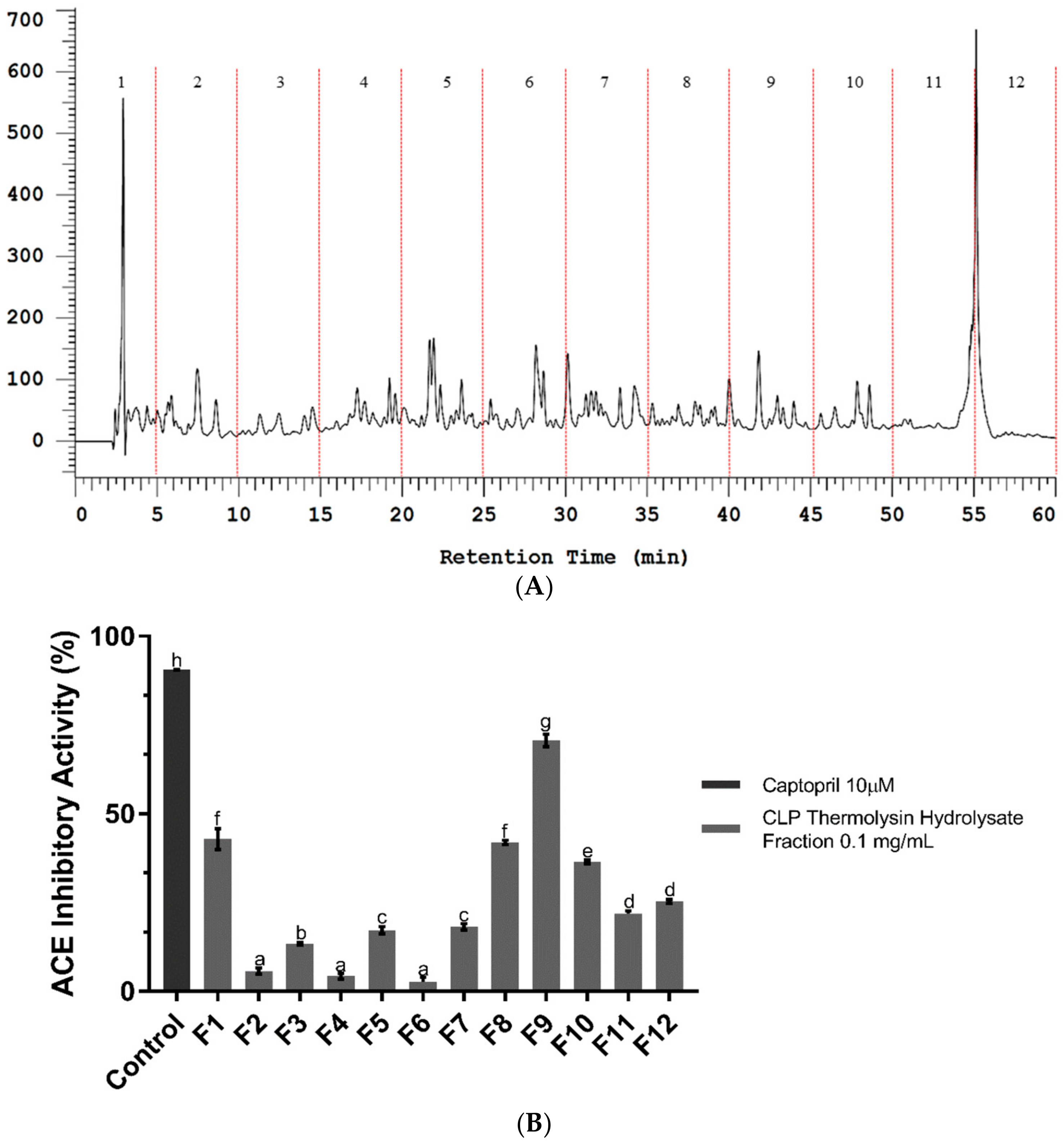
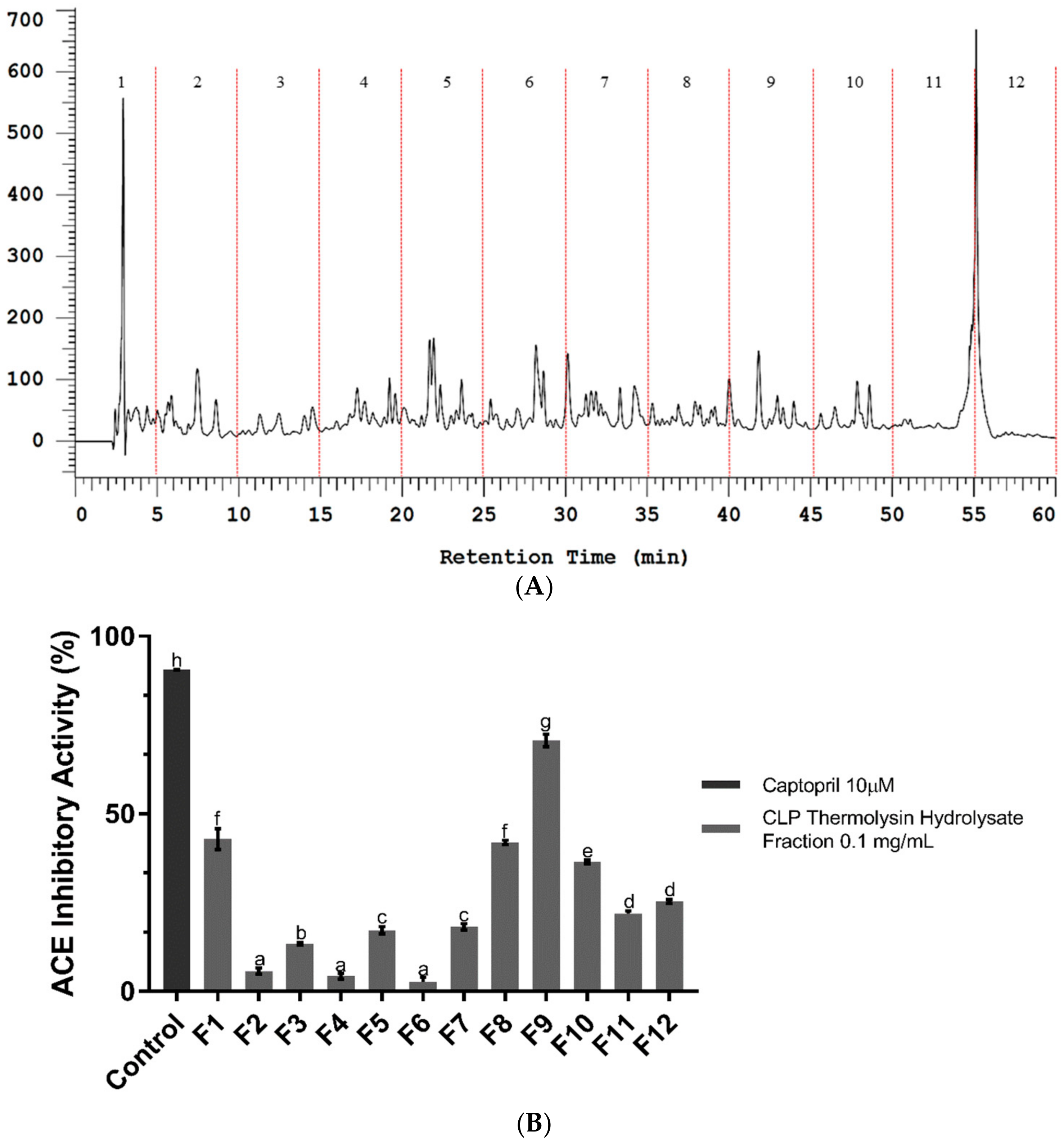
2.2. Bioassay-Guided Fractionation of CLP Hydrolysate Digested by Thermolysin
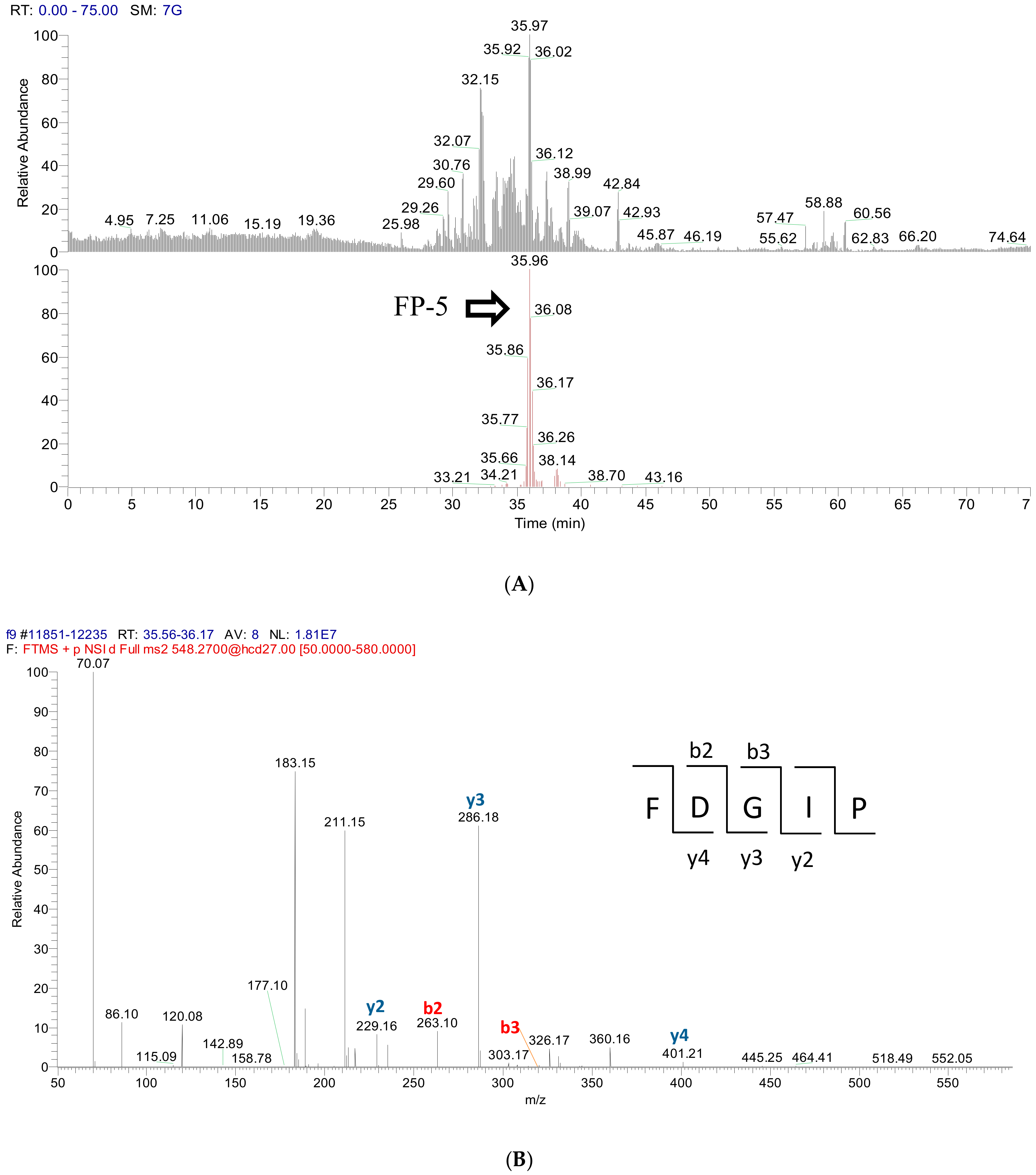
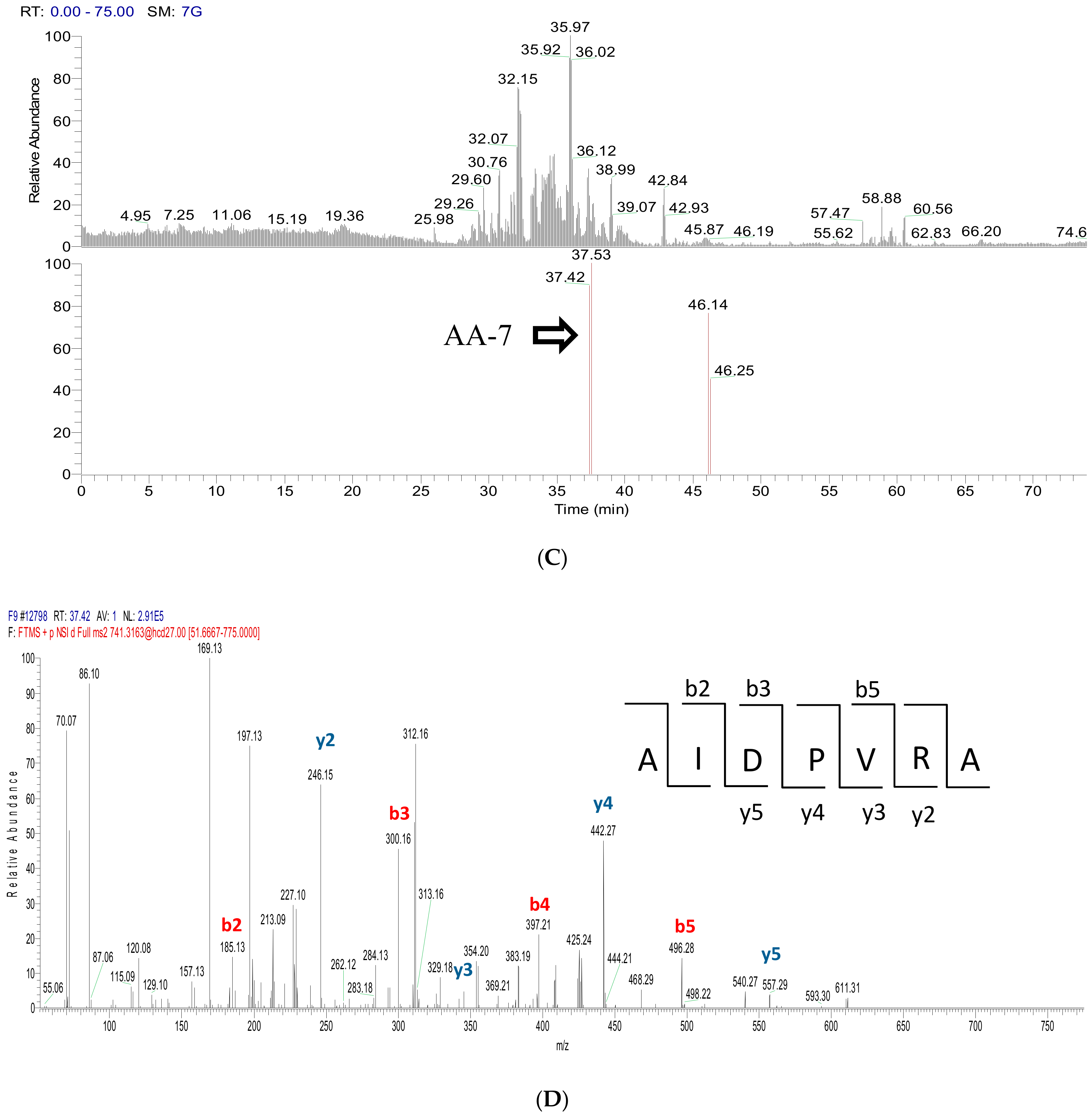
