Comparison of Chemical Profiles, Anti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Synthetic Borneol and Natural Borneol


Abstract
1. Introduction
2. Results
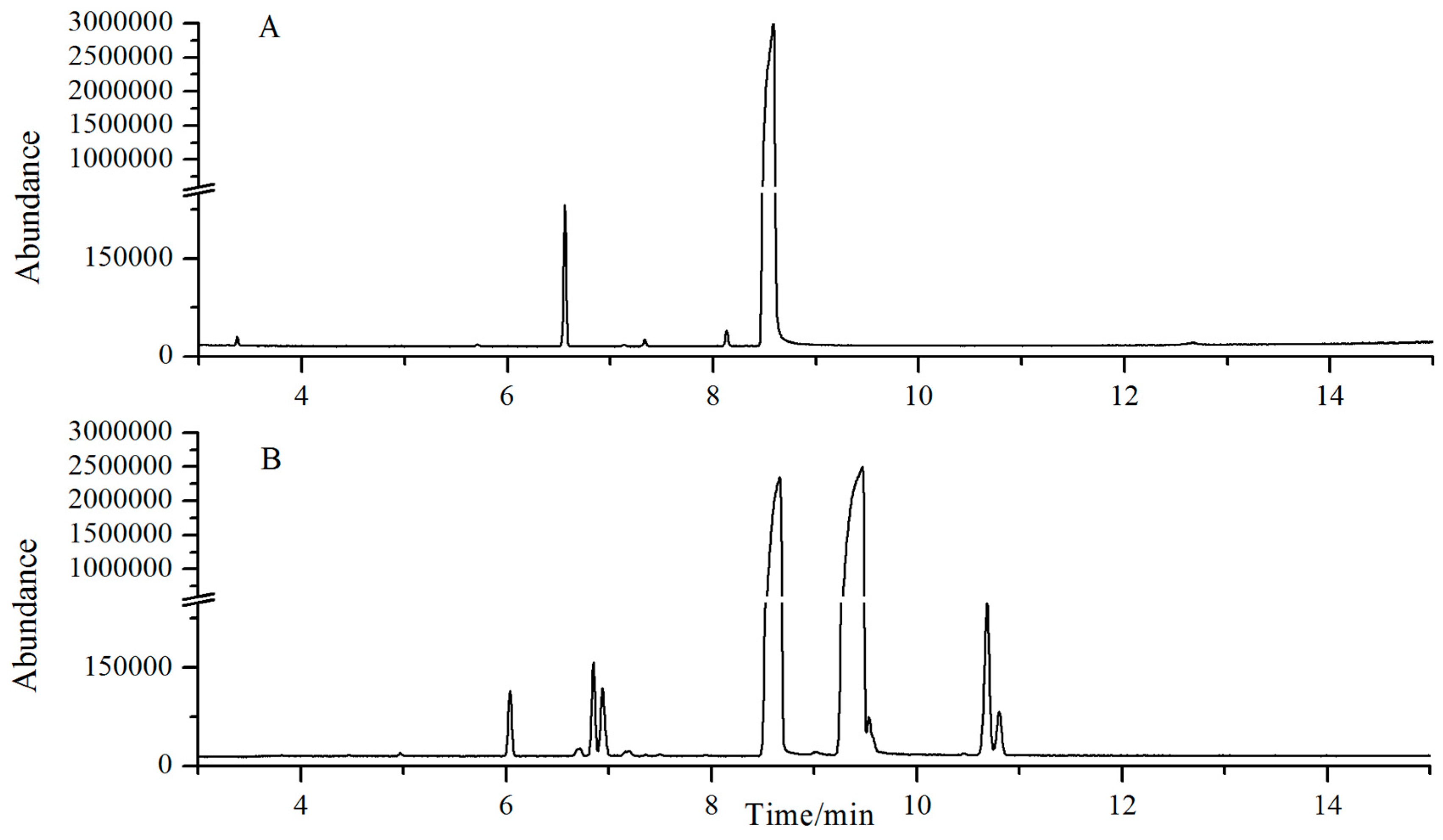
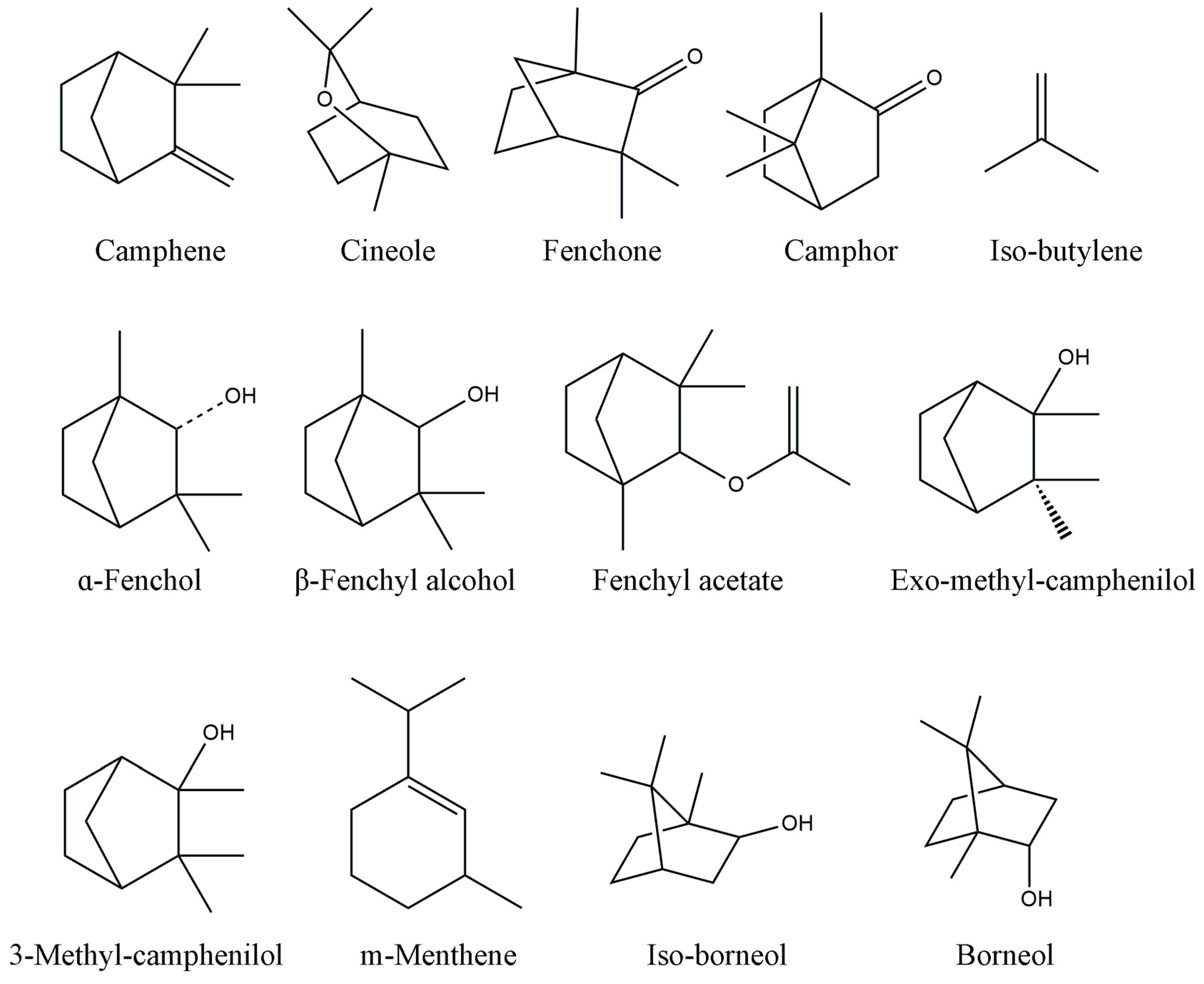
2.1. Chemical Profiles of SB and NB by GC-MS Analysis
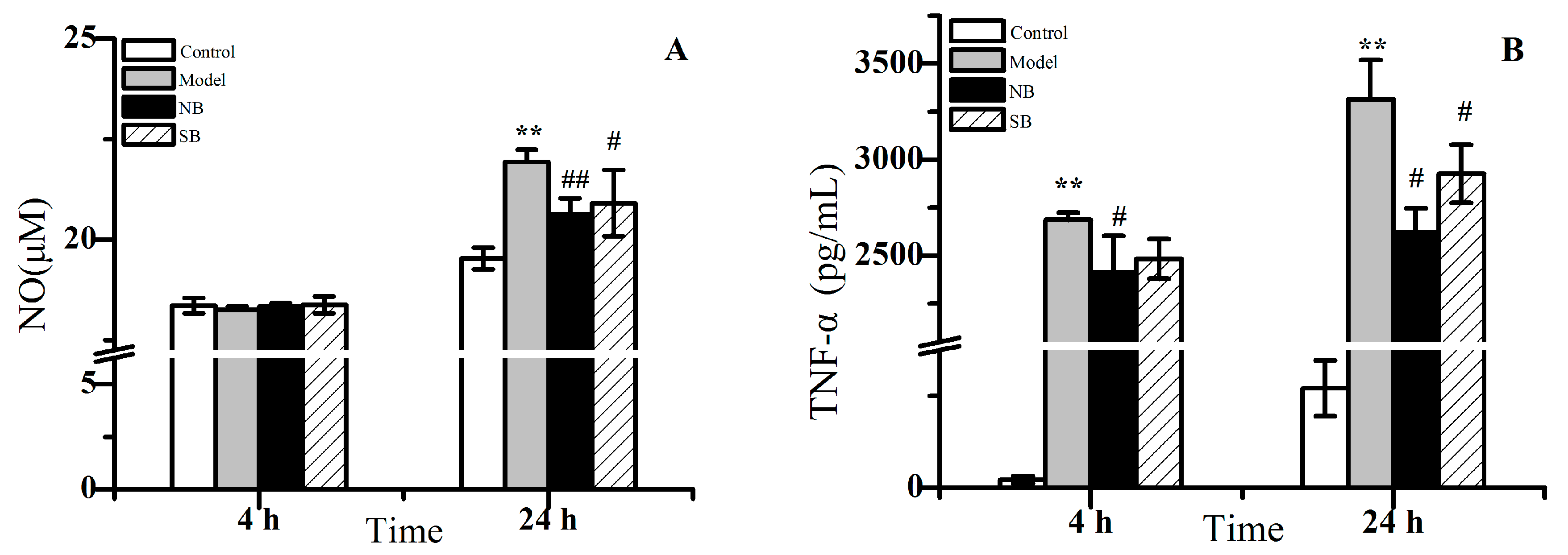
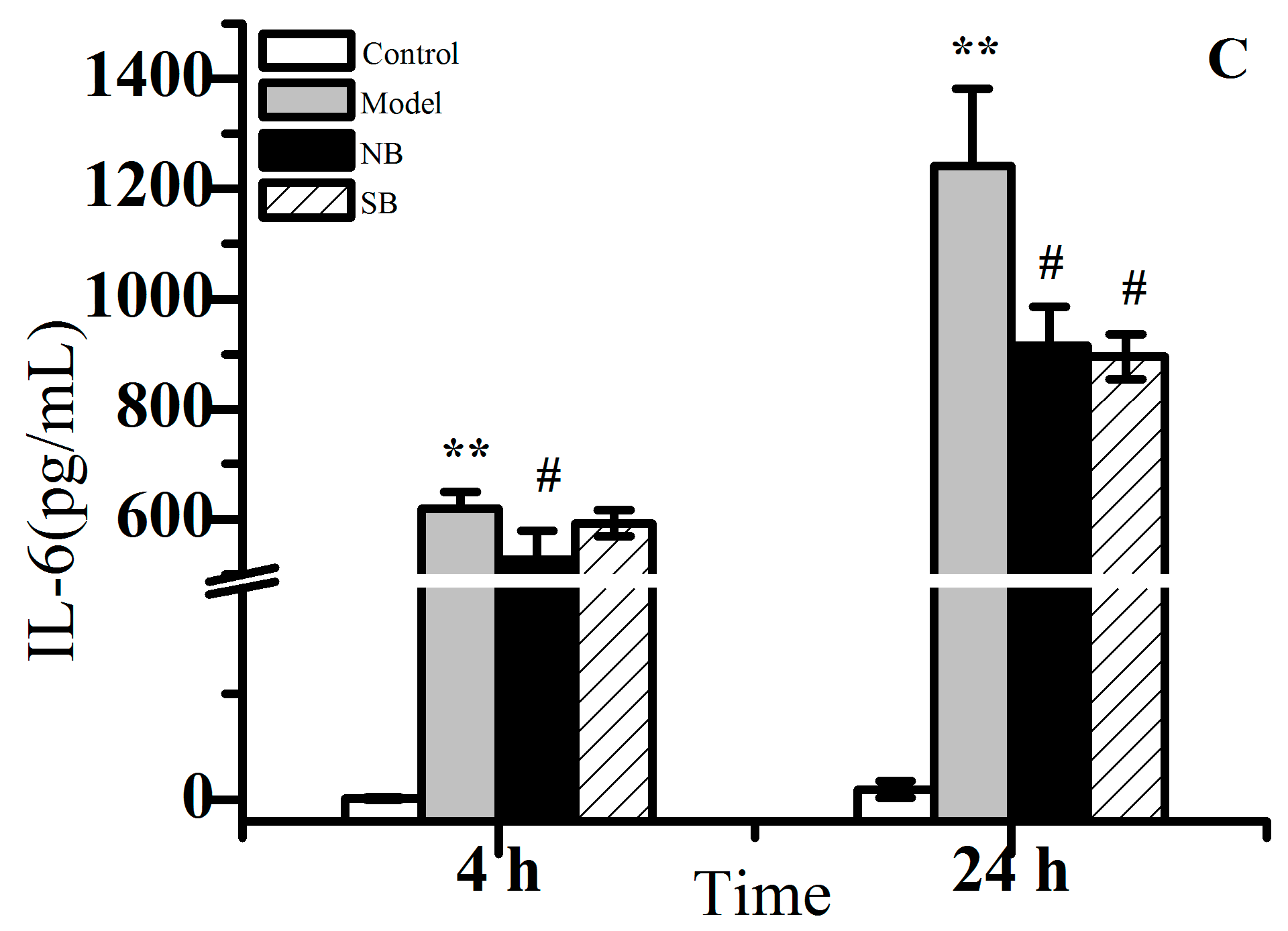
2.2. Anti-Inflammation Activity In Vitro
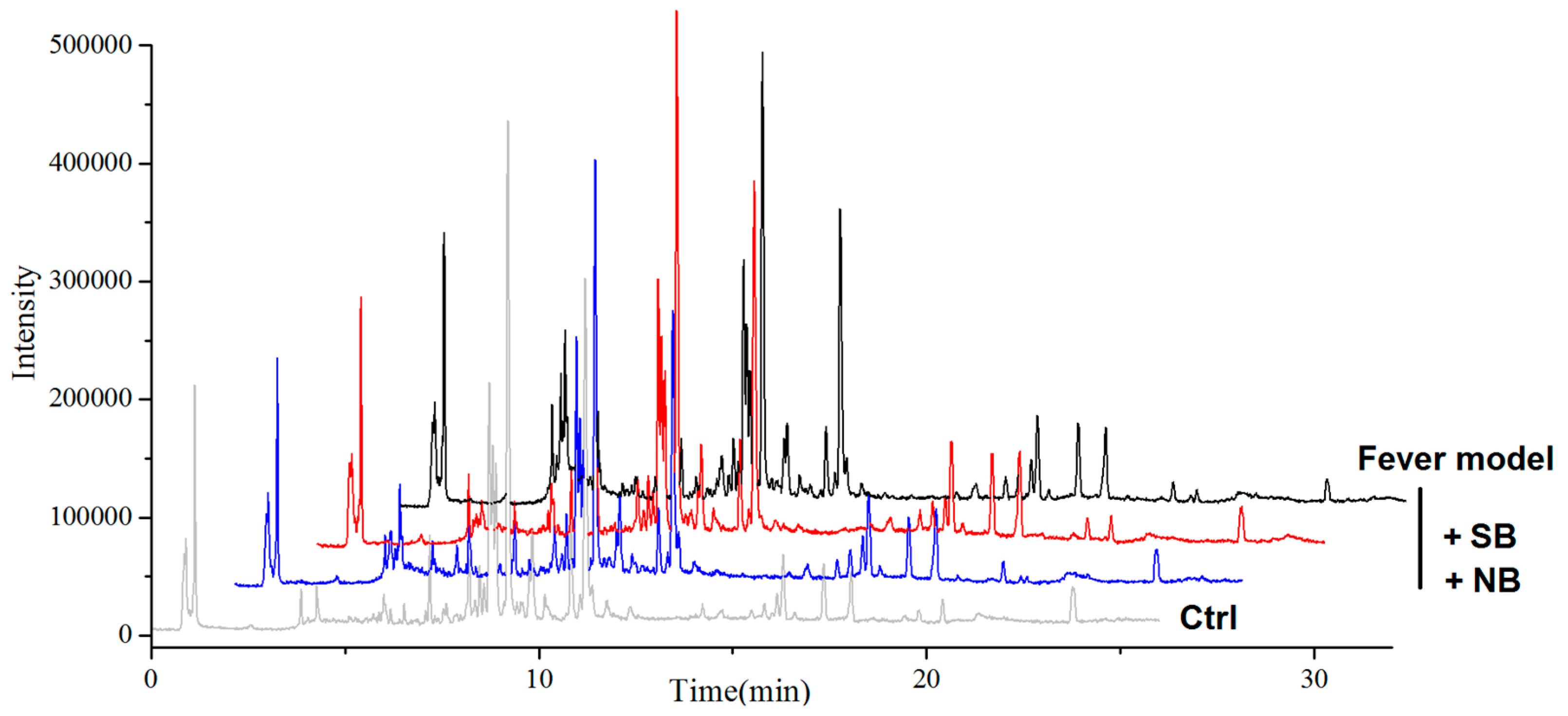
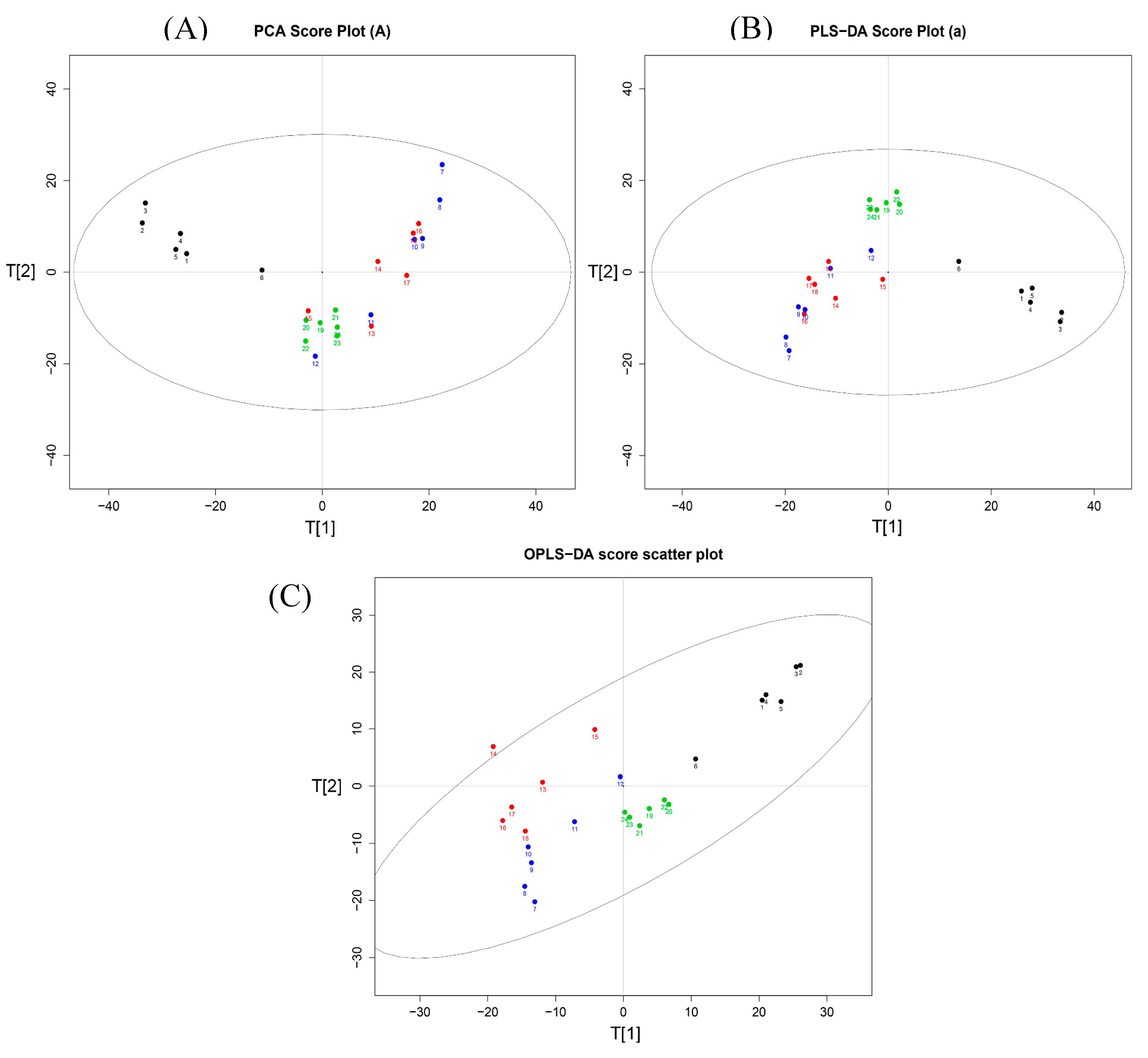
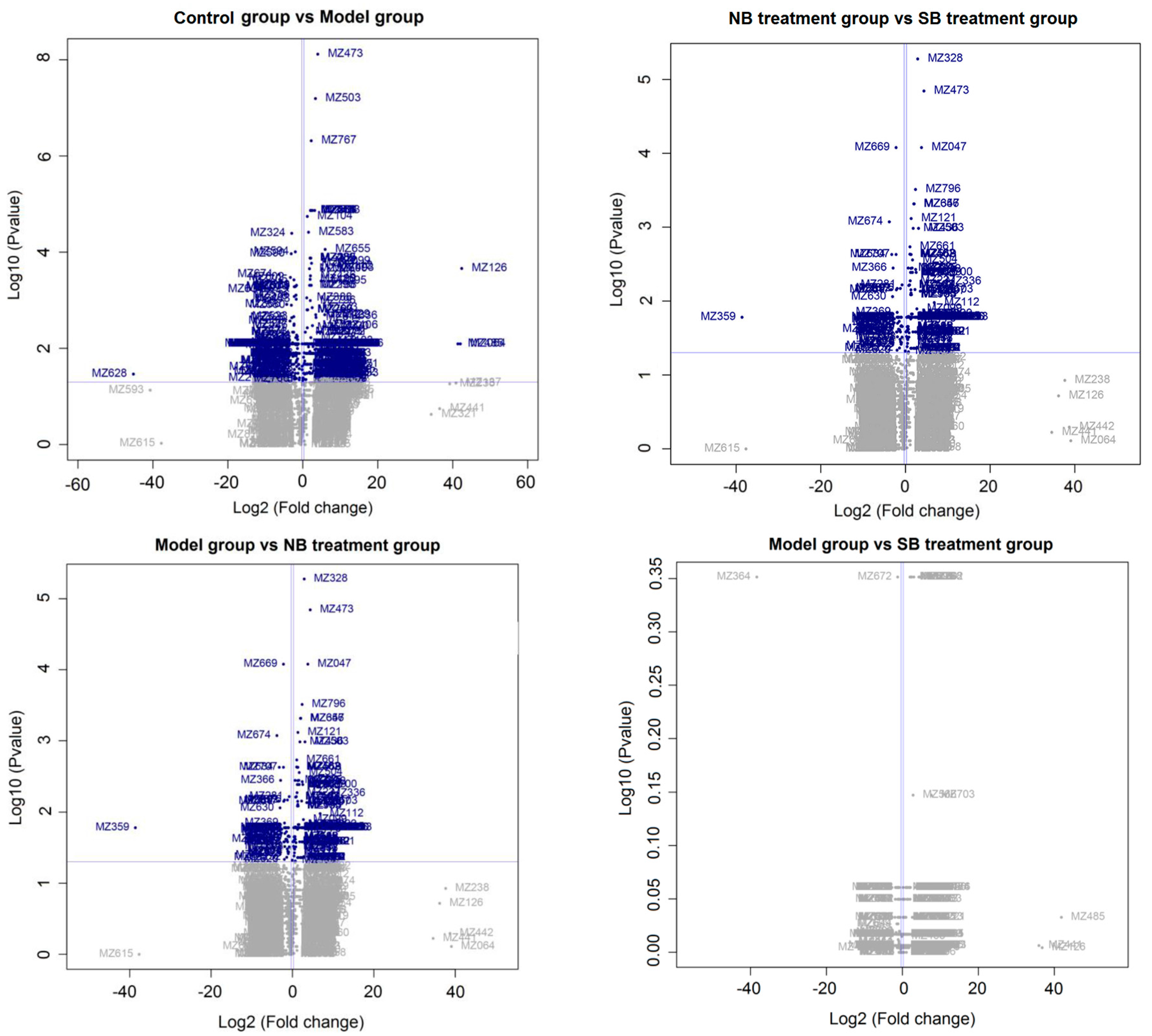
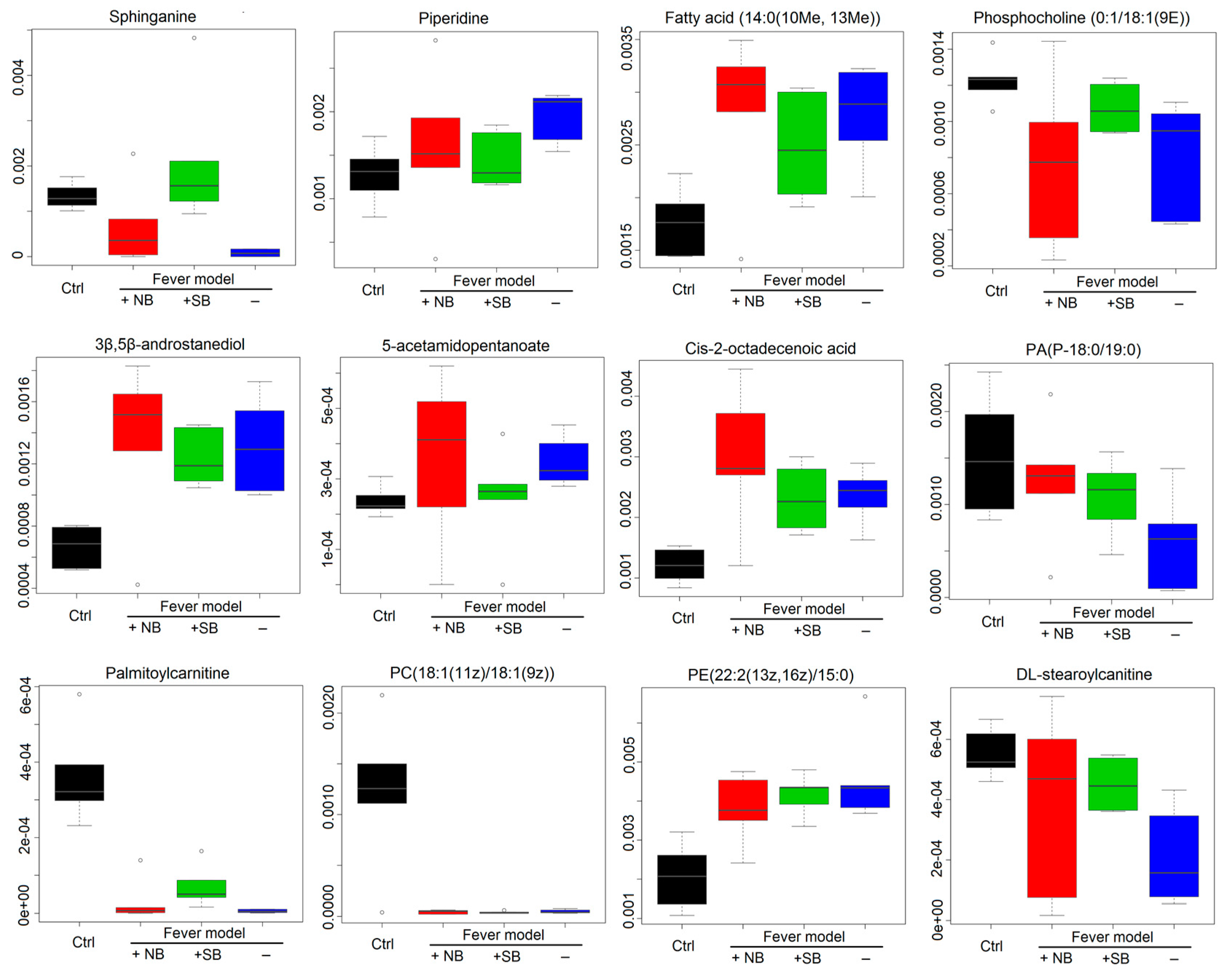
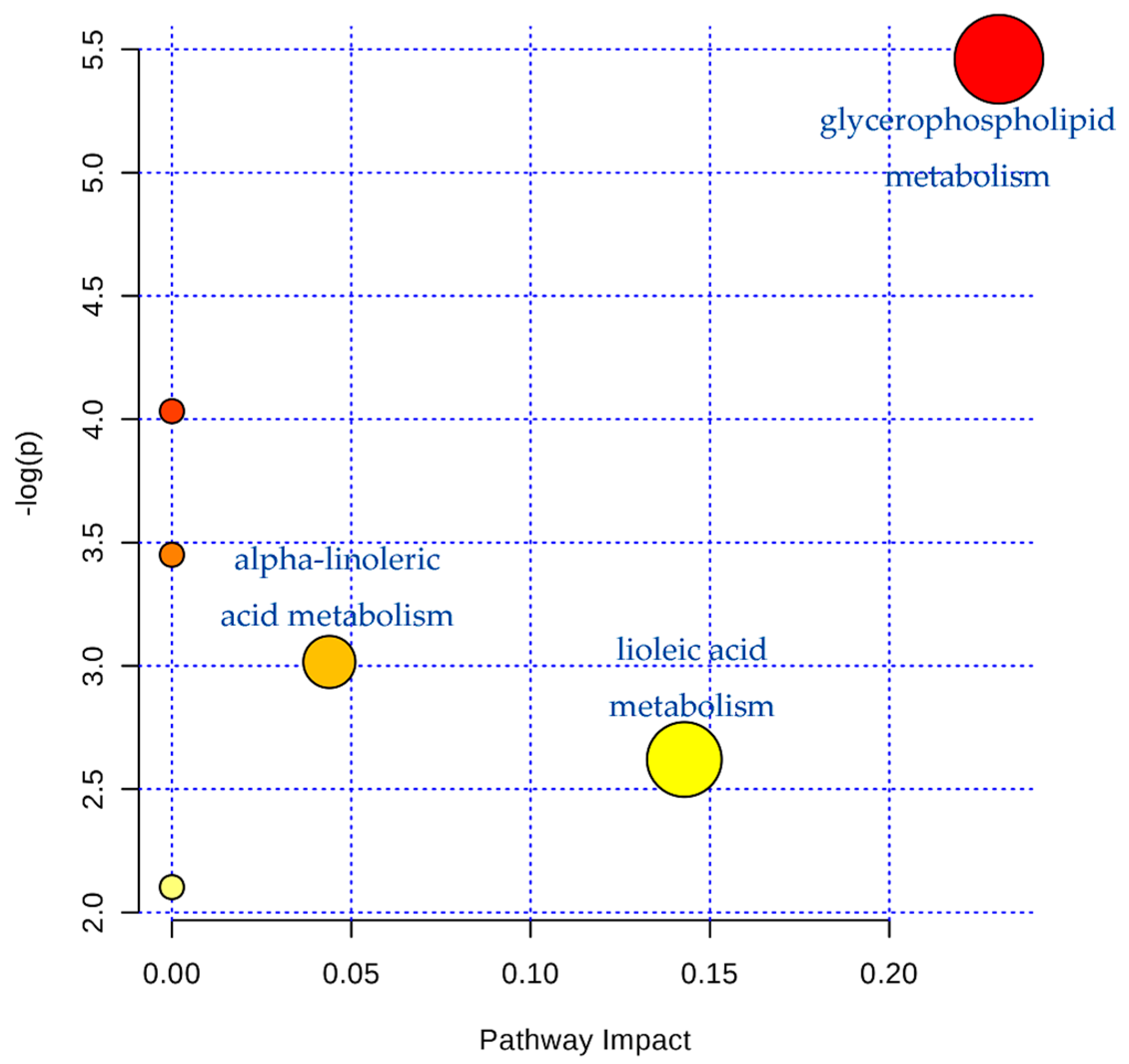
2.3. Serum Metabonomics of NB and SB in Endotoxic Fever Rats
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Chemical Profile by GC-MS Analysis
4.3. In Vitro Anti-Inflammation Effects on LPS-Induced RAW 264.7 Cells
4.4. Animal Handling for Endotoxic Fever Rat Model
4.5. Serum Pretreatment
4.6. UPLC-Q-TOF/MS Condition
4.7. Identification of Potential Biomarkers
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- State Pharmacopoeia Commission. Chinese Pharmacopoeia; Chinese Medical Science and Technology Press: Beijing, China, 2015; pp. 260–261. ISBN 9787506773379.
- Zhang, T.M. Clinical Chinese Pharmacy, 2nd ed.; Shanghai Scientific and Technical Press: Shanghai, China, 2012; pp. 308–309. ISBN 9787547813188. [Google Scholar]
- Liu, R.; Zhang, L.; Lan, X.; Li, L.; Zhang, T.T.; Sun, J.H.; Du, G.H. Protection by borneol on cortical neurons against oxygen-glucose deprivation/reperfusion: Involvement of anti-oxidation and anti-inflammation through nuclear transcription factor κappaB signaling pathway. Neuroscience 2011, 176, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A. Camphor in the Edo era-camphor and borneol for medicines. Yakushigaku Zasshi 2000, 35, 49–54. [Google Scholar] [PubMed]
