Antibacterial Activity of Polyphenols: Structure-Activity Relationship and Influence of Hyperglycemic Condition




Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of the Glycation Products
2.2. Antibacterial Susceptibilities of Polyphenols
2.3. Minimum Inhibitory Concentration (MIC) Values and Structure-Activity Relationships
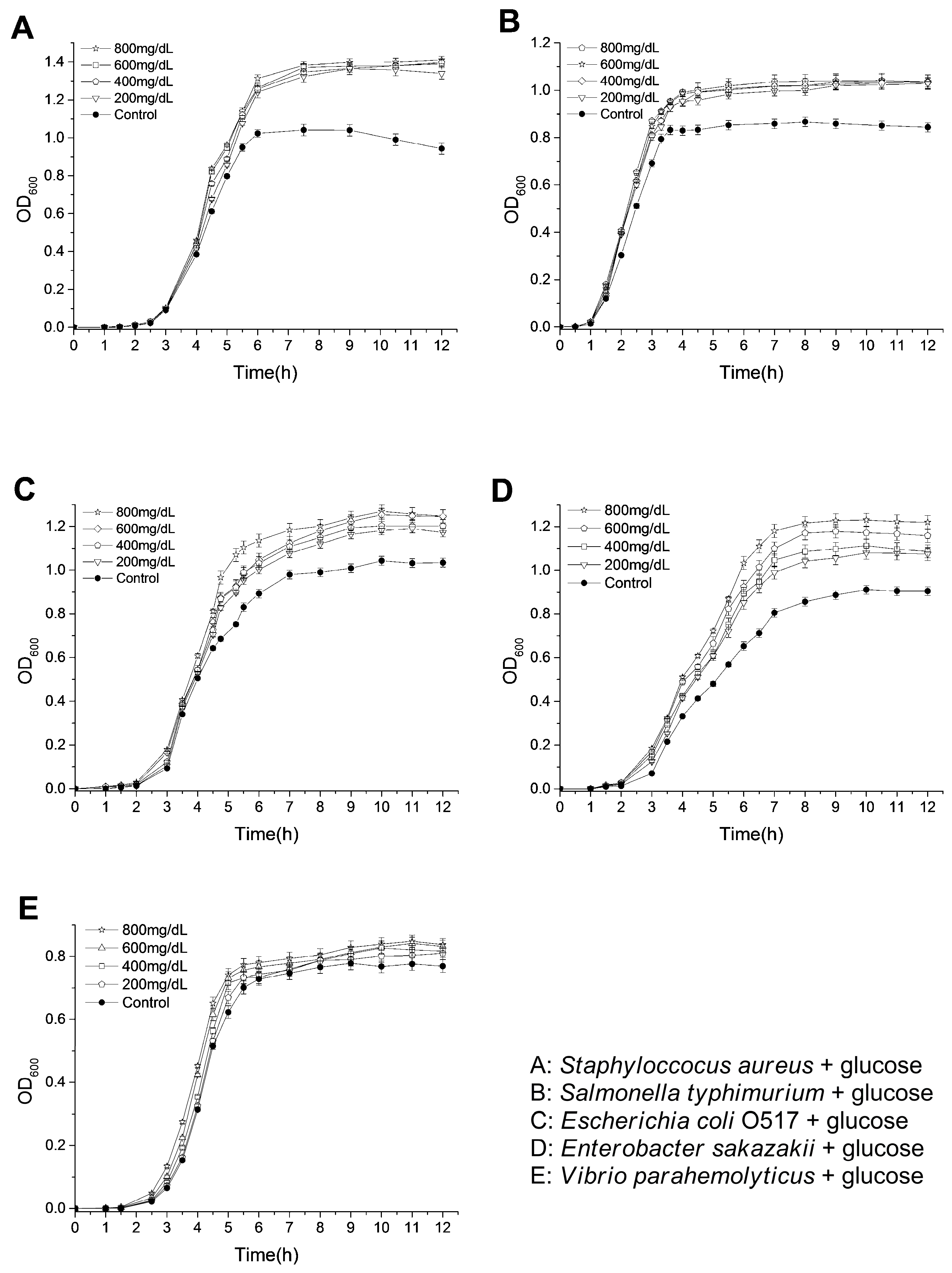
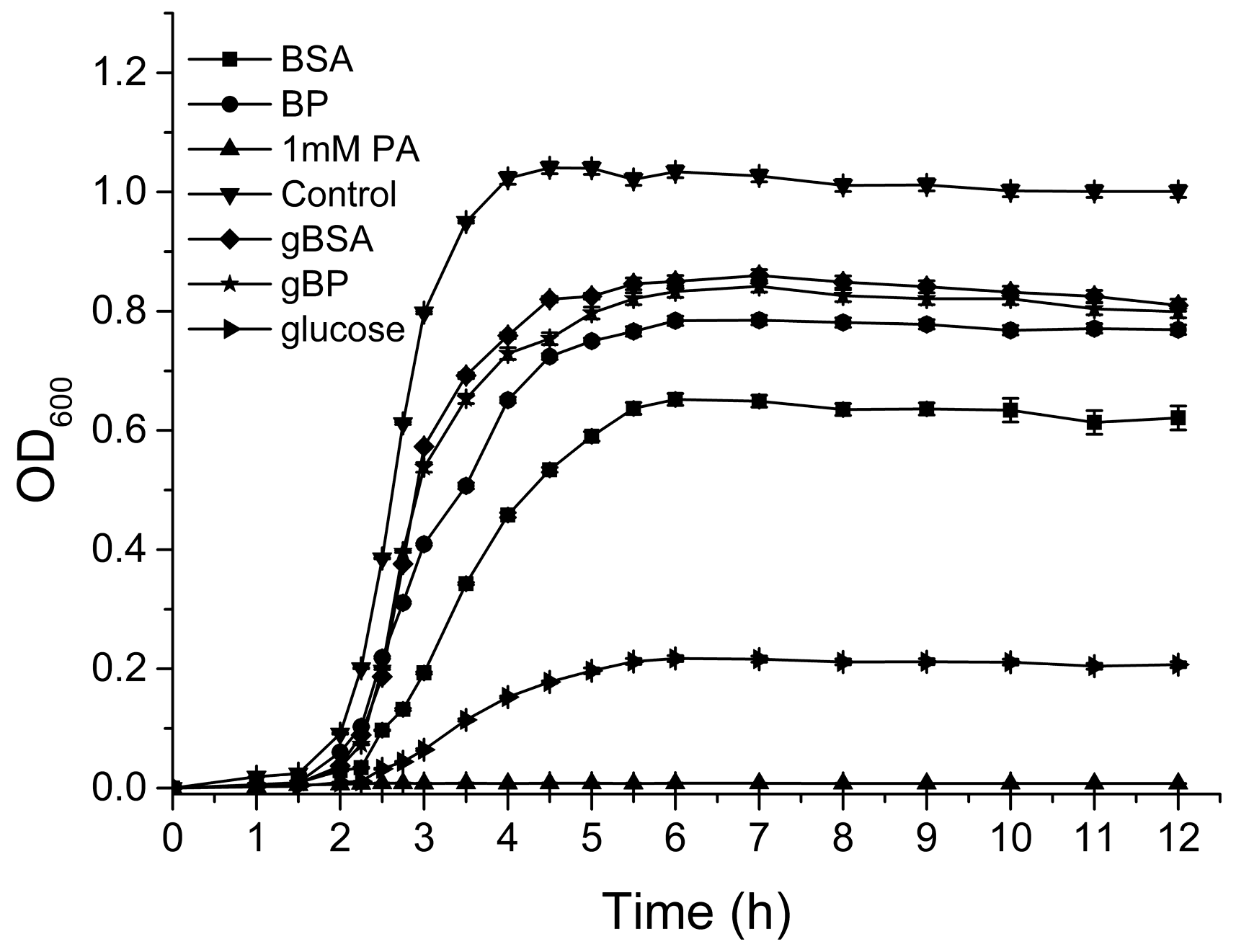
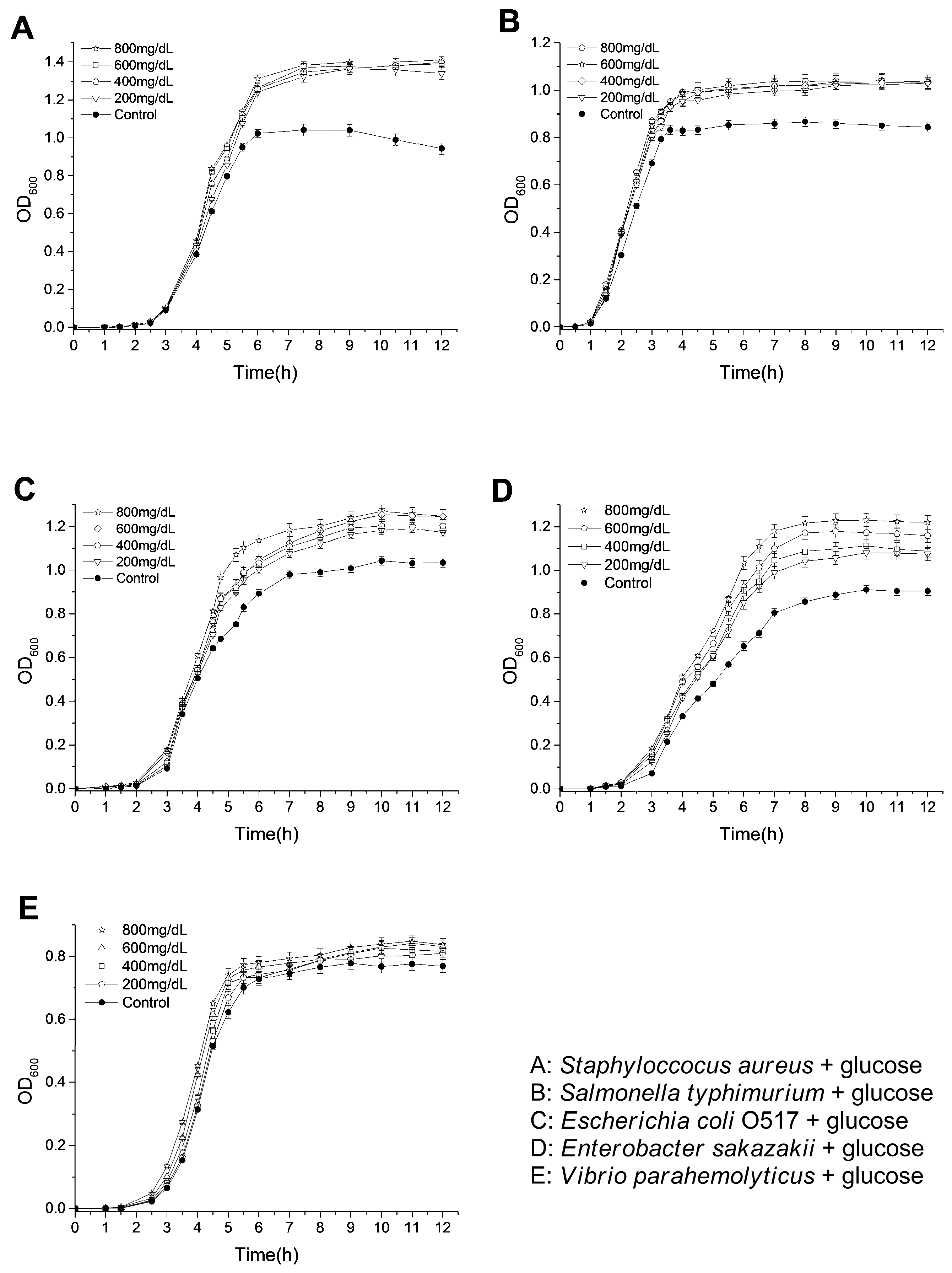
2.4. Effects of Glucose on the Bacterial Growth
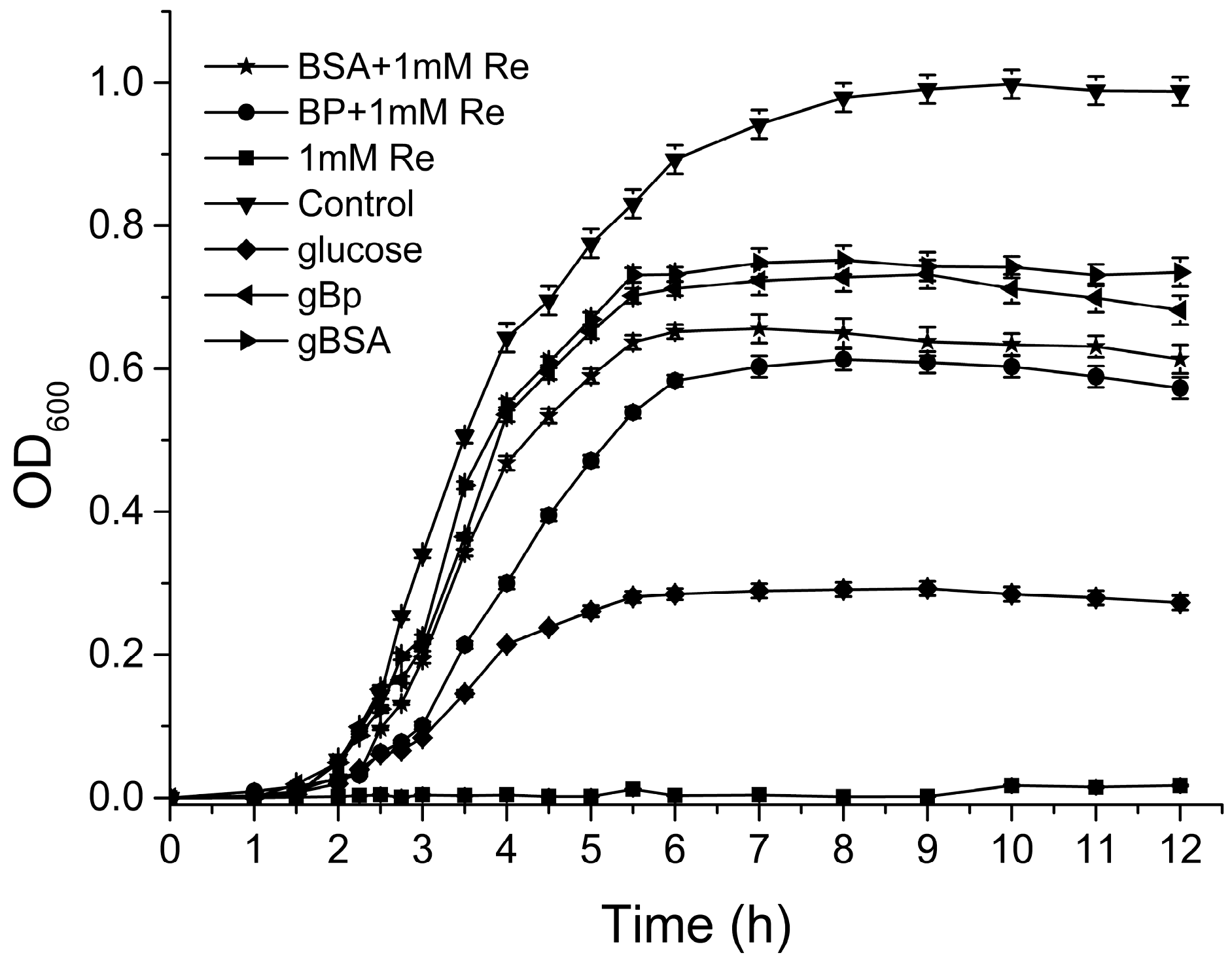
2.5. Antibacterial Activities of Polyphenols under Hyperglycemic Conditions
2.6. Effects of Polyphenols on the Growth of Bacteria under Hyperglycemic Conditions
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Apparatus
3.3. Glycations of BSA and BP
3.4. Determination of the Glycation Products
3.5. Antibacterial Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [PubMed]
- Xiao, J.B. Polyphenol-Plasma Proteins Interaction: Its Nature, Analytical Techniques, and Influence on Bioactivities of Polyphenols. Curr. Drug Metab. 2013, 14, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Kai, G. A Review of Dietary Polyphenol-Plasma Protein Interactions: Characterization, Influence on the Bioactivity, and Structure-Affinity Relationship. Crit. Rev. Food Sci. Nutr. 2012, 52, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Polyphenols and health: Update and perspectives. Arch. Biochem. Biophys. 2010, 501, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Varoni, E.M.; Lodi, G.; Sardella, A.; Carrassi, A.; Iriti, M. Plant Polyphenols and Oral Health: Old Phytochemicals for New Fields. Curr. Med. Chem. 2012, 19, 1706–1720. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.C.W.