1. Introduction
It is estimated that more than 2 billion people drink tea every day. More than 50 countries cultivates tea [
Camellia sinensis (L.) O. Kuntze] plants. China has cultivated tea for more than 2000 years. During the long cultivation history, numerous elite tea varieties have been cultivated for health benefits. To date, approximately 246 cultivars, such as Longjing and Shuchaozao, are grown for multiple nutraceutical benefits and aroma flavors [
1]. Based on reports from Chinese Government, approximately 9.78 million tons of fresh leaves were harvested from different cultivars to produce various tea products in 2016, and the economic value was approximately
$25.6 billion. Many tea products from China are sold to different regions all over the world.
A daily basis of drinking tea benefits human health. This is because tea contains high contents of polyphenols and other nutraceuticals. The primary tea polyphenols are composed of flavones, flavonols, flavan-3-ols, anthocyanins, proanthocyanidins (PAs), and other specialized forms of flavonoids, among which flavan-3-ols (catechins) account for 70–80% of the total content [
2]. Flavan-3-ols and PAs are the main astringent metabolites in tea beverages. Numerous studies have demonstrated that flavan-3-ols possess multiple pharmaceutical functions, such as antioxidative [
3], antimutagenic, anticarcinogenic [
4], antidiabetic [
5], antibacterial, anti-inflammatory, antihypertensive, and anticardiovascular disease activities [
6], UV-B protection, body weight control [
7], and prevention against Parkinson’s disease and other aging diseases [
8].
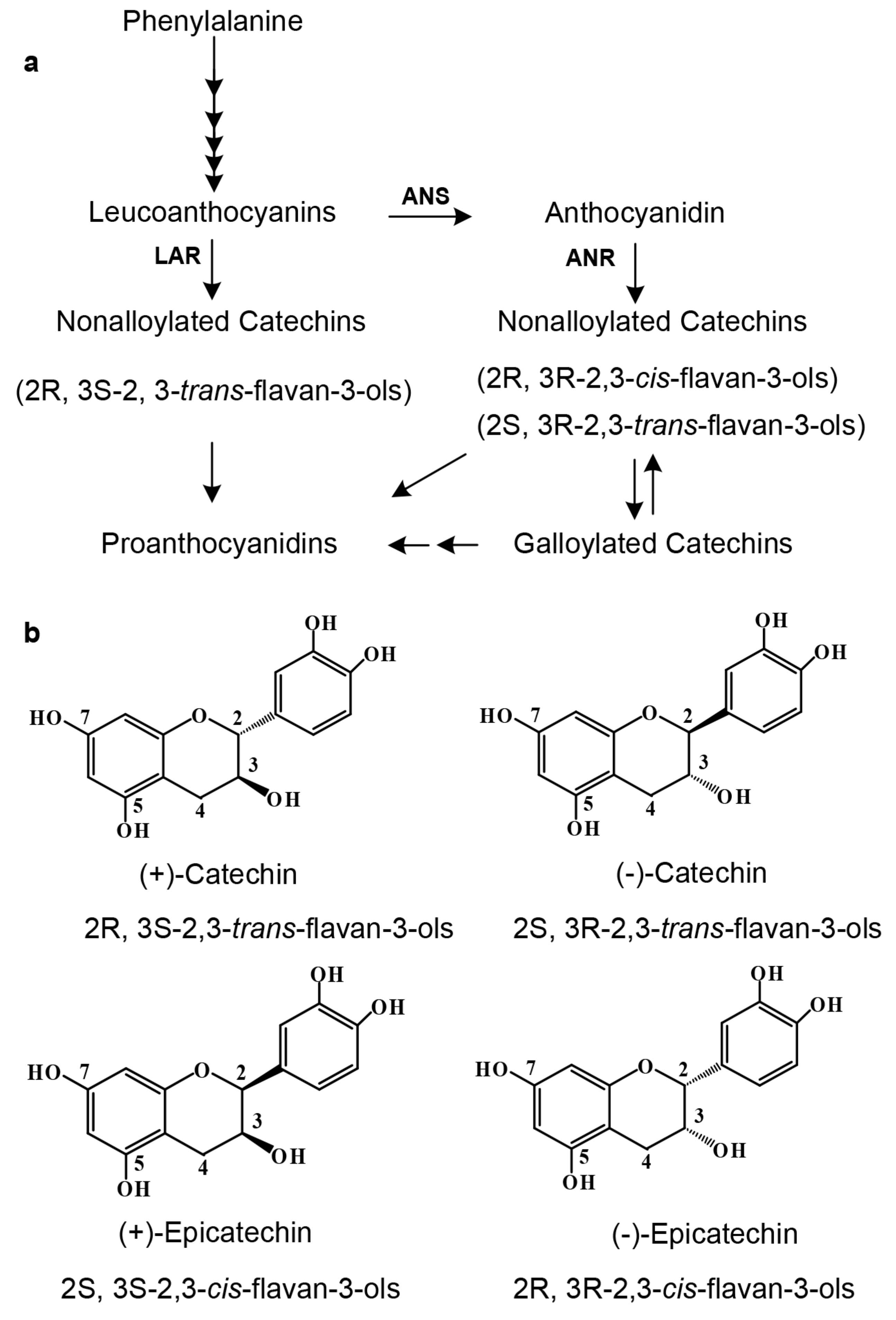
The main composition of tea flavan-3-ols has been appropriately characterized by numerous investigations. Primary aglycones of flavan-3-ols include (+)-catechin [(+)-Ca)] and (−)-epicatechin [(−)-EC)] (
Figure 1b), and (+)-gallocatechin [(+)-GC)] and (−)-epigallocatechin [(−)-EGC)] [
9]. Primary conjugates are composed of (−)-epciatechin-3-
O-gallate [(−)-ECG)] and (−)-epigallocatechin-3-
O-gallate [(−)-EGCG)]. The formation of (−)-ECG and (−)-EGCG was recently characterized by enzyme investigations. Two enzymes, namely UDP-glucose: galloyl-1-
O-β-
d-glucosyltransferase (UGGT) and 1-
O-galloyl-β-
d-glucose
O-galloyltransferase (ECGT) [
2], were isolated from tea leaves. These two enzymes were demonstrated to add gallic acid at the 3-
O positon of both (−)-EC and (−)-EGC to form (−)-ECG and (−)-EGCG.
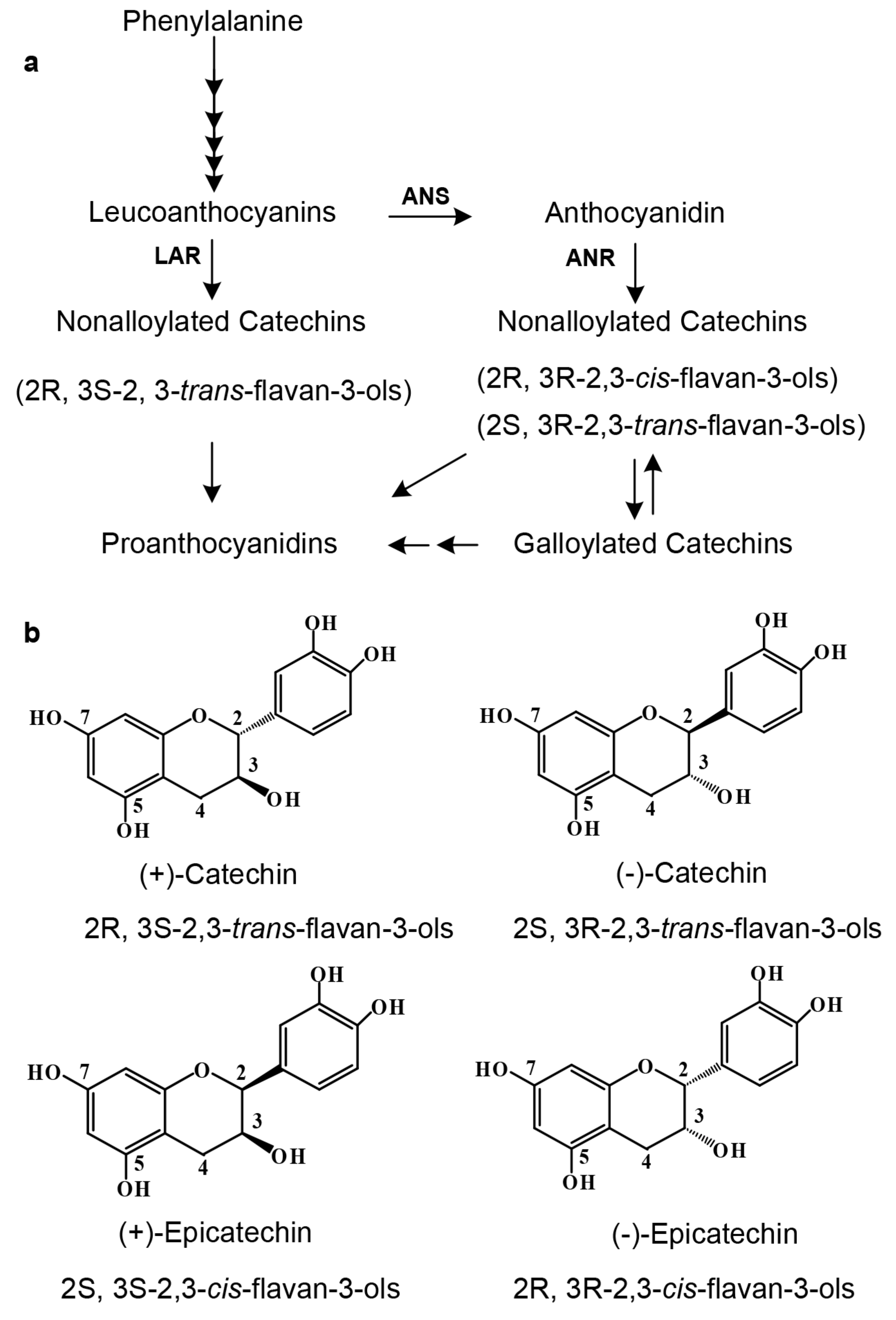
Since 2003, two biosynthetic pathways have been characterized to control the formation of flavan-3-ols, each with different stereo-configuration preference (
Figure 1a) [
9,
10,
11,
12]. One is the anthocyanidin reductase (ANR) pathway, which starts with leucoanthocyanidins and then undergoes anthocyanidins to three stereo types of flavan-3-ols, such as (−)-Ca, (−)-EC, and (+)-EC. This pathway has been shown to exist in the plant kingdom across ferns, gymnosperms, and angiosperms [
13]. This pathway has also been used to develop metabolic engineering technology for production of PAs in commercial crops, such as tobacco plant [
14,
15] and forage crops (such as
Medicago trucatula and
M. sativa) [
14,
16]. The other is the leucoanthocyanidin reductase (LAR) pathway, which is from leucoanthocyanidins to one stereo type of flavan-3-ols, such as (+)-Ca (
Figure 1b) [
13]. Although
Arabidopsis thaliana does not contain the LAR pathway [
10], multiple plants have been shown to include it [
13]. To date, both the ANR and LAR pathways have been demonstrated to associate with the formation of PAs in one tea cultivar [
17,
18]. Two
ANR homologs, namely
CsANR1 and
CsANR2, were recently cloned from a blister blight-resistant tea cultivar TRI2043 grown in Sri Lanka [
17]. Unlike ANRs from
A. thaliana and
M. trucatula, which catalyze cyanidin to (−)-EC and (−)-Ca, recombinant CsANR1 and CsANR2 were shown to catalyze this substrate to (+)-EC and (+)-Ca [
17]. In addition, the ectopic expression of
CsANR1 and
CsANR2 in tobacco plants were reported to lead to the formation of PAs in anthocyanin producing tissues. One LAR, namely
CsLAR, was also cloned and functionally characterized from this tea cultivar. On the one hand, the recombinant CsLAR was characterized to convert leucocyanidin to (+)-Ca. On the other hand, the ectopic expression of
CsLAR led to the formation of (−)-EC in anthocyanin producing tissues of tobacco plants [
17].
