Integrated Proteomics and Lipidomics Investigation of the Mechanism Underlying the Neuroprotective Effect of N-benzylhexadecanamide

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Viability Assay
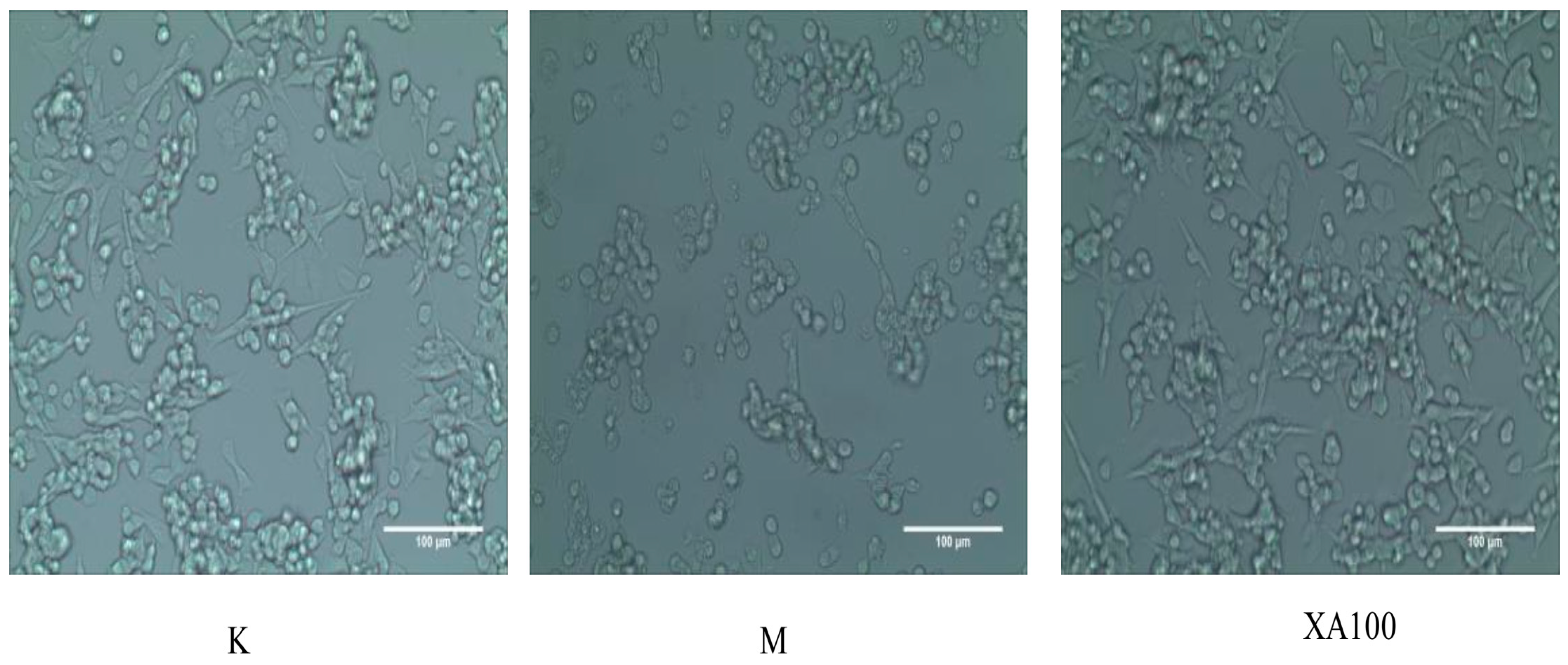
2.4. Observation of Cell Morphology
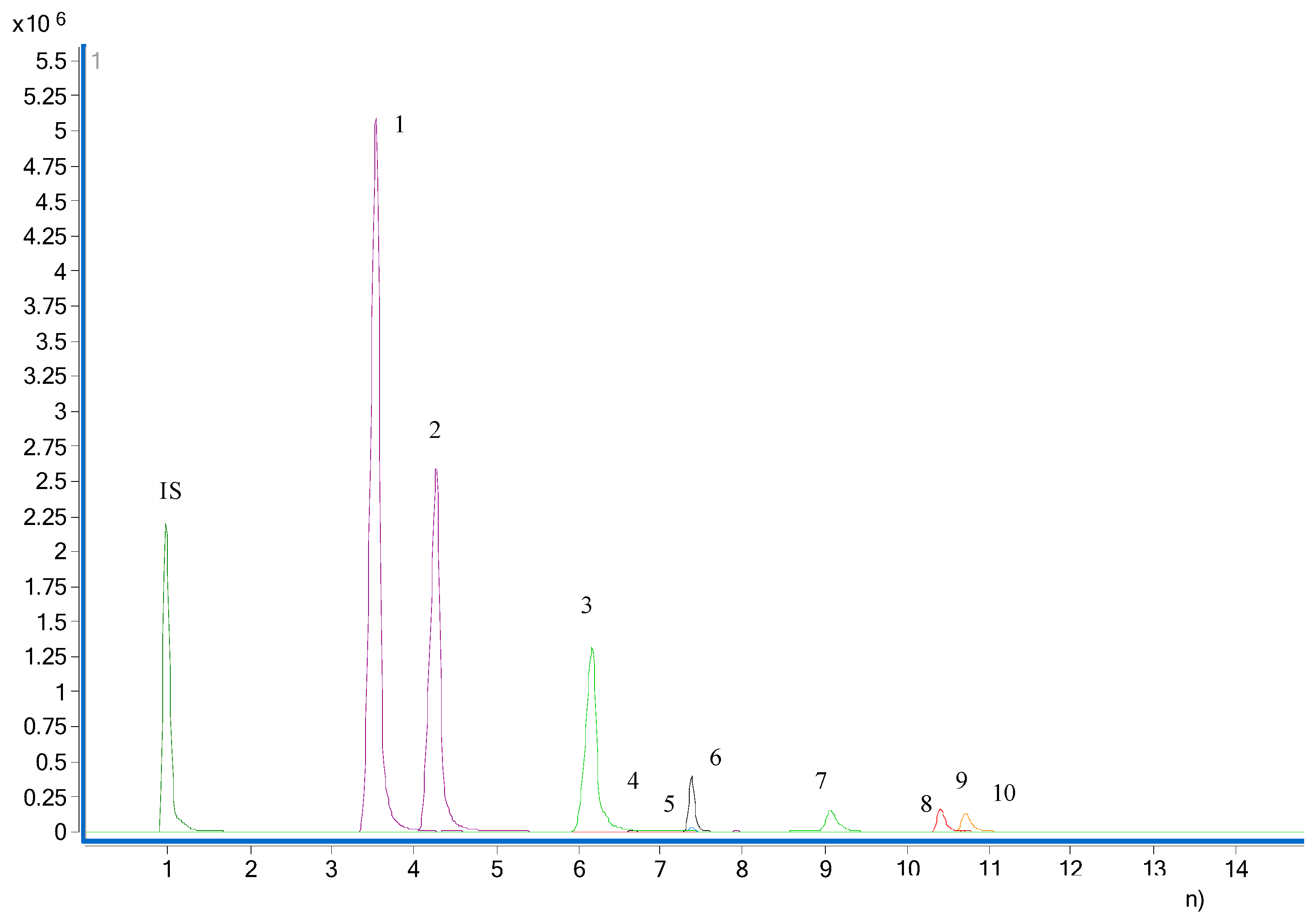
2.5. Measurement of Neurotransmitters
2.5.1. Preparation of Standards Solutions.
2.5.2. Pretreatment of Cell Supernatants and Cell Samples.
2.5.3. Chromatographic Conditions.
3. Lipidomic and Proteomic Analyses
3.1. Incubation Conditions for Lipidomic and Proteomic Studies.
3.2. Preparation of Lipid Samples.
3.3. Preparation of Protein Samples
3.4. Lipidomic Analysis Using LC-MS
3.5. Proteomic Analysis Using Nanorplc-MS/MS.
4. Results and Discussion
4.1. Neuroprotective Effect of XA on MPP +-Induced MN9D Cells
Optimization of Suitable Concentrations of MPP+ and XA
4.2. Absolute and Relative Quantification of Neurotransmitters
Method Validation
4.3. Quantification of Neurotransmitters
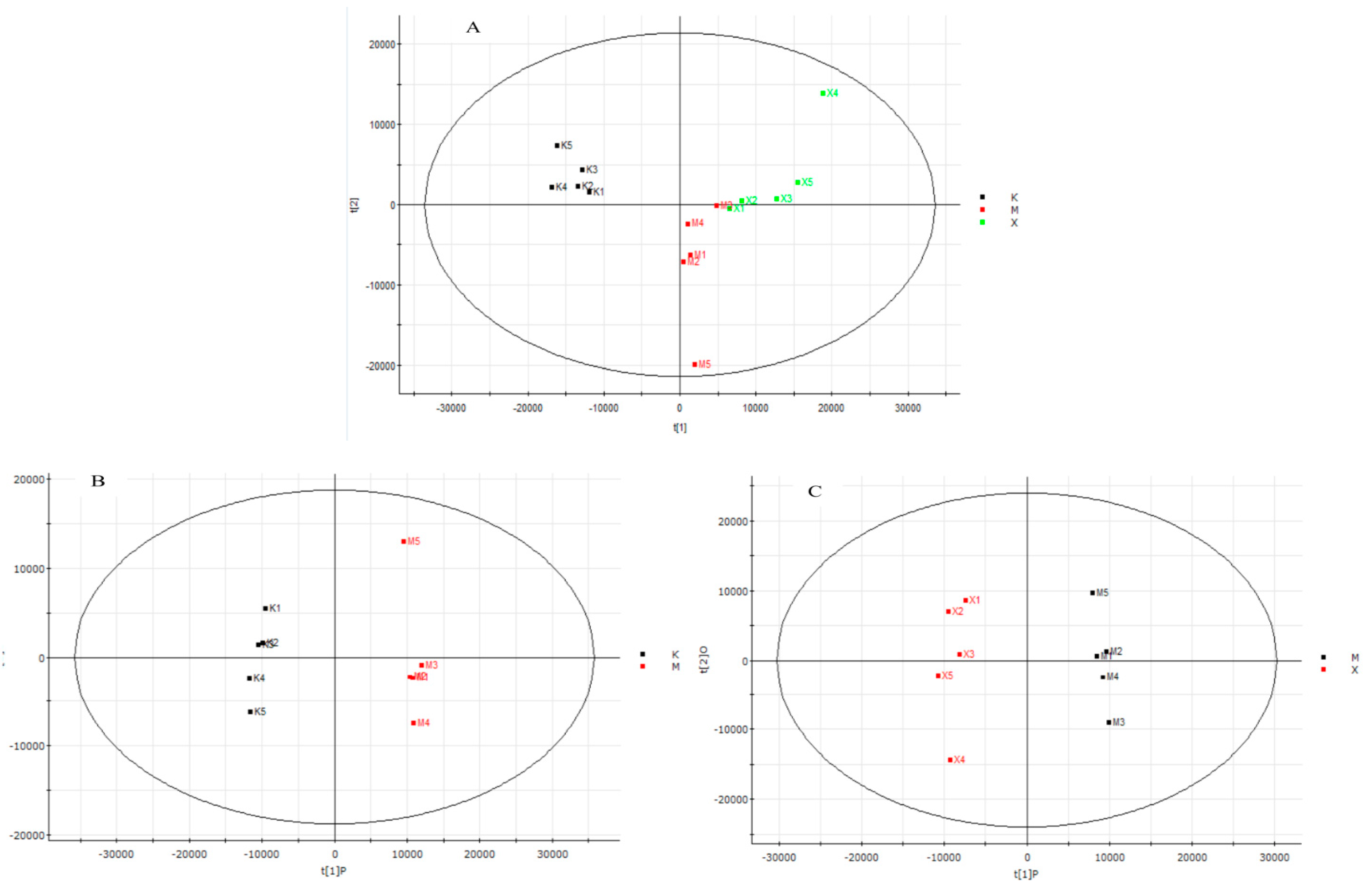
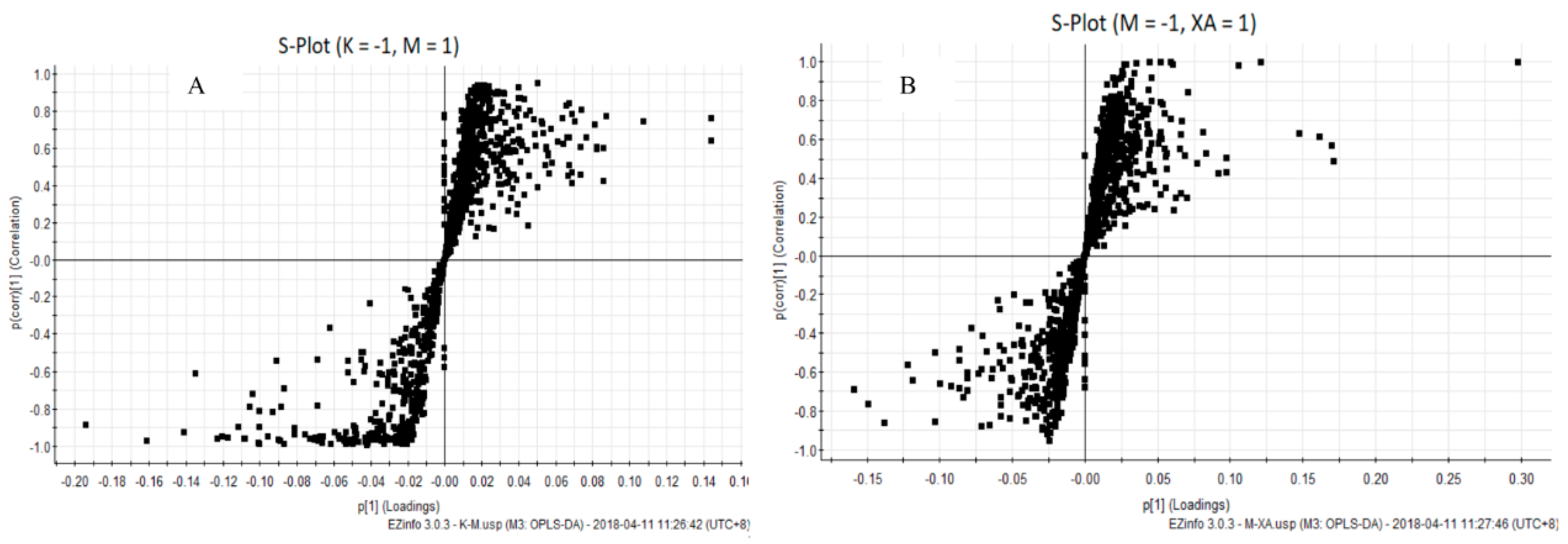
4.4. XA Regulates Distinct Lipid Profiles in MPP+-Induced MN9D Cell Model
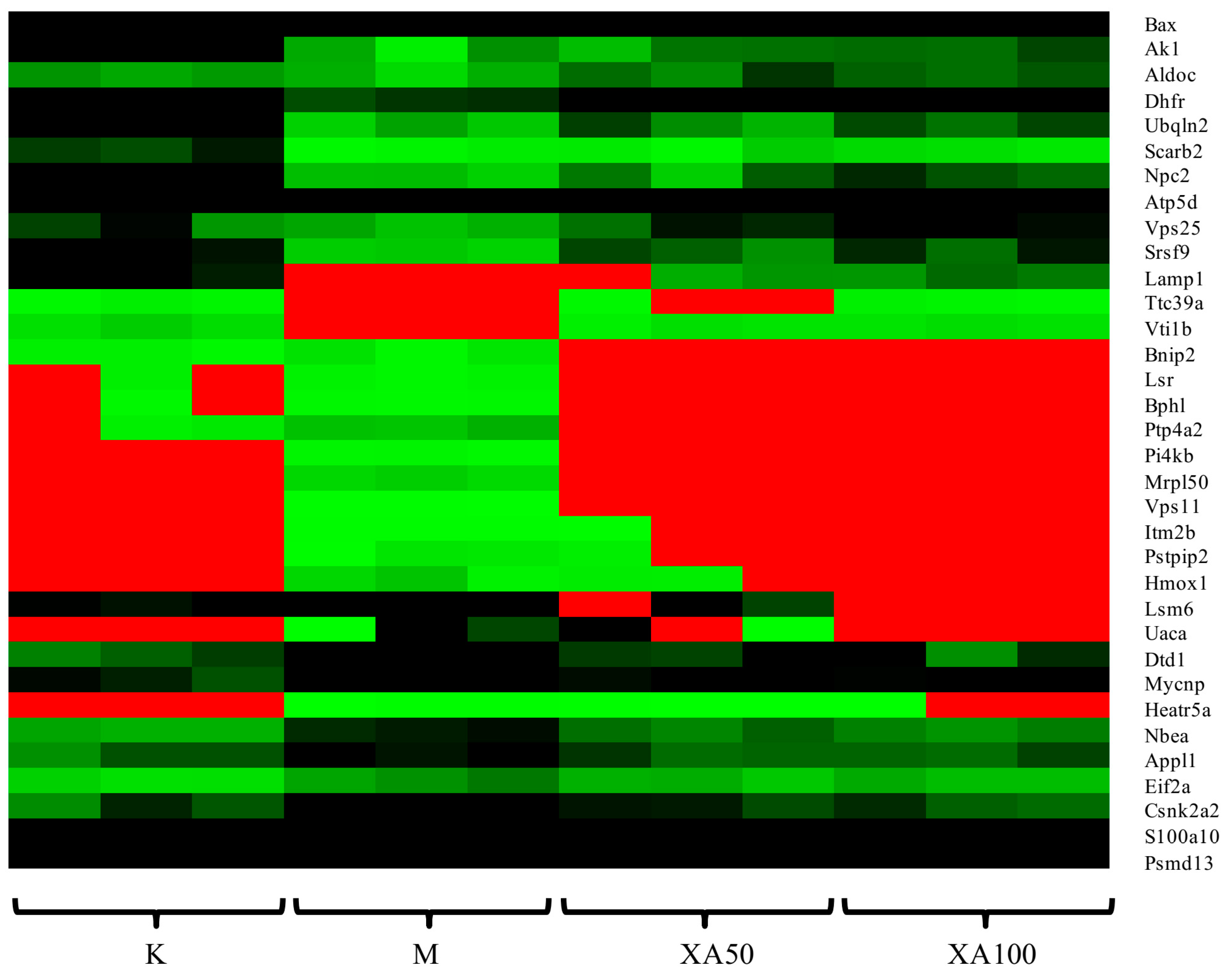
4.5. XA Regulates Distinct Protein Profiles in MPP+-induced MN9D Cell Model
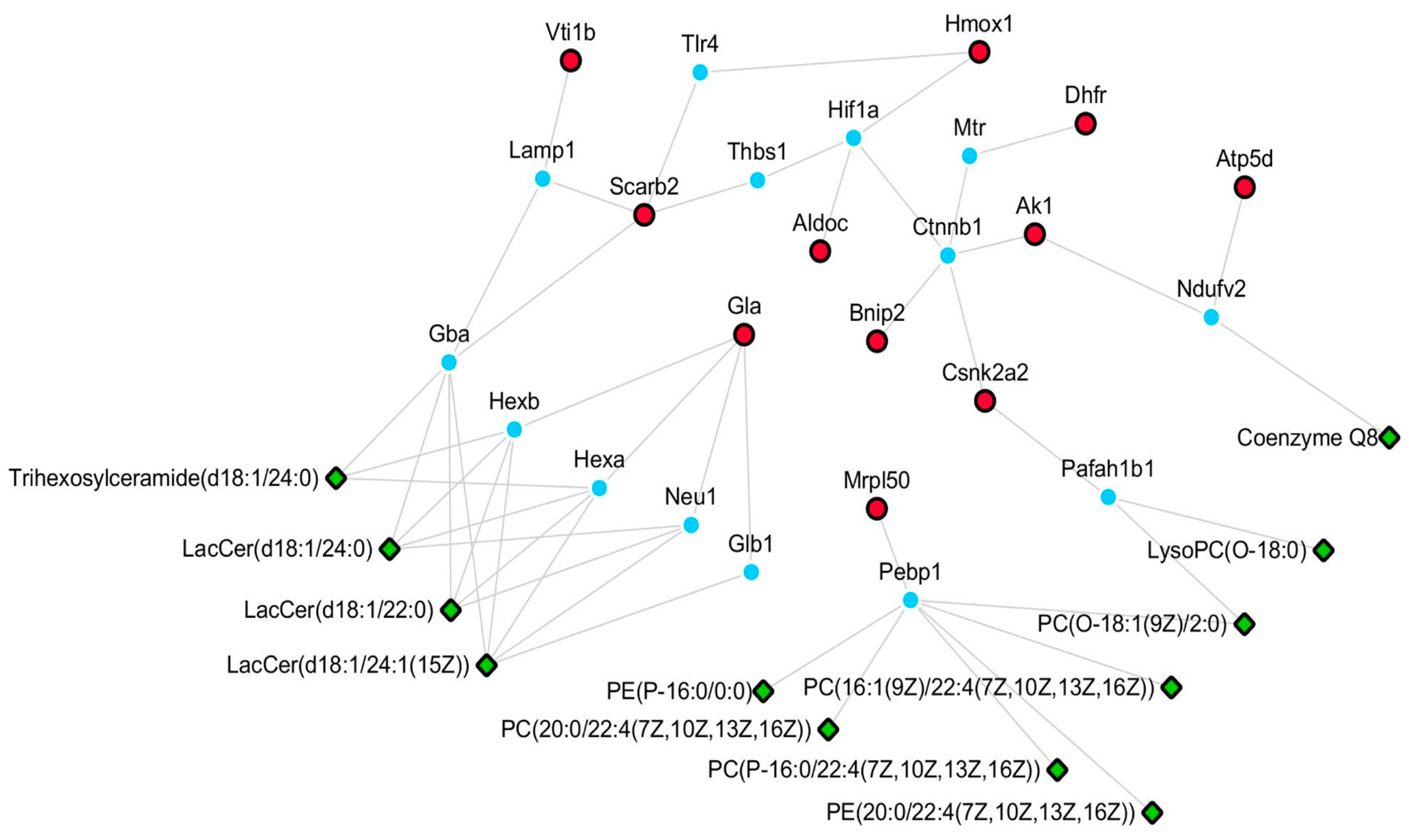
4.6. Potential Pathways Involved in the Neuroprotective Effect of XA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, Y.Y.; Zhao, H.Y.; Si, N.; Wang, H.J.; Bian, B.L. Recent researching progress of Lepidium meyenii (Maca). China J. Chin. Mater. Med. 2015, 40, 34–44. [Google Scholar]
- Guo, S.S.; Gao, X.F.; Gu, Y.R.; Wan, Z.X.; Lu, A.M.; Qin, Z.H.; Luo, L. Preservation of cognative function by Lepidium meyenii (Maca) is associated with improvement of mitochondrial activity and upregulation of autophay-related proteins in middle-aged mouse cortex. Evid.-Based Complement. Altern. 2016. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Jin, W.; Lv, X.; Dai, P.; Ao, Y.; Wu, M.; Deng, W.; Yu, L. Effects of macamides on endurance capacity and anti-fatigue property in prolonged swimming mice. Pharm. Biol. 2016, 54, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Huamán, A.; Casimiro-Gonzales, S.; ChávezPérez, J.A.; Gonzales-Arimborgo, C.; Cisneros-Fernández, R.; Aguilar-Mendoza, L.Á.; Gonzales, G.F. Antioxidant and neuroprotector effect of Lepidium meyenii (maca) methanol leaf extract against 6-hydroxy dopamine (6-OHDA)-induced toxicity in PC12 cells. Toxicol. Mech. Method 2017, 27, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Gugnani, K.S.; Vu, N.; Rondón-Ortiz, A.N.; Böhlke, M.; Maher, T.J.; Pino-Figueroa, A. Neuroprotective activity of macamides on manganese-induced mitochondrial disruption in U-87 MG gliblastoma cells. Toxicol. Appl. Pharm. 2018, 340, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Kelley, C.J.; Pino-Figueroa, A.; Vu, H.D.; Maher, T.J. Macamides and their synthetic analogs: Evaluation of in vitro FAAH inhibition. Bioorg. Med. Chem. 2013, 21, 5188–5197. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, P.; Brantner, A.; Wang, H.; Shu, X.; Yang, J.; Si, N.; Han, L.; Zhao, H.; Bian, B. Chemical profiling analysis of maca using UHPLC-ESI-Orbitrap MS coupled with UHPLC-ESI-QQQ MS and the neuroprotective study on its active ingredients. Sci. Rep. 2017, 7, 44660. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Won, L.A.; Konyur, P.J.; Hammond, D.N.; Fox, A.P.; Wainer, B.H.; Hoffmann, P.C.; Heller, A. Immortalization of embryonic mesencephlalic dopaminergic neurons by somatic cell fusion. Brain Res. 1991, 552, 67–76. [Google Scholar] [CrossRef]