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [PubMed]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic biochemical mechanisms behind the health benefits of polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, X. Structures Required of Polyphenols for Inhibiting Advanced Glycation end Products Formation. Curr. Drug Metab. 2013, 14, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Chen, X.; Xiao, J. Inhibition of flavonoids on acetylcholine esterase: Binding and structure-activity relationship. Food Funct. 2014, 5, 2582–2589. [Google Scholar] [CrossRef] [PubMed]
- Tangney, C.C.; Rasmussen, H.E. Polyphenols, Inflammation, and Cardiovascular Disease. Curr. Atheroscler. Rep. 2013, 15. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Kerner, W.; Bruckel, J. Definition, classification and diagnosis of diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2014, 122, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Pitocco, D.; Tesauro, M.; Alessandro, R.; Ghirlanda, G.; Cardillo, C. Oxidative Stress in Diabetes: Implications for Vascular and Other Complications. Int. J. Mol. Sci. 2013, 14, 21525–21550. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.; Gorter, K.J.; Hak, E.; Goudzwaard, W.L.; Schellevis, F.G.; Hoepelman, A.I.M.; Rutten, G. Increased risk of common infections in patients with type 1 and type 2 diabetes Mellitus. Clin. Infect. Dis. 2005, 41, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.J.; Martin, N.; Makepeace, A.; Sillars, B.A.; Davis, W.A.; Davis, T.M.E. Incidence and Predictors of Hospitalization for Bacterial Infection in Community-Based Patients with Type 2 Diabetes: The Fremantle Diabetes Study. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Tseng, C.C.; Wu, A.B.; Lin, W.H.; Teng, C.H.; Yan, J.J.; Wu, J.J. Bacterial characteristics and glycemic control in diabetic patients with Escherichia coli urinary tract infection. J. Microbiol. Immunol. Infect. 2013, 46, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Xie, Y.; Chen, X. Type 2 diabetes diminishes the benefits of dietary antioxidants: Evidence from the different free radical scavenging potential. Food Chem. 2015, 186, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.X.; Xiao, J.B.; Kai, G.Y.; Chen, X.Q. Glycation of plasma proteins in type II diabetes lowers the non-covalent interaction affinities for dietary polyphenols. Integr. Biol. 2012, 4, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Huang, S.-M.; Lin, J.-A.; Yen, G.-C. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011, 2, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Schmitt, J.; Munch, G.; Gasic-Milencovic, J. Characterization of advanced glycation end products for biochemical studies: Side chain modifications and fluorescence characteristics. Anal. Biochem. 2005, 338, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, R.; Carrizo Flores, R.; Ocampos, S.; Lucatti, A.; Flores Galleguillo, L.; Tonn, C.; Sosa, V. Flavonoids from Pterocaulon alopecuroides with Antibacterial Activity. Planta Med. 2008, 74, 1463–1467. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Chen, X.; Ren, L. Antibacterial Activities of Flavonoids: Structure-Activity Relationship and Mechanism. Curr. Med. Chem. 2014, 22, 132–149. [Google Scholar] [CrossRef]
