Anti-HIV Activity of Ocimum labiatum Extract and Isolated Pheophytin-a


Abstract
1. Introduction
2. Results
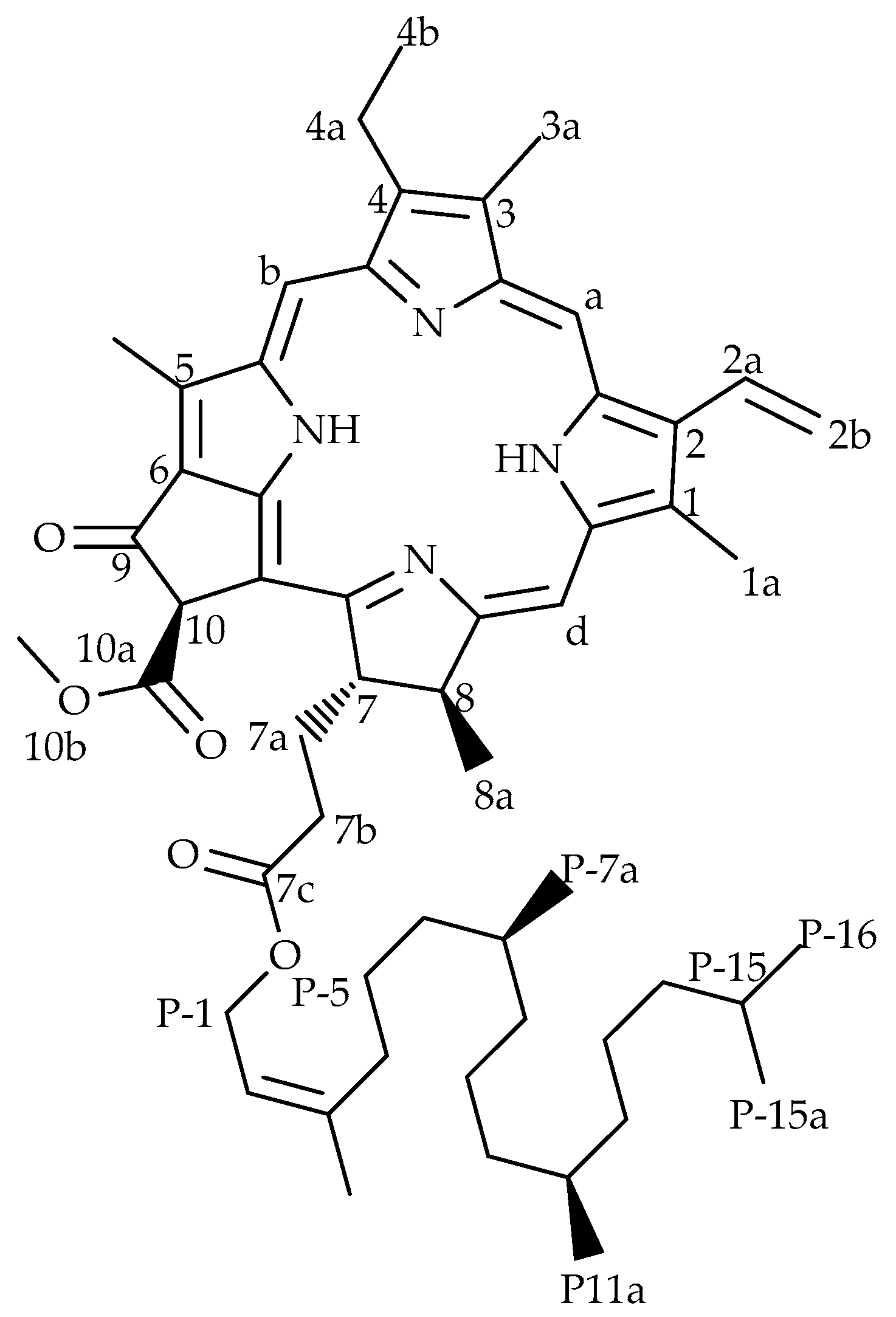
2.1. Structural Elucidation of Pheophytin-a
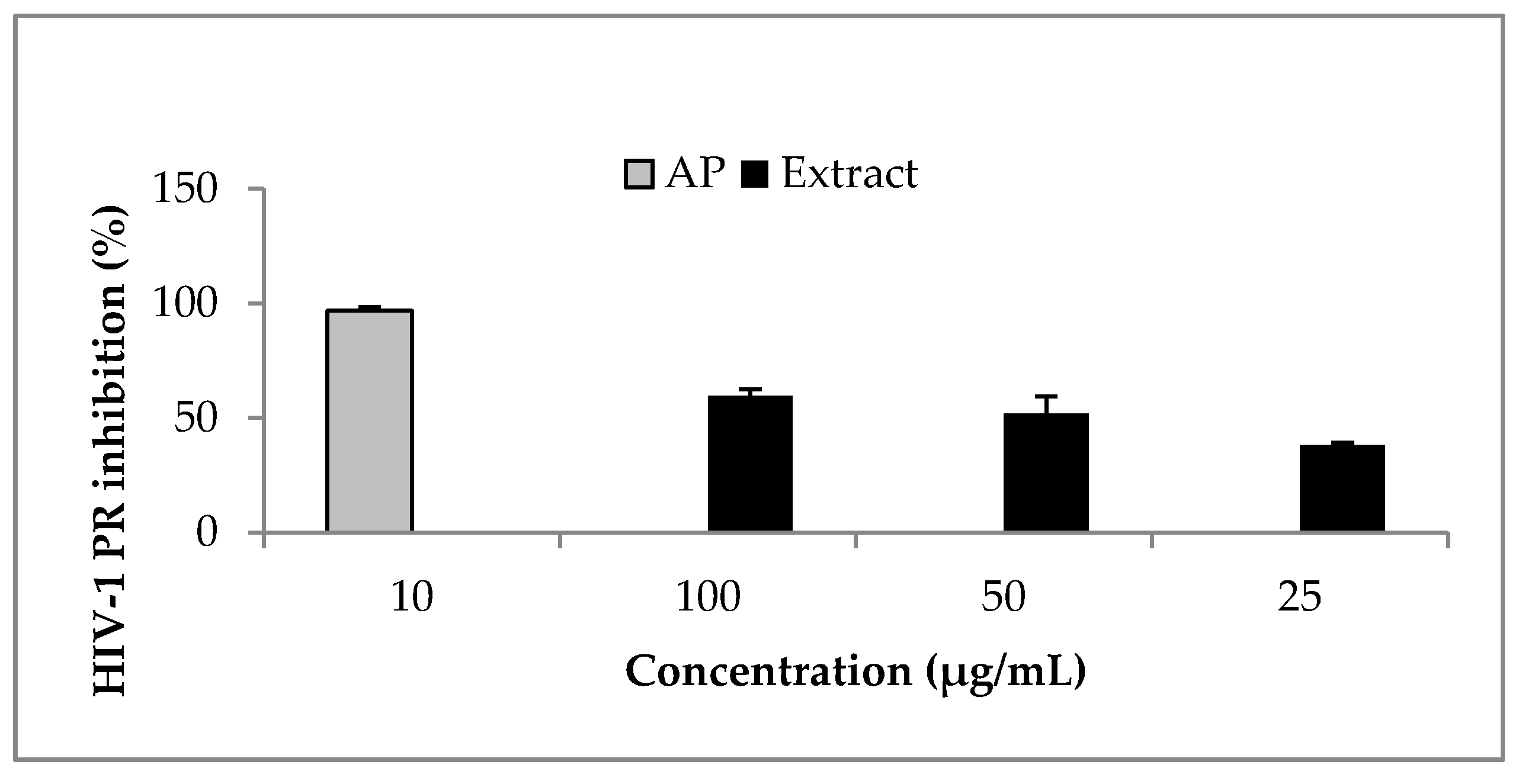
2.2. HIV-1 PR Inhibition
2.3. Inhibition of PMA-Induced HIV-1 Expression by O. labiatum
2.4. Cytotoxic Effect of O. labiatum Extract and Phy-a
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Compound Isolation
4.4. HIV-1 PR Assay
4.5. HIV-1 RT Assay
4.6. HIV-1 p24 Antigen ELISA
4.7. Cells
Cytotoxicity Analysis of the Extract and Isolated Compound
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.-C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Annu. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.A.; Lewis, W. Aging and HIV/AIDS: Pathogenetic role of therapeutic side effects. Lab. Investig. 2014, 94, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Kebaabetswe, P.M.; Stirratt, M.J.; McLellan-Lemal, E.; Henderson, F.L.; Gray, S.C.; Rose, C.E.; Williams, T.; Paxton, L.A. Factors associated with adherence and concordance between measurement strategies in an HIV daily oral tenofovir/emtricitibine as pre-exposure prophylaxis (Prep) clinical trial, Botswana, 2007–2010. AIDS Behav. 2015, 19, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Wensing, A.M.; Calvez, V.; Gunthard, H.F.; Johnson, V.A.; Paredes, R.; Pillay, D.; Shafer, R.W.; Richman, D.D. Special contribution 2014 Update of the drug resistance mutations in HIV-1. Top. Antivir. Med. 2014, 22, 642–650. [Google Scholar] [PubMed]
- Perera, C.; Efferth, T. Antiviral medicinal herbs and phytochemicals. J. Pharmacogn. 2012, 3, 45–48. [Google Scholar]
- Wang, S.-Y.; Tseng, C.-P.; Tsai, K.-C.; Lin, C.-F.; Wen, C.-Y.; Tsay, H.-S.; Sakamoto, N.; Tseng, C.-H.; Cheng, J.-C. Bioactivity-guided screening identifies pheophytin a as a potent anti-hepatitis C virus compound from Lonicera hypoglauca Miq. Biochem. Biophys. Res. Commun. 2009, 385, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-J.; Tan, G.T.; Hoang, V.D.; Hung, N.V.; Cuong, N.M.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.S. Natural anti-HIV agents. Part IV. Anti-HIV constituents from Vatica cinerea. J. Nat. Prod. 2003, 66, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Bouslama, L.; Hayashi, K.; Lee, J.