Thymbra capitata (L.) Cav. and Rosmarinus officinalis (L.) Essential Oils: In Vitro Effects and Toxicity on Swine Spermatozoa

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of the EOs
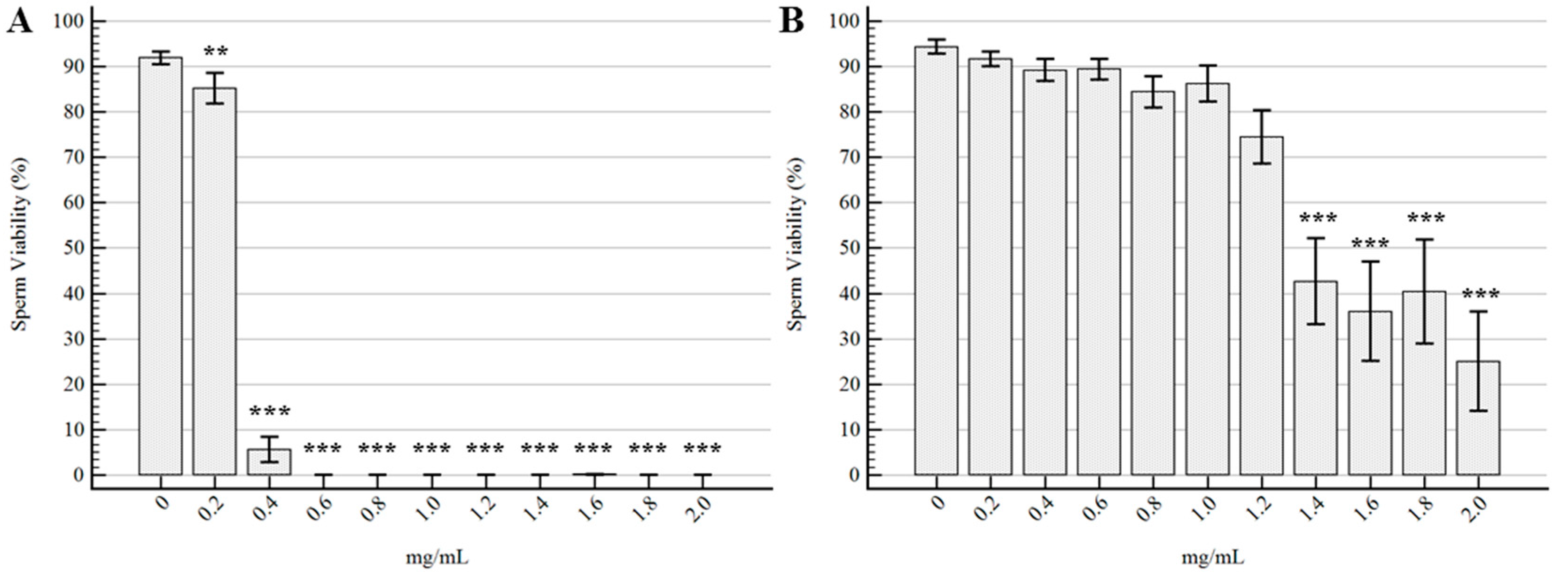
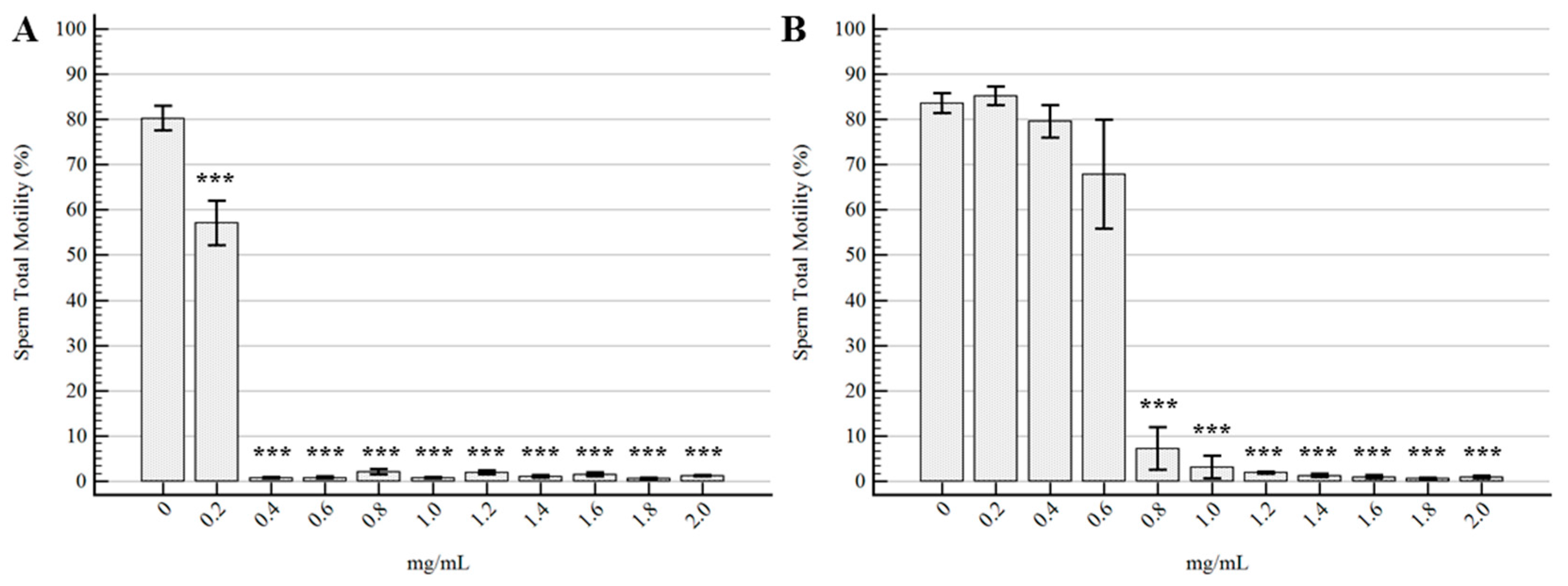
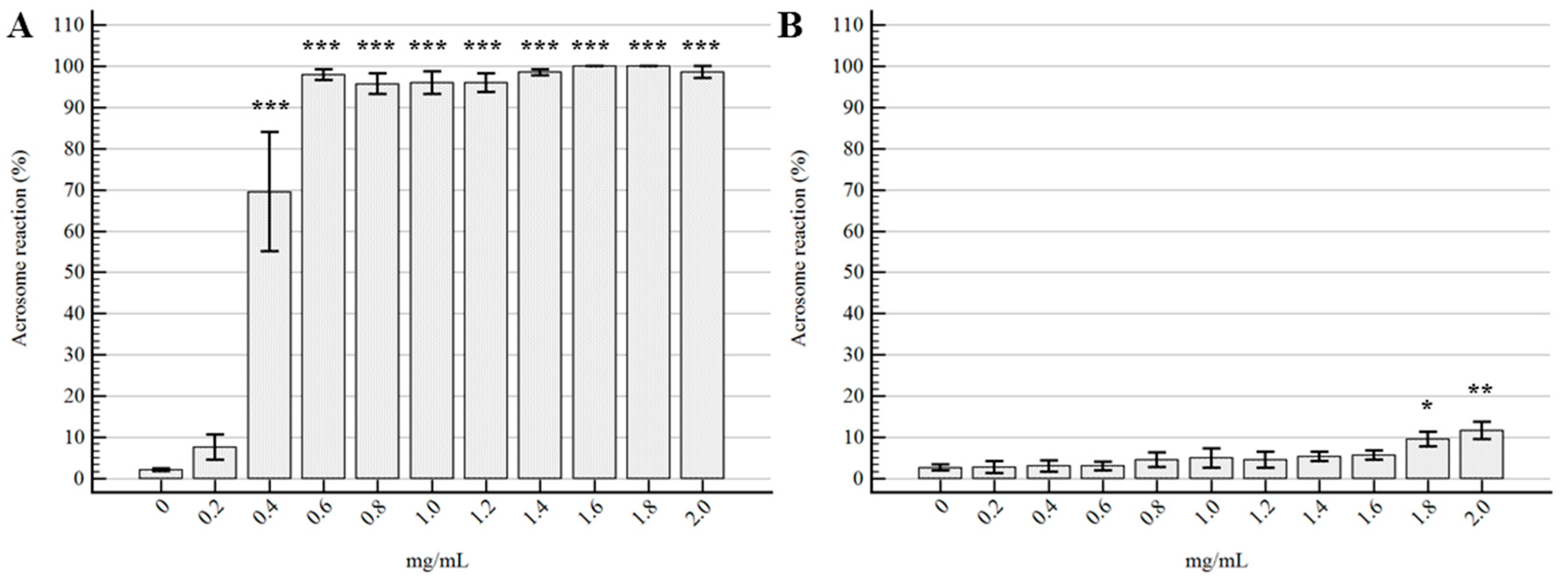
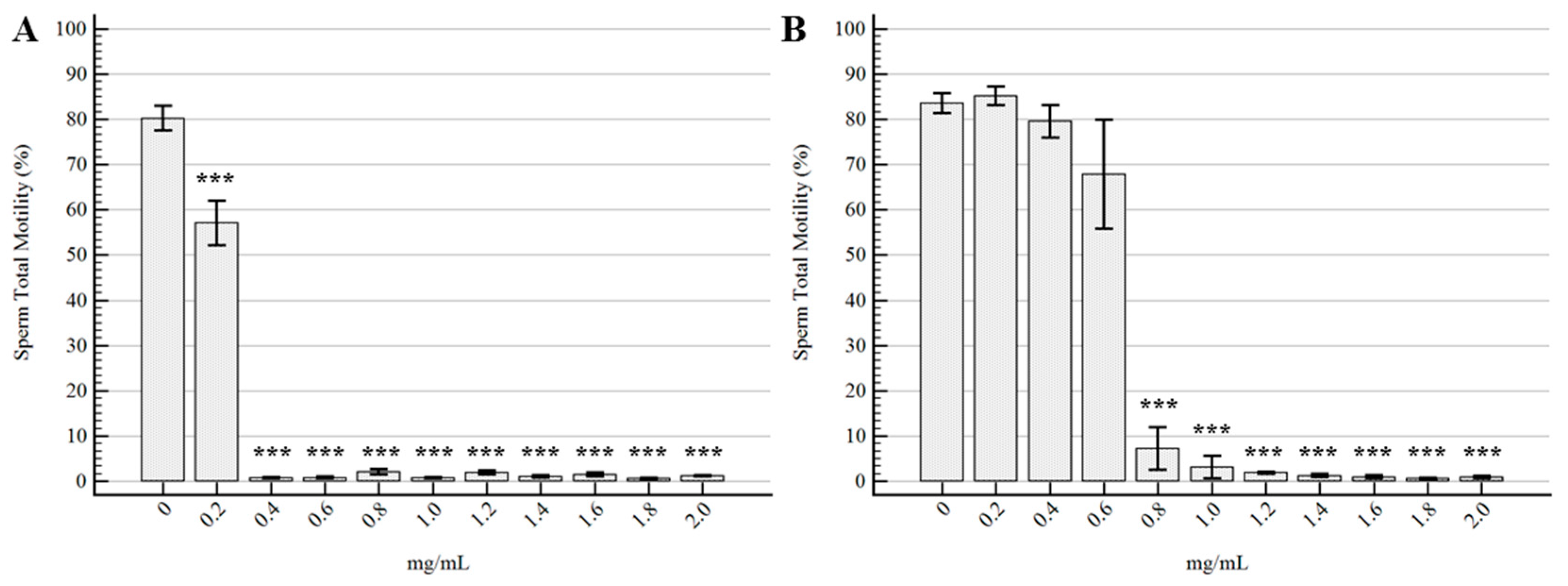
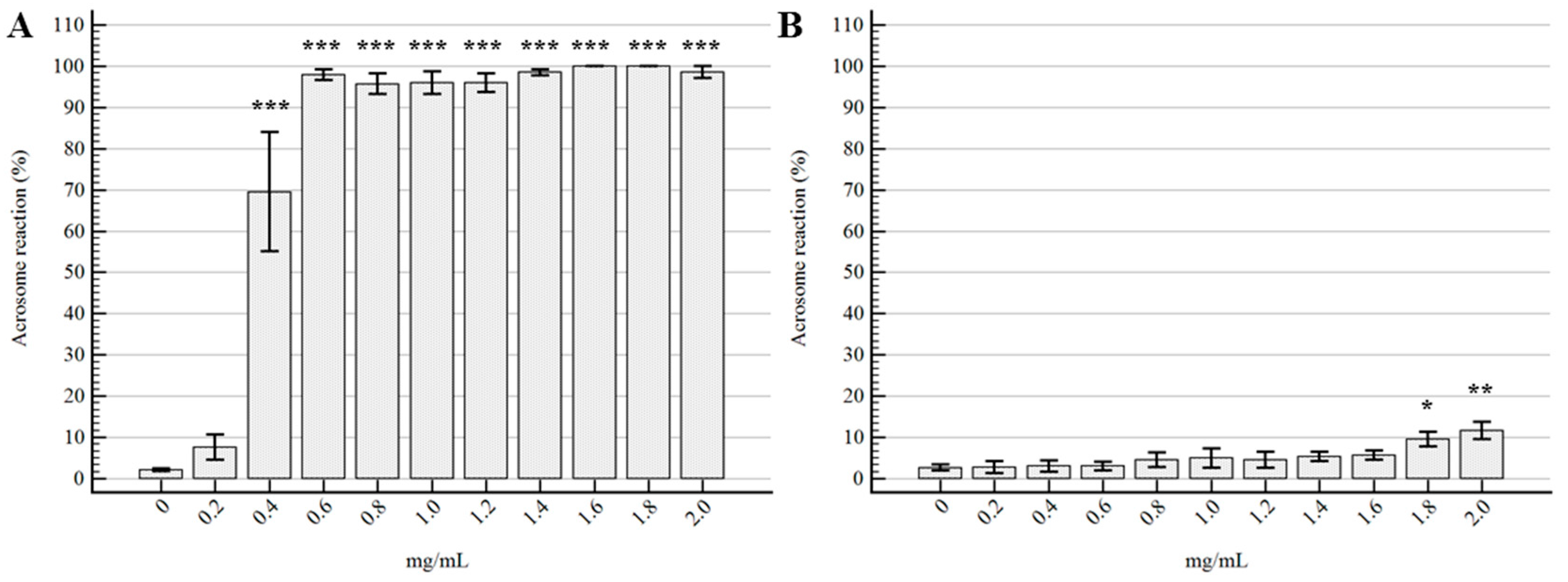
2.2. Semen Morpho-Functional Evaluations
2.3. Morphological Evaluation by SEM
3. Discussion
4. Materials and Methods
4.1. Chemo-Characterization of the EOs
4.1.1. Gas Chromatography-Mass Detector (GC-MS) Analysis
4.1.2. Gas Chromatography-Flame Ionization Detector (GC-FID) Analysis
4.1.3. Qualitative and Semi-Quantitative Analysis
4.2. Boars and Ejaculate Collection
4.3. Experimental Protocol
4.4. Semen Morpho-Functional Evaluations
4.5. Scanning Electron Microscopy (SEM)
4.6. Statistical Methods
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chikhoune, A.; Stouvenel, L.; Iguer-Ouada, M.; Hazzit, M.; Schmitt, A.; Lorès, P.; Wolf, J.P.; Aissat, K.; Auger, J.; Vaiman, D.; et al. In Vitro effects of Thymus munbyanus essential oil and thymol on human sperm motility and function. Reprod. Biomed. Online 2015, 31, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of lavender oil and its major components to human skin cells. Cell Prolif. 2004, 37, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Bag, A.; Chattopadhyay, R.R. Evaluation of Synergistic Antibacterial and Antioxidant Efficacy of Essential Oils of Spices and Herbs in Combination. PLoS ONE 2015, 10, e0131321. [Google Scholar] [CrossRef] [PubMed]