Shuchazao is an elite cultivar. It produces orchid-like aroma. To date, it is one of primary cultivars grown in Anhui and Henan provinces, China. The total cultivation field is around 14,000 hectares. The annual fresh leaf yield is 100,000–140,000 tons to produce different superior green tea products [Data from Tea Research Institute of China Academy of Agriculture Sciences AS (TRI, CAAS)] (
http://www.e-chinatea.cn/other_shujuku.aspx). Here, we report to characterize the ANR pathway in Shuchazao grown in the field. LC-MS based metabolic profiling identified six flavan-3-ol molecules including (+)-Ca, (−)-EC, (+)-GC, (−)-EGC, (−)-ECG, and (−)-EGCG. The contents of these six molecules were different in buds and four positional leaves, which are usually used as primary materials to produce tea products. Enzyme analysis showed that crude protein of fresh leaves converted cyanidin and delphinidin to (−)-EC and (−)-EGC as major products and (−)-Ca and (−)-GC as minor products, respectively. Two cDNAs, namely
CssANRa and
CssANRb, were cloned from young leaf tissues and expressed in
E. coli to induce recombinant proteins. Enzymatic analysis demonstrated that CssANRa catalyzed anthocyanidins to 2
S,3
R-2,3-
trans, 2
R,3
R-2,3-
cis-flavan-3-ols, and 2
S,3
S-2,3-
cis-flavan-3-ols such as (−)-Ca, (−)-EC and (+)-EC. CssANRb converted anthocyanidins to 2
S,3
R-2,3-
trans and 2
S,3
S-2,3-
cis-flavan-3-ols such as (−)-Ca and (+)-EC. Transgenic analysis further demonstrated that the overexpression of either of the two genes led to the formation of PAs in transgenic tissues. These data are informative to regulate flavan-3-ol biosynthesis in this elite tea for producing high quality tea products.
3. Discussion