- Li, X.Z.; Zhang, S.N.; Lu, F.; Liu, C.F.; Wang, Y.; Bai, Y.; Wang, N.; Liu, S.M. Cerebral metabonomics study on Parkinson’s disease mice treated with extract of Acanthopanax senticosus harms. Phytomedicine 2013, 20, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.Q.; Zhang, Z.; Hou, L.; Zhang, L.; Zhang, J.Y.; Wang, Y. Phytic acid attenuates inflammatory responses and the levels of NFκB and p-ERK in MPTP-induced Parkinson’s disease model of mice. Neurosci. Lett. 2015, 597, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Santos-Fandila, A.; Zafra-Gómez, A.; Barranco, A.; Navalón, A.; Rueda, R.; Ramírez, M. Quantification determination of neurotransmitters, metabolites and derivates in microdialysates by UHPLC-tandem mass spectrometry. Talanta 2013, 114, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Kondo, F.; Tachi, M.; Gosho, M.; Fukayama, M.; Yoshikawa, K. Changes in hypothalamic neurotransmitter and prostanoid levels in response to NMDA, CRF, and GLP-1 stimulation. Anal. Bioanal. Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Jamwal, S.; Kumar, P. Piperine enhances the protective effect of curcumin against 3-NP induced neurotoxicity: Possible neurotransmitters modulation mechanism. Neurochem. Res. 2015, 40, 1758–1766. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wei, J.; Yang, M. Simultaneous Analysis of Iridoid Glycosides and Anthraquinones in Morinda officinalis Using UPLC-QqQ-MS/MS and UPLC-Q/TOF-MSE. Molecules 2018, 23, 1070. [Google Scholar] [CrossRef] [PubMed]
- Dimitrakopoulos, C.; Hindupur, S.K.; Häfliger, L.; Behr, J.; Montazeri, H.; Hall, M.N.; Beerenwinkel, N. Network-based integration of multi-omics data for prioritizing cancer genes. Bioinformatics 2018. [Google Scholar] [CrossRef] [PubMed]
- Francescatto, M.; Chierici, M.; Rezvan Dezfooli, S.; Zandonà, A.; Jurman, G.; Furlanello, C. Multi-omics integration for neuroblastoma clinical endpoint perdition. Biol. Direct. 2018, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Del Boccio, P.; Pieragostino, D.; Di Ioia, M.; Petrucci, F.; Lugaresi, A.; De Luca, G.; Gamni, D.; Onofri, M.; Di Llio, C.; Saccetta, P.; et al. Lipidomics investigation for the characterization of circulating serum lipids in multiple sclerosis. J. Prot. 2011, 74, 2826–2836. [Google Scholar] [CrossRef] [PubMed]
- Martins, I.J. The role of clinical proteomics, lipidomics, and genomics in the diagnosis of Alzheimer’s disease. Proteomes 2016, 4, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.; Astiz, M.; Magnani, N.; Imsen, M.; Merino, F.; Álvarez, S.; Reinés, A.; Seilicovich, A. Hormorne deprivation alters mitochondrial function and lipid profile in the hippocampus. J. Endocrinol. 2017, 233, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rabiei, Z.; Bigdeli, M.R.; Rasoulian, B.; Ghassempour, A.; Mirzajani, F. The neuroprotection effect of pretreatment with olive leaf extract on brain lipidomics in rat stroke model. Phytomedicine 2012, 19, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Yang, C.; Zhao, J.; Tse, H.F.; Rong, J. Proteomic identification of calcium-binding chaperone calreticulin as a potential mediator for the neuroprotective and neuritogenic activities of fruit-derived glycoside amygdalin. J. Nutr. Biochem. 2015, 26, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.J.; Zhou, J.T.; Chen, X.; Lou, Y.X.; Liu, D.; Zou, X.J.; Yang, B.; Yin, Y.X.; Pan, Y. Quantitative proteomics study of the neuroprotective effects of B12 on hydrogen peroxide-induced apoptosis in SH-SY5Y cells. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Fang, L.; Yang, Y.; Jiang, H.; Yang, H.; Zhang, H.; Zhou, H. Quantitative proteomic analysis reveals the neuroprotective effects of huperzine A for amyloid beta treated neuroblastoma N2a cells. Proteomics 2013, 13, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Mohamadpour, A.H.; Mehri, S.; Abnous, K.; Hassanzadeh-Khayyat, M. Proteomics screening of molecular targets of granulocyte colony stimulating factor in the mouse brain and PC12 cell line. Life Sci. 2014, 108, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Zhao, J.P.; Dunbar, D.C.; Khan, I.A. Constituents of Lepidium meyenii (maca). Phytochemistry 2002, 59, 105–110. [Google Scholar] [CrossRef]
- Sjödin, L.; Nederman, T.; Olsson, P.A.; Viitanen, E. In-vitro bioassays for cholecystokinin. J. Pharm. Pharmacol. 1989, 41, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Schou-Pedersen, A.M.V.; Hansen, S.N.; Tveden-Nyborg, P.; Lykkesfeldt, J. Simultaneous quantification of monoamine neurotransmitters andtheir biogenic metabolites intracellularly and extracellularly in primary neuronal cell cultures and in sub-regions of guinea pig brain. J. Chromatogr. B 2016, 1028, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.Y.; Zhang, F.B.; Zhang, Y.; Cao, C.Y.; Li, X.Y.; Li, D.F.; Liu, X.; Wang, H.J.; Huang, L.Q. Proteomic investigation of signatures for geniposide-induced hepatotoxicity. J. Proteome Res. 2014, 13, 5724–5733. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Q.H.; Hu, C.X.; Yu, D.; Wang, L.C.; Zhou, Y.; Zhao, X.J.; Li, Q.; Hou, X.L.; Xu, G.W. Development of A High Coverage Pseudotargeted Lipidomics Method Based on Ultra-High Performance Liquid Chromatography-Mass Spectrometry. Anal. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Song, C.M.; Ma, H.B. The biological rule of taurine. J. Jilin Med. Coll. 2005, 26, 231–234. [Google Scholar]
- Kumari, N.; Prentice, H.; Wu, J.Y. Taurine and its neuroprotective role. Adv. Exp. Med. Biol. 2013, 775, 19–27. [Google Scholar] [PubMed]
- Jhang, K.A.; Lee, E.O.; Chong, Y.H. Norepinephrine provides short-term neuroprotection against Aβ1-42 by reducing oxidative stress independent of Nrf2 activation. Neurobiol. Aging 2014, 35, 2465–2473. [Google Scholar] [CrossRef] [PubMed]
- Huo, J. The Effects of the Electro-Acupunture of Nuchal on the Cholinergic Neuronal and Monoamine Neurotransmitters in Laboratory Rats; Heilongjiang University of Chinese Medicine: Harbin, China, 2012. [Google Scholar]
- Acon-Chen, C.; Koenig, J.A.; Smith, G.R.; Truitt, A.R.; Thomas, T.P.; Shih, T.M. Evaluation of acetylcholine, seizure activity and neuropathology following high-dose nerve agent exposure and delayed neuroprotective treatment drugs in freely moving rats. Toxicol. Mech. Method 2016, 26, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Wang, J.H.; Yau, L.F.; Ho, H.M.; Chan, C.L.; Hu, P.; Liu, L.; Jiang, Z.H. A cellular lipidomic study on the Aβ-induced neurotoxicity and neuroprotective effects of EGCG by using UPLC/MS-based glycerolipids profiling and multivariate analysis. Mol. BioSyst. 2012, 8, 3208–3215. [Google Scholar] [CrossRef] [PubMed]
- Shazeeb, M.S.; Feula, G.; Bogdanov, A. Liposome-encapsulated superoxide dismutase mimetic: Theranostic potential of an MR detectable and neuroprotective agent. Contrast Med. Mol. Imaging 2014, 9, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Wu, L.; Chu, M.; Feng, H.; Lu, C.; Wang, Q.; He, M.; Ge, X. Effects of breviscapine on amyloid beta 1-42 induced Alzheimer’s disease mice: A HPLC-QTOF-MS based plasma metabonomics study. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1057, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Ikeno, Y.; Cheon, S.H.; Konno, N.; Nakamura, A.; Kitamoto, K.; Arioka, M. Lysophosphatidylcholine protects cerebellar granule neurons from apoptotic cell death. J. Neurosci. Res. 2009, 87, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Megighian, A.; Rigoni, M.; Caccin, P.; Zordan, M.A.; Montecucco, C. A lysolecithin/fatty acid mixture promotes and then blocks neurotransmitter release at the Drosophila melanogaster larval neuromuscular junction. Neurosci. Lett. 2007, 416, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Cao, C.; Bao, W.; Yang, S.; Shi, H.; Hao, D.; Zhao, X.; Wu, Y. The plasma metabolic profiling of chronic acephate exposure in rats via an ultra-performance liquid chromatography-mass spectrometry based metabonomic method. Mol. Biosyst. 2015, 11, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Many ceramides. J. Biol. Chem. 2011, 286, 27855–27862. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, C.; Martinez-Martinez, P. Ceramide function in the brain: When a slight tilt is enough. Cell. Mol. Life Sci. 2013, 70, 181–203. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, O.; Futerman, A.H. The role of the ceramide acyl chain length in neurodegeneration: Involvement of ceramide synthases. Neuromol. Med. 2010, 12, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Caspi, M.; Perry, G.; Skalka, N.; Meisel, S.; Firsow, A.; Amit, M.; Rosin-Arbesfeld, R. Aldolase positively regulates of the canonical Wnt signaling pathway. Mol. Cancer 2014, 13, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Zahid, S.; Khan, R.; Oellerich, M.; Ahmed, N.; Asif, A.R. Differential S-nitrosylation of proteins in Alzheimer’s disease. Neuroscience 2014, 256, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Cario, H.; Smith, D.E.C.; Blom, H.; Blau, N.; Bode, H.; Holzmann, K.; Pannicke, U.; Hopfner, K.-P.; Rump, E.-M.; Ayric, Z.; et al. Dihydrofolate reductase deficiency due to a homozygous DHFR mutation causes megaloblastic anemia and cerebral folate deficiency leading to severe neurologic disease. Am. J. Hum. Genet. 2011, 88, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Xi, Y.; Gao, M.; Li, Z.; Xu, C.; Fan, S.; He, W. Gene expression profiles of entorhinal cortex in Alzheimer’s disease. Am. J. Alzheimers Dis. 2014, 29, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, R.N.; Levy, O.A.; Wolf, P.; Oliva, P.; Zhang, X.K.; Waters, C.H.; Fahn, S.; Kang, U.J.; Liong, C.; Ford, B.; et al. SCARB2 variants and glucocerebrosidase activity in Parkinson’s disease. Npj Parkinson’s Dis. 2016, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Motaghinejad, M.; Motevalian, M.; Abdollahi, M.; Heidari, M.; Madjd, Z. Topiramate confers neuroprotection against methylphenidate-induced neurodegeneration in dentate gyrus and CA1 regions of hippocampus via CREB/BDNF pathway in rats. Neurotox. Res. 2017, 31, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.J.; Rickmann, M.; Backofen, B.; Browski, S.M.; Rosenbusch, J.; Schöning, S.; Fleischmann, T.; Krieglstein, K.; Mollard, G.F. Lack of the endosomal SNAREs vti1a and vti1b led to significant impairments in neuronal development. Proc. Natl. Acad. Sci. USA 2011, 108, 2575–2580. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; O’Callaghan, N.J.; Fenech, M. Telomere length in white blood cells, buccal cells and brain tissue and its variation with ageing and alzheimer’s disease. Mech. Aging Dev. 2008, 129, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Bjonnes, A.; Prescott, J.; Dib, P.; Natt, P.; Lane, J.; Lerner, M.; Copper, J.A.; Ye, Y.; Li, K.W.; et al. Genome-wide association sudy Identifies variants in casein kinase II (CSNK2A2) to be associated with leukocyte telomere length in a punjabi sikh diabetic cohort. Circ. Cardiovasc. Gene 2014, 7, 287–310. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.Y.; Choi, B.O.; Jeong, J.H.; Kong, K.A.; Hwang, J.; Ahn, J.H. Amyloid beta-mediated hypomethylation of heme oxygenase 1 correlates with cognative impairment in Alzheimer’s disease. PLoS ONE 2016, e0153156. [Google Scholar] [CrossRef]
