Wound-Healing Peptides for Treatment of Chronic Diabetic Foot Ulcers and Other Infected Skin Injuries




Abstract
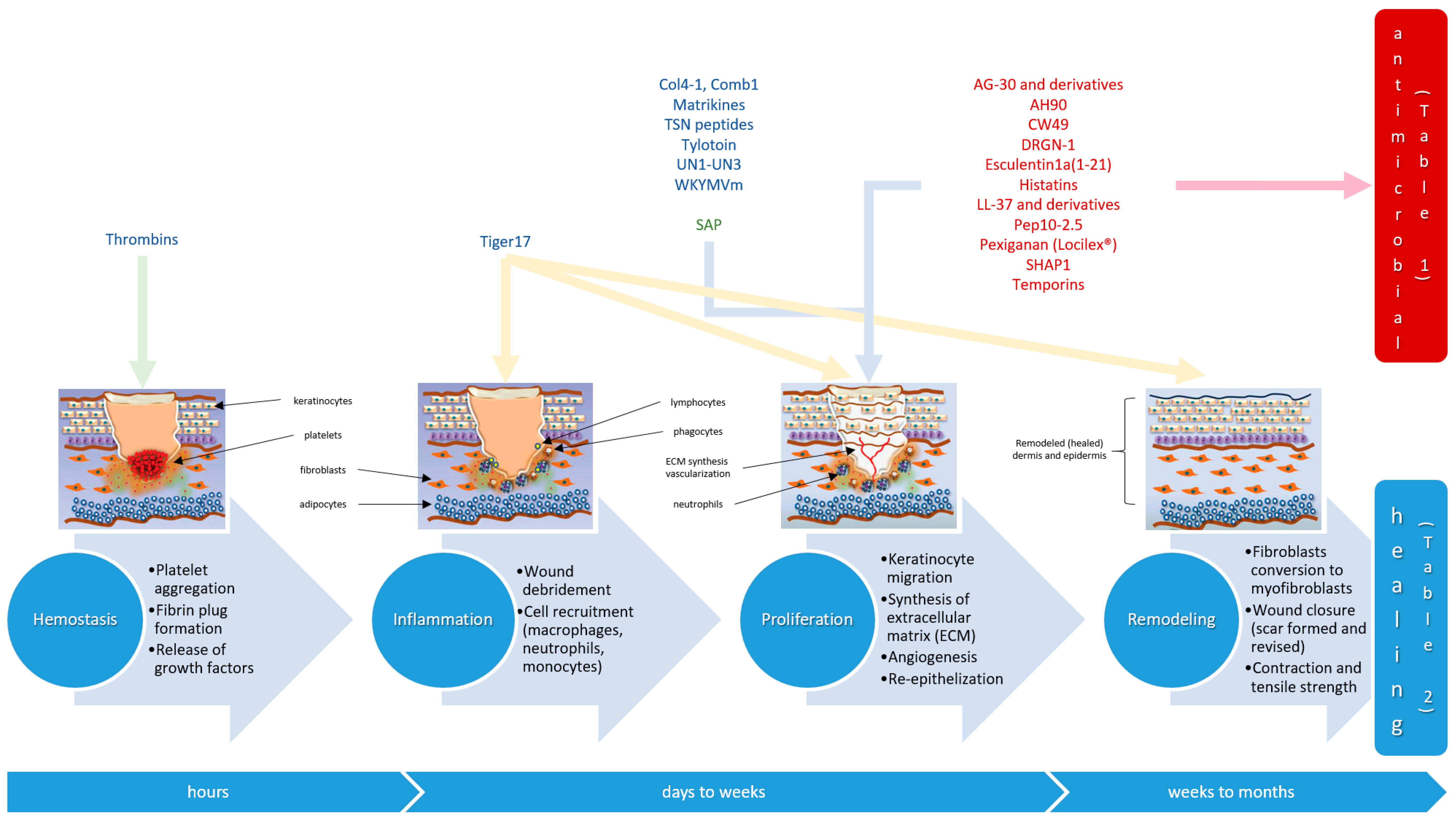
1. Introduction
2. Wound-Healing Antimicrobial Peptides
2.1. Human-Based Defence Peptides
2.2. Amphibian-Based Defence Peptides
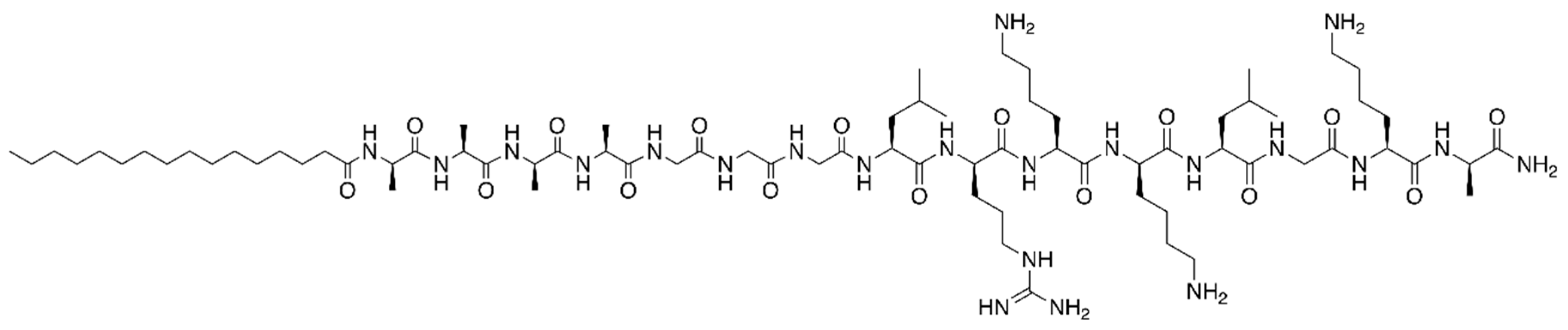
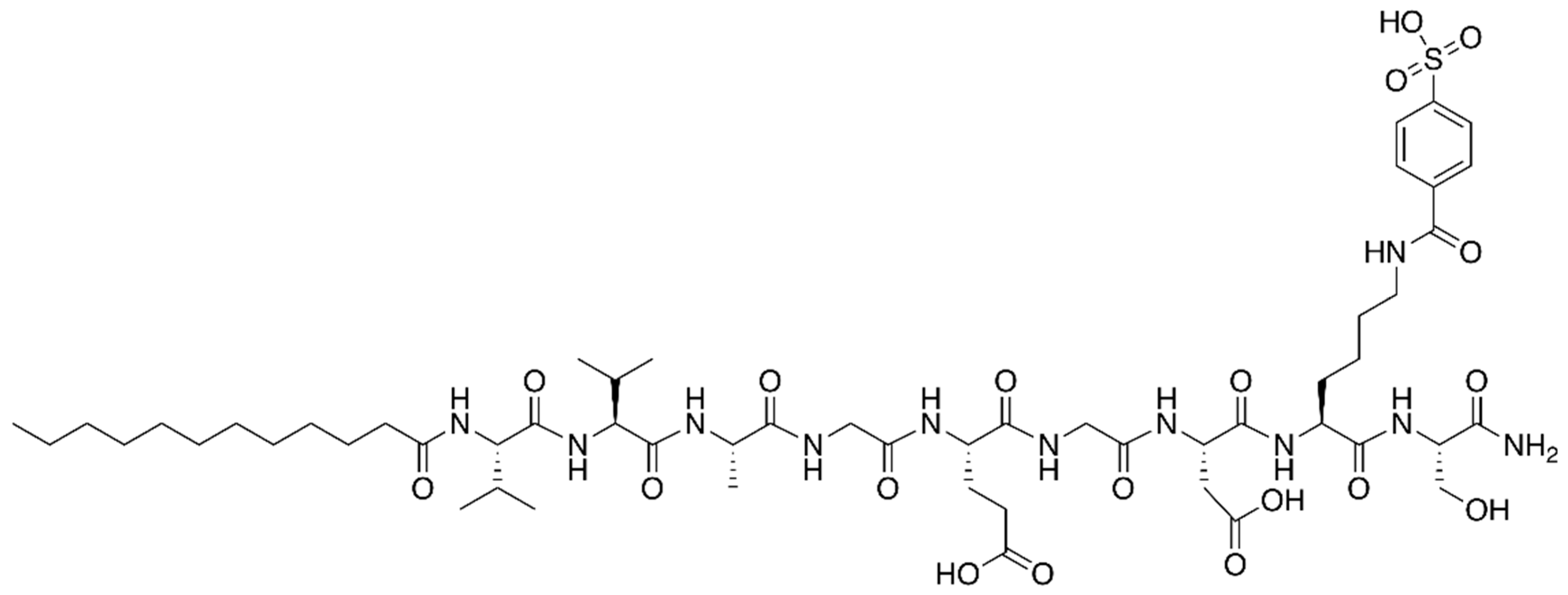
2.3. Synthetic Peptides
3. Tissue Regeneration Peptides
3.1. Human-Based Peptides
3.2. Amphibian-Based Peptides
3.3. Self-Assembling Peptides
4. Peptide-Based Wound Dressings
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- International Diabetes Federation. IDF Diabetes Atlas, 7th ed.; IDF: Brussels, Belgium, 2015. [Google Scholar]
- Uckay, I.; Gariani, K.; Pataky, Z.; Lipsky, B.A. Diabetic foot infections: State-of-the-art. Diabetes Obes. Metab. 2014, 16, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Alexiadou, K.; Doupis, J. Management of diabetic foot ulcers. Diabetes Ther. 2012, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Epicast Report: Diabetic Foot Ulcers-Epidemiology Forecast to 2025. Available online: https://www.researchandmarkets.com/research/l9r7nt/epicast_report (accessed on 28 July 2017).