- Freires, I.A.; Denny, C.; Benso, B.; de Alencar, S.M.; Rosalen, P.L. Antibacterial Activity of Essential Oils and Their Isolated Constituents against Cariogenic Bacteria: A Systematic Review. Molecules 2015, 20, 7329–7358. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Kamiya, M.; Hayashi, T. Virucidal effects of the steam distillate from Houttuynia cordata and its components on HSV-1, influenza virus, and HIV. Planta Med. 1995, 61, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, P.; Schuhmacher, A.; Astani, A.; Reichling, J. Melissa officinalis oil affects infectivity of enveloped herpesviruses. Phytomedicine 2008, 15, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Tampieri, M.P.; Galuppi, R.; Macchioni, F.; Carelle, M.S.; Falcioni, L.; Cioni, P.L.; Morelli, I. The inhibition of Candida albicans by selected essential oils and their major components. Mycopathologia 2005, 159, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V.; Nielsen, J.B. A review of the toxicity of Melaleuca alternifolia (tea tree) oil. Food Chem. Toxicol. 2006, 44, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Kang, S.C. Studies on the viability and membrane integrity of human spermatozoa treated with essential oil of Trachyspermum ammi (L.) Sprague ex Turrill fruit. Andrologia 2012, 44 (Suppl. 1), 117–125. [Google Scholar] [CrossRef] [PubMed]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and electron microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H. Review: In vitro spermicidal tests. Contraception 1996, 54, 131–147. [Google Scholar] [CrossRef]