- Geerlings, S.E.; Brouwer, E.C.; Gaastra, W.; Verhoef, J.; Hoepelman, A.I. Effect of glucose and pH on uropathogenic and non-uropathogenic Escherichia coli: Studies with urine from diabetic and non-diabetic individuals. J. Med. Microbiol. 1999, 48, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Garnett, J.P.; Baker, E.H.; Naik, S.; Lindsay, J.A.; Knight, G.M.; Gill, S.; Tregoning, J.S.; Baines, D.L. Metformin reduces airway glucose permeability and hyperglycaemia-induced Staphylococcus aureus load independently of effects on blood glucose. Thorax 2013, 68, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Stingley, R.L.; Liu, H.; Mullis, L.B.; Elkins, C.A.; Hart, M.E. Staphylococcus aureus toxic shock syndrome toxin-1 (TSST-1) production and Lactobacillus species growth in a defined medium simulating vaginal secretions. J. Microbiol. Methods 2014, 106, 57–66. [Google Scholar] [CrossRef] [PubMed]
- You, Y.B.; Xue, T.; Cao, L.Y.; Zhao, L.P.; Sun, H.P.; Sim, B.L. Staphylococcus aureus glucose-induced biofilm accessory proteins, GbaAB, influence biofilm formation in a PIA-dependent manner. Int. J. Med. Microbiol. 2014, 304, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.D.; Rowley, G.; Hinton, J.C.D.; Thompson, A. Glucose and Glycolysis Are Required for the Successful Infection of Macrophages and Mice by Salmonella enterica Serovar Typhimurium. Infect. Immun. 2009, 77, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Hironaka, I.; Iwase, T.; Sugimoto, S.; Okuda, K.-I.; Tajima, A.; Yanaga, K.; Mizunoe, Y. Glucose Triggers ATP Secretion from Bacteria in a Growth-Phase-Dependent Manner. Appl. Environ. Microbiol. 2013, 79, 2328–2335. [Google Scholar] [CrossRef] [PubMed]
- Zeitlinger, M.A.; Derendorf, H.; Mouton, J.W.; Cars, O.; Craig, W.A.; Andes, D.; Theuretzbacher, U. Protein Binding: Do We Ever Learn? Antimicrob. Agents Chemother. 2011, 55, 3067–3074. [Google Scholar] [CrossRef] [PubMed]