Shuchaozao is an elite commercial tea cultivar. The ANR pathway in Shuchaozao leaves for tea products lacks an appropriate characterization, although, to date, this pathway in numerous other plant species has been solidly substantiated in genetics, biochemistry, metabolic engineering, and evolution [
12,
13,
13,
19,
20], and two
ANR homologs have also been reported in a blister-resistance tea cultivar. In this study, we focused on characterization of the ANR pathway in field-grown plants’ leaves that are used for commercial tea products. Metabolic profiling revealed that young leaves (
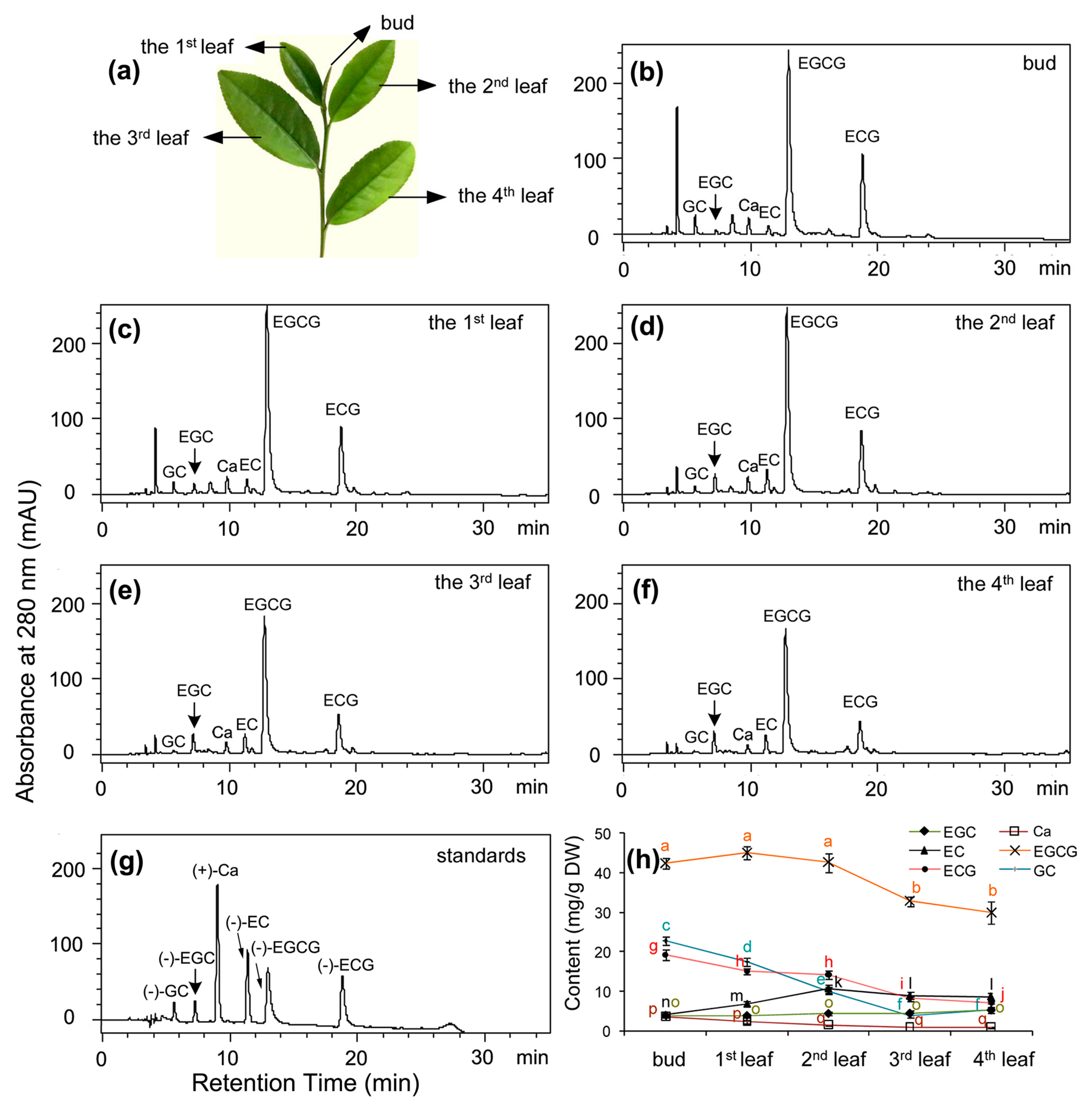
Figure 2a) used for tea products were rich in total flavan-3-ols (
Figure 2h), which were composed of (−)-EGCG, (−)-ECG, (+)-GC, (−)-EC, (−)-EGC, and (+)-Ca. Of these metabolites, (−)-EGCG accounted for approximately 50% of total flavan-3-ols in five positional leaves although its contents decreased in the fully expanded leaves such as the 3rd and 4th leaves. (−)-EGC accounted for 5–10% of total flavan-3-ols in these leaves. Therefore, (−)-EGC and (−)-EGCG together accounted for 55–60% of total leaf flavan-3-ols. In addition, (−)-EC and (−)-ECG accounted for 25–28% of total flavan-3-ols. As we reported previously, (−)-ECG and (−)-EGCG are derived from galloylation of (−)-EC and (−)-EGC [
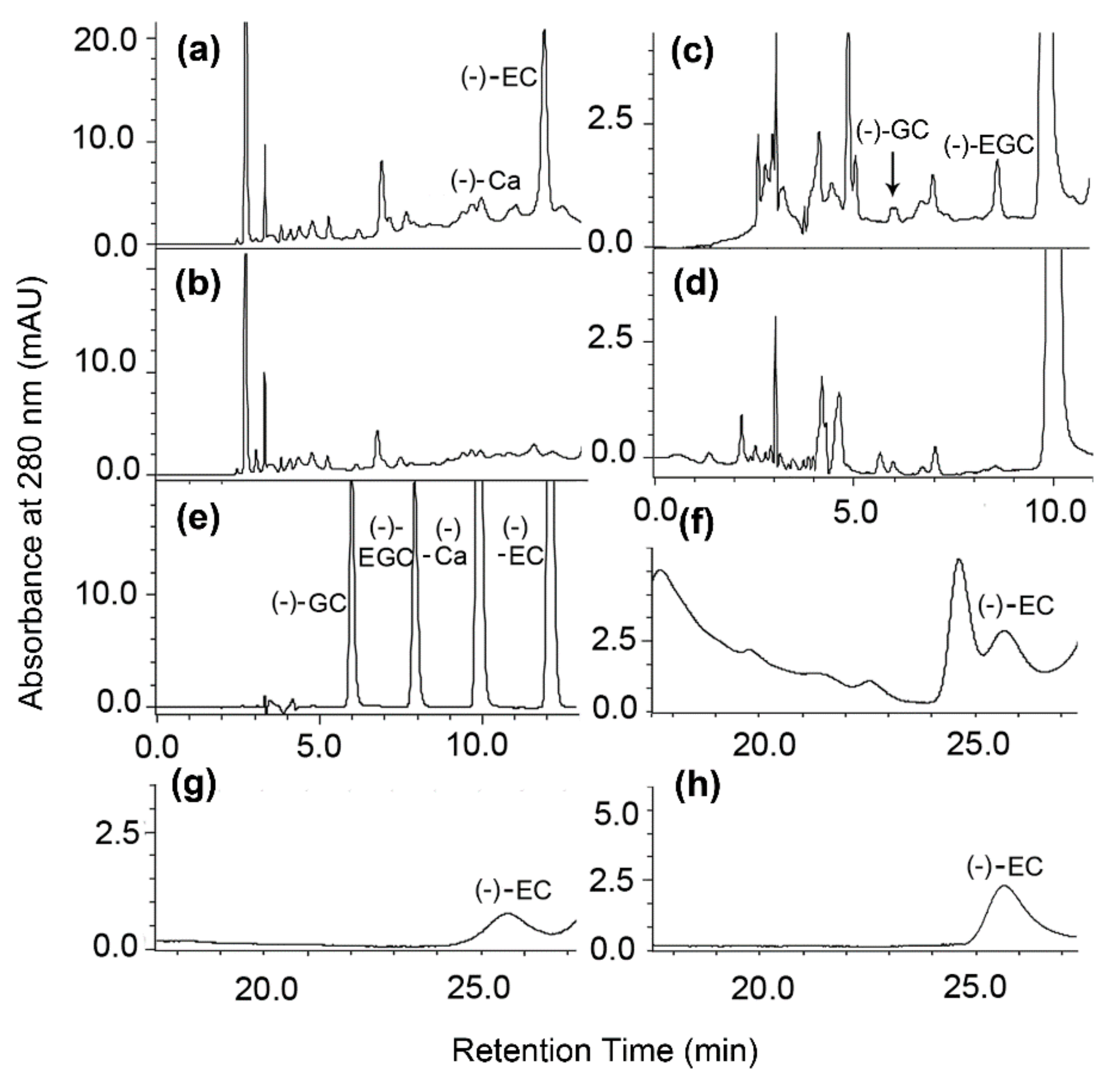
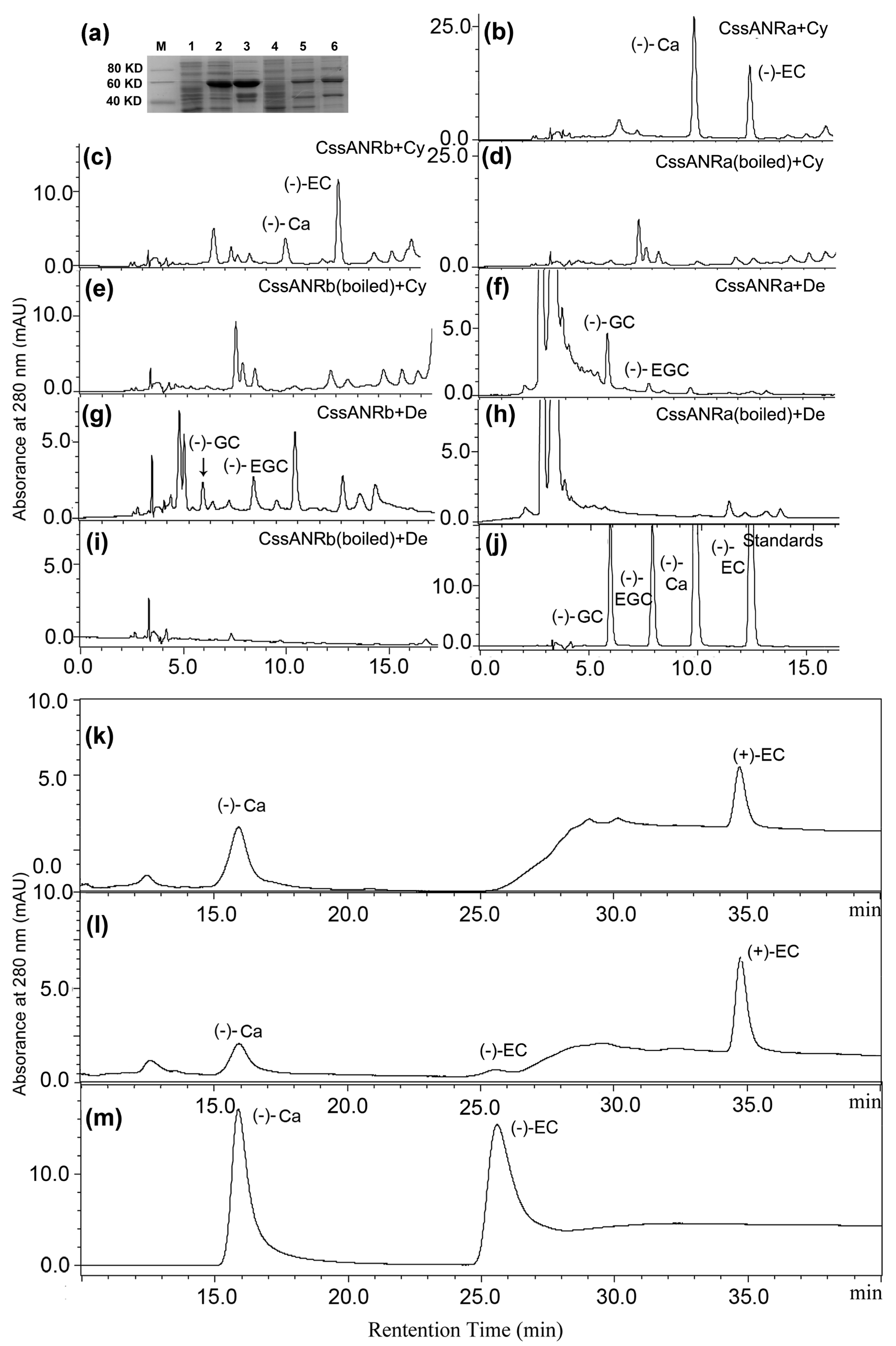
2], which are only derived from the ANR catalysis. Herein, leaf ANR extracts converted cyanidin and delphinidin to (−)-EC and (−)-Ca, and (−)-EGC and (−)-GC, respectively (
Figure 3a–d). Chiral analysis further showed that crude leaf ANR extracts produced (−)-EC but not (+)-EC (
Figure 3f–h). Therefore, these data reveal that the ANR pathway in leaves of Shuchaozao is actively responsible for the high total contents of flavan-3-ols. Furthermore, these data indicate that the ANR pathway is closely associated with flavan-3-ols based high quality products.