- Kong, Q.X.; Wu, Z.Y.; Chu, X.; Liang, R.Q.; Xia, M.; Li, L. Study on the anti-cerebral ischemia effect of borneol and its mechanism. Afr. J. Tradit. Complement. Altern. Med. 2013, 11, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Cui, Y.; Yu, Q.; Xie, X.; Liu, Y.; Wei, M.; Ci, X.; Peng, L. Modulation of LPS-stimulated pulmonary inflammation by borneol in murine acute lung injury model. Inflammation 2014, 37, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Du, S.; Bai, J.; Li, P.; Wen, R.; Zhao, X. Bioavailability and brain-targeting of geniposide in gardenia-borneol co-compound by different administration routes in mice. Int. J. Mol. Sci. 2012, 13, 14127–14135. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gong, X.; Lu, Y.; Du, S.; Yang, Z.; Bai, J.; Li, P.; Wu, H. Enhancing effect of borneol and muscone on geniposide transport across the human nasal epithelial cell monolayer. PLoS ONE 2014, 9, e101414. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Cao, L.; Ge, H.; Duanmu, W.; Tan, L.; Yuan, J.; Tunan, C.; Li, F.; Hu, R.; Gao, F.; Feng, H. L-Borneol induces transient opening of the blood-brain barrier and enhances the therapeutic effect of cisplatin. Neuroreport 2017, 28, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Liu, X.W.; Du, F.F.; Li, M.J.; Xu, F.; Wang, F.Q.; Liu, Y.; Li, C.; Sun, Y. Sensitive assay for measurement of volatile borneol, isoborneol, and the metabolite camphor in rat pharmacokinetic study of Borneolum (Bingpian) and Borneolum syntheticum (synthetic Bingpian). Acta Pharmacol. Sin. 2013, 34, 1337–1348. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Du, S.; Yao, Z.; Zhao, P.; Zhai, Y. Study on natural borneol and synthetic borneol affecting mucosal permeability of gardenia extract. China J. Chin. Mater. Med. 2009, 34, 1207–1210. [Google Scholar]
- Luo, D.D.; Chen, X.Y.; Zhang, Z.B.; Sun, C.Y.; Zheng, Y.F.; Liu, Y.H.; Wang, X.F.; Wang, Q.; Zhan, J.Y.; Su, Z.R. Different effects of (+)‑borneol and (−)‑borneol on the pharmacokinetics of osthole in rats following oral administration. Mol. Med. Rep. 2017, 15, 4239–4246. [Google Scholar] [CrossRef] [PubMed]
- Lao, Y.M.; Jiang, J.G.; Yan, L. Application of metabonomic analytical techniques in the modernization and toxicology research of traditional Chinese medicine. Br. J. Pharmacol. 2009, 157, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Wrotek, S.; Jędrzejewski, T.; Nowakowska, A.; Kozak, W. Glutathione deficiency attenuates endotoxic fever in rats. Int. J. Hyperth. 2015, 31, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Yang, J.; Huang, Y.; Zhang, K.; Bo, Y.; Lu, X.; Su, G.; Ma, J.; Yang, J.; Zhao, L.; Wu, C. Serum metabonomics study of anti-depressive effect of Xiao-Chai-Hu-Tang on rat model of chronic unpredictable mild stress. J. Chromatogr. B 2016, 1029, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Feng, G.; Zhao, B.; Zhao, J.; Pi, Z.; Liu, S.; Song, F.; Liu, Z. A non-target urinary and serum metabolomics strategy reveals therapeutical mechanism of Radix Astragali on adjuvant-induced arthritis