- Burian, A.; Wagner, C.; Stanek, J.; Manafi, M.; Boehmdorfer, M.; Jager, W.; Zeitlinger, M. Plasma protein binding may reduce antimicrobial activity by preventing intra-bacterial uptake of antibiotics, for example clindamycin. J. Antimicrob. Chemother. 2011, 66, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Or, I.; Katz, C.; Ron, E.Z. AGEs Secreted by Bacteria Are Involved in the Inflammatory Response. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Molecular mechanisms of antibiotic resistance. Chem. Commun. 2011, 47, 4055–4061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, S.; Chen, F.; Wang, M. Treatment of proteins with dietary polyphenols lowers the formation of AGEs and AGE-induced toxicity. Food Funct. 2014, 5, 2656–2661. [Google Scholar] [CrossRef] [PubMed]
- Zwang, T.J.; Gormally, M.V.; Johal, M.S.; Sazinsky, M.H. Enhanced iron availability by protein glycation may explain higher infection rates in diabetics. Biometals 2012, 25, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Bhatwadekar, A.D.; Ghole, V.S. Rapid method for the preparation of an AGE-BSA standard calibrator using thermal glycation. J. Clin. Lab. Anal. 2005, 19, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.N.; Metcalf, P.A.; Baker, J.R. Fructosamine: A new approach to the estimation of serum glycosylprotein. An index of diabetic control. Clin. Chim. Acta 1983, 127, 87–95. [Google Scholar] [CrossRef]
- Eloff, J.N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta Med. 1998, 64, 711–713. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |

{kind=link}
{kind=link}
{kind=link}
| Samples | Fluorescence Intensities (FI 460 nm) | UV Absorbance | EDMF (mM) | ||
|---|---|---|---|---|---|
| 330 nm | 360 nm | 400 nm | |||
| BSA | 22 * | 0.054 | 0.023 | 0.006 | 1.84 |
| gBSA | 390 | 0.129 | 0.071 | 0.021 | 20.71 |