- Belcredito, S.; Vegeto, E.; Brusadelli, A.; Ghisletti, S.; Mussi, P.; Ciana, P.; Maqqi, A. Estrogen neuroprotection: The involvement of the Bcl-2 binding protein BNIP2. Brain Res. Rev. 2001, 37, 335–342. [Google Scholar] [CrossRef]
- Moon, J.I.; Birren, S.J. Target-dependent inhibition of sympathetic neuron growth via modulation of a BMP signaling pathway. Dev. Biol. 2008, 315, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Javitch, J.A.; D’mato, R.J.; Strittmatter, S.M.; Snyder, S.H. Parkinsonism-inducing neurotoxin, MPTP: Uptake of the metabolite MPP by dopamine neurons explains selective toxicity. Proc. Natl. Acad. Sci. USA 1985, 82, 2173–2177. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.E.; Heales, S.J.R.; Davies, S.E.C.; Boakye, P.; Clark, J.B. Effects of 1-methyl-4-phenylpyridium on isolated rat brain mitochondria: Evidence for a primary involvement of energy depletion. J. Neurochem. 1994, 63, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Kong, Y.; Zhang, H. Oxidative stress, mitochondrial dysfunction, and aging. J. Signal. Transduct. 2012. [CrossRef] [PubMed]
- Perier, C.; Vila, M. Mitochondrial Biology and Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2012, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: The compound N-benzylhexadecanamide (XA) was isolated from Maca and identified by the authors (purity ≥ 95.0%). |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Chemical Compounds | Quant/Qual Ion Pair | Quant/Qual CE(V) |
|---|---|---|---|
| 0 | diazepam | 285→193/285→154 | 37/30 |
| 1 | aspartic acid | 134→88/134→116 | 6/6 |
| 2 | asparagine | 133→87.1/133→74.1 | 6/16 |
| 3 | serine | 106→60.1/106→88.1 | 9/6 |
| 4 | taurine | 126→108/126→44.2 | 9/19 |
| 5 | tyrosine | 182→165.0/182→136.0 | 16/12 |
| 6 | noradrenaline | 170→152.0/170→134.0 | 6/18 |
| 7 | homovanillic acid | 183→137.0/183→137.0 | 14/14 |
| 8 | choline | 104→59.9/104→45.1 | 18/25 |
| 9 | acetyl choline | 146→87.0/146→60.2 | 10/6 |
| 10 | butyrylcholine | 174→115.0/174→71 | 16/20 |
| 11 | ornithine | 133→70/133→116 | 21/6 |
| 12 | lysine | 130→84/130→56 | 15/37 |
| 13 | phenylalanine | 166→120/166→103 | 15/25 |
| 14 | tryptophan | 205→188/205→146 | 10/15 |
| 15 | L-leucine | 132→86/132→30 | 9/12 |
| 16 | methionine | 150→104/150→56 | 12/9 |
| 17 | dimethylglycine | 104→58/104→42 | 15/37 |
| 18 | proline | 116→69.9/116→69.9 | 3/3 |
| 19 | histamine | 112→94.9/112→67.9 | 12/21 |
| 20 | threonine | 120→74/120→101.9 | 6/6 |
| 21 | citrulline | 176→159/176→70 | 6/24 |
| 22 | arginine | 175→70/175→116 | 18/15 |
| 23 | serotonin | 177.1→160/177.1→115.0 | 10/28 |
| 24 | adrenaline | 184→166.0/184→135.1 | 6/16 |
| 25 | dopamine | 154→137.0/154→91 | 10/29 |
| 26 | levodopa | 198→181.0/198→152.0 | 6/19 |
| 27 | γ-aminobutyric acid | 104→87.1/104→69.1 | 9/16 |
| Treatment | Survival Rate (%) |
|---|---|
| Control | 100 |
| 1000 μM MPP+ | 41.34 ± 1.16 |
| 500 μM MPP+ | 50.44 ± 1.05 |
| 250 μM MPP+ | 72.16 ± 2.36 |
| 125 μM MPP+ | 79.62 ± 2.25 |
| 67.5 μM MPP+ | 80.49 ± 2.71 |
| Concentration | Survival Rate (%) |
|---|---|
| Control group | 100 |
| Model group | 49.94 ± 0.26 |
| 200 μM XA | 62.07 ± 1.78 * |
| 100 μM XA | 79.27 ± 2.23 ** |
| 50 μM XA | 54.56 ± 1.35 |
| 25 μM XA | 50.75 ± 2.01 |
| 12.5 μM XA | 54.91 ± 2.28 |
| Analyte | Regression Equation | R2 | LOD (ng/mL) | LOQ (ng/mL) | Linear Range (ng/mL) |