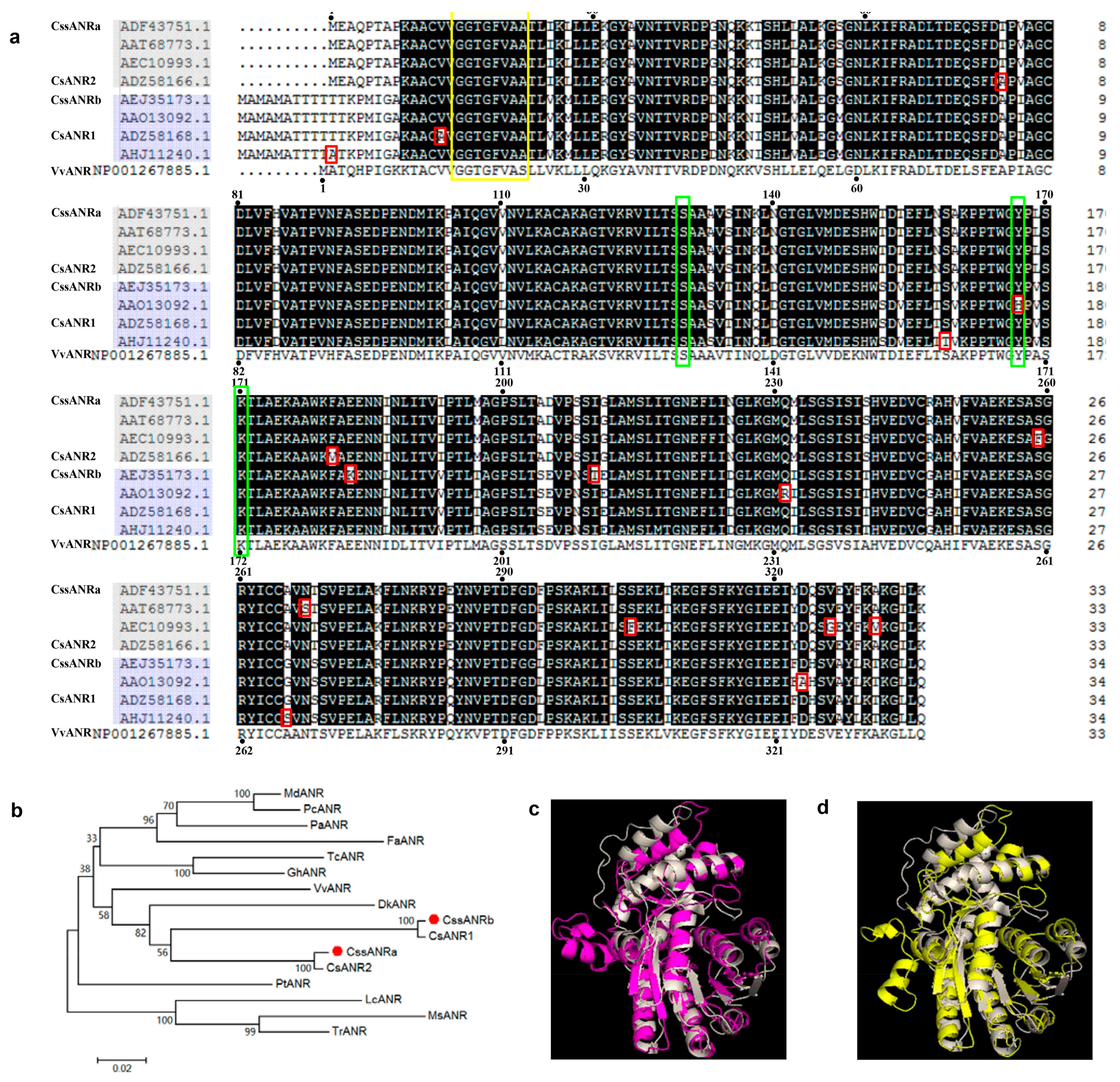
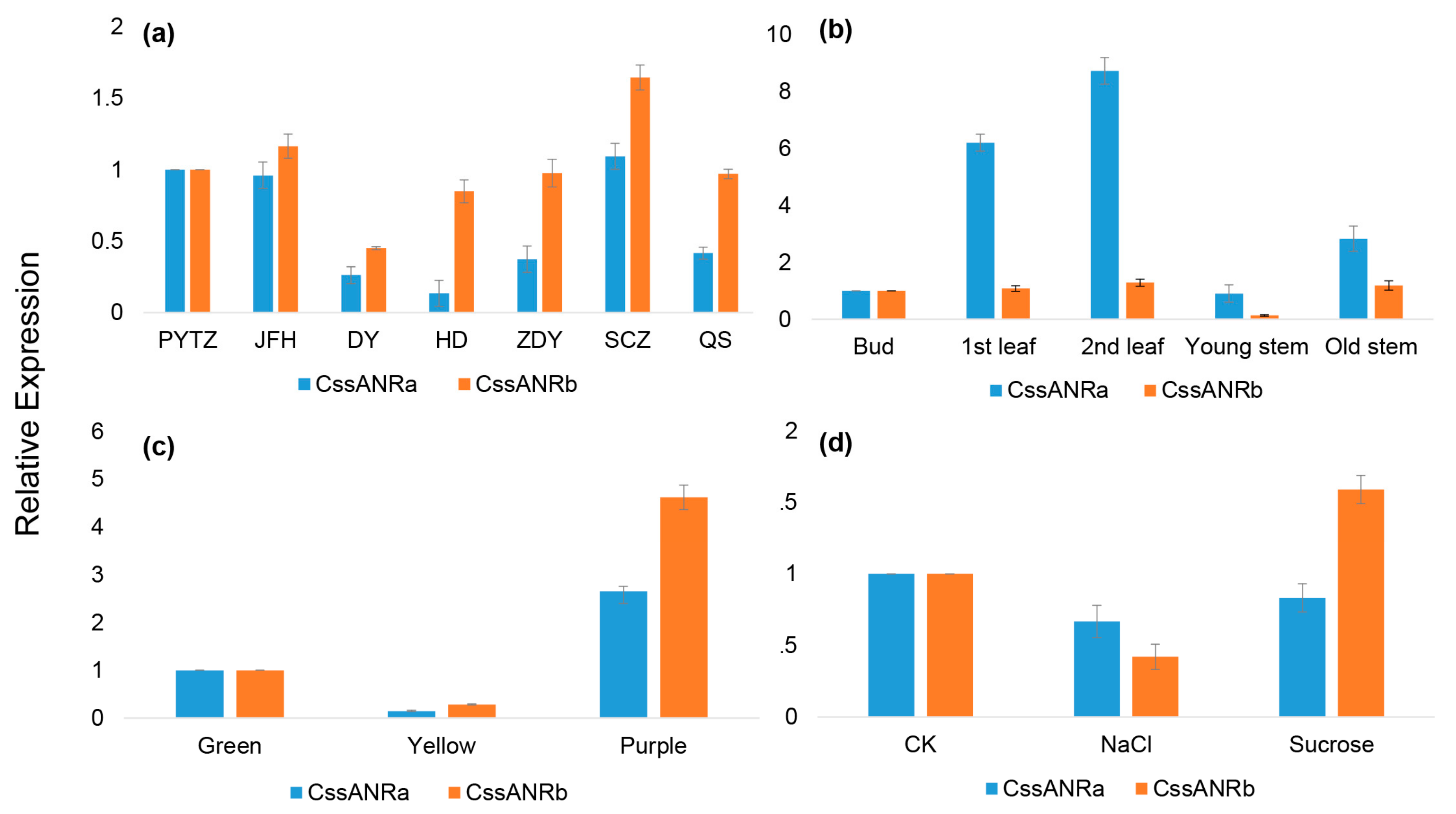
To further understand the role of the ANR pathway in this cultivar, gene specific primers were designed to clone full length of cDNAs. Two
ANR homologs,
CssANRa and
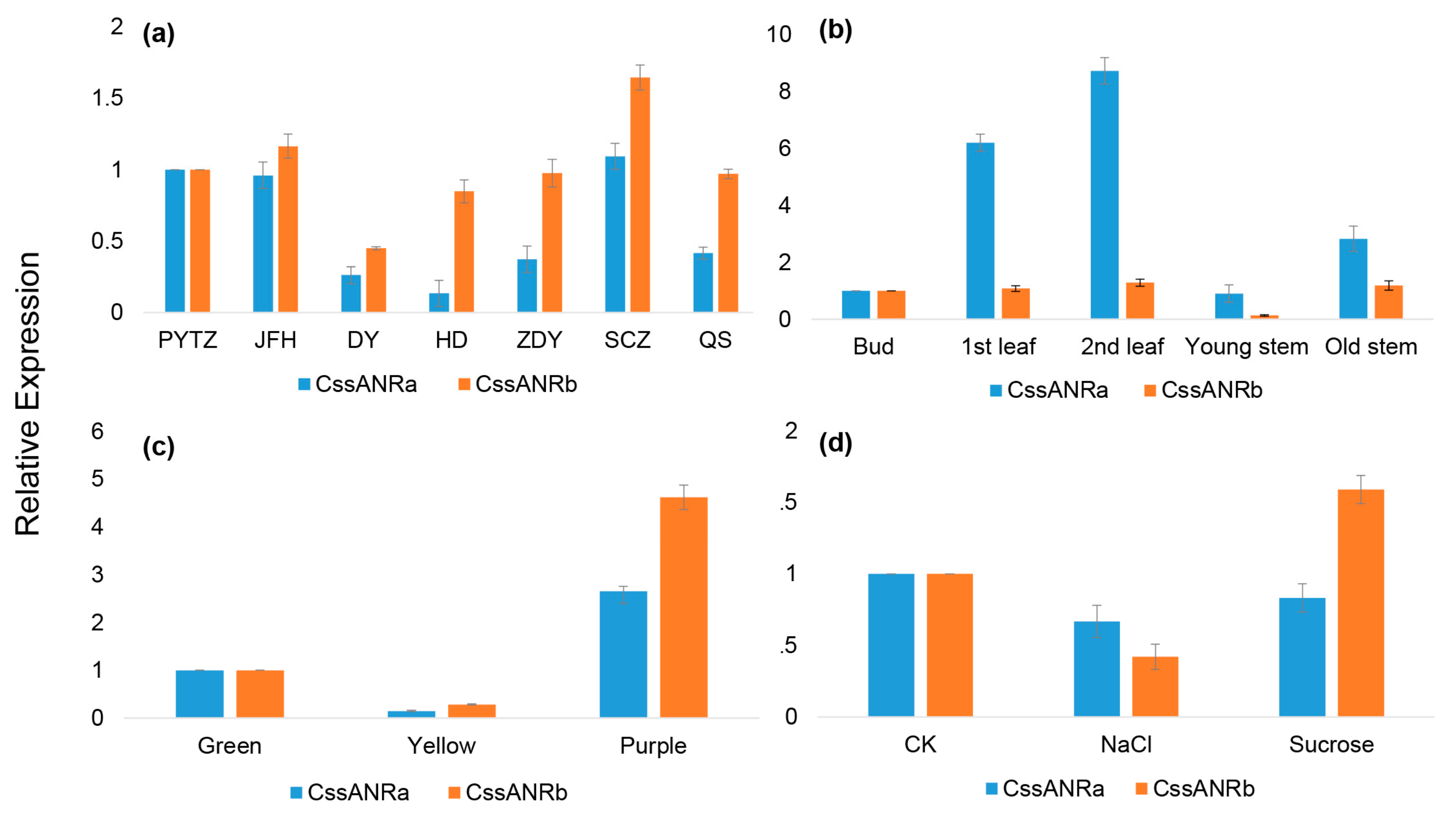
CssANRb, were isolated from young leaves. Results from qRT-PCR analysis showed that the expression levels of
CssANRa were more apparently variable than those of
CssANRb in different tea varieties and different leaf development stages (
Figure 5a,b). By contrast, qRT-PCR analysis characterized that the expression levels of
CssANRb was more obviously variable than those of
CssANRa in differently pigmented leaves and in those leaves under NaCl and sucrose treatments (
Figure 5c,d). In addition, it was interesting that heterogeneous expression in
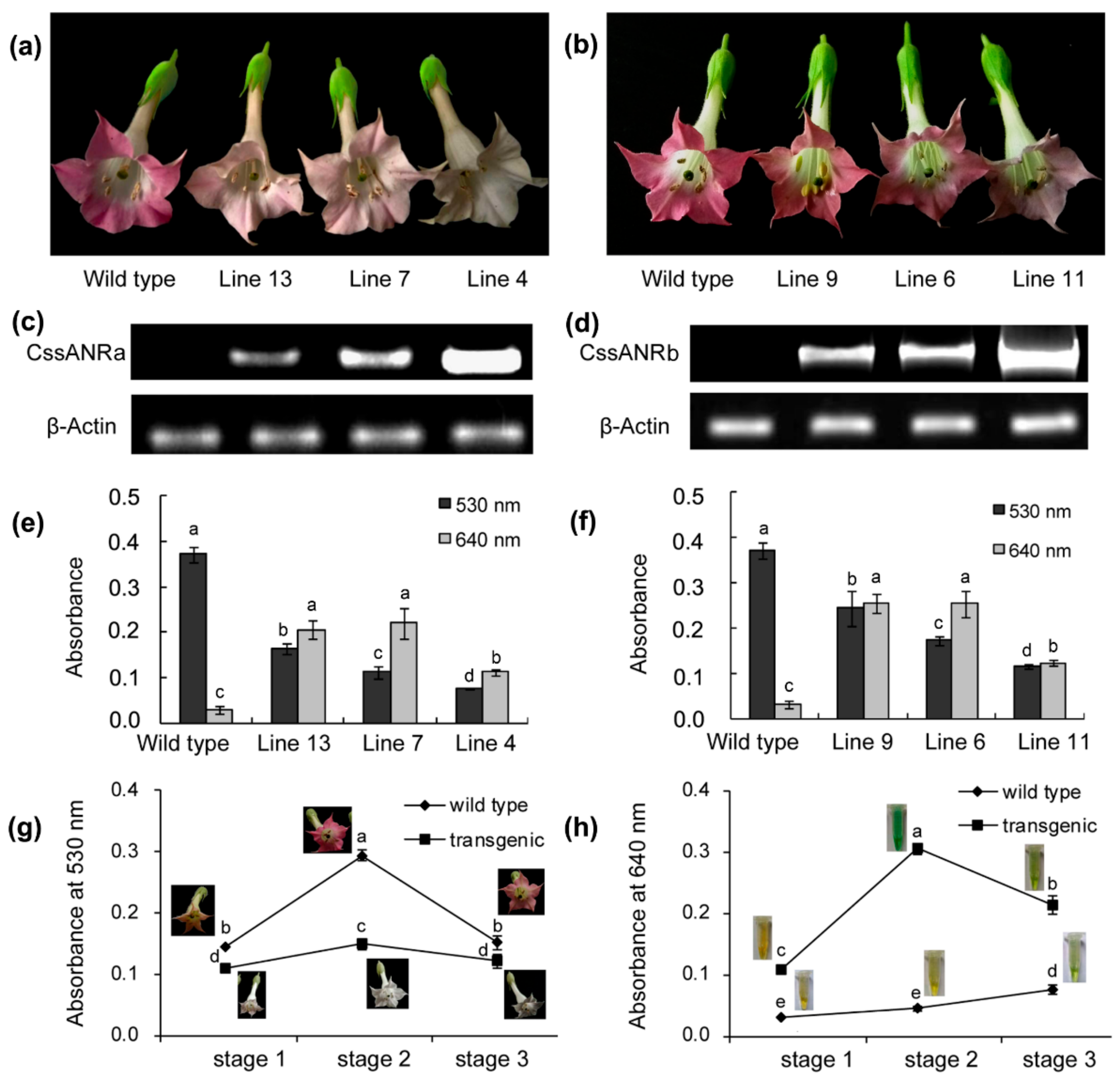
E. coli revealed that
CssANRa and
CssANRb exhibited different plasmid preferences to produce recombinant proteins. The pET31a
+ plasmid was appropriate to induce a recombinant CssANRa but not CssANRb. Instead, the recombinant CssANRb was induced when the pET31 SUMO plasmid was used (
Figure 6a). In addition, recombinant CssANRa and CssANRb showed different pH optima. By contrast, the temperature optima of the two recombinant enzymes were the same. As those results obtained from the
CsANR1 and
CsANR2 overexpression reported by Pang et al. [
17], the overexpression of
CssANRa and
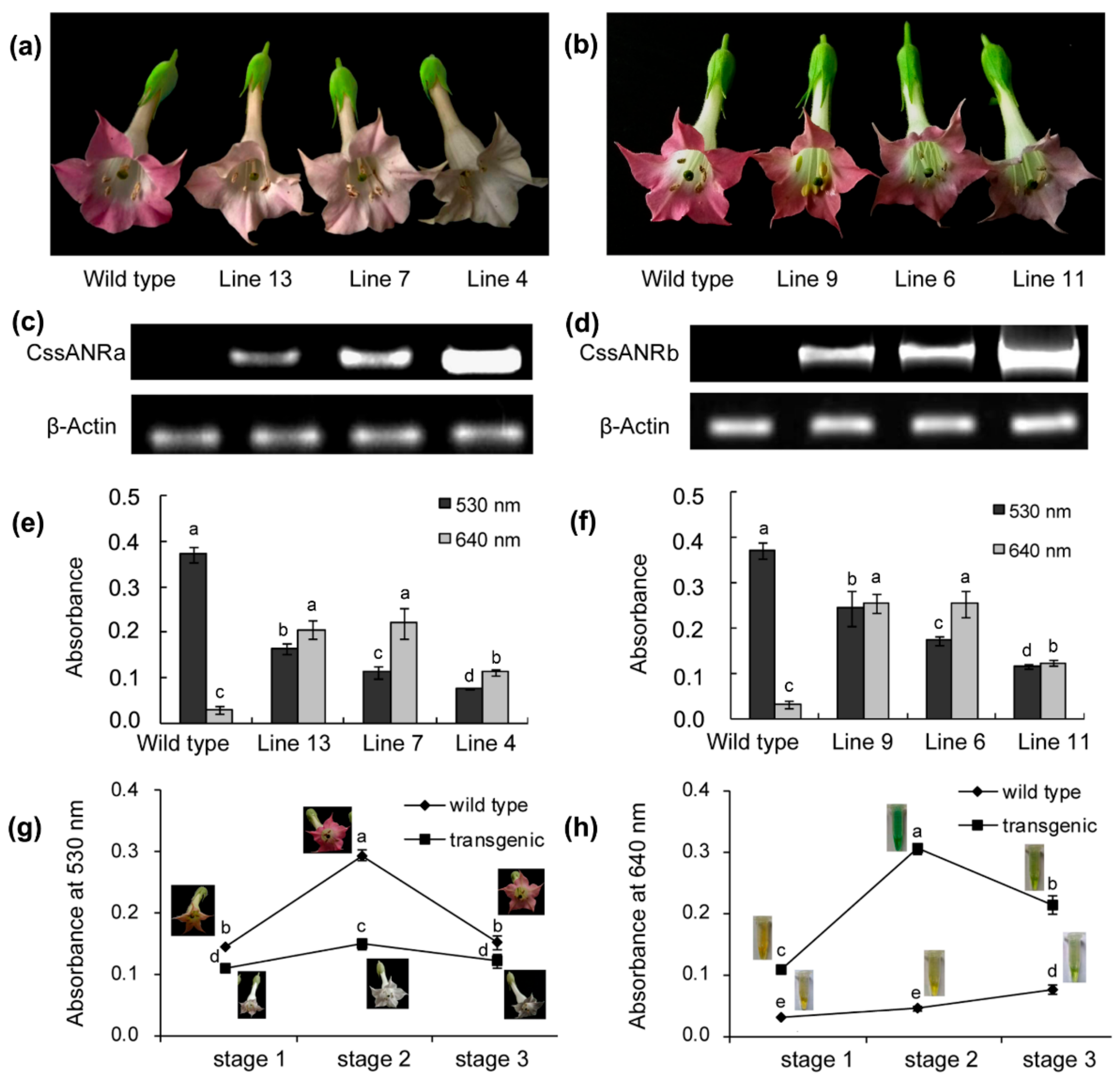
CssANRb also led to the formation of PAs in anthocyanin-producing flowers and reduction of anthocyanins during flower development (
Figure 7). These transgenic data provide evidence that
CssANRa and
CssANRb are involved in the biosynthesis of PAs.