- Hingorani, A.; LaMuraglia, G.M.; Henke, P.; Meissner, M.H.; Loretz, L.; Zinszer, K.M.; Driver, V.R.; Frykberg, R.; Carman, T.L.; Marston, W.; et al. The management of diabetic foot: A clinical practice guideline by the society for vascular surgery in collaboration with the american podiatric medical association and the society for vascular medicine. J. Vasc. Surg. 2016, 63, 3S–21S. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, K.V.; Tiwari, S.; Purandare, V.B.; Khedkar, S.; Bhosale, S.S.; Unnikrishnan, A.G. Choice of wound care in diabetic foot ulcer: A practical approach. World J. Diabetes 2014, 5, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.S.; Evans, G.R. Advances in wound healing: A review of current wound healing products. Plast. Surg. Int. 2012, 2012, 190436. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.; Wrobel, J.S.; Maceachern, M.P.; Boles, B.R. Collagen-based wound dressings for the treatment of diabetes-related foot ulcers: A systematic review. Diabetes Metab. Syndr. Obes. 2013, 6, 17–29. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Theato, P. Collagen and collagen mimetic peptide conjugates in polymer science. Eur. Polym. J. 2013, 49, 2986–2997. [Google Scholar] [CrossRef]
- Abu Samah, N.H.; Heard, C.M. Topically applied kttks: A review. Int. J. Cosmet. Sci. 2011, 33, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S.; Salza, R. Matricryptins and matrikines: Biologically active fragments of the extracellular matrix. Exp. Dermatol. 2014, 23, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Mai, S.; Mauger, M.T.; Niu, L.N.; Barnes, J.B.; Kao, S.; Bergeron, B.E.; Ling, J.Q.; Tay, F.R. Potential applications of antimicrobial peptides and their mimics in combating caries and pulpal infections. Acta Biomater. 2017, 49, 16. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.L. Antimicrobial peptides stage a comeback. Nat. Biotechnol. 2013, 31, 379. [Google Scholar] [CrossRef] [PubMed]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Kim, C.; Seo, C.H.; Park, Y. The therapeutic applications of antimicrobial peptides (AMPs): A patent review. J. Microbiol. 2017, 55, 1. [Google Scholar] [CrossRef] [PubMed]
- Fuente-Nuñez, C.; Silva, O.N.; Lu, T.K.; Franco, O.L. Antimicrobial peptides: Role in human disease and potential as immunotherapies. Pharmacol. Ther. 2017, 178, 132. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host defense antimicrobial peptides as antibiotics: Design and application strategies. Curr. Opin. Chem. Biol. 2017, 38, 87. [Google Scholar] [CrossRef] [PubMed]
- Travkova, O.; Moehwald, H.; Berezinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521. [Google Scholar] [CrossRef] [PubMed]
- Suleman, L. Extracellular bacterial proteases in chronic wounds: A potential therapeutic target? Adv. Wound Care New Rochelle 2016, 5, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Barton, G.M. A calculated response: Control of inflammation by the innate immune system. J. Clin. Investig. 2008, 118, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Usui, M.L.; Lippman, S.I.; James, G.A.; Stewart, P.S.; Fleckman, P.; Olerud, J.E. Biofilms and inflammation in chronic wounds. Adv. Wound Care 2013, 2, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Nishikawa, T.; Tamura, N.; Maeda, A.; Hibino, H.; Mochizuki, M.; Shimosato, T.; Moriya, T.; Morishita, R.; Tamai, K.; et al. Modification of a novel angiogenic peptide, ag30, for the development of novel therapeutic agents. J. Cell. Mol. Med. 2012, 16, 1629–1639. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Mu, L.; Tang, J.; Shen, C.; Gao, C.; Rong, M.; Zhang, Z.; Liu, J.; Wu, X.; Yu, H.; et al. A potential wound healing-promoting peptide from frog skin. Int. J. Biochem. Cell Biol. 2014, 49, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Duan, Z.; Tang, J.; Lv, Q.; Rong, M.; Lai, R. A short peptide from frog skin accelerates diabetic wound healing. FEBS J. 2014, 281, 4633–4643. [Google Scholar] [CrossRef] [PubMed]