- EUR-Lex-52015XC0911(01)-EN-EUR-Lex. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52015XC0911%2801%29 (accessed on 11 October 2017).
- Hung, P.H.; Baumber, J.; Meyers, S.A.; VandeVoort, C.A. Effects of environmental tobacco smoke in vitro on rhesus monkey sperm function. Reprod. Toxicol. 2007, 23, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.A.; La Vignera, S.; Giacone, F.; Iacoviello, L.; Vicari, E.; Mongioi’, L.; Calogero, A.E. In vitro effects of nicotine on sperm motility and bio-functional flow cytometry sperm parameters. Int. J. Immunopathol. Pharmacol. 2013, 26, 739–746. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, O.J.; Vassilev, A.; Uckun, F.M. Evaluation of boar sperm as a model system to study the mechanism of spermicidal activity of vanadocenes. Biochem. Biophys. Res. Commun. 2000, 270, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Althouse, G. Artificial insemination in swine: Boar stud management. In Current Therapy in Large Animal Theriogenology; Youngquist, R.S., Threlfall, W.R., Eds.; Elsevier Health Sciences: New York, NY, USA, 2006; pp. 731–738. ISBN 978-1-4377-1340-4. [Google Scholar]
- Elmi, A.; Ventrella, D.; Barone, F.; Benvenuti, S.; Scozzoli, M.; Bacci, M.L. Preliminary evaluations of the effects of Cuminum cyminum and Coriandrum sativum essential oils on swine spermatozoa. JSM Sex. Med. 2017, 2, 1012. [Google Scholar]