In our experiments, we observed an interesting different result between leaf ANR and recombinant ANR assays. (+)-EC formed from the recombinant ANR assay (
Figure 6k,m) was not observed from the leaf ANR assay (
Figure 3f,h). The biochemical mechanism behind this observation is unclear. One possibility might be associated with the difference of native leaf ANR extract and recombinant ANR. Native ANRs were extracted from
Lotus corniculatus,
Desmodium uncinatum,
Hordeum vulgare,
Vitis vinifera,
Vitis bellula,
Parthenocissus heterophylla,
Cerasus serrulata, and
Dryopteris pycnopteroides [
13]. Enzymatic analysis demonstrated that native ANRs from all of these plant species catalyzed cyanidin to (−)-EC as a major product and (−)-Ca as a minor product. Another interesting possibility may be associated with recombinant vectors. Xie et al. reported that both recombinant AtANR and MtANR fused with pMAL converted cyanidin to (−)-EC and (−)-Ca but no (+)-EC [
21]. Although Xie et al. [
11] did not use a chiral column in their study, they identified chirality via a dichroism spectrum analysis, which resolved the absolute configurations of flavan-3-ols. It was interesting that (+)-EC was observed in reactions catalyzed by His-tag fused ANR. Gargouri et al. demonstrated that a recombinant
V. vinifera ANR fused with a His-Tag performed a dual catalytic activity, reduction and epimerization [
19]. The recombinant VvANR converted cyanidin to (−)-EC, then to (−)-Ca and (+)-EC. Pang et al. [
17] also fused CsANR1 and CsANR2 with a His-Tag, which resulted in the formation of (+)-EC. In our report herein, CssANRa and CssANRb were also fused with His-Tag. These interesting differences indicate that the final understanding of absolute configurations of flavan-3-ols needs more studies of both native plant ANRs and recombinant ANRs.