rats. J. Chromatogr. B 2017, 1048, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Cauwels, A. Nitric oxide in shock. Kidney Int. 2007, 72, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Temiz-Resitoglu, M.; Kucukkavruk, S.P.; Guden, D.S.; Cecen, P.; Sari, A.N.; Tunctan, B.; Gorur, A.; Tamer-Gumus, L.; Buharalioglu, C.K.; Malik, K.U.; et al. Activation of mTOR/IκB-α/NF-κB pathway contributes to LPS-induced hypotension and inflammation in rats. Eur. J. Pharmacol. 2017, 802, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Mollace, V.; Muscoli, C.; Masini, E.; Cuzzocrea, S.; Salvemini, D. Modulation of prostaglandin biosynthesis by nitric oxide and nitric oxide donors. Pharmacol. Rev. 2005, 57, 217–252. [Google Scholar] [CrossRef] [PubMed]
- Puhr, M.; De Marzo, A.; Isaacs, W.; Lucia, M.S.; Sfanos, K.; Yegnasubramanian, S.; Culig, Z. Inflammation, Microbiota, and Prostate Cancer. Eur. Urol. Focus 2016, 2, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lee, M.K.; Lim, S.Y.; Sung, S.H.; Kim, Y.C. Inhibition of inducible NO synthase, cyclooxygenase-2 and interleukin-1beta by torilin is mediated by mitogenactivated protein kinases in microglial BV2 cells. Br. J. Pharmacol. 2009, 156, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Unuma, K.; Aki, T.; Noritake, K.; Funakoshi, T.; Uemura, K. A CO-releasing molecule prevents annexin A2 down-regulation and associated disorders in LPS-administered rat lung. Biochem. Biophys. Res. Commun. 