- Song, D.W.; Kim, S.H.; Kim, H.H.; Lee, K.H.; Ki, C.S.; Park, Y.H. Multi-biofunction of antimicrobial peptide-immobilized silk fibroin nanofiber membrane: Implications for wound healing. Acta Biomater. 2016, 39, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Cappiello, F.; Imanishi, A.; Mastrofrancesco, A.; Picardo, M.; Paus, R.; Mangoni, M.L. The frog skin-derived antimicrobial peptide esculentin-1a(1-21) NH2 promotes the migration of human hacat keratinocytes in an egf receptor-dependent manner: A novel promoter of human skin wound healing? PLoS ONE 2015, 10, e0128663. [Google Scholar] [CrossRef] [PubMed]
- Dhople, V.; Krukemeyer, A.; Ramamoorthy, A. The human beta-defensin-3, an antibacterial peptide with multiple biological functions. Biochim. Biophys. Acta 2006, 1758, 1499–1512. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; Trujillo, V.; Montoya, A.; Gonzalez-Curiel, I.; Castaneda-Delgado, J.; Cardenas, A.; Rincon, K.; Hernandez, M.L.; Hernandez-Pando, R. Expression of antimicrobial peptides in diabetic foot ulcer. J. Dermatol. Sci. 2012, 65, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Oudhoff, M.J.; Bolscher, J.G.; Nazmi, K.; Kalay, H.; van’t Hof, W.; Amerongen, A.V.; Veerman, E.C. Histatins are the major wound-closure stimulating factors in human saliva as identified in a cell culture assay. FASEB J. 2008, 22, 3805–3812. [Google Scholar] [CrossRef] [PubMed]
- Steinstraesser, L.; Hirsch, T.; Schulte, M.; Kueckelhaus, M.; Jacobsen, F.; Mersch, E.A.; Stricker, I.; Afacan, N.; Jenssen, H.; Hancock, R.E.; et al. Innate defense regulator peptide 1018 in wound healing and wound infection. PLoS ONE 2012, 7, e39373. [Google Scholar] [CrossRef] [PubMed]
- Duplantier, A.J.; van Hoek, M.L. The human cathelicidin antimicrobial peptide ll-37 as a potential treatment for polymicrobial infected wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [PubMed]
- Lamb, H.M.; Wiseman, L.R. Pexiganan acetate. Drugs 1998, 56, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Pfalzgraff, A.; Heinbockel, L.; Su, Q.; Gutsmann, T.; Brandenburg, K.; Weindl, G. Synthetic antimicrobial and lps-neutralising peptides suppress inflammatory and immune responses in skin cells and promote keratinocyte migration. Sci. Rep. 2016, 6, 31577. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.; Silva, J.P.; Rodrigues, A.C.; Costa, R.; Guardao, L.; Schmitt, F.; Soares, R.; Vilanova, M.; Domingues, L.; Gama, M. Wound healing activity of the human antimicrobial peptide ll37. Peptides 2011, 32, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Lee, Y.W.; Park, M.K.; Shin, J.R.; Lim, K.J.; Cho, J.H.; Kim, S.C. Efficacy of the designer antimicrobial peptide shap1 in wound healing and wound infection. Amino Acids 2014, 46, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, H.; Nakagami, H.; Tenma, A.; Saito, Y.; Kaga, T.; Kanamori, T.; Tamura, N.; Tomono, K.; Kaneda, Y.; Morishita, R. Novel anti-microbial peptide SR-0379 accelerates wound healing via the PI3 kinase/Akt/mTOR pathway. PLoS ONE 2014, 9, e92597. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Luca, V.; Segev-Zarko, L.A.; Shai, Y.; Mangoni, M.L. Temporins a and b stimulate migration of hacat keratinocytes and kill intracellular Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 2520–2527. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Gallo, R.L. Antimicrobial peptides in human skin disease. Eur. J. Dermatol. 2008, 18, 11–21. [Google Scholar] [PubMed]
