AetMYC1, the Candidate Gene Controlling the Red Coleoptile Trait in Aegilops tauschii Coss. Accession As77

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
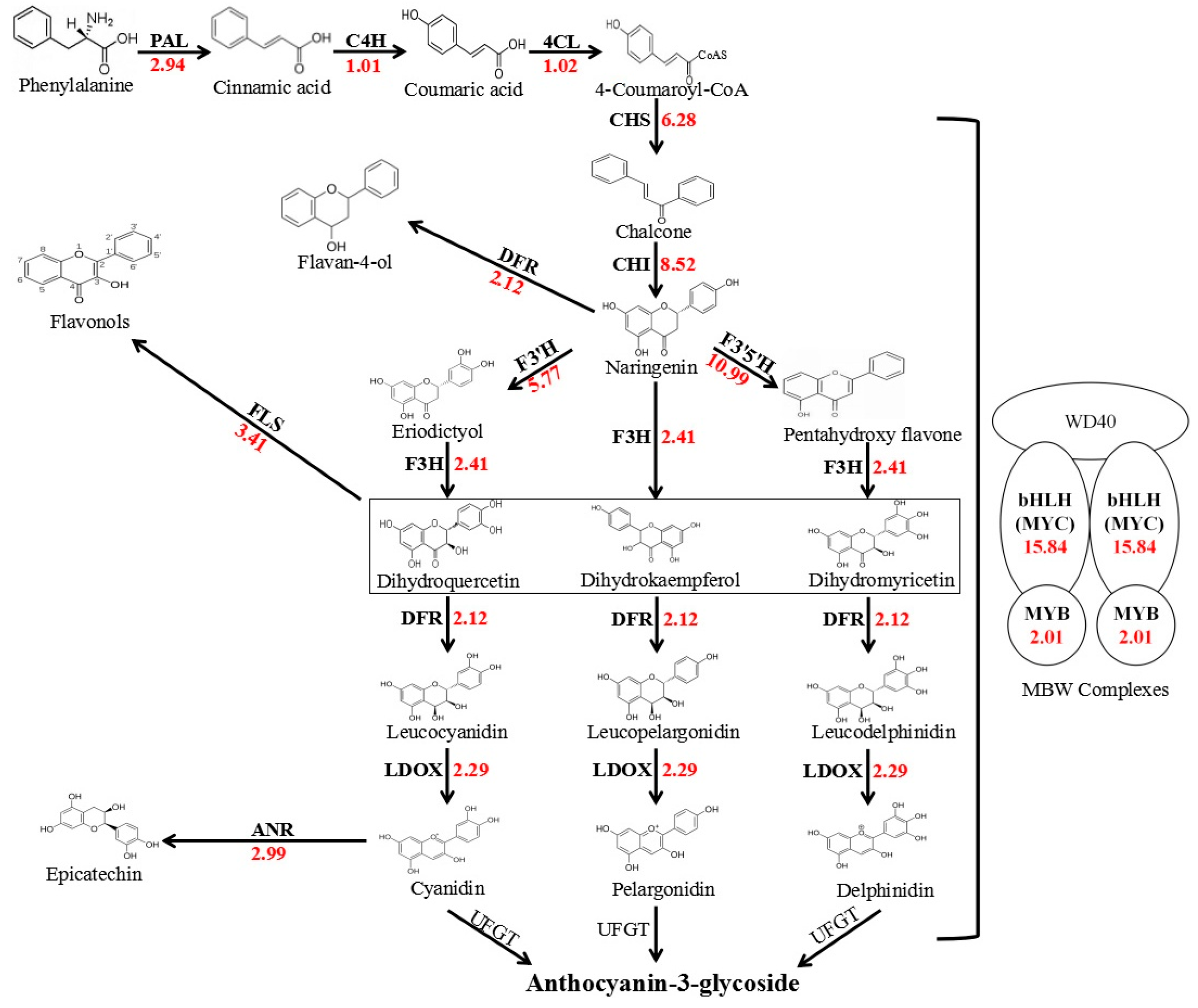
2. Results
2.1. Transcriptome Analyses of Red and White Coleoptiles
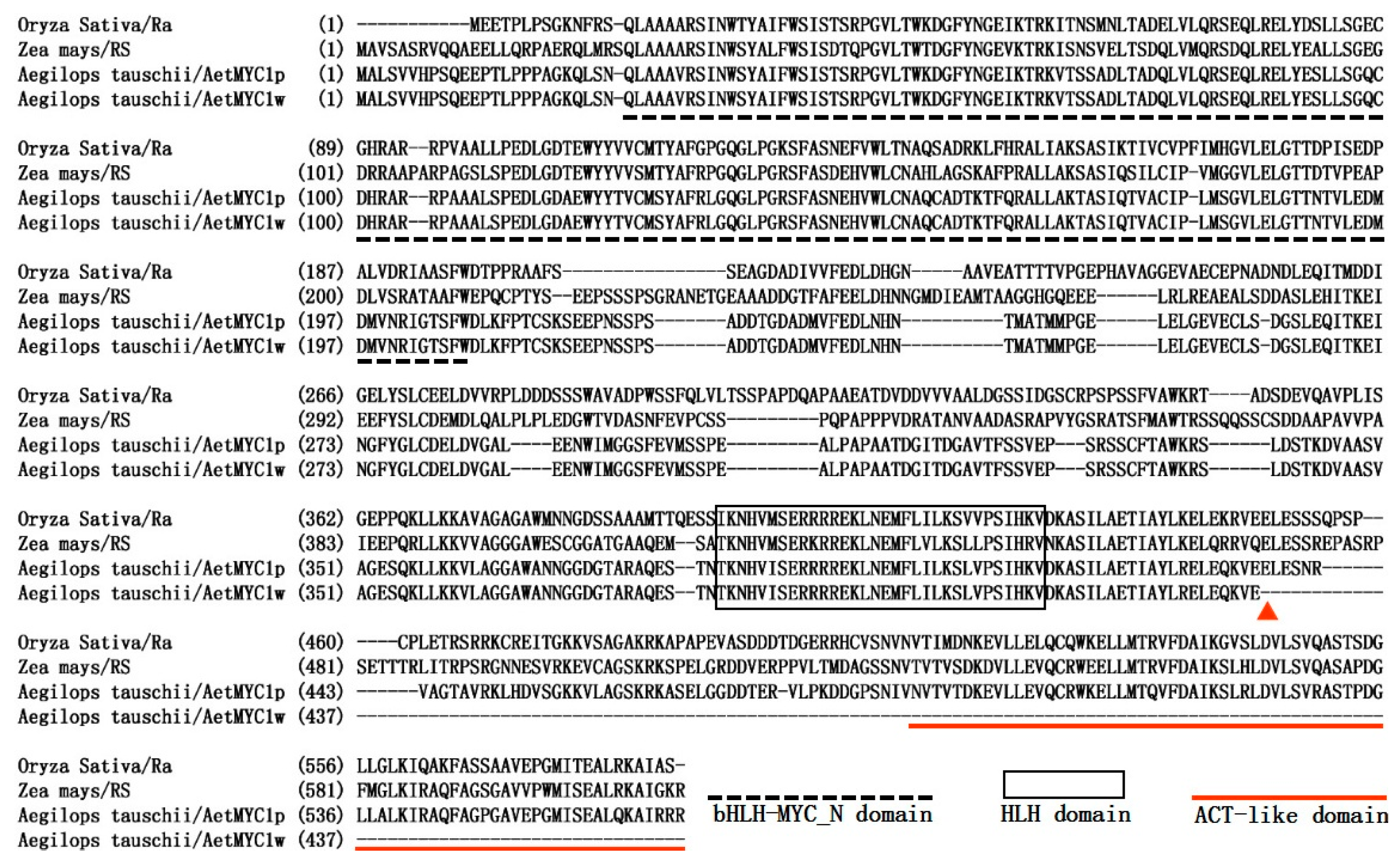
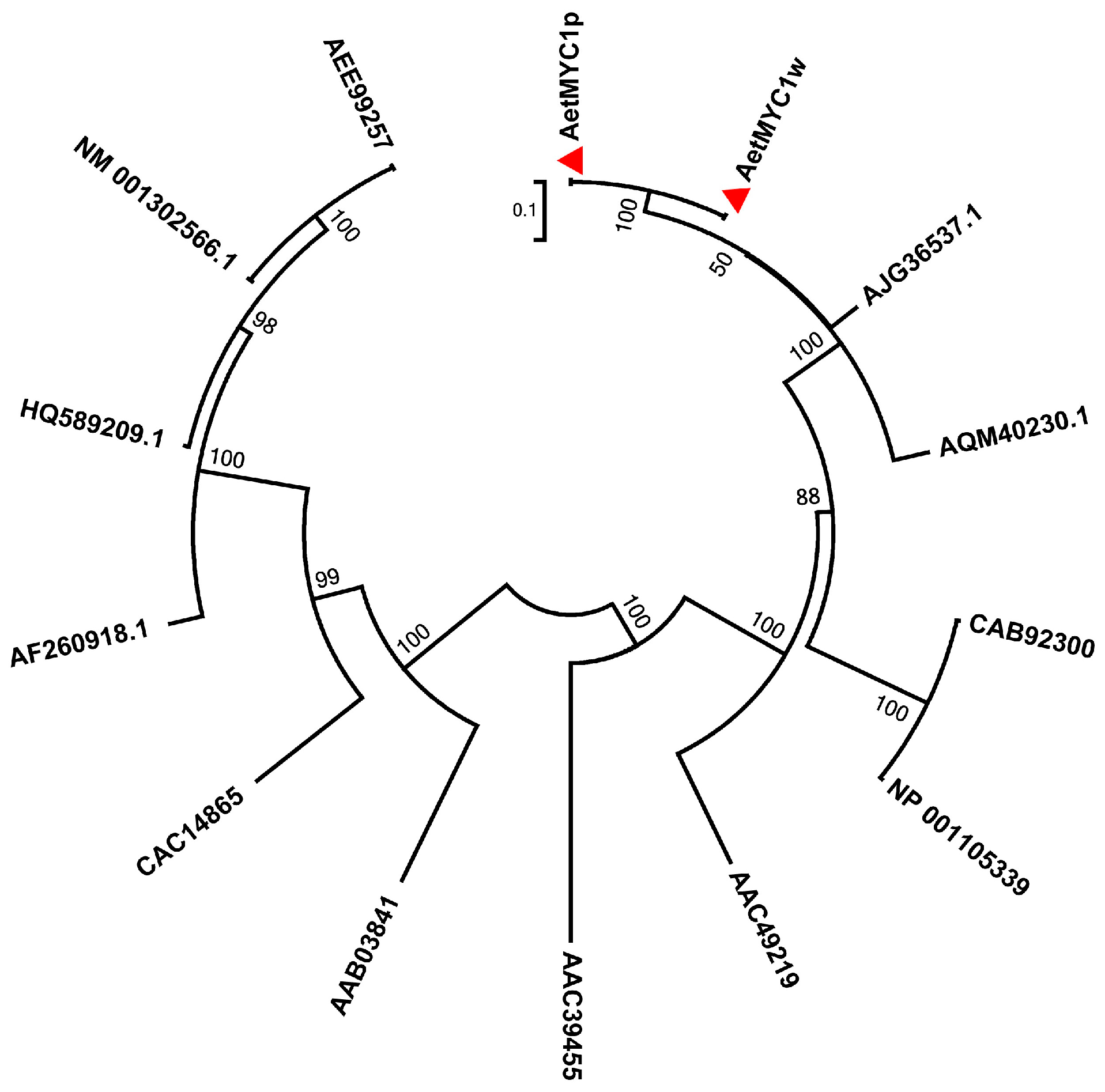
2.2. Molecular Characteristics of AetMYB7D and AetMYC1
2.3. Differences of Transcript Abundance and Regulatory Activity of AetMYB7D and AetMYC1
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Phenotype Scans
4.2. Genomic DNA, Total RNA, and cDNA Preparation
4.3. Transcriptome Analysis
4.4. Gene Isolation
4.5. Transient Expression
4.6. Expression Profiles
4.7. Bioinformatic Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shoeva, O.Y.; Gordeeva, E.I.; Arbuzova, V.S.; Khlestkina, E.K. Anthocyanins participate in protection of wheat seedlings from osmotic stress. Cereal Res. Commun. 2017, 45, 1–10. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Kihara, H.; Yamashita, K.; Tanaka, M. Morphological, physiological, genetical and cytological studies in Aegilops and Triticum collected from Pakistan, Afghanistan and Iran. In Committee of the Kyoto University Scientific Expedition to theKarakoram and Hindukush; Kyoto University: Kyoto, Japan, 1965. [Google Scholar]
- Khlestkina, E.K.; Pestsova, E.G.; Roder, M.S.; Borner, A. Molecular mapping, phenotypic expression and geographical distribution of genes determining anthocyanin pigmentation of coleoptiles in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2002, 104, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Ye, G.; Zong, Y.; Zhang, B.; Chen, W.; Liu, B.; Zhang, H. Diversity and distribution of Rc1 alleles in Triticum urartu from the Middle East. Genet. Resour. Crop Evolut. 2017. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, P.B.; Raasch, J.A.; Kramer, L.C.; Neumann, P.; Wielgus, S.M.; Austin-Phillips, S.; Jiang, J. Sgt1, but not Rar1, is essential for the RB-mediated broad-spectrum resistance to potato late blight. BMC Plant Biol. 2008, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Styles, E.D.; Ceska, O.; Seah, K.T. Developmental differences in action of R and B alleles in Maize. Can. J. Genet. Cytol. 1973, 15, 59–72. [Google Scholar] [CrossRef]
- Atchley, W.R.; Wollenberg, K.R.; Fitch, W.M.; Terhalle, W.; Dress, A.W. Correlations among amino acid sites in bHLH protein domains: An information theoretic analysis. Mol. Biol. Evolut. 2000, 17, 164–178. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Himi, E.; Nisar, A.; Noda, K. Colour genes (R and Rc) for grain and coleoptile upregulate flavonoid biosynthesis genes in wheat. Genome 2005, 48, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Himi, E.; Taketa, S. Isolation of candidate genes for the barley Ant1 and wheat Rc genes controlling anthocyanin pigmentation in different vegetative tissues. Mol. Genet. Genom. 2015, 290, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Hou, X.J.; Zhang, B.; Chen, W.J.; Liu, D.C.; Liu, B.L.; Zhang, H.G. Identification of a candidate gene for Rc-D1, a locus controlling red coleoptile colour in wheat. Cereal Res. Commun. 2016, 44, 35–46. [Google Scholar] [CrossRef]
- Ye, G.J.; Wei, L.; Chen, W.J.; Zhang, B.; Liu, B.L.; Zhang, H.G. Frame-shift mutation causes the function loss of TaMYB-A1 regulating anthocyanin biosynthesis in Triticum aestivum. Cereal Res. Commun. 2017, 45, 35–46. [Google Scholar] [CrossRef]