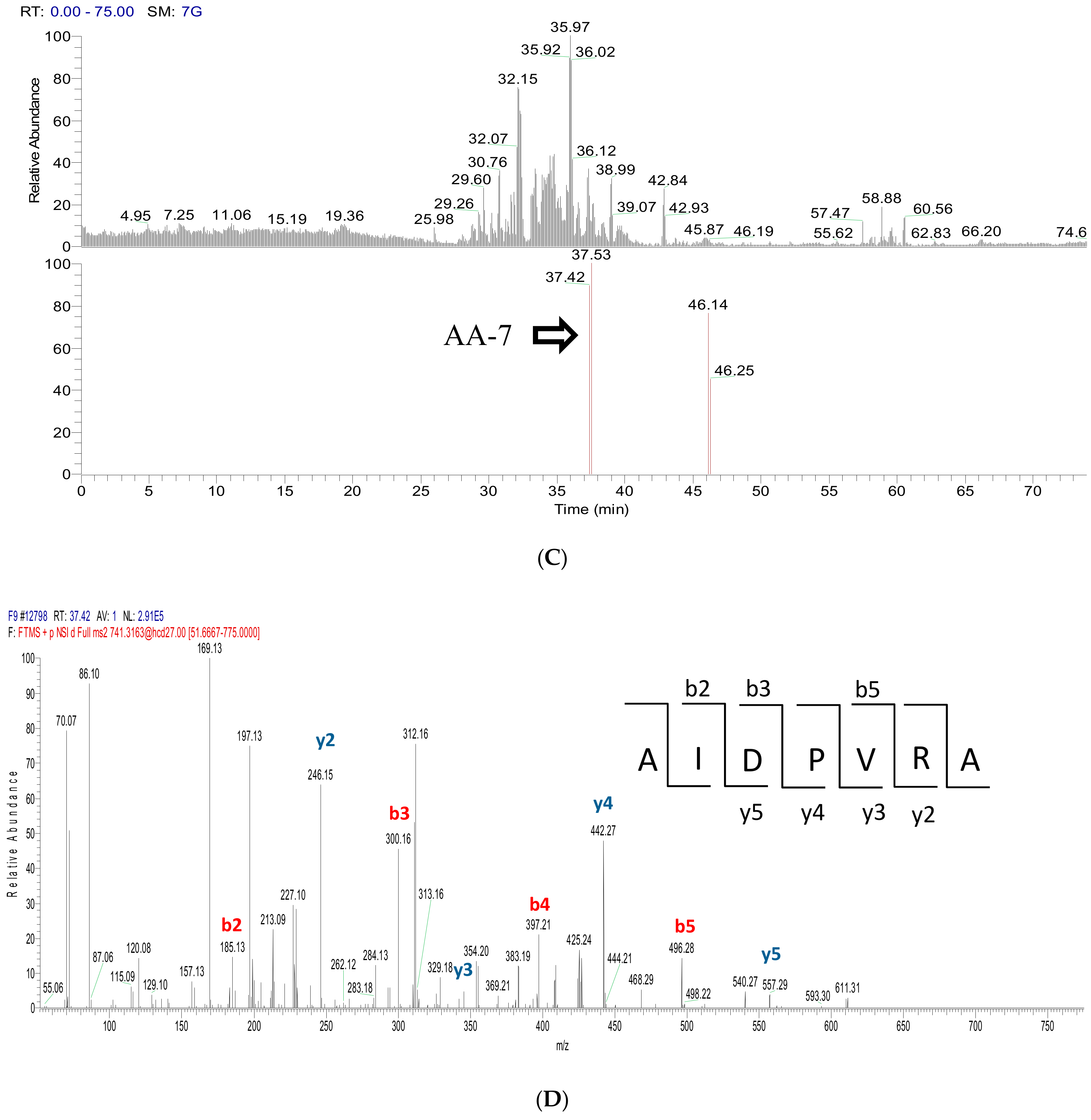
2.3. Peptide Sequence Identification
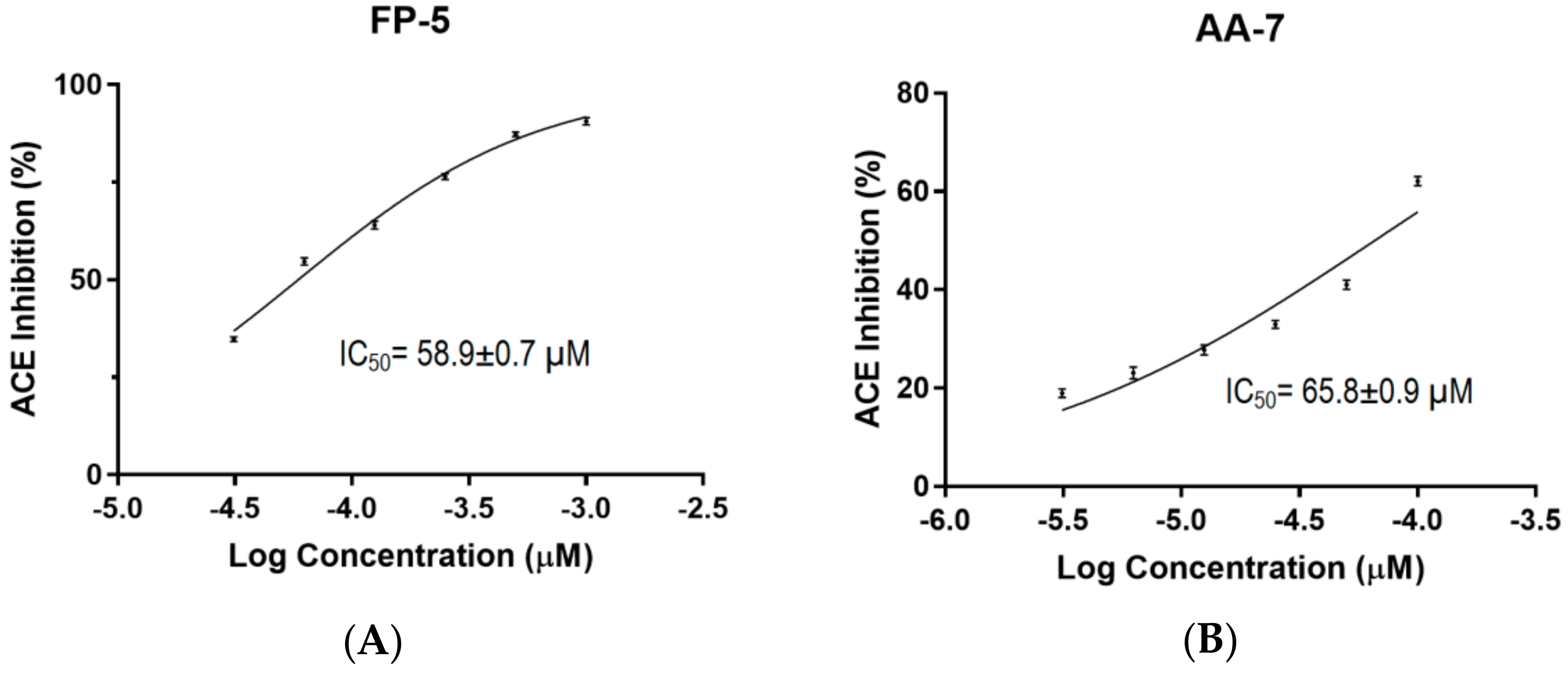
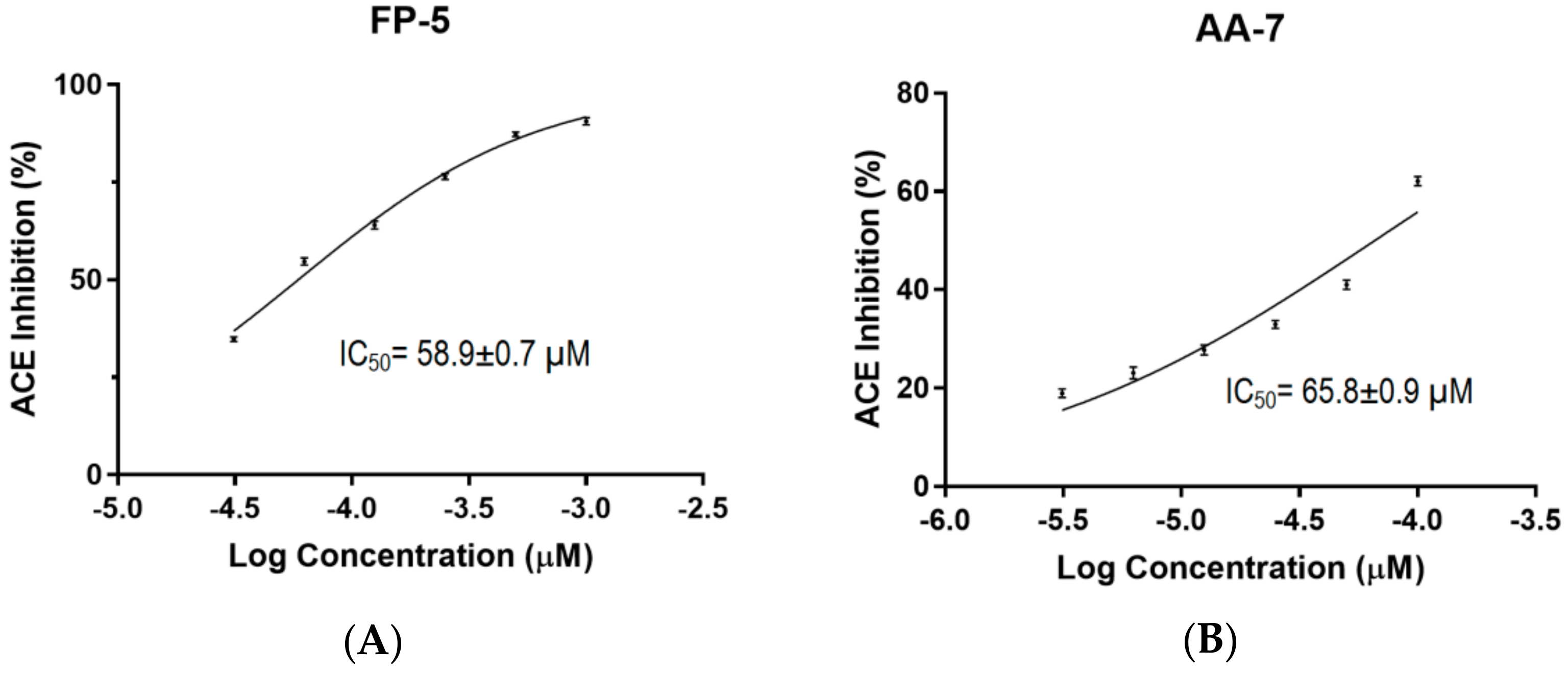
2.4. Determination of IC50 Values for FP-5 and AA-7
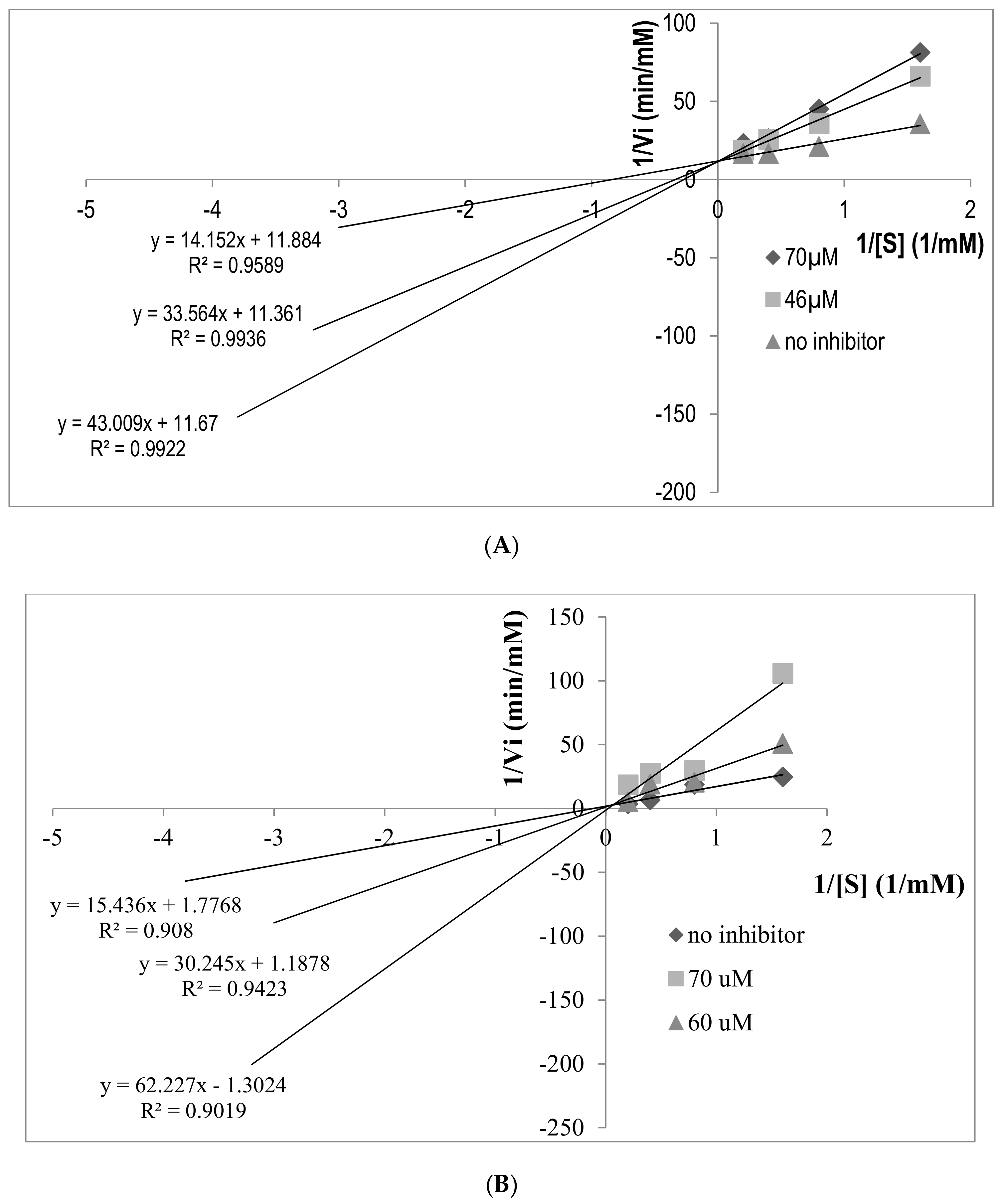
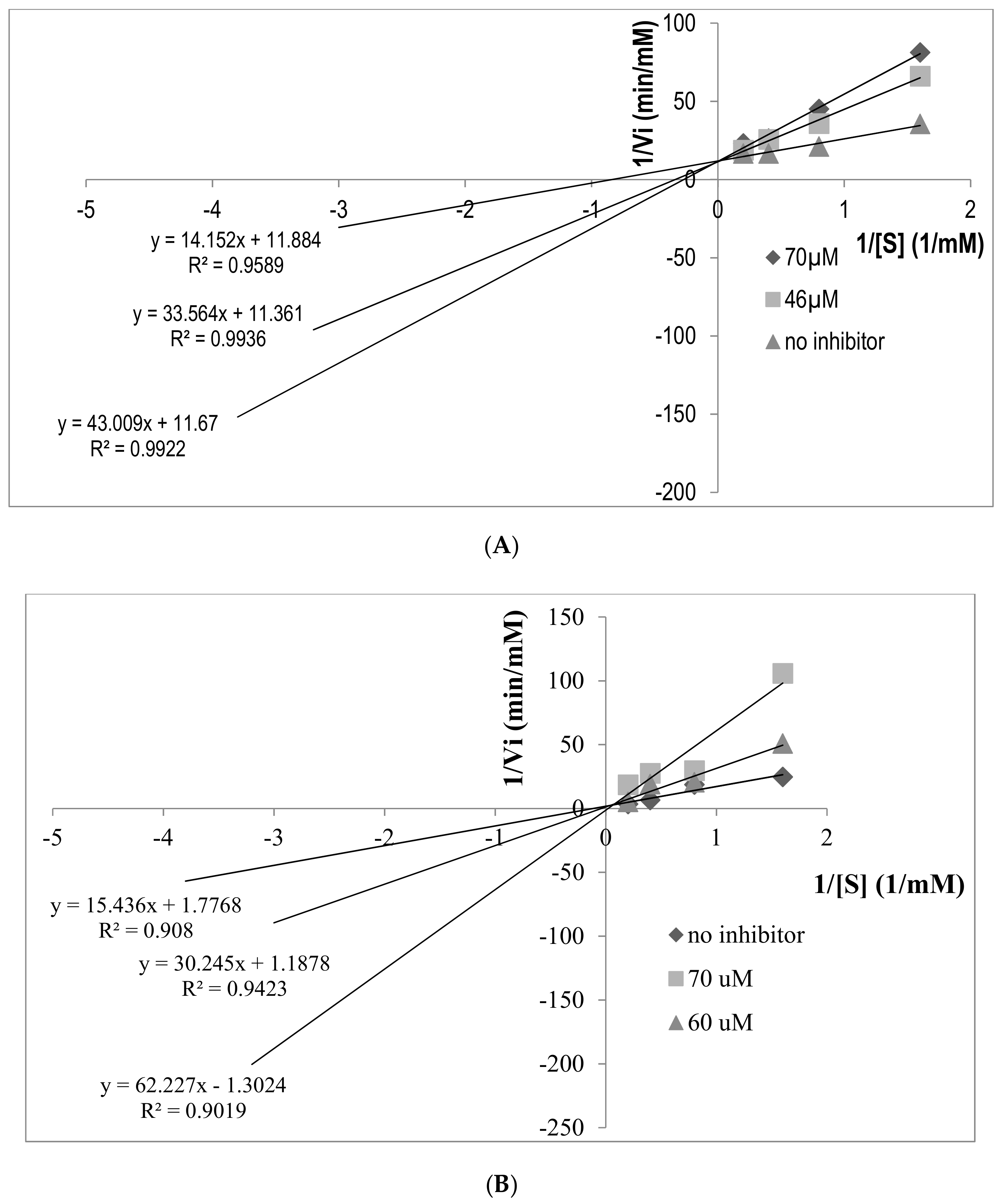