|---|---|---|---|---|---|
| aspartic acid | Y = (0109 × X) + 2.431 × 10−4 | 0.9961 | 2.510 | 10.040 | 10.040–2008 |
| asparagine | Y = (1.087 × X) + 3.066 × 10−4 | 0.9999 | 1.925 | 4.813 | 4.813–3850 |
| serine | Y = (2.239 × X) + 6.326 × 10−3 | 0.9971 | 2.436 | 4.871 | 4.871–1948.5 |
| taurine | Y = (0.137 × X) + 2.844 × 10−4 | 0.9960 | 2.471 | 4.941 | 4.941–1976.5 |
| tyrosine | Y = (1.880 × X) + 2.142 × 10−3 | 0.9988 | 1.988 | 4.971 | 4.971–3976.5 |
| noradrenaline | Y = (2.765 × X) − 7.771 × 10−5 | 0.9995 | 1.065 | 2.663 | 2.663–2130.5 |
| homovanillic acid | Y = (0.610 × X) + 3.685 × 10−4 | 0.9986 | 2.471 | 2.471 | 2.471–1976.5 |
| choline | Y = (10.050 × X) + 3.276 × 10−2 | 0.9972 | 5.711 | 11.423 | 11.423–4569 |
| acetylcholine | Y = (39.80 × X) + 0.040 | 0.9972 | 1.018 | 2.544 | 2.544–2035.5 |
| butyrylcholine | Y = (74.40 × X) + 0.045 | 0.9993 | 0.954 | 2.386 | 2.386–1909 |
| Analyte | S | C | ||||||
|---|---|---|---|---|---|---|---|---|
| K | M | XA50 | XA100 | K | M | XA50 | XA100 | |
| aspartic acid | 2395.355 ± 96.529 | 5510.444 ± 467.377 ** | 14274.770 ± 1087.576 **## | 23712.580 ± 4528.323 *# | 1944.144 ± 109.031 | 1375.344 ± 83.746 ** | 3700.921 ± 185.859 **## | 4927.835 ± 288.260 **## |
| asparagine | 6884.435 ± 1015.190 | 14,272.680 ± 1663.724 * | 31,556.410 ± 3124.580 **## | 40,738.920 ± 2586.551 **## | 10,116.610 ± 1352.500 | 4504.988± 290.293 * | 4213.556 ± 73.306 * | 3824.943 ± 190.385 *# |
| serine | 13,975.260 ± 1087.511 | 39,178.200 ± 5911.520 * | 60,529.650 ± 3594.469 **# | 80,162.380 ± 1577.983 **## | 12,317.940 ± 1561.040 | 6873.225 ± 1076.321 ** | 5501.029 ± 18.212 * | 5555.612 ± 229.131 * |
| taurine | 471.331 ± 99.991 | 142.098 ± 10.374 * | 432.399 ± 71.048 # | 1105.402 ± 156.281 *## | 20,115.850 ± 2635.606 | 9444.964 ± 807.414 * | 12,097.300 ± 261.632 *# | 17,170.847 ± 831.887 *## |
| tyrosine | 236.723 ± 8.283 | 487.634 ± 6.951 ** | 591.228 ± 29.906 **# | 707.278 ± 67.708 **# | 708.870 ± 81.875 | 294.214 ± 34.469 ** | 509.265 ± 33.050 # | 395.586 ± 29.330 * |
| noradrenaline | 118.122 ± 6.944 | 113.990 ± 4.223 | 125.690 ± 4.836 | 136.567 ± 3.149 # | 90.074 ± 6.222 | 66.727 ± 2.370 * | 68.401 ± 1.072 * | 69.707 ± 1.770 *# |
| homovanillic acid | 320.472 ± 9.119 | 235.988 ± 10.898 ** | 317.552 ± 20.965 **# | 396.112 ± 31.864 **# | 480.939 ± 84.673 | 205.297 ± 22.389 * | 271.848 ± 15.054 # | 461.287 ± 35.156 # |
| choline | 115.893 ± 21.214 | 61.222 ± 7.910 * | 254.697 ± 5.314 *## | 266.724 ± 9.861 **## | 6.203 ± 1.699 | 5.953 ± 0.353 * | - | - |
| acetylcholine | 423.041 ± 64.848 | 227.654 ± 101.324 * | 630.147 ± 44.006 ## | 1108.037± 101.248 **## | 1600.918 ± 143.872 | 380.128 ± 31.322 ** | 440.118 ± 10.453 ** | 582.330 ± 5.916 **# |
| butyrylcholine | - | - | 4.309 ± 0.752 **## | 7.731 ± 0.434 **## | 9.904 ± 0.761 | 8.152 ± 0.566 | 10.206 ± 0.807 | 28.001 ± 2.426 **## |
| Analyte | S | C | ||||||
|---|---|---|---|---|---|---|---|---|
| K | M | XA50 | XA100 | K | M | XA50 | XA100 | |
| ornithine | 600.781 ± 100.501 | 1517.943 ± 117.101 ** | 2062.528 ± 122.675 **## | 1548.961 ± 48.987 ** | 32.226 ± 3.387 | 21.717 ± 0.813 ** | 19.457 ± 0.142 **## | 18.397 ± 1.329 **# |
| lysine | 27.421 ± 4.076 | 14.576 ± 0.558 ** | 11.510 ± 0.050 **# | 11.346 ± 0.043 **## | 0.051 ± 2.57 × 10−4 | 0.041 ± 5.78 × 10−3 * | 0.050 ± 7.93 × 10−3 | 0.043 ± 6.83 × 10−3 |
| phenylalanine | 21.522 ± 4.642 | 90.387 ± 6.830 ** | 130.374 ± 12.928 **## | 111.406 ± 7.036 **## | 7.909 ± 2.583 | 5.551 ± 0.305 | 4.707 ± 0.193 # | 4.252 ± 0.259 ## |
| tryptophan | 0.474 ± 6.62×10−2 | 0.258 ± 7.24 × 10−3 ** | 0.289 ± 4.87 × 10−2 * | 0.198 ± 2.97 × 10−3 **## | 6.70 × 10−2 ± 6.25 × 10−3 | 3.30×10−2 ± 2.33 × 10−3 ** | 2.65×10−2 ± 1.81 × 10−3 **# | 3.26×10−2 ± 2.17 × 10−3 ** |