2017, 487, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Lin, M.W.; Korivi, M.; Wu, J.J.; Liao, C.H.; Chang, C.T.; Liao, J.W.; Hseu, Y.C. Coenzyme Q0 regulates NFκB/AP-1 activation and enhances Nrf2 stabilization in attenuation of LPS-induced inflammation and redox imbalance: Evidence from in vitro and in vivo studies. Biochim. Biophys. Acta 2016, 1859, 246–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ding, H.W.; Li, H.D.; Huang, H.M.; Li, X.F.; Yang, Y.; Zhang, Y.L.; Pan, X.Y.; Huang, C.; Meng, X.M.; Li, J. Hesperetin derivative-14 alleviates inflammation by activating PPAR-γ in mice with CCl4-induced acute liver injury and LPS-treated RAW 264.7 cells. Toxicol. Lett. 2017, 274, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Kim, S.S.; Oh, T.H.; Lee, N.H.; Hyun, C.G. Abies koreana Essential Oil Inhibits Drug-Resistant Skin Pathogen Growth and LPS-Induced Inflammatory Effects of Murine Macrophage. Lipids 2009, 44, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, R.M.C.; Leite, F.C.; Leite, J.A.; Mascarenhas, S.R.; Rodrigues, L.C.; Piuvezam, M.R. Synthesis, acute toxicity and anti-inflammatory effect of bornyl salicylate, a salicylic acid derivative. Immunopharmacol. Immunotoxicol. 2012, 34, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Ocaña, A.; Reglero, G. Effects of Thyme Extract Oils (from Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on Cytokine Production and Gene Expression of oxLDL-Stimulated THP-1-Macrophages. J. Obes. 2012, 2012, e104706. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shen, Y.Y.; Li, J.; Lin, Y.H.; Luo, C.X.; Zhu, D.Y. (+)-Borneol alleviates mechanical hyperalgesia in models of chronic inflammatory and neuropathic pain in mice. Eur. J. Pharmacol. 