2.5. Determination of ACE Inhibition Pattern
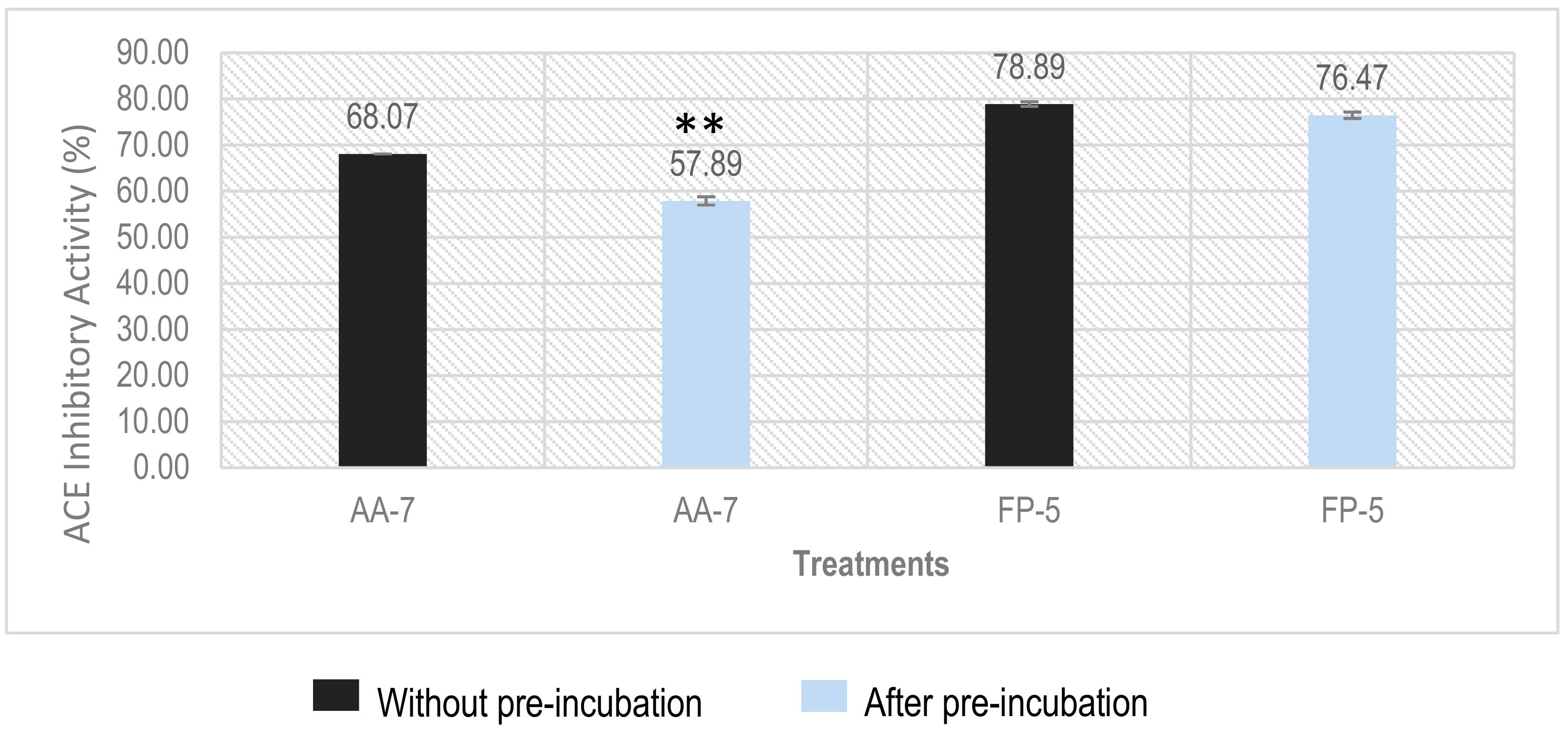
2.6. Determination of Inhibition Types of ACEI Peptides Using Pre-Incubation Experiment
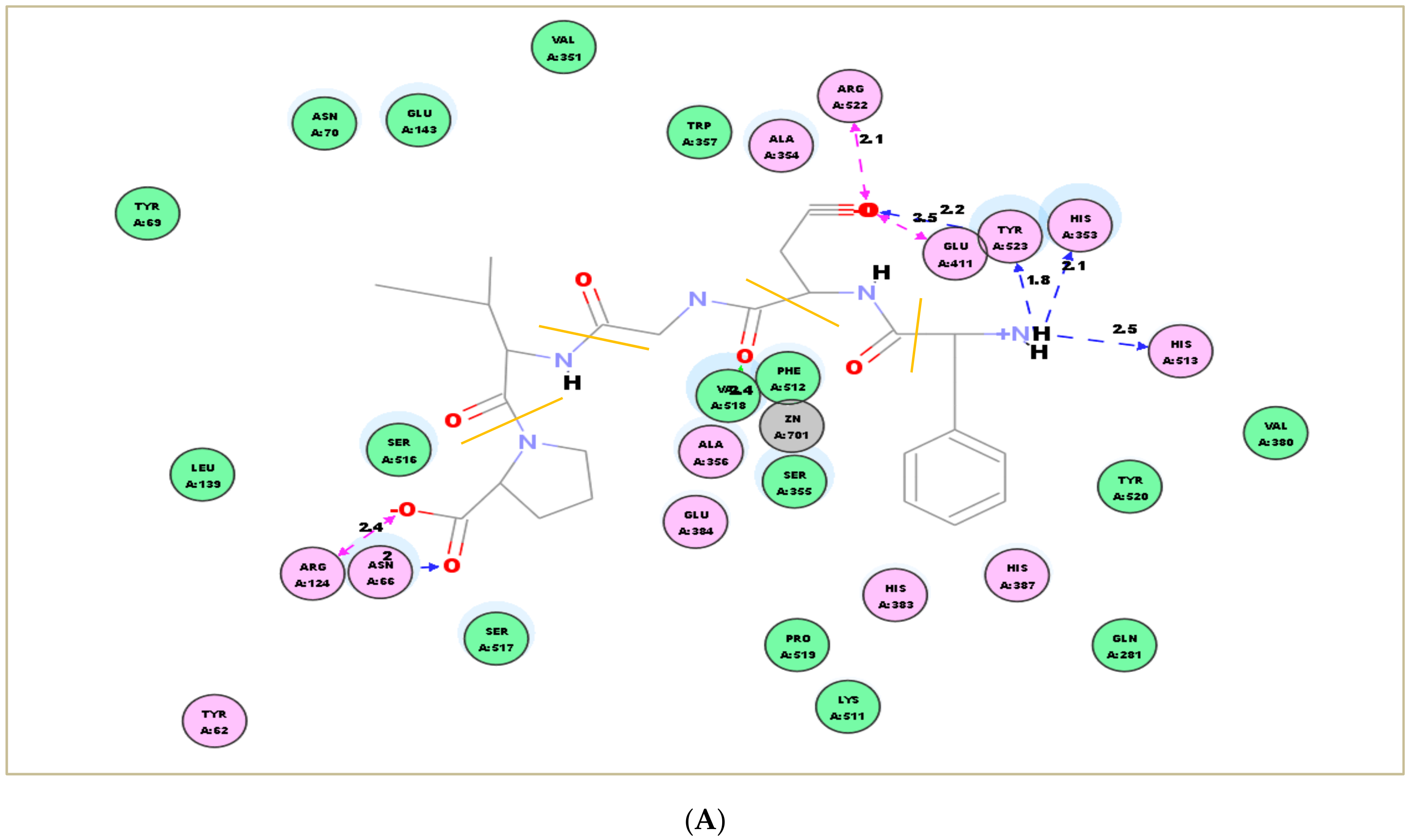
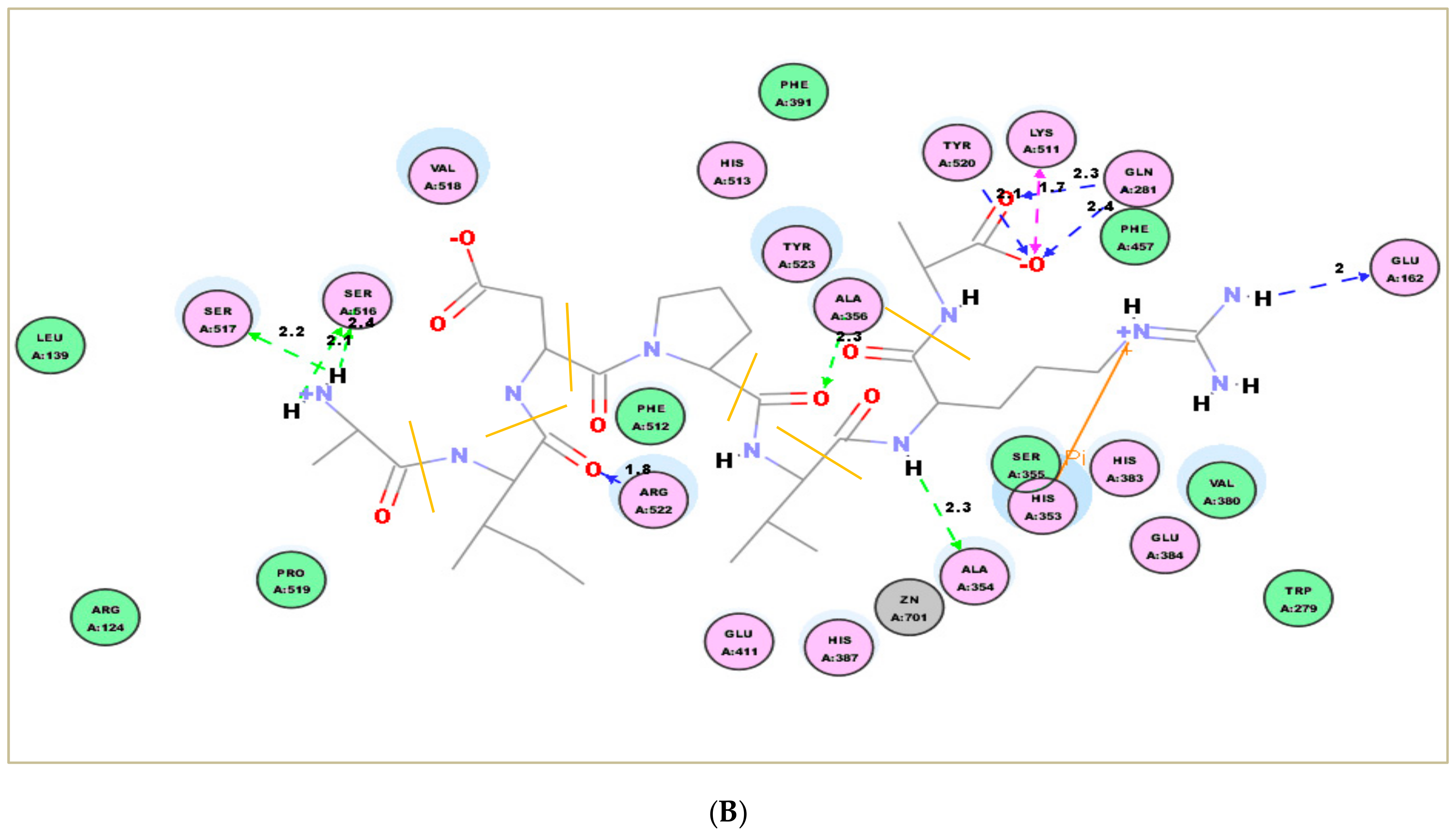