- Kumar, R.; Ichihashi, Y.; Kimura, S.; Chitwood, D.H.; Headland, L.R.; Peng, J.; Maloof, J.N.; Sinha, N.R. A high-throughput method for Illumina RNA-seq library preparation. Front. Plant Sci. 2012, 3, 202. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Milos, P.M. RNA sequencing: Advances, challenges and opportunities. Nat. Rev. Genet. 2011, 12, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Shoeva, O.Y.; Gordeeva, E.I.; Khlestkina, E.K. The regulation of anthocyanin synthesis in the wheat pericarp. Molecules 2014, 19, 20266–20279. [Google Scholar] [CrossRef] [PubMed]
- Shoeva, O.Y.; Mock, H.-P.; Kukoeva, T.V.; Andreas, B.; Khlestkina, E.K. Regulation of the flavonoid biosynthesis pathway genes in purple and black grains of hordeum vulgare. PLoS ONE 2016, 11, e0163782. [Google Scholar]
- Hu, J.; Anderson, B.; Wessler, S.R. Isolation and characterization of rice R genes: Evidence for distinct evolutionary paths in rice and maize. Genetics 1996, 142, 1021–1031. [Google Scholar] [PubMed]
- Petroni, K.; Cominelli, E.; Consonni, G.; Gusmaroli, G.; Gavazzi, G.; Tonelli, C. The developmental expression of the maize regulatory gene Hopi determines germination-dependent anthocyanin accumulation. Genetics 2000, 155, 323–336. [Google Scholar] [PubMed]
- Strygina, K.V.; Khlestkina, E.K. MYC gene family in cereals: Transformations during evolution of hexaploid bread wheat and its relatives. Mol. Biol. 2017, 51, 772–779. [Google Scholar] [CrossRef]
- Ahmed, N.; Maekawa, M.; Utsugi, S.; Himi, E.; Ablet, H.; Rikiishi, K.; Noda, K. Transient expression of anthocyanin in developing wheat coleoptile by maize C1 and B-peru regulatory genes for anthocyanin synthesis. Breed. Sci. 2003, 53, 29–34. [Google Scholar] [CrossRef]