- Satyal, P.; Jones, T.H.; Lopez, E.M.; McFeeters, R.L.; Ali, N.A.; Mansi, I.; Al-Kaf, A.G.; Setzer, W.N. Chemotypic Characterization and Biological Activity of Rosmarinus officinalis. Foods 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Bounatirou, S.; Smiti, S.; Miguel, M.G.; Faleiro, L.; Rejeb, M.N.; Neffati, M.; Costa, M.M.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Chemical composition, antioxidant and antibacterial activities of the essential oils isolated from Tunisian Thymus capitatus Hoff. et Link. Food Chem. 2007, 105, 146–155. [Google Scholar] [CrossRef]
- Carrasco, A.; Perez, E.; Cutillas, A.-B.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Origanum vulgare and Thymbra capitata Essential Oils from Spain: Determination of Aromatic Profile and Bioactivities. Nat. Prod. Commun. 2016, 11, 113–120. [Google Scholar] [PubMed]
- Sathananthan, P.A.H.; Deen, F. Microscopic Images of Human Sperm, Oocytes and Embryos for Assisted Reproductive Technology Including Embryonic Stem Cells: (Multi-Author); Henry Sathananthan, A., Ed.; Professor Arunachalam Henry Sathananthan: Melbourne, Australia, 2014; ISBN 978-0-9942165-0-2. [Google Scholar]
- Gallucci, M.N.; Oliva, M.; Casero, C.; Dambolena, J.; Luna, A.; Zygadlo, J.; Demo, M. Antimicrobial combined action of terpenes against the food-borne microorganisms Escherichia coli, Staphylococcus aureus and Bacillus cereus. Flavour Fragr. J. 2009, 24, 348–354. [Google Scholar] [CrossRef]
- Björndahl, L.; Söderlund, I.; Johansson, S.; Mohammadieh, M.; Pourian, M.R.; Kvist, U. Why the WHO recommendations for eosin-nigrosin staining techniques for human sperm vitality assessment must change. J. Androl. 