-B.; Ghorbel, A.; Hayashi, T. Potent virucidal effect of pheophorbide a and pyropheophorbide a on enveloped viruses. J. Nat. Med. 2011, 65, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Labra, M.; Miele, M.; Ledda, B.; Grassi, F.; Mazzei, M.; Sala, F. Morphological characterization, essential oil composition and DNA genotyping of Ocimum basilicum L. cultivars. Plant Sci. 2004, 167, 725–731. [Google Scholar] [CrossRef]
- Chandrasekaran, C.V.; Srikanth, H.S.; Anand, M.S.; Allan, J.J.; Viji, M.M.H.; Amit, A. Evaluation of the mutagenic potential and acute oral toxicity of standardized extract of Ocimum sanctum. Hum. Exp. Toxicol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, K.S.; Lobo, R.; Shirwaikar, A.A.; Shirwaikar, A. Ocimum gratissimum: A review of its chemical, pharmacological and ethnomedicinal properties. Open Complement. Med. J. 2009, 1, 1–15. [Google Scholar] [CrossRef]
- Lamorde, M.; Tabuti, J.R.S.; Obua, C.; Kukunda-byobona, C.; Lanyero, H.; Byakika-kibwika, P.; Bbosa, G.S.; Lubega, A.; Ogwal-okeng, J.; Ryan, M.; et al. Medicinal plants used by traditional medicine practitioners for the treatment of HIV/AIDS and related conditions in Uganda. J. Ethnopharmacol. 2010, 130, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Kisangau, D.P.; Lyaruu, H.V.M.; Hosea, K.M.; Joseph, C.C. Use of traditional medicines in the management of HIV/AIDS opportunistic infections in Tanzania: A case in the Bukoba rural district. J. Ethnobiol. Ethnomed. 2007, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gurrapu, S.; Mamidala, E. Medicinal plants used by traditional medicine practitioners in the management of HIV/AIDS-related diseases in tribal areas of Adilabad district, Telangana region. Am. J. Sci. Med. Res. 2016, 2, 239–245. [Google Scholar]
- Ayisi, N.K.; Nyadedzor, C. Comparative in vitro effects of AZT and extracts of Ocimum gratissimum, Ficus polita, Clausena anisata, Alchornea cordifolia, and Elaeophorbia drupifera against HIV-1 and HIV-2 infections. Antivir. Res. 2003, 58, 25–33. [Google Scholar] [CrossRef]
- Tibyangye, J.; Okech, M.A.; Nyabayo, J.M.; Nakavuma, L.; Sciences, B.; Resources, A. In vitro antibacterial activity of Ocimum suave essential oils against Uropathogens isolated from patients in selected hospitals in Bushenyi district, Uganda. Br. Microbiol. Res. J. 2015, 8, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Rege, A.A.; Ambaye, R.Y.; Deshmukh, R.A. In Vitro testing of anti-HIV activity of some medicinal plants. Indian J. Nat. Prod. Resour. 2010, 1, 193–199. [Google Scholar]