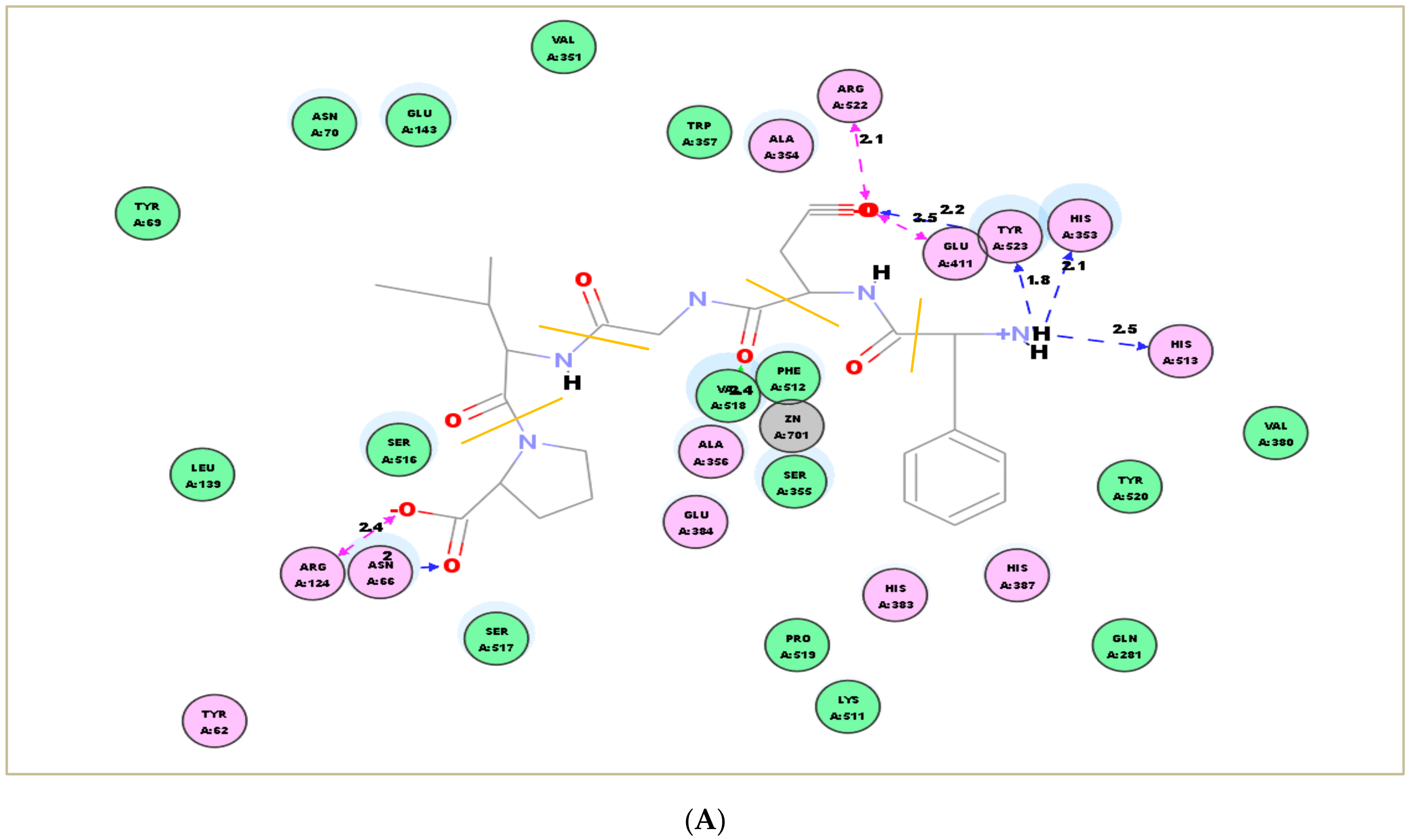
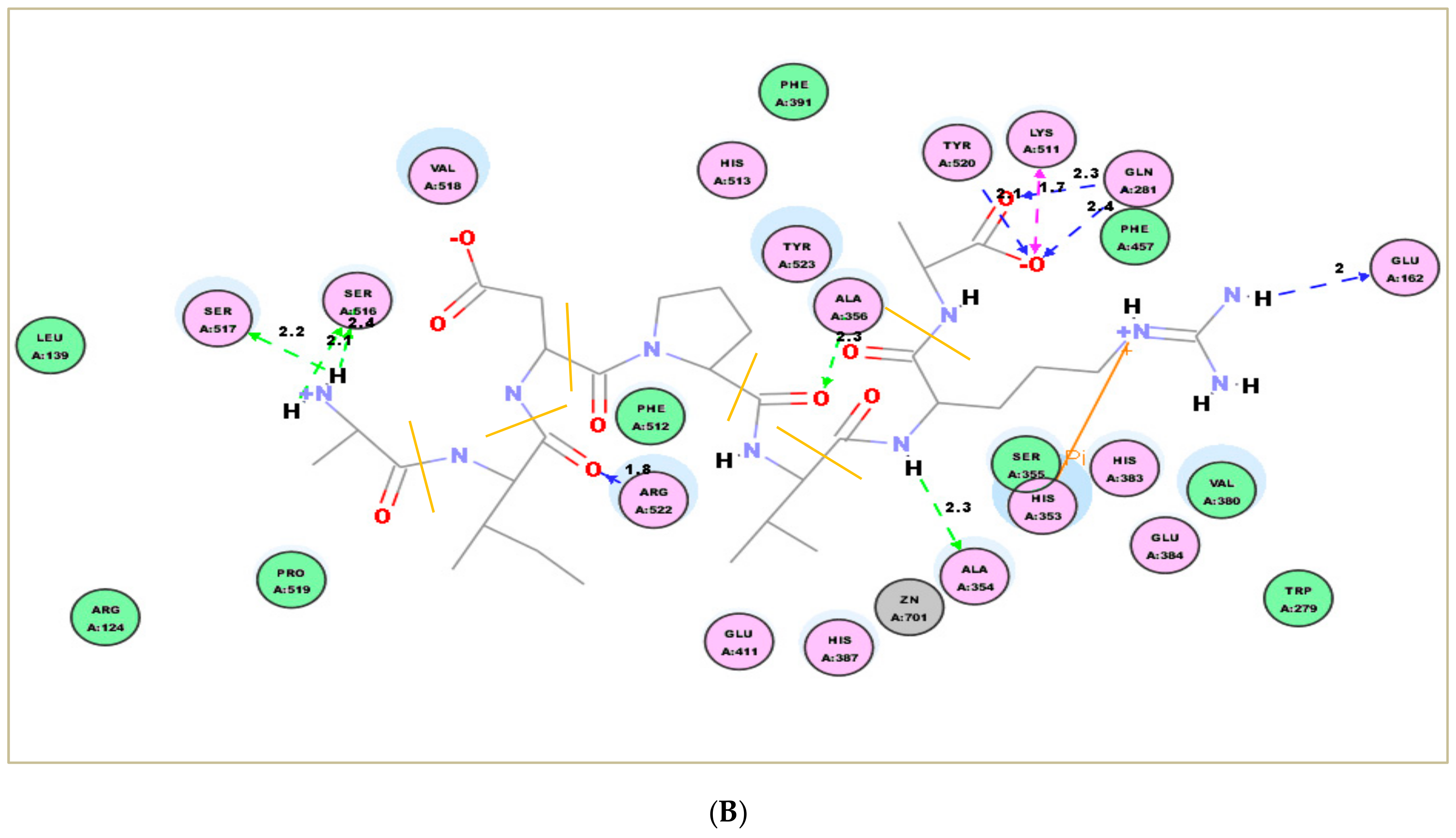
2.7. Molecular Docking Simulation
3. Materials and Methods
3.1. Materials and Chemical Reagents
3.2. Preparation of Protein from C. lentillifera
3.3. Enzymatic Digestion of CLP
3.4. Fractionation of CLP Hydrolysate
3.5. Measurement of ACE Inhibitory Activity
3.6. Identification of Peptide Sequences Using LC-MS/MS and De Novo Sequencing
3.7. Preparation of Synthetic Peptides
3.8. Determination of Inhibition Pattern by Inhibition Kinetics
3.9. Pre-Incubation Experiment to Determine the Inhibitor Type of ACEI Peptide
3.10. Molecular Docking Simulation
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gabb, G.M.; Mangoni, A.; Anderson, C.S.; Cowley, D.; Dowden, J.S.; Golledge, J.; Hankey, G.J.; Howes, F.S.; Leckie, L.; Perkovic, V.; et al. Guideline for the Diagnosis and Management of Hypertension in Adults-2016. Available online: https://www.mja.com.au/system/files/issues/10.5694mja16.00526.pdf (accessed on 12 February 2018).