- Carretero, M.; Escámez, M.J.; García, M.; Duarte, B.; Holguín, A.; Retamosa, L.; Jorcano, J.L.; Río, M.D.; Larcher, F. In vitro and in vivo wound healing-promoting activities of human cathelicidin ll-37. J. Investig. Dermatol. 2008, 128, 223–236. [Google Scholar] [CrossRef] [PubMed]
- McCrudden, M.T.C.; McLean, D.T.F.; Zhou, M.; Shaw, J.; Linden, G.J.; Irwin, C.R.; Lundy, F.T. The host defence peptide ll-37 is susceptible to proteolytic degradation by wound fluid isolated from foot ulcers of diabetic patients. Int. J. Pept. Res. Ther. 2014, 20, 457–464. [Google Scholar] [CrossRef]
- Gonzalez-Curiel, I.; Trujillo, V.; Montoya-Rosales, A.; Rincon, K.; Rivas-Calderon, B.; deHaro-Acosta, J.; Marin-Luevano, P.; Lozano-Lopez, D.; Enciso-Moreno, J.A.; Rivas-Santiago, B. 1,25-dihydroxyvitamin D3 induces LL-37 and HBD-2 production in keratinocytes from diabetic foot ulcers promoting wound healing: An in vitro model. PLoS ONE 2014, 9, e111355. [Google Scholar] [CrossRef] [PubMed]
- Gronberg, A.; Mahlapuu, M.; Stahle, M.; Whately-Smith, C.; Rollman, O. Treatment with LL-37 is safe and effective in enhancing healing of hard-to-heal venous leg ulcers: A randomized, placebo-controlled clinical trial. Wound Repair Regen. 2014, 22, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Nakagami, H.; Maeda, A.; Morishita, R.; Miyazaki, N.; Ogawa, T.; Tabata, Y.; Kikuchi, Y.; Hayashi, H.; Tatsu, Y.; et al. Development of a novel antimicrobial peptide, ag-30, with angiogenic properties. J. Cell. Mol. Med. 2009, 13, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Oudhoff, M.J.; Kroeze, K.L.; Nazmi, K.; van den Keijbus, P.A.; van’t Hof, W.; Fernandez-Borja, M.; Hordijk, P.L.; Gibbs, S.; Bolscher, J.G.; Veerman, E.C. Structure-activity analysis of histatin, a potent wound healing peptide from human saliva: Cyclization of histatin potentiates molar activity 1000-fold. FASEB J. 2009, 23, 3928–3935. [Google Scholar] [CrossRef] [PubMed]
- Boink, M.A.; Roffel, S.; Nazmi, K.; van Montfrans, C.; Bolscher, J.G.; Gefen, A.; Veerman, E.C.; Gibbs, S. The influence of chronic wound extracts on inflammatory cytokine and histatin stability. PLoS ONE 2016, 11, e0152613. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Magainins, a class of antimicrobial peptides from xenopus skin: Isolation, characterization of two active forms, and partial cdna sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Holroyd, K.J.; Zasloff, M. Topical versus systemic antimicrobial therapy for treating mildly infected diabetic foot ulcers: A randomized, controlled, double-blinded, multicenter trial of pexiganan cream. Clin. Infect. Dis 2008, 47, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Gottler, L.M.; Ramamoorthy, A. Structure, membrane orientation, mechanism, and function of pexiganan--a highly potent antimicrobial peptide designed from magainin. Biochim. Biophys. Acta 2009, 1788, 1680–1686. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Fernandes, M.; Pinheiro, M.; Maia, S.; Seabra, C.L.; Ferreira-da-Silva, F.; Costa, F.; Reis, S.; Gomes, P.; Martins, M.C. Antimicrobial properties of membrane-active dodecapeptides derived from msi-78. Biochim. Biophys. Acta 2015, 1848, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Pinheiro, M.; Fernandes, M.; Maia, S.; Seabra, C.L.; Ferreira-da-Silva, F.; Reis, S.; Gomes, P.; Martins, M.C. A 17-mer membrane-active msi-78 derivative with improved selectivity toward bacterial cells. Mol. Pharm. 2015, 12, 2904–2911. [Google Scholar] [CrossRef] [PubMed]
- Newswire, P. Dipexium Pharmaceuticals Announces Issuance of Locilex® Patent by European Union. Available online: http://www.prnewswire.com/news-releases/dipexium-pharmaceuticals-announces-issuance-of-locilex-patent-by-european-union-300323725.html (accessed on 28 July 2017).