- Silprasit, K.; Seetaha, S.; Pongsanaraku, P.; Hannongbua, S.; Choowongkomon, K. Anti-HIV-1 reverse transcriptase activities of hexane extracts from some asian medicinal plants. J. Med. Plant Res. 2011, 5, 4899–4906. [Google Scholar]
- Hussein, A.A.; Meyer, J.J.M.; Jimeno, L. Bioactive diterpenes from Orthosiphon labiatus and Salvia africana-lutea. J. Nat. Prod. 2007, 70, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Kapewangolo, P.; Omolo, J.J.; Bruwer, R.; Fonteh, P.; Meyer, D. Antioxidant and anti-inflammatory activity of Ocimum labiatum extract and isolated labdane diterpenoid. J. Inflamm. 2015, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Goff, D.A.; Abraham, R.J. The NMR spectra of porphyrins. Org. Magn. Reson. 1984, 22, 779–783. [Google Scholar] [CrossRef]
- Ina, A.; Hayashi, K.-I.; Nozaki, H.; Kamei, Y. Pheophytin a, a low molecular weight compound found in the marine brown alga Sargassum fulvellum, promotes the differentiation of PC12 cells. Int. J. Dev. Neurosci. 2007, 25, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kalebic, T.; Kinter, A.; Poli, G.; Anderson, M.E.; Meister, A.; Fauci, A.S. Suppression of human immunodeficiency virus expression in chronically infected monocytic cells by glutathione, glutathione ester, and N-acetylcysteine. Proc. Natl. Acad. Sci. USA 1991, 88, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, P.; Meyer, D. In vitro reactivation of latent HIV-1 by cytostatic bis(thiosemicarbazonate) gold(III) complexes. BMC Infect. Dis. 2014, 14, 680. [Google Scholar] [CrossRef] [PubMed]
- Atienzar, F.A.; Tilmant, K.; Gerets, H.H.; Toussaint, G.; Speeckaert, S.; Hanon, E.; Depelchin, O.; Dhalluin, S. The use of real-time cell analyzer technology in drug discovery: Defining optimal cell culture conditions and assay reproducibility with different adherent cellular models. J. Biomol. Screen. 2011, 16, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Geuenich, S.; Goffinet, C.; Venzke, S.; Nolkemper, S.; Baumann, I.; Plinkert, P.; Reichling, J.; Keppler, O.T. Aqueous extracts from peppermint, sage and lemon balm leaves display potent anti-HIV-1 activity by increasing the virion density. Retrovirology 2008, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kapewangolo, P.; Hussein, A.A.; Meyer, D. Inhibition of HIV-1 enzymes, antioxidant and anti-inflammatory activities of Plectranthus barbatus. J. Ethnopharmacol. 