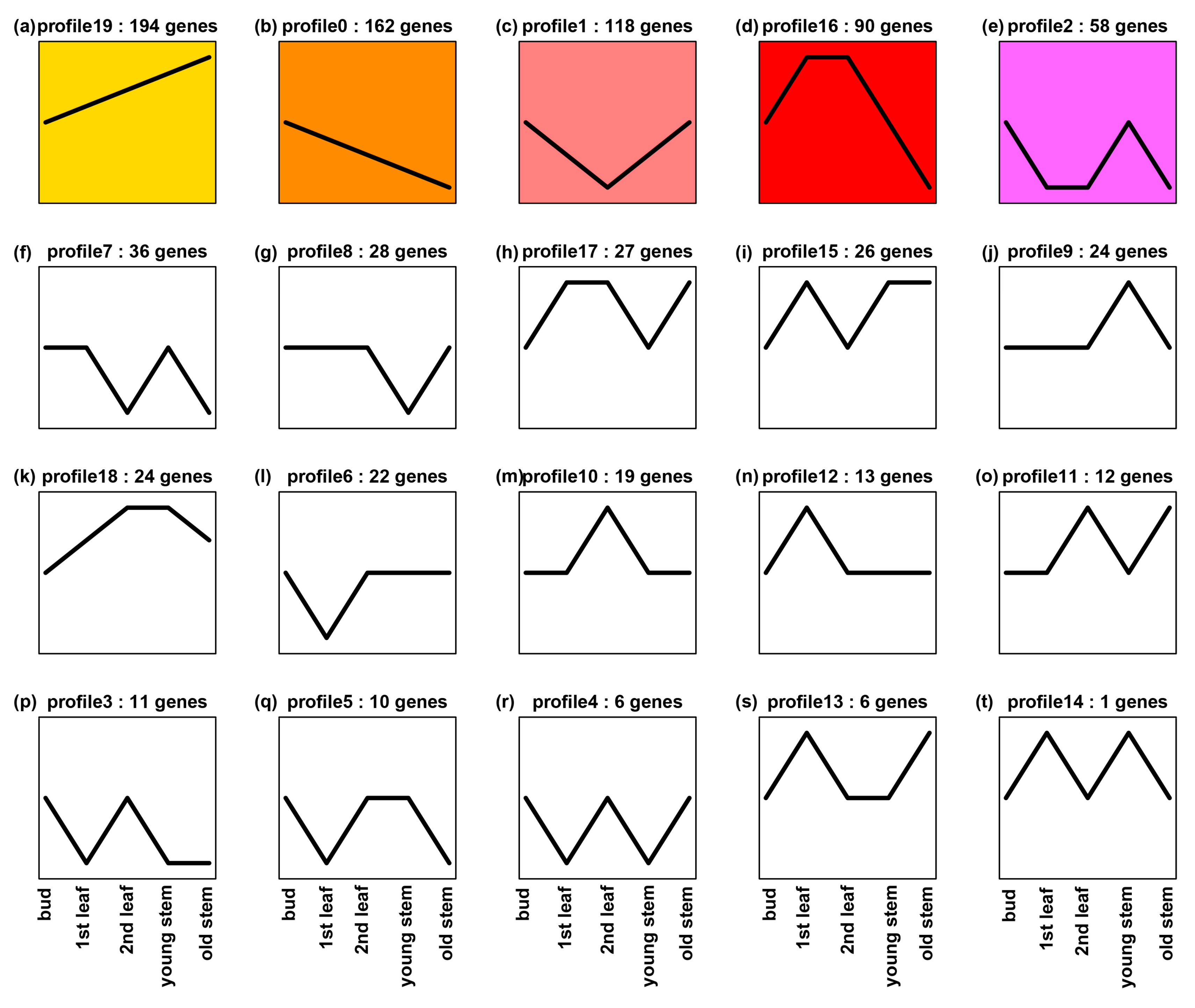
To enhance general understanding of
ANR expression in tea tissues, we further mined
ANR transcripts (without separate two paralogs) in tissues of two other elite green tea varieties, “Pingyangtezao (PYTZ)” and “Ruixue”. Five transcriptomes were sequenced for buds, the 1st leaf, the 2nd leaf, young stem, and old stem of PYTZ, respectively. We obtained transcripts of ANR from all five transcriptomes. In addition, we annotated 1420 potential transcription factor (TF) cDNAs. The transcript of each TF was also obtained from all five transcriptomes. The transcripts of ANR and each TF were used for association analysis of expression profile trend in five tissues. The resulting data revealed the coupled expression of 885 TFs cDNAs and
ANR in each tissue. A further expression trend analysis using 885 TF genes and
ANR resulted in 20 expression profiles in buds, the 1st leaf, the 2nd leaf, young stem, and old stem (
Figure 8). The transcript trend of
ANR was the same as those of 23 TF genes, which was characterized in profile 18 (
Figure 8k). Further annotation analysis characterized the 23 TFs in 13 families (
Supplementary Figure S2), which included four bHLH and 4 MYB members. These data indicate that the expression of
ANR is positively associated with bHLH and 4 MYB members. Meanwhile, we analyzed gene expression association between
ANR and TF cDNAs in flowers. Three transcriptomes were sequenced for three developmental stages of flowers of Ruixue. We also obtained transcripts of
ANR from all three transcriptomes. Moreover, we annotated 1325 TF genes from the three flower transcriptomes and obtained their transcripts in each transcriptome. Based on transcripts of these TF genes and
ANR, we performed expression association analysis to obtain 1100 TF genes, the expressions of which were coupled with that of
ANR. Based on transcript of each TF gene and
ANR, eight expression trend profiles were established (
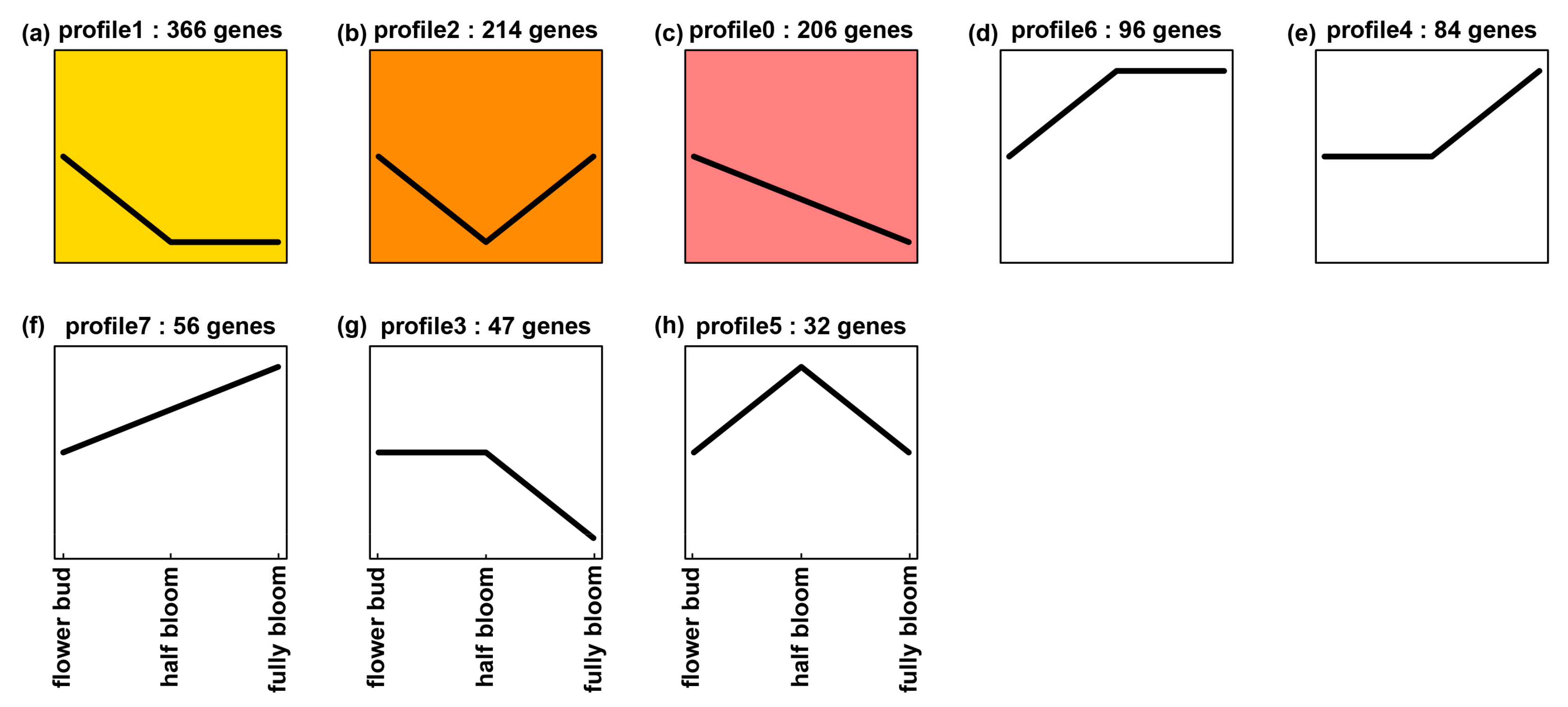
Figure 9). The expression trend of
ANR was the same as those of 205 TF genes, which was characterized in profile 0 (
Figure 9c). The 205 TF genes were annotated to 29 families, which included 74 MYB (MYB family) and 12 bHLH (bHLH family) members (
Supplementary Figure S3). These data indicate that the transcript trend of
ANR is closely associated with those of MYB and bHLH gene members. All of these data will be useful for us to further study the regulation of the ANR pathway in Suchazao and other green tea varieties.
4. Materials and Methods
4.1. Field Growth of Shuchazao and Other Varieties for Sampling
Shuchazao (
Camellia sinensis (L.) O. Kuntze) plants are grown in an experimental tea garden (research station) at Anhui Agricultural University (latitude 31.86° N, longitude 117.27° E, altitude 20 m above mean sea level) and in the farming field in Hefei, Anhui, China. Plants grown in the tea garden is used for numerous research purposes, while plants grown in the field are used to harvest leaves to produce tea products for sale. Plants were grown 10 years when we performed all experiments reported here. Buds, the 1st leaf, the 2nd leaf, the 3rd leaf, and the 4th leaf (
Figure 2a) of new shoots were collected from plants at these two locations. When samples were harvested, they were immediately frozen in liquid nitrogen, transported to laboratory, and then stored in a −80 °C freezer until use for gene cloning and metabolic analysis. In addition, leaves were collected from one year old Shuchazao seedlings and then separated into three groups, which were immediately treated 4 h with 200 mM NaCl and 200 mM sucrose (dissolved in double deionized water) and dd-water as control. Leaf samples treated were immediately frozen in liquid nitrogen and then stored in a −80 °C freezer until RNA isolation and qRT-PCR analysis described below.
In addition, 15 other tea varieties with 10 years old are grown in the same garden for numerous research purposes. Seven were selected to comparatively understand
ANR gene expression. These are
C. sinensis cv. Pingyangtezao (PYTZ),
C. sinensis cv. Jinfenghuang (JFH),
C. sinensis cv. Dayewulong (DY),
C. sinensis cv. Huangdan (HD),
C. sinensis cv. Zhengdayin (ZDY),
C. sinensis cv. Shuchazao (SCZ),
C. sinensis cv. Queshe (QS). The 2nd leaf (
Figure 2a) was sampled from new shoots of these varieties. Eight other varieties were used to compare
ANR expression in three different types of pigmented leaves, green, yellow, and purple. The green group included
C. sinensis cv. LongJing, and
C. sinensis cv. Tieguanyin. For sample collection, Shuchazao was added to form three varieties in this green group. The yellow group included
C. sinensis cv. Huangjinya,
C. sinensis cv. Zhonghuangyihao, and
C. sinensis cv. Zhonghuangerhao. The purple group consisted of
C. sinensis cv. Zijuan,
C. sinensis cv. Ziyan, and
C. sinensis cv. Sunrouge). The 2nd leaf of new shoots (
Figure 2a) was also sampled from each variety in each group for qRT-PCR analysis.