2004, 25, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Broekhuijse, M.L.; Šoštarić, E.; Feitsma, H.; Gadella, B.M. Application of computer-assisted semen analysis to explain variations in pig fertility. J. Anim. Sci. 2012, 90, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Chaftar, N.; Girardot, M.; Quellard, N.; Labanowski, J.; Ghrairi, T.; Hani, K.; Frère, J.; Imbert, C. Activity of Six Essential Oils Extracted from Tunisian Plants against Legionella pneumophila. Chem. Biodivers. 2015, 12, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corp: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- Fantinati, P.; Zannoni, A.; Bernardini, C.; Forni, M.; Tattini, A.; Seren, E.; Bacci, M.L. Evaluation of swine fertilisation medium (SFM) efficiency in preserving spermatozoa quality during long-term storage in comparison to four commercial swine extenders. Animal 2009, 3, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Barone, F.; Ventrella, D.; Zannoni, A.; Forni, M.; Bacci, M.L. Can Microfiltered Seminal Plasma Preserve the Morphofunctional Characteristics of Porcine Spermatozoa in the Absence of Antibiotics? A Preliminary Study. Reprod. Domest. Anim. Zuchthyg. 2016, 51, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Bacci, M.L.; Zannoni, A.; De Cecco, M.; Fantinati, P.; Bernardini, C.; Galeati, G.; Spinaci, M.; Giovannoni, R.; Lavitrano, M.; Seren, E.; et al. Sperm-mediated gene transfer-treated spermatozoa maintain good quality parameters and in vitro fertilization ability in swine. Theriogenology 2009, 72, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Bucci, D.; Isani, G.; Giaretta, E.; Spinaci, M.; Tamanini, C.; Ferlizza, E.; Galeati, G. Alkaline phosphatase in boar sperm function. Andrology 2014, 2, 100–106. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | LRI 1 | Area % |