2013, 149, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Klos, M.; Van de Venter, P.J.; Milne, P.J.; Traore, H.N.; Meyer, D.; Oosthuizen, V. In vitro anti-HIV activity of five selected South African medicinal plant extracts. J. Ethnopharmacol. 2009, 124, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Farsani, M.S.; Behbahani, M.; Isfahani, H.Z. The effect of root, shoot and seed extracts of the Iranian Thymus L. (family: Lamiaceae) species on HIV-1 replication and CD4 expression. Cell J. 2016, 18, 255–262. [Google Scholar]
- Fang, H.; Chokkalingam, U.; Chiou, S.; Hwang, T. Bioactive chemical constituents from the brown alga Homoeostrichus formosana. Int. J. Mol. Sci. 2015, 16, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Lanfer-Marquez, U.M.; Barros, R.M.C.; Sinnecker, P. Antioxidant activity of chlorophylls and their derivatives. Food Res. Int. 2005, 38, 885–891. [Google Scholar] [CrossRef]
- Liu, C.M.; Kao, C.L.; Wu, H.M.; Li, W.J.; Huang, C.T.; Li, H.T.; Chen, C.Y. Antioxidant and anticancer aporphine alkaloids from the leaves of Nelumbo nucifera Gaertn. cv. Rosa-plena. Molecules 2014, 19, 17829–17838. [Google Scholar] [CrossRef] [PubMed]
- Makatini, M.M.; Petzold, K.; Arvidsson, P.I.; Honarparvar, B.; Govender, T.; Maguire, G.E.M.; Parboosing, R.; Sayed, Y.; Soliman, M.E.S.; Kruger, H.G. Synthesis, screening and computational investigation of pentacycloundecane-peptoids as potent CSA-HIV PR inhibitors. Eur. J. Med. Chem. 2012, 57, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Sakdarat, S.; Shuyprom, A.; Pientong, C.; Ekalaksananan, T.; Thongchai, S. Bioactive constituents from the leaves of Clinacanthus nutans Lindau. Bioorg. Med. Chem. 2009, 17, 1857–1860. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.L.; Lam, M.L.; Au, T.K.; Ip, D.T.M.; Ng, T.B.; Fong, W.P.; Wan, D.C.C. A comparison of human immunodeficiency virus type-1 protease inhibition activities by the aqueous and methanol extracts of Chinese medicinal herbs. Life Sci. 2000, 67, 2889–2896. [Google Scholar] [CrossRef]