| BP | 12 | 0.024 | 0.012 | 0.003 | 1.04 |
| gBP | 95 | 0.079 | 0.034 | 0.011 | 3.23 |
| Subclass | (1 μM)/Bacteria | Subtitutions | Inhibition Zone (Φ, mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| OH | OCH3 | Others | EC | SA | ST | ES | VP | ||
| Flavones | Flavone | 9 * | 8 | 8 | 8 | 9 | |||
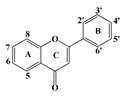 | 6-hydroxyflavone | 6 | - | - | - | - | - | ||
| 6-mehoxyflavone | 6 | - | - | - | - | - | |||
| 7-hydroxyflavone | 7 | - | - | - | - | - | |||
| 3-hydroxyflavone | 3 | - | - | - | - | - | |||
| Bacalein | 5, 6, 7 | 8 | 10 | - | - | 9 | |||
| Bacalin | 5, 6 | 7-β-d-glucuronide | - | - | - | - | 10 | ||
| Chrysin | 5, 7 | - | - | - | - | - | |||
| Apigenin | 5, 7, 4′ | - | - | - | - | - | |||
| Luteolin | 5, 7, 3′, 4′ | - | - | - | - | - | |||
| Flavanones | Flavanone | - | - | - | - | - | |||
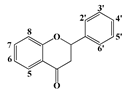 | 6-hydroxyflavanone | 6 | - | - | - | - | - | ||
| 4′-hydroxyflavanone | 4′ | - | - | - | - | - | |||
| 6-methoxyflavanone | 6 | - | - | - | - | - | |||
| Narigenin | 5, 7, 4′ | 8 | 11 | 8 | 9.5 | 11.5 | |||
| Narigin | 5, 4′ | 7-neohesperidose | - | - | - | - | - | ||
| Hesperitin | 5, 7, 3′ | 4′ | 8.5 | - | 8 | 8.5 | 10 | ||
| Hesperidin | 5, 3′ | 4′ | 7-rutinose | - | - | - | - | - | |
| Taxifolin | 3, 5 ,7 ,3′ ,4′ ,5′ | 6.5 | 6.5 | 7.5 | - | 9.5 | |||
| Neohesperidin-dihydrochalcone | 5, 7, 4′ | - | - | - | - | - | |||
| Flavonols | Quercetin | 3, 5, 7, 3′, 4′ | 8 | 9 | 8 | 8 | - | ||
 | Myricetin | 3, 5, 7, 3′, 4′, 5′ | 8 | 10 | 8 | 8 | 12 | ||
| Quercetrin | 5, 7, 3′, 4′ | 3-o-β-d-Glucoside | - | - | - | - | - | ||
| Fisetin | 3, 7, 3′, 4′ | - | - | - | - | - | |||
| Rutin | 5, 7, 3′, 4′ | - | - | - | - | 8 | |||
| Isoflavones | Genistein | 5, 7, 4′ | - | - | - | - | 7 | ||