4.2. Extraction and Analysis of Flavan-3-ols in Leaves from Field-Grown Shuchazao Plants
Fresh samples, including buds, the 1st leaf, the 2nd leaf, the 3rd leaf, and the 4th leaf (
Figure 2a), were grounded into fine powder in liquid nitrogen. A powdered sample (100 mg) was suspended in 1 mL extraction solution (methanol: ddH
2O, 80:20) in a 1.5 mL tube and sonicated for 10 min at room temperature. After centrifugation at 4000×
g for 15 min, the resulting upper clear phase was pipetted to a new 1.5 mL tube. The remaining pellet was extracted a second time. Supernatants were combined and the final volume was adjusted to 2 mL. All extracts were filtered through a 0.22 µm membrane for HPLC analysis.
Extracts from different tissues were analyzed using high performance of liquid chromatography (HPLC) on WATERS600 (Waters, Milford, MA, USA) with a photodiode array detector. Metabolites were separated with a reverse-phase column (Synergi 4u Fusion-RP8 0.5 μm, 250 × 4.6 mm) (Phenomenex, Torrance, CA, USA). Elution solvents were composed of A (1% acetic acid) and B (acetonitrile). A gradient elution program used to separate metabolite was composed of: 0–30 min with 13–30% B, 30–32 min with 13% B, and 32–35 min with 0% B. The flow rate was set 1 mL/min. Ultraviolet (UV) spectra was recorded at 280 nm for identification of flavan-3-ol components. (−)-EC, (+)-Ca, (+)-GC, (−)-EGC, (−)-ECG and (−)-EGCG standards were used as positive controls to identify metabolites from plant tissues and enzyme assays described below.
4.3. Extraction of Crude Enzyme from Field-Grown Tea Leaves and Enzyme Assay
Frozen tea leaves (approximately 25 g, a mixture of the 1st–4th leaves) were ground into fine powder in liquid nitrogen. The powdered sample was completely suspended in 200 mL phosphate buffer (0.1 M, pH 7.4) supplemented with 25 g polyvinyl polypyrrolidone (PVPP) and 5 mM β-mercaptoethanol in a 250 mL centrifugation tube. The mixture was placed on ice for 1 h and then was centrifuged at 12,000× g for 15 min at 4 °C. The resulting supernatant (80 mL) was pipetted into a new tube, followed by addition of ammonia sulfate (from 0% to 40%) to precipitate proteins. The mixture was centrifuged at 12,000× g for 15 min at 4 °C and the resulting supernatant was disposed to a waste container. The remained residue was dissolved in 5 mL phosphate-buffer (0.1 M, pH 7.0) and centrifuged at 12,000× g for 10 min at 4 °C. The resulting supernatant containing crude proteins was used for ANR activity assay described below.
Enzyme assay for crude enzyme extract was carried out by following a method reported by Xie et al. [
21] and Zhang et al. [
22] with slight modifications. In brief, enzyme assay was carried out in 15 mL capped polypropylene tubes. The reaction volume was 3.5 mL consisting of 100 mM phosphate buffer (pH 6.5), 2 mM NADPH, 0.5 mM substrate (cyanidin or delphinidin), and 300 μg crude enzyme extract. Boiled crude enzyme extract was used to replace crude enzyme in the reaction as one control. In addition, reactions without adding substrates were carried out as the other control. The enzymatic reactions were incubated for 40 min at 40 °C. All reactions were stopped by addition of 1 mL ethyl acetate (EA) to reaction tubes and vortexing, followed by centrifugation at 5000×
g for 5 min. The resulting ethyl acetate supernatant phase was pipetted into a new tube. This step was repeated two times. Three times of EA extractions was pooled together to obtain 3 mL extract. EA was dried off via a rotary evaporation at room temperature. The remained residues were completely suspended in 100 μL HPLC-grade methanol, followed by centrifugation at 10,000×
g for 2 min. The methanol phase was transferred to a new tube for HPLC analysis, which was as described above in the analysis of flavan-3-ols in leaves from field-grown Shuchazao plants. (−)-EC, (−)-Ca, (−)-GC, and (−)-EGC standards were used to identify metabolites from enzymatic reactions.
4.4. Isolation of Shuchazao ANR cDNA (CssANR) and Quantitative Reverse Transcription-Poly Chain Reaction (qRT-PCR) Analysis
Total RNA was isolated from mixed samples of buds and leaves (
Figure 2a) using the RNAiso-mate for Plant Tissue Kit according to the manufacturer’s protocol (Takara, Tokyo, Japan). The first strand cDNA was synthesized using PrimeScript
® RT reagent Kit according to the manufacturer’s protocol (Takara, Tokyo, Japan). According to two cDNA sequences, KY615701.1 (CssANRa) and KY615702.1 (CssANRb), which we previously submitted to GenBank, the first pair of primers consisting of forward 5’-ATGGAAGCCCAACCGACA-3’ and reverse 5’-TCAATTCTTCAAAATCCC3’ was designed to amply for
CssANRa. The second pair of primers consisting of forward 5’-ATGGCAATGGCAATGGCAACAAC-3’ and reverse 5’-TCAGTTCTGCAAAAGCCCCTTAG-3’ was designed to amplify
CssANRb. The open reading frame (ORF) of
CssANRa and
CssANRb were amplified using a thermal gradient program that was composed of 5 min at 94 °C, 30 cycles of 30 s at 94 °C, 30 s at 62 °C, and 1 min at 72 °C, followed by a 10-min extension at 72 °C. The products of PCR were gel-purified using Takara MiniBEST Agarose Gel Extraction Kit (Takara) and ligated into the pMD18-T vector (Takara, Tokyo, Japan). The ligated products were transformed into
E. coli strain DH5α competent cells using electroporation. Transformed cells were streaked on LB medium supplemented with 50 mg/L ampicillin. Positive colonies were selected to isolate recombinant plasmids. The resulting plasmids were termed pMD18-T-
CssANRa and pMD18-T-
CssANRb for sequencing at BGI (
http://www.bgitechsolutions.com/).
In addition, primer pairs were designed for qRT-PCR analysis that was conducted using SYBR-Green PCR Master mix (Invitrogen, Shanghai, China) and were carried out on a LightCycler
® 480 System (Roche Diagnostics, Indianapolis, IN, USA). The primer pair for
CssANRa included forward 5’-GAGTACTTCAAGGCTAAGGGG AT-3’ and reverse 5’-CAAGCAAACCAAGCAAAACC-3’. The primer pair for
CssANRb included forward 5’-CTGGCAATCCAAGGAGTGC-3’ and 5’-GCCCCGTTCCATCAAGC-3’. The glyceraldehyde-3-phosphate dehydrogenase gene (
GAPDH) is housekeeping gene in tea plants. It was used for normalization. The primer pair used for this gene consisted of forward 5’-TTGGCATCGTTGAGGGTCT-3’ and reverse 5’-CAGTGGGAACACGGAAAGC-3’. Values were normalized against the expression level of
GAPDH [
23]. Relative expression values were calculated with the 2
−△△Ct method. Values of gene expression were means of four replicates.
4.5. Analysis of ANR Sequences
Available
C. sinensis ANR sequences curated at NCBI was identified and their amino acid sequences were aligned using ClustalX. In addition, four
C. sinensis and 12 other species’ ANR amino acid sequences were used to construct a phylogenetic tree via Molecular Evolutionary Genetics Analysis version 5.0 (MEGA5.0; MegaSoftware, Tempe, AZ, USA) [
24] using a neighbor–joining statistical method (with bootstrapping 1000 replicates) [
25]. These 12 ANRs are MsANR (
Medicago sativa, ADK95116.1, LcANR (
Lotus corniculatus, ABC71333.1), FaANR (
Fragaria ananassa, ABG76842.1), PaANR (
Prunus avium, ADY15312.1), MdANR (
Malus domestica, AEL79861.1), TcANR (
Theobroma cacao, ADD51353.1), PtANR (
Populus trichocarpa, XP_002305639.1), VvANR (
Vitis vinifera, CAD91911), DkANR (
Diospyros kaki, BAF56654.1), PcANR (
Pyrus communi, ABB77695.1), GhANR (
Gossypium hirsutum, ABM64802.1), TrANR (
Trifolium repens, ADV31321.1). Four
C. sinensis ANRs were CsANR1 (ADZ58168.1) and CsANR2 (ADZ58166.1) reported previously as well as CssANRa (ASU87432.1) and CssANRb (ASU87433.1) described here. The three-dimensional structures of CssANRa and CssANRb were modelled using homology modeling methods provided by Swiss Model (
https://swissmodel.expasy.org/).
4.6. Induction of Recombinant CssANRa and CssANRb in E. coli
The pMD18-T-CssANRa and -CssANRb vectors containing the ORFs of CssANRa and CssANRb were digested using XbaI and SnaBI restriction enzymes. The resulting ORFs of CssANRa and CssANRb were separated on agarose gel using electrophoresis and then purified using the MiniBEST Plasmid Purification Kit (Takara) according to the manufacturer’s protocol. In addition, the pET31a+ vector was digested using the same restriction enzymes. Then, the ORFs of CssANRa and CssANRb were separately ligated to pET31a+ overnight using T4 DNA Ligase (Takara) under 15 °C. The ligation products were introduced to BL21 (DE3) competent cells using a common electroporation method. Transformed BL21 cells were streaked on agar-solidified medium containing 50 mg/L ampicillin. Antibiotic-resistant colonies were selected to confirm the presence of ORFs in the plasmids and positive colonies were used to induce protein expression. The new plasmids were named as pET31a+-CssANRa and -CssANRb, respectively. Furthermore, the pET31a+ vector was also introduced into BL21 cells as protein induction control. In addition to pET31a+, the ORF of CssANRb was ligated to the T7 SUMO vector (Invitrogen, K300-01) using the same ligation method. The ligation products were introduced to One Shot Mach1™-T1R Competent Cells (Invitrogen, K300-01) to select positive colonies on LB medium supplemented with 50 mg/L kanamycin. Kanamycin resistant colonies were selected to confirm the presence of CssANRb. The resulting new plasmid was named as T7 SUMO-CssANRb. In addition, T7 SUMO was introduced to the same competent cells as vector control for induction of recombinant protein expression.