- Tonelli, C.; Consonni, G.; Dolfini, S.F.; Dellaporta, S.L.; Viotti, A.; Gavazzi, G. Genetic and molecular analysis of Sn, a light-inducible, tissue specific regulatory gene in maize. Mol. Gen. Genet. 1991, 225, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Procissi, A.; Dolfini, S.; Ronchi, A.; Tonelli, C. Light-dependent spatial and temporal expression of pigment regulatory genes in developing maize seeds. Plant Cell 1997, 9, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shu, Q. Fine mapping and candidate gene analysis of purple pericarp gene Pb in rice (Oryza sativa L.). Chin. Sci. Bull. 2007, 52, 3097–3104. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Wing, J.F.; van der Woude, K.; Mol, J.N.; Koes, R. Analysis of bHLH and MYB domain proteins: Species-specific regulatory differences are caused by divergent evolution of target anthocyanin genes. Plant J. 1998, 13, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Wang, X.; Gao, J.; Guo, Y.; Huang, Z.; Du, Y. The tomatohoffman’s anthocyaninless gene encodes a bHLH transcription factor involved in anthocyanin biosynthesis that is developmentally regulated and induced by low temperatures. PLoS ONE 2016, 11, e0151067. [Google Scholar] [CrossRef] [PubMed]
- Mujeebkazi, A.; Rosas, V.; Roldan, S. Conservation of the genetic variation of Triticum tauschii (coss.) schmalh. (Aegilops squarrosa auct. non L.) in synthetic hexaploid wheats (T. turgidum L. s.lat. x T. tauschii; 2n=6x=42, AABBDD) and its potential utilization for wheat improvement. Genet. Resour. Crop. Evolut. 1996, 43, 129–134. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Abdalla, O.; Mujeeb-Kazi, A.; Kazi, A.G.; Xu, S.S.; Gosman, N.; Lagudah, E.S.; Bonnett, D.; Sorrells, M.E.; Tsujimoto, H. Synthetic hexaploids: Harnessing species of the primary gene pool for wheat improvement. J. Plant Breed. Rev. 2013, 37, 35–122. [Google Scholar]
- GRIN NPGS Plant collection sites. Available online: https://www.ars-grin.gov/npgs/sitelist.html (accessed on 18 September 2015).