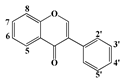 | Formononetin | 7 | 4′ | - | - | - | - | - | |
| Biochanin A | 5, 7 | 4′ | - | - | - | - | 7 | ||
| Puerarin | 7, 4′ | - | - | - | - | - | |||
| Dadazein | 7, 4′ | - | - | - | - | - | |||
| Dadazin | 7-glucoside | - | - | - | - | - | |||
| Stilbenes | Piceid | 5, 4′ | 3-glucoside | - | - | - | - | - | |
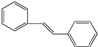 | |||||||||
| Resveratrol | 3, 5, 4′ | 11 | 9 | 9 | - | 12 | |||
| Phenolic acids | Gallic acid | R = H | 7 | - | 8 | - | - | ||
 | Propyl gallate | R = Propyl | 10 | 7 | - | 10 | 19 | ||
| Methyl gallate | R = Methyl | 9 | 7 | - | 8 | 12 | |||
| Ethyl gallate | R = Ethyl | 9 | - | - | 8 | 20 | |||
| Pyrogallic acid | 14 | 12 | 8 | 8 | 22 | ||||
| Antibiotics | Kana (0.5 μM) | 22 | 20 | 25 | 22 | 20 | |||
| Carbenicilin (0.5 μM) | 18 | 30 | 30 | 18 | 15 | ||||
| Polypehols/MIC (mmol/L) | E. coli O517 | S. aureus | ST | ES | VP |
|---|---|---|---|---|---|
| Flavone | 2 * | 2 | 2 | 2 | 1 |
| Bacalein | 0.5 | 1 | 1.5 | 1 | 0.5 |
| Bacalin | 2.5 | >2.5 | 2.5 | >2.5 | >2.5 |
| Quercetin | 2 | 2 | 2 | 2 | 1 |
| Myricetin | 0.5 | 1 | 2 | 2.5 | 0.5 |
| Taxin | >2.5 | >2.5 | >2.5 | >2.5 | 2 |
| Rutin | >2.5 | >2.5 | >2.5 | >2.5 | 2.5 |
| Narigenin | 2.5 | 2.5 | 2.5 | 2.5 | 1 |
| Hesperitin | 1 | >2.5 | >2.5 | >2.5 | 2 |
| Resveratrol | 1 | 2.5 | 2.5 | 2.5 | 1 |
| Gallic acid | >2.5 | >2.5 | >2.5 | >2.5 | 2.5 |
| Propyl gallate | 2 | >2.5 | >2.5 | 2.5 | 1.5 |
| Methyl gallate | 2.5 | >2.5 | >2.5 | >2.5 | 2.5 |
| Ethyl gallate | 2 | >2.5 | >2.5 | 2.5 | 1.5 |
| Pyrogallic acid | 1 | 1.5 | 1.5 | 1.5 | 0.5 |
| Kana | 0.1 | 0.25 | 0.25 | 0.1 | 0.05 |
| Carbenicilin | 0.1 | 0.25 | 0.25 | 0.1 | 0.05 |
| Bacteria | Polyphenol | Control | MIC (mmol/L) | ||||