2015, 757, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Gao, Y.; Chen, J.; Huang, Y.; Zhang, Z.; Xu, F. Time-resolved metabolomics analysis of individual differences during the early stage of lipopolysaccharide-treated rats. Sci. Rep. 2016, 6, e34136. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Misra, V.; Holman, A.; Kamat, A.; Morgello, S.; Gabuzda, D. Plasma metabolomics identifies lipid abnormalities linked to markers of inflammation, microbial translocation, and hepatic function in HIV patients receiving protease inhibitors. BMC Infect. Dis. 2013, 13, e203. [Google Scholar] [CrossRef] [PubMed]
- Petropoulou, P.I.; Berbée, J.F.; Theodoropoulos, V.; Hatziri, A.; Stamou, P.; Karavia, E.A.; Spyridonidis, A.; Karagiannides, I.; Kypreos, K.E. Lack of LCAT reduces the LPS-neutralizing capacity of HDL and enhances LPS-induced inflammation in mice. Biochim. Biophys. Acta 2015, 1852, 2106–2115. [Google Scholar] [CrossRef] [PubMed]
- Balvers, M.G.; Verhoeckx, K.C.; Bijlsma, S.; Rubingh, C.M.; Meijerink, J.; Wortelboer, H.M.; Witkamp, R.F. Fish oil and inflammatory status alter the n-3 to n-6 balance of the endocannabinoid and oxylipin metabolomes in mouse plasma and tissues. Metabolomics 2012, 8, 1130–1147. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, M.; Manivannan, J.; Sivasubramanian, J.; Silambarasan, T.; Balamurugan, E.; Raja, B. Molecular metabolic fingerprinting approach to investigate the effects of borneol on metabolic alterations in the liver of nitric oxide deficient hypertensive rats. Mol. Cell. Biochem. 2012, 362, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Peng, J.; Xue, Z.; Wei, Y. Construction and analysis of correlation networks based on gas chromatography-mass spectrometry metabonomics data for lipopolysaccharide-induced inflammation and intervention with volatile oil from Angelica sinensis in rats. Mol. Biosyst. 2015, 11, 3174–3187. [Google Scholar] [CrossRef] [PubMed]