|---|---|---|
| α-Thujene | 927 | 0.71 |
| α-Pinene | 933 | 0.96 |
| Camphene | 948 | 0.15 |
| β-Pinene | 976 | 0.13 |
| β-Myrcene | 993 | 1.37 |
| α-Phellandrene | 1006 | 0.16 |
| α-Terpinene | 1017 | 1.31 |
| p-Cymene | 1026 | 12.28 |
| Limonene | 1029 | 0.45 |
| γ-Terpinene | 1060 | 5.62 |
| trans-sabinene hydrate | 1067 | 0.09 |
| α-Terpinolene | 1089 | 0.21 |
| Linalool | 1102 | 2.37 |
| Borneol | 1167 | 0.18 |
| Terpinen-4-ol | 1179 | 0.7 |
| Thymol | 1296 | 3.49 |
| Carvacrol | 1312 | 65.2 |
| β-Caryophyllene | 1426 | 1.92 |
| α-Humulene | 1456 | 0.11 |
| Caryophyllene oxide | 1594 | 0.12 |
| Total | 97.54 |
| Compounds | LRI 1 | Area % |
|---|---|---|
| α-Pinene | 936 | 23.55 |
| Camphene | 949 | 10.16 |
| β-Pinene | 977 | 5.39 |
| β-Myrcene | 993 | 1.88 |
| p-Cymene | 1026 | 2.8 |
| 1,8-Cineole | 1034 | 21.36 |
| Linalool | 1103 | 0.96 |
| Camphor | 1145 | 22.03 |
| Borneol | 1168 | 2.84 |
| Terpinen-4-ol | 1180 | 0.05 |
| α-Terpineol | 1193 | 2.45 |
| Bornyl acetate | 1290 | 1.38 |
| β-Caryophyllene | 1427 | 1.19 |
| Caryophyllene oxide | 1594 | 0.17 |
| Total | 96.22 |
| Parameters | T. capitata EO | R. officinalis EO |
|---|---|---|
| β (95% C.I.) p Value | β (95% C.I.) p Value | |
| V % | −0.013 (−0.017; −0.009) p < 0.0001 | −0.018 (−0.022; −0.015) p < 0.0001 |
| TotM % | −0.015 (−0.020: −0.010) p < 0.0001 | −0.014 (−0.017; −0.012) p < 0.0001 |