- Li, Y.; Starr, S.E.; Lisziewicz, J.; Ho, W.Z. Inhibition of HIV-1 replication in chronically infected cell lines and peripheral blood mononuclear cells by retrovirus-mediated antitat gene transfer. Gene Ther. 2000, 7, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Kudo, E.; Taura, M.; Matsuda, K.; Shimamoto, M.; Kariya, R.; Goto, H.; Hattori, S.; Kimura, S.; Okada, S. Inhibition of HIV-1 replication by a tricyclic coumarin GUT-70 in acutely and chronically infected cells. Bioorg. Med. Chem. Lett. 2013, 23, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Mirabelli, C.K.; Johnson, R.K.; Sung, C.M.; Sung, C.; Faucette, L.; Muirhead, K.; Crooke, S.T. Evaluation of the in vivo antitumor activity and in vitro cytotoxic properties of auranofin, a coordinated gold compound, in murine tumor models. Cancer Res. 1985, 45, 32–39. [Google Scholar] [PubMed]
Sample Availability: Samples of the extract and compound are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Isolated 1H δ ppm | Literature 1H δ ppm [23] |
|---|---|---|
| α | 9.38 | 9.38 |
| β | 9.52 | 9.52 |
| δ | 8.57 | 8.55 |
| 1a | 3.4 | 3.4 |
| 2a | 8 | 8 |
| 2b | 6.31 | 6.28 |
| 2 | 6.17 | 6.18 |
| 3a | 3.23 | 3.23 |
| 4a | 3.69 | 3.68 |
| 4b | 1.69 | 1.69 |
| 7 | 4.21 | 4.21 |
| 7a | 2.63 | 2.63 |
| 7a′ | 2.33 | 2.34 |
| 7b | 2.47 | 2.49 |
| 7b′ | 2.19 | 2.19 |
| 8 | 4.47 | 4.46 |
| 8a | 1.81 | 1.8 |
| 10 | 6.26 | 6.26 |
| 10b | 3.89 | 3.88 |
| P-5/P-15 | 1.1, 1.3 | 1.0–1.3 |
| P-7a, P-11a | 0.86 | 0.85 |
| NH | −1.6 | −1.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapewangolo, P.; Kandawa-Schulz, M.; Meyer, D. Anti-HIV Activity of Ocimum labiatum Extract and Isolated Pheophytin-a. Molecules 2017, 22, 1763. https://doi.org/10.3390/molecules22111763
Kapewangolo P, Kandawa-Schulz M, Meyer D. Anti-HIV Activity of Ocimum labiatum Extract and Isolated Pheophytin-a. Molecules. 2017; 22(11):1763. https://doi.org/10.3390/molecules22111763
Chicago/Turabian StyleKapewangolo, Petrina, Martha Kandawa-Schulz, and Debra Meyer. 2017. "Anti-HIV Activity of Ocimum labiatum Extract and Isolated Pheophytin-a" Molecules 22, no. 11: 1763. https://doi.org/10.3390/molecules22111763
APA StyleKapewangolo, P., Kandawa-Schulz, M., & Meyer, D. (2017). Anti-HIV Activity of Ocimum labiatum Extract and Isolated Pheophytin-a. Molecules, 22(11), 1763. https://doi.org/10.3390/molecules22111763