- Newswire, P. Dipexium Pharmaceuticals Announces Issuance of Locilex® Patent in Japan. Available online: http://www.prnewswire.com/news-releases/dipexium-pharmaceuticals-announces-issuance-of-locilex-patent-in-japan-300241111.html (accessed on 28 July 2017).
- Newswire, P. Dipexium Announces Top-Line Data from Onestep Phase 3 Trials with Locilex® in Mild Diabetic Foot Infection Did not Meet Primary Clinical Endpoint of Superiority Versus Vehicle Plus Standardized Wound Care. Available online: http://www.prnewswire.com/news-releases/dipexium-announces-top-line-data-from-onestep-phase-3-trials-with-locilex-in-mild-diabetic-foot-infection-did-not-meet-primary-clinical-endpoint-of-superiority-versus-vehicle-plus-standardized-wound-care-300350302.html (accessed on 28 July 2017).
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef]
- Mu, L.; Tang, J.; Liu, H.; Shen, C.; Rong, M.; Zhang, Z.; Lai, R. A potential wound-healing-promoting peptide from salamander skin. FASEB J. 2014, 28, 3919–3929. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.M.C.; Dean, S.N.; Propst, C.N.; Bishop, B.M.; van Hoek, M.L. Komodo dragon-inspired peptide DRGN-1 promotes wound-healing of a mixed-biofilm infected wound. NPJ Biofilms Microb. 2017, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.W.; Heo, S.C.; Lee, T.W.; Park, G.T.; Yoon, J.W.; Jang, I.H.; Kim, S.C.; Ko, H.C.; Ryu, Y.; Kang, H.; et al. N-acetylated proline-glycine-proline accelerates cutaneous wound healing and neovascularization by human endothelial progenitor cells. Sci. Rep. 2017, 7, 43057. [Google Scholar] [CrossRef] [PubMed]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. GHK peptide as a natural modulator of multiple cellular pathways in skin regeneration. Biomed. Res. Int. 2015, 2015, 648108. [Google Scholar] [CrossRef] [PubMed]
- Demidova-Rice, T.N.; Geevarghese, A.; Herman, I.M. Bioactive peptides derived from vascular endothelial cell extracellular matrices promote microvascular morphogenesis and wound healing in vitro. Wound Repair Regen. 2011, 19, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Suguna, L.; Shanthi, C. Wound healing activity of a collagen-derived cryptic peptide. Amino Acids 2015, 47, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Liu, H.; Gao, C.; Mu, L.; Yang, S.; Rong, M.; Zhang, Z.; Liu, J.; Ding, Q.; Lai, R. A small peptide with potential ability to promote wound healing. PLoS ONE 2014, 9, e92082. [Google Scholar] [CrossRef] [PubMed]
- Fife, C.; Mader, J.T.; Stone, J.; Brill, L.; Satterfield, K.; Norfleet, A.; Zwernemann, A.; Ryaby, J.T.; Carney, D.H. Thrombin peptide chrysalin® stimulates healing of diabetic foot ulcers in a placebo-controlled phase i/ii study. Wound Repair Regen. 2007, 15, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sheets, A.R.; Demidova-Rice, T.N.; Shi, L.; Ronfard, V.; Grover, K.V.; Herman, I.M. Identification and characterization of novel matrix-derived bioactive peptides: A role for collagenase from santyl(r) ointment in post-debridement wound healing? PLoS ONE 2016, 11, e0159598. [Google Scholar] [CrossRef] [PubMed]
- Demidova-Rice, T.N.; Wolf, L.; Deckenback, J.; Hamblin, M.R.; Herman, I.M. Human platelet-rich plasma- and extracellular matrix-derived peptides promote impaired cutaneous wound healing in vivo. PLoS ONE 2012, 7, e32146. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.W.; Heo, S.C.; Jang, I.H.; Jeong, G.O.; Yoon, J.W.; Mun, J.H.; Kim, J.H. Stimulation of cutaneous wound healing by an fpr2-specific peptide agonist wkymvm. Wound Repair Regen. 2015, 23, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Sheets, A.R.; Massey, C.J.; Cronk, S.M.; Iafrati, M.D.; Herman, I.M. Matrix- and plasma-derived peptides promote tissue-specific injury responses and wound healing in diabetic swine. J. Transl. Med. 2016, 14, 197. [Google Scholar] [CrossRef] [PubMed]