- Minkler, P.E.; Kerner, J.; North, K.N.; Hoppel, C.L. Quantitation of long-chain acylcarnitines by HPLC/fluorescence detection: Application to plasma and tissue specimens from patients with carnitine palmitoyltransferase-II deficiency. Clin. Chim. Acta 2005, 352, 81–92. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds synthetic borneol and natural borneol are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | R.T. (min) | Identity | Content% (SB) | Content% (NB) |
|---|---|---|---|---|
| 1 | 2.16 | Camphene | 0.0016 | 0.0071 |
| 2 | 3.37 | Cineole | - | 0.0447 |
| 3 | 4.46 | Fenchone | 0.0038 | - |
| 4 | 6.04 | Camphor | 0.325 | 0.8095 |
| 5 | 6.73 | Iso-butylene | 0.0015 | - |
| 6 | 6.83 | α-fenchol | 0.4557 | 0.0116 |
| 7 | 6.93 | β-fenchyl alcohol | 0.3749 | - |
| 8 | 7.15 | fenchyl acetate | - | 0.0399 |
| 9 | 7.16 | exo-methyl-camphenilol | 0.0482 | - |
| 10 | 7.20 | 3-methyl-camphenilol | 0.0151 | 0.001 |
| 11 | 7.49 | m-menthene | 0.0128 | - |
| 12 | 8.13 | iso-borneol | 35.77 | 0.0918 |
| 13 | 8.49 | Borneol | 61.39 | 98.96 |
| Peak No. | R.T. (min) | Formula | Mass (m/z) | Biomarkers |
|---|---|---|---|---|
| 1 | 6.04 | C16H35NO2 | 274.2668 | Sphinganine |
| 2 | 1.14 | C5H11N | 85.0891 | Piperidine |
| 3 | 17.42 | C16H32O2 | 256.2402 | Fatty acid(14:0(10Me, 13Me)) |
| 4 | 9.64 | C26H52NO7P | 521.3554 | PC(0:1/18:1(9E)) |
| 5 | 15.90 | C20H34O2 | 306.2559 | 3β,5β-androstanediol |