|---|---|---|---|---|---|---|---|
| Glucose | BSA | gBSA | BP | gBP | |||
| E. coli O517 | Myricetin | 1 * | 1 | 1.5 | 1.5 | 1.5 | 2.5 |
| Bacalein | 0.5 | 0.5 | 0.5 | 0.75 | 0.75 | 1.5 | |
| Pyrogallic acid | 0.75 | 0.75 | 0.75 | 1 | 1 | 2 | |
| Resveratrol | 1 | 1.5 | 1.5 | 1.5 | 1.5 | 1.5 | |
| E. sakazakii | Pyrogallic acid | 1.5 | 2 | 1.5 | 2 | 2 | >2.5 |
| Bacalein | 0.5 | 1 | 1 | 1 | 1 | 2.5 | |
| S. typhimurium | Pyrogallic acid | 0.75 | 1 | 0.75 | 1.5 | 1 | 2.5 |
| Myricetin | 1 | 1 | 1 | 1.5 | 1.5 | 2 | |
| S. aureus | Pyrogallic acid | 1 | 1.5 | 1.5 | 2 | 1.5 | >2.5 |
| Bacalein | 0.25 | 0.5 | 0.5 | 0.75 | 0.5 | 2 | |
| V. parahemolyticus | Pyrogallic acid | 0.75 | 1 | 0.75 | 1.5 | 1 | >2.5 |
| Naringenin | 0.75 | 0.75 | 1 | 1 | 1 | >2.5 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Y.; Chen, J.; Xiao, A.; Liu, L. Antibacterial Activity of Polyphenols: Structure-Activity Relationship and Influence of Hyperglycemic Condition. Molecules 2017, 22, 1913. https://doi.org/10.3390/molecules22111913
Xie Y, Chen J, Xiao A, Liu L. Antibacterial Activity of Polyphenols: Structure-Activity Relationship and Influence of Hyperglycemic Condition. Molecules. 2017; 22(11):1913. https://doi.org/10.3390/molecules22111913
Chicago/Turabian StyleXie, Yixi, Jing Chen, Aiping Xiao, and Liangliang Liu. 2017. "Antibacterial Activity of Polyphenols: Structure-Activity Relationship and Influence of Hyperglycemic Condition" Molecules 22, no. 11: 1913. https://doi.org/10.3390/molecules22111913
APA StyleXie, Y., Chen, J., Xiao, A., & Liu, L. (2017). Antibacterial Activity of Polyphenols: Structure-Activity Relationship and Influence of Hyperglycemic Condition. Molecules, 22(11), 1913. https://doi.org/10.3390/molecules22111913