| ProgM % | −0.027 (−0.037; −0.016) p < 0.0001 | −0.026 (−0.031; −0.021) p < 0.0001 |
| AR % | 0.013 (0.010; 0.016) p < 0.0001 | 0.086 (0.052; 0.119) p < 0.0001 |
| pH | 0.508 (−1.168; 2.185) p = 0.546 | −0.018 (−2.275; 2.238) p = 0.987 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmi, A.; Ventrella, D.; Barone, F.; Filippini, G.; Benvenuti, S.; Pisi, A.; Scozzoli, M.; Bacci, M.L. Thymbra capitata (L.) Cav. and Rosmarinus officinalis (L.) Essential Oils: In Vitro Effects and Toxicity on Swine Spermatozoa. Molecules 2017, 22, 2162. https://doi.org/10.3390/molecules22122162
Elmi A, Ventrella D, Barone F, Filippini G, Benvenuti S, Pisi A, Scozzoli M, Bacci ML. Thymbra capitata (L.) Cav. and Rosmarinus officinalis (L.) Essential Oils: In Vitro Effects and Toxicity on Swine Spermatozoa. Molecules. 2017; 22(12):2162. https://doi.org/10.3390/molecules22122162
Chicago/Turabian StyleElmi, Alberto, Domenico Ventrella, Francesca Barone, Gianfranco Filippini, Stefania Benvenuti, Annamaria Pisi, Maurizio Scozzoli, and Maria L. Bacci. 2017. "Thymbra capitata (L.) Cav. and Rosmarinus officinalis (L.) Essential Oils: In Vitro Effects and Toxicity on Swine Spermatozoa" Molecules 22, no. 12: 2162. https://doi.org/10.3390/molecules22122162
APA StyleElmi, A., Ventrella, D., Barone, F., Filippini, G., Benvenuti, S., Pisi, A., Scozzoli, M., & Bacci, M. L. (2017). Thymbra capitata (L.) Cav. and Rosmarinus officinalis (L.) Essential Oils: In Vitro Effects and Toxicity on Swine Spermatozoa. Molecules, 22(12), 2162. https://doi.org/10.3390/molecules22122162