| 6 | 1.12 | C7H13NO3 | 159.0895 | 5-acetamidopentanoate |
| 7 | 18.13 | C18H34O2 | 282.2559 | cis-2-octadecenoic acid |
| 8 | 21.44 | C40H79O7P | 702.5563 | PA(P-18:0/19:0) |
| 9 | 8.78 | C23H45NO4 | 399.3349 | Palmitoylcarnitine |
| 10 | 23.82 | C44H84NO8P | 785.6007 | PC(18:1(11Z)/18:1(9Z)) |
| 11 | 18.12 | C42H80NO8P | 757.5622 | PE(22:2(13Z, 16Z)/15:0) |
| 12 | 10.33 | C25H49NO4 | 427.3662 | DL-stearoylcanitine |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, L.; Zhang, Y.; Li, W.; Zhang, J.; Wang, D.; Fu, J.; Wang, P. Comparison of Chemical Profiles, Anti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Synthetic Borneol and Natural Borneol. Molecules 2017, 22, 1446. https://doi.org/10.3390/molecules22091446
Zou L, Zhang Y, Li W, Zhang J, Wang D, Fu J, Wang P. Comparison of Chemical Profiles, Anti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Synthetic Borneol and Natural Borneol. Molecules. 2017; 22(9):1446. https://doi.org/10.3390/molecules22091446
Chicago/Turabian StyleZou, Liang, Yan Zhang, Wei Li, Jinming Zhang, Dan Wang, Jia Fu, and Ping Wang. 2017. "Comparison of Chemical Profiles, Anti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Synthetic Borneol and Natural Borneol" Molecules 22, no. 9: 1446. https://doi.org/10.3390/molecules22091446
APA StyleZou, L., Zhang, Y., Li, W., Zhang, J., Wang, D., Fu, J., & Wang, P. (2017). Comparison of Chemical Profiles, Anti-Inflammatory Activity, and UPLC-Q-TOF/MS-Based Metabolomics in Endotoxic Fever Rats between Synthetic Borneol and Natural Borneol. Molecules, 22(9), 1446. https://doi.org/10.3390/molecules22091446