| L-leucine | 63.876 ± 10.329 | 733.788 ± 219.994 ** | 1042.663 ± 180.334 ** | 857.797 ± 37.087 ** | 94.712 ± 4.736 | 86.116 ± 1.129 * | 79.111 ± 1.922 **## | 70.187 ± 8.315 *# |
| methionine | 0.784 ± 0.129 | 3.676 ± 0.805 ** | 7.124 ± 0.925 **## | 5.232 ± 0.601 ** | 0.388 ± 0.048 | 0.289 ± 0.016 * | 0.300 ± 0.013 * | 0.246 ± 0.028 * |
| dimethylglycine | 4.397 ± 0.655 | 9.808 ± 1.845 ** | 12.696 ± 1.353 ** | 12.398 ± 0.745 ** | 1.505 ± 0.003 | 1.400 ± 0.091 | 1.497 ± 0.057 | 1.490 ± 0.086 |
| proline | 34.412 ± 2.978 | 26.567 ± 3.802 | 55.397 ± 6.297 | 62.614 ± 2.243 | 17.092 ± 2.391 | 5.750 ± 0.444 | 6.133 ± 0.622 | 8.218 ± 0.561 |
| histamine | 0.192 ± 2.94 × 10−2 | 0.174 ± 3.80 ×10−3 | 0.256 ± 1.42 × 10−2*## | 0.281 ± 1.73×10−2 # | 1.91×10−2 ± 7.11 × 10−3 | 0.92×10−2± 1.14 × 10−3 | 1.09×10−2 ± 1.75 × 10−3 | 1.09×10−2 ± 5.62 × 10−4 |
| threonine | 13.792 ± 1.807 | 42.426 ± 0.420 ** | 58.133 ± 2.274 **## | 42.037 ± 4.344 ** | 7.283 ± 2.135 | 3.710 ± 0.640 * | 3.766 ± 8.74×10−2 * | 4.032 ± 0.281 |
| citrulline | 52.390 ± 4.242 | 163.451 ± 23.996 ** | 238.912 ± 31.804 **# | 224.588 ± 20.786 **# | 6.605 ± 1.205 | 2.862 ± 0.056 ** | 3.027 ± 0.085 **# | 2.853 ± 0.165 ** |
| arginine | 6.172 ± 0.028 | 8.938 ± 0.888 ** | 12.233 ± 0.267 ** | 11.151 ± 0.956 **# | 0.380 ± 0.060 | 0.888 ± 0.021 * | 0.267 ± 0.021 * | 0.252 ± 0.03 * |
| serotonin | 4.91 × 10−3 ± 7.27 × 10−4 | 7.14 × 10−4 ± 8.65 × 10−5 ** | 1.20 ×10−3± 2.67 × 10−4* | 4.89×10−3 ± 4.33 × 10−4 ## | 1.25×10−3 ± 3.16 × 10−4 | 9.70×10−4 ± 7.74 × 10−5 | 1.15×10−3 ± 1.61 × 10−5 | 1.29×10−3 ± 1.01 × 10−4 |
| adrenaline | 4.57 × 10−3 ± 7.95e × 10−4 | 1.24 × 10−3 ± 1.43e × 10−4 * | 1.36×10−3 ± 1.02 × 10−4 * | 1.82×10−3 ± 9.45 × 10−5 *## | 8.65×10−4 ± 8.35 × 10−5 | 4.37×10−4 ± 5.54 × 10−5** | 4.90×10−4 ± 5.25 × 10−5 ** | 1.25×10−3 ± 1.76×10−4 *## |
| dopamine | 2.40 × 10−3 ± 2.68e × 10−4 | 2.07 × 10−3 ± 7.92 × 10−5 | 2.68×10−3 ± 1.67 × 10−4# | 1.44×10−3 ± 9.89 × 10−5 *# | 3.41×10−3 ± 7.10 × 10−4 | 1.71×10−4 ± 1.71 × 10−4 * | 2.70×10−3 ± 6.64 × 10−4 | 1.10×10−3 ± 7.37 × 10−5 * |
| levodopa | 1.41 × 10−3 ± 1.42 × 10−4 | 1.17 × 10−3 ± 2.85 × 10−4 | 1.77×10−3 ± 1.97 × 10−4 | 1.84×10−3 ± 1.69 × 10−4 | 1.63×10−3 ± 2.31 × 10−4 | 5.80×10−4± 8.57 × 10−5 * | 1.14×10−3 ± 8.99 × 10−5 ## | 1.43×10−3 ± 6.93×10−5 *## |
| γ-aminobutyric acid | 2.1 × 10−2 ± 4.22 × 10−3 | 2.83 × 10−3 ± 1.70 × 10−3 * | 3.30×10−3 ± 5.53 × 10−4 * | 3.23×10−3 ± 1.52 × 10−4 * | 7.47×10−4 ± 1.52 × 10−4 | 6.14×10−4 ± 1.90 × 10−5 | 7.81×10−4 ± 1.03 × 10−4 | 7.40×10−4 ± 8.50 × 10−4 |
| No. | Potential Biomarkers | tR/Min | m/z | Formula | VIP | Type | Change Multiplier (M/K) | Change Multiplier (XA/M) | Post-Model Trend | |
|---|---|---|---|---|---|---|---|---|---|---|
| K-M | XA-M | |||||||||
| L1 | PC(O-18:1(9Z)/2:0) | 2.82 | 550.3870 | C32H64NO7P | 1.31 | 1.57 | PC | 0.46 | 2.07 | ↓ |
| L2 | PC (0:0/18:0) | 2.70 | 524.3714 | C26H54NO7P | 4.76 | 4.47 | PC | 0.48 | 1.67 | ↓ |
| L3 | PC (24:0/0:0) | 6.59 | 608.4654 | C32H66NO7P | 2.29 | 1.74 | PC | 0.50 | 1.46 | ↓ |
| L4 | PC(P-18:0/0:0) | 3.08 | 508.3761 | C26H54NO6P | 3.47 | 1.27 | PC | 0.79 | 1.87 | ↓ |
| L5 | PC (18:2(9Z,12Z)/22:2(13Z,16Z)) | 12.39 | 860.6162 | C48H88NO8P | 1.73 | 2.05 | PC | 1.57 | 0.66 | ↑ |
| L6 | PC (P-16:0/22:4(7Z,10Z,13Z,16Z)) | 10.64 | 816.5897 | C46H84NO7P | 2.17 | 2.09 | PC | 2.13 | 0.63 | ↑ |
| L7 | PC (18:3(6Z,9Z,12Z)/22:2(13Z,16Z)) | 10.02 | 858.6007 | C48H86NO8P | 1.01 | 1.14 | PC | 2.77 | 0.42 | ↑ |
| L8 | PC (16:1(9Z)/22:4(7Z,10Z,13Z,16Z)) | 7.50 | 830.5687 | C46H82NO8P | 1.59 | 1.76 | PC | 1.57 | 0.67 | ↑ |