- Maquart, F.X.; Pasco, S.; Ramont, L.; Hornebeck, W.; Monboisse, J.C. An introduction to matrikines: Extracellular matrix-derived peptides which regulate cell activity. Implication in tumor invasion. Crit. Rev. Oncol. Hematol. 2004, 49, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Arul, V.; Gopinath, D.; Gomathi, K.; Jayakumar, R. Biotinylated ghk peptide incorporated collagenous matrix: A novel biomaterial for dermal wound healing in rats. J. Biomed. Mater. Res. B Appl. Biomater. 2005, 73, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Arul, V.; Kartha, R.; Jayakumar, R. A therapeutic approach for diabetic wound healing using biotinylated ghk incorporated collagen matrices. Life Sci. 2007, 80, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.; Goktas, M.; Tekinay, A.B.; Guler, M.O.; Hauser, C.A.; Mitraki, A. Self-assembled proteins and peptides as scaffolds for tissue regeneration. Adv. Healthc. Mater. 2015, 4, 2557–2586. [Google Scholar] [CrossRef] [PubMed]
- Rajangam, K.; Behanna, H.A.; Hui, M.J.; Han, X.; Hulvat, J.F.; Lomasney, J.W.; Stupp, S.I. Heparin binding nanostructures to promote growth of blood vessels. Nano Lett. 2006, 6, 2086–2090. [Google Scholar] [CrossRef] [PubMed]
- Mammadov, R.; Mammadov, B.; Toksoz, S.; Aydin, B.; Yagci, R.; Tekinay, A.B.; Guler, M.O. Heparin mimetic peptide nanofibers promote angiogenesis. Biomacromolecules 2011, 12, 3508–3519. [Google Scholar] [CrossRef] [PubMed]
- Senturk, B.; Mercan, S.; Delibasi, T.; Guler, M.O.; Tekinay, A.B. Angiogenic peptide nanofibers improve wound healing in stz-induced diabetic rats. ACS Biomater. Sci. Eng. 2016, 2, 1180–1189. [Google Scholar] [CrossRef]
- Han, G.; Ceilley, R. Chronic wound healing: A review of current management and treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Raines, R.T. Review collagen-based biomaterials for wound healing. Biopolymers 2014, 101, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, Y.; Han, H.; De Loera, R.; Wen, Y.; Zhang, X.; Sun, T.; Mora-Solano, C.; Collier, J.H. Peptide biomaterials raising adaptive immune responses in wound healing contexts. J. Biomed. Mater. Res. Part A 2016, 104, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Cereceres, S.; Touchet, T.; Browning, M.B.; Smith, C.; Rivera, J.; Höök, M.; Whitfield-Cargile, C.; Russell, B.; Cosgriff-Hernandez, E. Chronic wound dressings based on collagen-mimetic proteins. Adv. Wound Care 2015, 4, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Collagenase Diabetic Foot Ulcer Study Group; Tallis, A.; Motley, T.A.; Wunderlich, R.P.; Dickerson, J.E., Jr.; Waycaster, C.; Slade, H.B. Clinical and economic assessment of diabetic foot ulcer debridement with collagenase: Results of a randomized controlled study. Clin. Ther. 2013, 35, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Choudary, D.; Insen, S.G.; Goyal, S.; Chabbra, U.; Singal, G. A comparative study of collagen dressings versus conventional dressings in wound healing in chronic ulcers. J. Evol. Med. Dent. Sci. 2017, 6, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Ikram, S. Chitosan based scaffolds and their applications in wound healing. Achiev. Life Sci. 2016, 10, 27–37. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, M.; Shao, X.; Wang, X.; Zhang, L.; Xu, P.; Zhong, W.; Zhang, L.; Xing, M.; Zhang, L. A laminin mimetic peptide sikvav-conjugated chitosan hydrogel promoting wound healing by enhancing angiogenesis, re-epithelialization and collagen deposition. J. Mater. Chem. B 2015, 3, 6798–6804. [Google Scholar] [CrossRef]