- Yan, Z.H.; Wan, Y.F.; Liu, K.F.; Zheng, Y.L.; Wang, D.W. Identification of a novel HMW glutenin subunit and comparison of its amino acid sequence with those of homologous subunits. Chin. Sci. Bull. 2002, 47, 220–225. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Romualdi, C.; Bortoluzzi, S.; D’Alessi, F.; Danieli, G.A. Ideg6: A web tool for detection of differentially expressed genes in multiple tag sampling experiments. Physiol. Genom. 2003, 12, 159–162. [Google Scholar] [CrossRef] [PubMed]
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: http://www.genome.jp/kegg/ (accessed on 3 May 2017).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evolut. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of DNA of all Aegilops tauschii accessions are available from the authors. |




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, D.; Ye, G.; Zong, Y.; Zhang, B.; Chen, W.; Liu, B.; Zhang, H. AetMYC1, the Candidate Gene Controlling the Red Coleoptile Trait in Aegilops tauschii Coss. Accession As77. Molecules 2017, 22, 2259. https://doi.org/10.3390/molecules22122259
Cao D, Ye G, Zong Y, Zhang B, Chen W, Liu B, Zhang H. AetMYC1, the Candidate Gene Controlling the Red Coleoptile Trait in Aegilops tauschii Coss. Accession As77. Molecules. 2017; 22(12):2259. https://doi.org/10.3390/molecules22122259
Chicago/Turabian StyleCao, Dong, Guangji Ye, Yuan Zong, Bo Zhang, Wenjie Chen, Baolong Liu, and Huaigang Zhang. 2017. "AetMYC1, the Candidate Gene Controlling the Red Coleoptile Trait in Aegilops tauschii Coss. Accession As77" Molecules 22, no. 12: 2259. https://doi.org/10.3390/molecules22122259
APA StyleCao, D., Ye, G., Zong, Y., Zhang, B., Chen, W., Liu, B., & Zhang, H. (2017). AetMYC1, the Candidate Gene Controlling the Red Coleoptile Trait in Aegilops tauschii Coss. Accession As77. Molecules, 22(12), 2259. https://doi.org/10.3390/molecules22122259