| L9 | PC (18:1(9Z)/22:4(7Z,10Z,13Z,16Z)) | 10.44 | 858.6012 | C46H86NO8P | 1.93 | 1.37 | PC | 1.84 | 0.38 | ↑ |
| L10 | PC (20:0/22:4(7Z,10Z,13Z,16Z)) | 13.33 | 888.6476 | C50H92NO8P | 2.28 | 1.20 | PC | 3.82 | 0.70 | ↑ |
| L11 | LysoPC (24:1(15Z)) | 4.87 | 606.4498 | C32H64NO7P | 1.53 | 1.18 | LysoPC | 0.46 | 2.70 | ↓ |
| L12 | LysoPC (O-18:0) | 3.63 | 552.4031 | C26H56NO6P | 1.40 | 2.03 | LysoPC | 0.50 | 1.70 | ↓ |
| L13 | PE (P-16:0/0:0) | 2.31 | 438.2983 | C21H44NO6P | 1.10 | 1.04 | PE | 0.41 | 1.86 | ↓ |
| L14 | PE (20:0/22:4(7Z,10Z,13Z,16Z)) | 13.21 | 824.6181 | C47H86NO8P | 2.60 | 2.21 | PE | 1.67 | 0.71 | ↑ |
| L15 | LacCer (d18:1/24:0) | 14.70 | 974.7503 | C54H103NO13 | 1.54 | 3.79 | Cer | 0.62 | 3.23 | ↓ |
| L16 | Trihexosylceramide (d18:1/24:0) | 14.51 | 1136.8030 | C60H113NO18 | 2.11 | 1.80 | Cer | 0.59 | 3.99 | ↓ |
| L17 | LacCer (d18:1/22:0) | 14.16 | 946.7188 | C52H99NO13 | 2.00 | 1.64 | Cer | 0.45 | 3.62 | ↓ |
| L18 | LacCer (d18:1/24:1(15Z)) | 14.14 | 972.7347 | C54H101NO13 | 1.40 | 1.72 | Cer | 0.50 | 3.98 | ↓ |
| L19 | DG (16:0/18:1(9Z)/0:0) | 13.07 | 603.5351 | C37H70O5 | 1.58 | 1.55 | DG | 1.82 | 0.67 | ↑ |
| L20 | Coenzyme Q8 | 15.33 | 727.5657 | C49H74O4 | 1.31 | 1.08 | Other | 1.46 | 0.69 | ↑ |
| L21 | Montecristin | 10.41 | 575.5036 | C37H66O4 | 1.43 | 1.51 | Other | 1.62 | 0.68 | ↑ |
| L22 | Calcitriol | 18.31 | 369.3520 | C27H44O3 | 3.76 | 2.44 | Other | 0.44 | 1.45 | ↓ |
| NO. | Protein Name | Description | Ratio | Post-Model Trend | ||
|---|---|---|---|---|---|---|
| M/K | XA50/M | XA100/M | ||||
| 1 | Aldoc | Fructose-bisphosphate aldolase C | 0.68 | 2.30 | 2.33 | ↓ |
| 2 | Dhfr | Dihydrofolate reductase | 0.42 | 2.19 | 2.49 | ↓ |
| 3 | Atp5d | ATP synthase subunit delta, mitochondrial | 0.32 | 2.88 | 3.04 | ↓ |
| 4 | Scarb2 | Lysosome membrane protein 2 | 0.063 | 2.12 | 2.63 | ↓ |
| 5 | Ak1 | Adenylate kinase isoenzyme 1 | 0.13 | 1.62 | 2.23 | ↓ |
| 6 | Vti1b | Vesicle transport through interaction with t-SNAREs homolog 1B | 0 | - | - | ↓ |
| 7 | Csnk2a2 | Casein kinase II subunit alpha’ | 1.92 | 0.66 | 0.54 | ↑ |
| 8 | Hmox1 | Heme oxygenase 1 | - | 0.34 | 0 | ↑ |
| 9 | Bnip2 | BCL2/adenovirus E1B 19 kDa protein-interacting protein 2 | 1.89 | 0 | 0 | ↑ |
| 10 | Mrpl50 | 39S ribosomal protein L50, mitochondrial | - | 0 | 0 | ↑ |
| 11 | Gla | Alpha-galactosidase A | - | 0.69 | 0.61 | ↑ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Wang, H.; Guo, F.; Si, N.; Brantner, A.; Yang, J.; Han, L.; Wei, X.; Zhao, H.; Bian, B. Integrated Proteomics and Lipidomics Investigation of the Mechanism Underlying the Neuroprotective Effect of N-benzylhexadecanamide. Molecules 2018, 23, 2929. https://doi.org/10.3390/molecules23112929
Zhou Y, Wang H, Guo F, Si N, Brantner A, Yang J, Han L, Wei X, Zhao H, Bian B. Integrated Proteomics and Lipidomics Investigation of the Mechanism Underlying the Neuroprotective Effect of N-benzylhexadecanamide. Molecules. 2018; 23(11):2929. https://doi.org/10.3390/molecules23112929
Chicago/Turabian StyleZhou, Yanyan, Hongjie Wang, Feifei Guo, Nan Si, Adelheid Brantner, Jian Yang, Lingyu Han, Xiaolu Wei, Haiyu Zhao, and Baolin Bian. 2018. "Integrated Proteomics and Lipidomics Investigation of the Mechanism Underlying the Neuroprotective Effect of N-benzylhexadecanamide" Molecules 23, no. 11: 2929. https://doi.org/10.3390/molecules23112929
APA StyleZhou, Y., Wang, H., Guo, F., Si, N., Brantner, A., Yang, J., Han, L., Wei, X., Zhao, H., & Bian, B. (2018). Integrated Proteomics and Lipidomics Investigation of the Mechanism Underlying the Neuroprotective Effect of N-benzylhexadecanamide. Molecules, 23(11), 2929. https://doi.org/10.3390/molecules23112929