- Ahluwali, M.; Bangalore, S. Management of hypertension in 2017: Targets and therapies. Curr. Opin. Cardiol. 2017, 32, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.; Dominguez, J.; Nguyen, L.; Gullapalli, N. Hypertension management: An update. Am. Health Drug Benefits 2010, 3, 47–56. [Google Scholar] [PubMed]
- Esther, C.R., Jr.; Marine, E.M.; Bernstein, K.E. The role of angiotensin-converting enzyme in blood pressure control, renal function, and male fertility. Trends Endocrinol. Metab. 1997, 8, 181–186. [Google Scholar] [CrossRef]
- Katzung, B.G.; Masters, S.B.; Trevor, A.J. Basic and Clinical Pharmacology, 12th ed.; McGraw-Hill: New York, NY, USA, 2012. [Google Scholar]
- Ogihara, T.; Mikami, H.; Katahira, K.; Otsuka, A. Comparative study of the effects of three angiotensin converting enzyme inhibitors on the cough reflex. Am. J. Hypertens. 1991, 4, S46–S51. [Google Scholar] [CrossRef]
- Saleh, A.S.; Zhang, Q.; Shen, Q. Recent research in antihypertensive activity of food protein-derived hydrolyzates and peptides. Crit. Rev. Food Sci. Nutr. 2016, 56, 760–787. [Google Scholar] [CrossRef] [PubMed]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting enzyme (ACE)-inhibitory peptides from plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; del Mar Contreras, M.; Recio, I. Antihypertensive peptides: Production, bioavailability and incorporation into foods. Adv. Colloid Interface Sci. 2011, 165, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.; Luo, M.; Sheng, Y.; Li, Z.; Wu, Y.; Liu, C. The antihypertensive effect of peptides: A novel alternative to drugs? Peptides 2008, 29, 1062–1071. [Google Scholar]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- Diego, A.; Vital, L.; Mojica, L.; de Mejía, E.G.; Mendoza, S.; Loarca-Piña, G. Biological potential of protein hydrolysates and peptides from common bean (Phaseolus vulgaris). Food Res. Int. 2015, 76, 39–50. [Google Scholar]
- Manilal, A.; Sujith, S.; Kiran, G.S.; Selvin, J.; Shakir, C.; Gandhimathi, R. Bio-potentials of seaweeds collected from Southwest Coast of India. J. Mar. Sci. Technol. 2009, 17, 67–73. [Google Scholar]
- Cha, S.H.; Lee, K.W.; Jeon, Y.J. Screening of extracts from red algae in Jeju for potentials marine angiotensin-I converting enzyme (ACE) inhibitory activity. Algae 2006, 21, 343–348. [Google Scholar] [CrossRef]
- Mary, A.; Mary, V.; Lorella, A.; Matias, J.R. Rediscovery of naturally occurring seagrape Caulerpa lentillifera from the gulf of mannar and its mariculture. Curr. Sci. 2009, 97, 1418–1420. [Google Scholar]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K. Nutrient content of tropical edible seaweeds, Eucheuma cottonii, Caulerpa lentillifera, and Sargassum polycystum. J. Appl. Phycol. 2009, 21, 75–80. [Google Scholar] [CrossRef]
- Maeda, R.; Ida, T.; Ihara, H.; Sakamoto, T. Induction of apoptosis in MCF-7 cells by β-1,3-xylooligosaccharides prepared from Caulerpa lentillifera. Biosci. Biotechnol. Biochem. 2012, 76, 1032–1034. [Google Scholar] [CrossRef] [PubMed]
- Wetli, H.A.; Brenneisen, R.; Tschudi, I.; Langos, M.; Bigler, P.; Sprang, T.; Schürch, S.; Mühlbauer, R.C. A gamma-glutamyl peptide isolated from onion (Allium cepa L.) by bioassay-guided fractionation inhibits resorption activity of osteoclasts. J. Agric. Food Chem. 2005, 53, 3408–3414. [Google Scholar] [CrossRef] [PubMed]
- Pank, M.; Kirret, O.; Paberit, N.; Aaviksaar, A. Hydrophobic interaction in thermolysin specificity. FEBS Lett. 1982, 142, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, K.; Chiba, H.; Yoshikawa, M. Peptide inhibitors for angiotensin I-converting enzyme from thermolysin digest of dried bonito. Biosci. Biotechnol. Biochem. 1992, 56, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Kamath, V.; Niketh, S.; Chandrashekar, A.; Rajini, P.S. Chymotryptic hydrolysates of α-kafirin, the storage protein of sorghum (Sorghum bicolor) exhibited angiotensin converting enzyme inhibitory activity. Food Chem. 2007, 100, 306–311. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Angiotensin I-converting enzyme (ACE) inhibitory activity, antioxidant properties, phenolic content and amino acid profiles of Fucus spiralis L. protein hydrolysate fractions. Mar. Drugs 2017, 15, 311. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.G. A unifying review of bioassay-guided fractionation, effect-directed analysis and related techniques. Sensors 2012, 12, 9181–9209. [Google Scholar] [CrossRef] [PubMed]
- Millioni, R.; Franchin, C.; Pivato, M.; Tessari, P.; Arrigoni, G. Sample loading influences studies comparing isoelectric focusing vs. strong cation exchange peptide fractionation. J. Chromatogr. A 2013, 1307, 207–208. [Google Scholar] [CrossRef] [PubMed]
- Priyanto, A.D.; Doerksen, R.J.; Chang, C.I.; Sung, W.C.; Widjanarko, S.B.; Kusnadi, J.; Lin, Y.C.; Wang, T.C.; Hsu, J.L. Screening, discovery, and characterization of angiotensin I-converting enzyme inhibitory peptides derived from proteolytic hydrolysate of bitter melon seed proteins. J. Proteomics 2015, 128, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Aluko, R.E.; Hossain, M.; Rai, D.K.; Hayes, M. Potential of a renin inhibitory peptide from the red seaweed Palmaria palmata as a functional food ingredient following confirmation and characterization of a hypotensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2014, 62, 8352–8356. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Chen, J-R. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Morihara, K.; Tswzuki, H. Thermolysin: Kinetic study with oligopeptides. Eur. J. Biochem. 1970, 15, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Zhou, H.; Qian, H. Enzymatic preparation and functional properties of wheat gluten hydrolysates. Food Chem. 2007, 101, 615–620. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-originating ACE inhibitors, including antihypertensive peptides, as preventive food components in blood pressure reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Pihlanto-Leppälä, A. Bioactive peptides derived from bovine whey proteins: Opioid and ace-inhibitory peptides. Trends Food Sci. Technol. 2000, 11, 347–356. [Google Scholar] [CrossRef]
- Natesh, R.; Schwager, S.L.; Sturrock, E.D.; Acharya, K.R. Crystal structure of the human angiotensin-converting enzyme lisinopril complex. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Bioactive Peptides. In Functional Foods and Nutraceuticals, 1st ed.; Springer: New York, NY, USA, 2012; pp. 37–61. [Google Scholar]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Ruiz, J.A.G.; Ramos, M.; Recio, I. Angiotensin converting enzyme-inhibitory activity of peptides isolated from Manchego cheese. Stability under simulated gastrointestinal digestion. Int. Dairy J. 2004, 14, 1075–1080. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joel, C.H.; Sutopo, C.C.Y.; Prajitno, A.; Su, J.-H.; Hsu, J.-L. Screening of Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from Caulerpa lentillifera. Molecules 2018, 23, 3005. https://doi.org/10.3390/molecules23113005
Joel CH, Sutopo CCY, Prajitno A, Su J-H, Hsu J-L. Screening of Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from Caulerpa lentillifera. Molecules. 2018; 23(11):3005. https://doi.org/10.3390/molecules23113005
Chicago/Turabian StyleJoel, Cesarea Hulda, Christoper C. Y. Sutopo, Arief Prajitno, Jui-Hsin Su, and Jue-Liang Hsu. 2018. "Screening of Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from Caulerpa lentillifera" Molecules 23, no. 11: 3005. https://doi.org/10.3390/molecules23113005
APA StyleJoel, C. H., Sutopo, C. C. Y., Prajitno, A., Su, J.-H., & Hsu, J.-L. (2018). Screening of Angiotensin-I Converting Enzyme Inhibitory Peptides Derived from Caulerpa lentillifera. Molecules, 23(11), 3005. https://doi.org/10.3390/molecules23113005