Five colonies each containing one of pET31a+-CssANRa, pET31a+-CssANRb, T7 SUMO-CssANRb, pET31a+, and T7 SUMO plasmids were used to induce protein expression. Each colony was inoculated into 10 mL LB broth containing 50 mg/L antibiotics (ampicillin for pET31a+-CssANRa and pET31a+ vector, and kanamycin for T7 SUMO-CssANRb and T7 SUMO vector) in an E-flask. All flasks were placed on a shake with a speed of 250 rpm at 37 °C. When cells were grown to a concentration indicated by an absorbent value about 0.6 that was recorded at 600 nm on an UV spectrophotometer, each flask was added isopropyl β-d-1-thiogalactopyranoside (IPTG) to a final concentration of 0.4 mM. Each flask was continuously shaken 3 h at 37 °C. Cells were harvested by centrifugation at 4000× g for 15 min at 4 °C. The remained cell pellet was suspended in 10 mL 100 mM phosphate buffer (pH 6.5) by vortexing. A T4 lyase was added to the suspension mixture with a final concentration of 100 µM. Then, the mixture was sonicated for 1 min on ice, followed by 20 min of centrifugation at 10,000× g at 4 °C. The supernatant was transferred to a new tube. Crude protein extracts were further treated using the MagnetHis Protein Purification System (Promega, Madison, WI, USA) to remove excessive proteins of E. coli according to the manufacturer’s protocol. Partially purified protein dissolved in 100 mM phosphate buffer (pH 6.5) was measured to estimate concentrations with the Bio-Rad Protein Assay system (Bio-RAD, California, CA, USA) and were stored in −80 °C until enzyme assay described below.
4.7. Enzyme Assays
Partially purified recombinant CssANRa and CssANRb (
Figure 6a) were assayed using the method reported by Xie et al. [
21] to optimize temperature and pH values. In all assays, experiments were carried out in 1 mL volume. Three replicates were completed for each reaction. The incubation time was 40 min. Boiled enzyme was used as control. In addition, reactions without addition of substrates were used as another control.
For optimization of temperature, enzyme reactions were carried out in 2 mL volume including 100 mM phosphate buffer (pH 6.5), 2 mM NADPH, 0.5 mM cyanidin, and 150 μg recombinant proteins. Tested temperature values included 0, 10, 20, 30, 40, 50 and 60 °C.
To optimize pH value, enzyme reactions were performed in 2 mL volume consisting of a buffer, 2 mM NADPH, 0.5 mM cyanidin, and 150 μg recombinant CssANRa or CssANRb. Buffers tested included 100 mM citric acid phosphate buffer (pH 4.0~6.0), 100 mM phosphate buffer (pH 6.0~7.2), and 100 mM Tris-HCl buffer (pH 7.5~9.0) were prepared to obtain different pH values from 4.0 to 9.0. Tested pH values included 4.0. 4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0 and 9.0. The incubation temperature was 40 °C.
Methods for stopping all reactions, extracting metabolites, and HPLC analysis were as described for crude enzyme assay.
4.8. Chiral Analysis of Products from Enzyme Reactions
HPLC using a normal-phase chiral column (catalog No. 80325; Chiral Technologies; 5 μm, 4.6 × 250 mm; Tokyo, Japan) was performed to determine the chirality of enzyme reaction products from cyanidin substrate according to a method reported by Pang et al. [
17]. Elution buffer was composed of solvent A (hexane:acetic acid, 99.5:0.5,
v/
v,) and solvent B (ethanol:acetic acid, 99.5:0.5,
v/
v). Isomers were separated using a gradient program, which was composed of 0–20 min with 20% solvent B, 20–23 min with 20–50% B, 23–38 min with 50% B, and 38–40 min with 50–20% B. The flow rate was set at 1 mL/min. UV spectrum was recorded at 280 nm to characterize isomers. (−)-EC and (−)-Ca standards were used as control.
4.9. Construction of Binary Vectors, Tobacco Transformation, and Flower Sample Harvest
The Gateway
® Cloning System (Invitrogen, New York, NY, USA) was used to construct binary vectors according to a method reported by Lei et al. [
26]. In brief, pairs of primers containing attB Gateway sequence were designed to amplify
CssANRa and
CssANRb to develop binary vectors. The primer pairs were CssANRa-attB (attB1 and attB2) and CssANRb-attB (attB1 and attB2) (
Table S1). The pMD18-T-
CssANRa and -
CssANRb plasmids were used as template for PCR. The thermal program used for PCR was composed of 95 °C × 2 min, 35 cycles of 94 °C × 1 min, 56 °C × 1 min, and 72 °C × 1.5 min, and 72 °C for 10 min extension. The resulting PCR products were purified and then cloned to the entry vector pDONR207 with Gateway
® BP Clonase
® Enzyme mix according to the manufacturer’s instructions (Invitrogen). The resulting ligation products were introduced to competent DH5α cells, which were selected on agar-solidified LB plates containing gentamycin. Antibiotic-resistant colonies were selected and cultured to isolate recombinant entry vector. The positive entry vectors were incubated with the Gateway plant transformation destination vector pCB2004 using Gateway
® LR ClonaseTM enzyme (Invitrogen) according to the manufacturer’s protocol. The incubation mixtures were then introduced into competent DH5α cells, which were further selected on agar-solified LB medium containing 50 µg/mL kanamycin. Positive colonies were selected to isolate plasmids. Finally, two plasmids, namely pCB2004-
CssANRa and -
CssANRb, were created, in which two genes were controlled by a 35S promoter. These two vectors and empty pCB2004 (as control) were introduced into competent
Agrobacterium tumefaciens strain EHA105 by electroporation, respectively. One positive colony was obtained for each vector. Positive colonies, namely EHA105-pCB2004-
CssANRa, EHA105-pCB2004-
CssANRb, EHA105-empty pCB2004, were selected on agar-solified LB medium containing 50 µg/mL kanamycin for genetic transformation.
A single colony, each for EHA105-pCB2004-CssANRa, EHA105-pCB2004-CssANRb, and EHA105-empty pCB2004, was inoculated into 20 mL liquid LB medium containing 50 mg/L kanamycin and 50 mg/L spectinomycin in 100 mL E-flask. Three flasks were placed on a rotary shaker at 200 rpm in the dark at 28 °C for overnight until OD value was about 0.6 at 600 nm. Cultured cells were transferred to a 50 mL sterile polypropylene centrifuge tube, followed by centrifugation at 4500× g for 10 min and disposal of supernatant. The remaining pellet was suspended in 20 mL liquid MS medium containing 100 μM acetosyringone (Sigma-Aldrich, St. Louis, MO, USA R40456), followed by centrifugation as above. This step was repeated once and the pellet was suspended in 20 mL liquid MS medium containing 100 μM acetosyringone to obtain activated cells for genetic transformation.
Sterile seedlings of tobacco (
Nicotiana tabacum ‘G28′) plants were grown on agar-solidified MS medium. Leaves of seedlings were cut into 1 × 1 cm discs. Infection of leaf discs with activated
A. tumefaciens strain EHA105 described above and selection of transgenic plants on regeneration medium containing 25 mg/L phosphinothricin followed a protocol reported previously [
27]. Gene specific primer pairs (CssANRa-test and CssANRb-test) were designed to screen transgenic plants (
Table S1). RT-PCR as described above for expression profiling analysis was carried out to identify positive transgenic candidates. Positive transgenic vs. wild-type tobacco plants were planted into pot soil and then grown in a glass house, which was provided with natural light and maintained at 22–25 °C. Flower pigmentation was photographed every day. Flowers were collected from different developmental stages and then frozen in liquid nitrogen immediately. Frozen samples were stored at −80 °C until PA and anthocyanin analysis described below.
4.10. Analysis of PAs and Anthocyanins in Transgenic vs. Wild-Type Tobacco Flowers
Flowers were ground into fine powder in liquid nitrogen to extract anthocyanins and PAs. Anthocyanins were extracted from 100 mg powdered tobacco flower samples as described previously [
23]. Anthocyanin levels were estimated using absorbent values recorded at 530 nm on a UV spectrophotometer (HITACHI, Tokyo, Japan) [
23]. PAs in flowers were extracted as reported previously [
28]. The PA levels were determined using blue coloration via a reaction of PAs with 0.2% DMACA solution [0.2% (
w/
v) dissolved in methanol:concentrated HCl, 9:1]. After reaction for five min, samples were measured at 640 nm on a UV spectrophotometer (HITACHI, Tokyo, Japan) [
17]. Absorbent values at 640 nm were used to compare PAs in transgenic vs. wild-type tobacco flower samples.
4.11. ANR Transcript Abundance Trend in Tissues
Recently, eight transcriptomes were sequenced from buds, the 1st leaf, the 2nd leaf, young stem, old stem, and three developmental stage flower samples of two commercial verities, “Pingyangtezao (PYTZ)” and “Ruixue”. Transcriptomes of buds, the 1st leaf, the 2nd leaf, young stem, and old stem of PYTZ were sequenced by Gene Denovo Biotechnology Co. (Guangzhou, China). Although these transcriptomes have not been published, we were allowed to mine transcripts of tea
ANR and transcription factor genes. Three developmental stages of flowers Ruixue were alabastrum (S1), half opened (bloom) flower (S2) and full bloom (S3). Transcriptomes from these flower stages were sequenced by the Beijing Genomics Institute (BGI,
http://www.genomics.cn/, Shenzhen, China) [
29]. Transcripts of total
ANR cDNAs were extracted from these transcriptomes to compare its abundance in different tissues. Moreover, all unigenes in these transcriptomes were blasted against transcription factor (TF) database from plant (
http://planttfdb.cbi.pku.edu.cn/, version 4.0) to annotate potential TFs. The resulting TF genes were further blasted against tree genomes (NCBI Sequence Read Archive Database No. PRJNA381277) [
30] to unify sequences based on BLASTN 2.6.0+ (NCBI, U.S. National Library of Medicine, Bethesda, MD, USA) [
31]. Transcripts of all TF genes were extracted from all eight transcriptomes and then analyzed using the omicshare cloud platform (
http://www.omicshare.com/tools/Home/Soft/trend) to understand each TF gene expression trend pattern, those TF genes which with the same trend were grouped together to be tagged as one group. As a result, different profile groups were numerated into profile 1, 2, and so on.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}