- Silva, J.P.; Dhall, S.; Garcia, M.; Chan, A.; Costa, C.; Gama, M.; Martins-Green, M. Improved burn wound healing by the antimicrobial peptide llkkk18 released from conjugates with dextrin embedded in a carbopol gel. Acta Biomater. 2015, 26, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Reis, L.A.; Feric, N.; Knee, E.J.; Gu, J.; Cao, S.; Laschinger, C.; Londono, C.; Antolovich, J.; McGuigan, A.P.; et al. Diabetic wound regeneration using peptide-modified hydrogels to target re-epithelialization. Proc. Natl. Acad. Sci. USA 2016, 113, E5792–E5801. [Google Scholar] [CrossRef] [PubMed]
- Grek, C.L.; Prasad, G.M.; Viswanathan, V.; Armstrong, D.G.; Gourdie, R.G.; Ghatnekar, G.S. Topical administration of a connexin43-based peptide augments healing of chronic neuropathic diabetic foot ulcers: A multicenter, randomized trial. Wound Repair Regen. 2015, 23, 203–212. [Google Scholar] [CrossRef] [PubMed]
- National Library of Medicine. A Study of Granexin Gel in the Treatment of Diabetic Foot Ulcer (Identifier: Nct02667327). Available online: https://clinicaltrials.gov/ct2/show/NCT02667327 (accessed on 14 June 2017).
- Seo, E.; Lim, J.S.; Jun, J.B.; Choi, W.; Hong, I.S.; Jun, H.S. Exendin-4 in combination with adipose-derived stem cells promotes angiogenesis and improves diabetic wound healing. J. Transl. Med. 2017, 15, 35. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
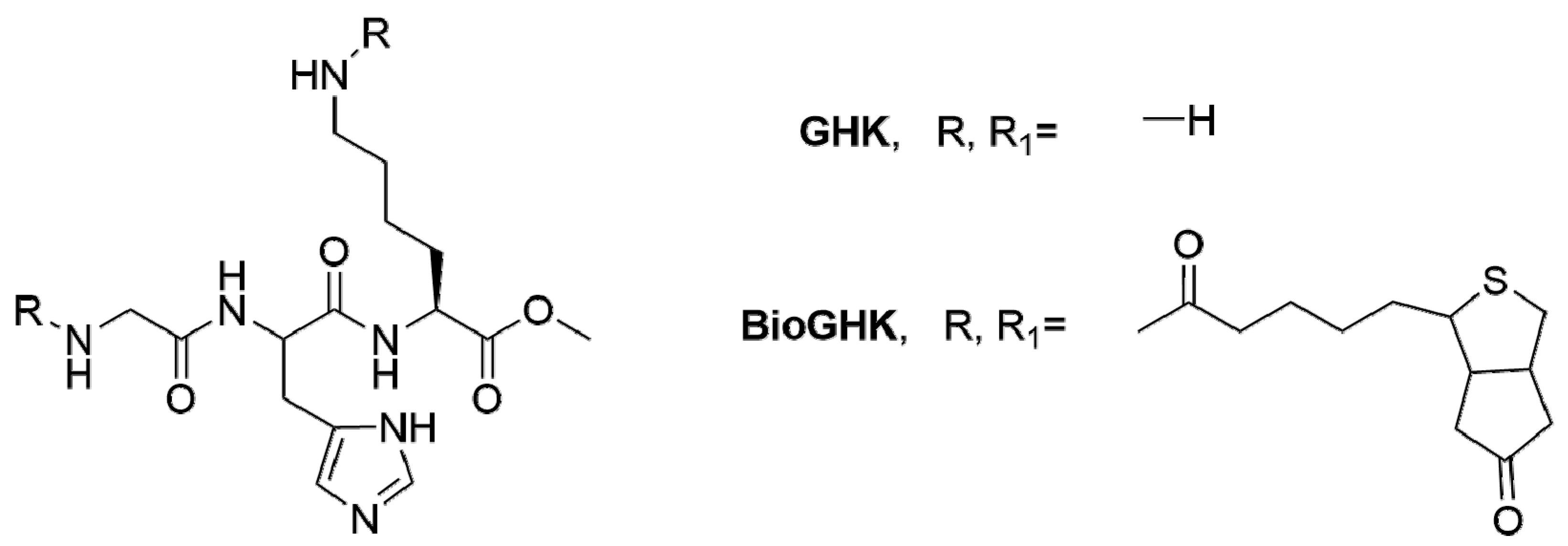{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Peptide Sequence | Reference |
|---|---|---|
| AG30 | MLSLIFLHRLKSMRKRLDRKLRLWHRKNYP | [26] |
| AG30/5C | MLKLIFLHRLKRMRKRLKRKLRLWHRKRYK | |
| AH90 | ATAWDFGPHGLLPIRPIRIRPLCG | [27] |
| CW49 | APFRMGICTTN | [28] |
| Cys-KR12 | KRIVKRIKKWLR | [29] |
| Esculentin-1a(1-21) | GIFSKLAGKKIKNLLISGLKG | [30] |
| hBD-1 | DHYNCVSSGGQCLYSACPIFTKIQGTCYRGKAKCCK (Disulfide bridges: 5-34, 12-27, 17-35) | [31,32] |
| hBD-2 | GIGDPVTCLKSGAICHPVFCPRRYKQIGTCGLPGTKCCKKP (Disulfide bridges: 8-37, 15-30, 20-38) | |
| hBD-3 | GIINTLQKYYCRVRGGRCAVLSCLPKEEQIGKCSTRGRKCCRRKK (Disulfide bridges: 11-40, 18-33, 23-41) | |
| hBD-4 | ELDRICGYGTARCRKKCRSQEYRIGRCPNTYACCLRK (Disulfide bridges: 6-33; 13-27; 17-34) | |
| Histatin-1 | DSHEKRHHGYRRKFHEKHHSHREFPFYGDYGSNYLYDN | [33] |
| Histatin-2 | RKFHEKHHSHREFPFYGDYGSNYLYDN | |
| Histain-3 | DSHAKRHHGYKRKFHEKHHSHRGYRSNYLYDN | |
| IDR-1018 | VRLIVAVRIWRR | [34] |
| LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | [35] |
| MSI-78 (pexiganan) | GIGKFLKKAKKFGKAFVKILKK | [36] |
| Pep19-2.5 | GCKKYRRFRWKFKGKFWFWG | [37] |
| PLL-37 | PLLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | [38] |
| SHAP1 | APKAMKLLKKLLKLQKKGI | [39] |
| SR-0007 | MLKLIFLHRLKRMRKRLKRK | [40] |
| SR-0379 | MLKLIFLHRLKRMRKRLkRK 1 | |
| Temporin A | FLPLIGRVLSGIL | [41] |
| Temporin B | LLPIVGNLLKSLL |
| Peptide Name | Peptide Sequence | Reference |
|---|---|---|
| Ac-PGP | N-acetylated-PGP | [61] |
| BioGHK | Biotinylated-GHK | [62] |
| Col4-1 | MFRKPIPSTVKA | [63] |
| Comb1 | DINECEIGAPAGEETEVTVEGLEPG | |
| E1 | GETGPAGPAGPIGPVGARGPAGPQGPRGDKGETGEQ | [64] |
| Tiger17 | WCKPKPKPRCH | [65] |
| TP-508 | AGYKPDEGKRGDACEGDSGGPFV | [66] |
| TSN1 | NFQGVQNRFVFGTP | [67] |
| TSN2 | MENAELDVPIQSVFTR | |
| TSN3 | NTDNIYPESSC | |
| TSN4 | PYLGYVFK | |
| TSN5 | MQTVAQLFKTVSSLSLST | |
| TSN6 | HSPDIQLQKGLTFEPIQIK | |
| TSN7 | STITQPYKTLNNARSP | |
| TSN8 | RPGPSPEGTGQSYNY | |
| TSN9 | MENAELDPPYLGYVFK | |
| TSN10 | TGQSYNQYSQRPYLGVYVFK | |
| TSN11 | LYGQTPLETL | |
| TSN12 | ELADSPALEIG | |
| TSN13 | LYGQTPLETLELADSPALEIG | |
| TSN14 | VSGNTVEYALPTLE | |
| TSN15 | LDSPTAPTVQSTALTWRP | |
| TSN16 | LDGSAPGPLYTGSALDF | |
| TSN17 | GSEGVRSGRSG | |
| TSN18 | QPQPLPSPGVGGKN | |
| Tylotoin | KCVRQNNKRVCK | [59] |
| UN1 | ELLESYIDGR | [68] |
| UN2 | TATSEYQTFFNPR | |
| UN3 | ELLESYIDGRPTATSEYQTFFNPR | |
| WKYMVm | WKYMVm 1 | [69] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, A.; Teixeira, C.; Ferraz, R.; Prudêncio, C.; Gomes, P. Wound-Healing Peptides for Treatment of Chronic Diabetic Foot Ulcers and Other Infected Skin Injuries. Molecules 2017, 22, 1743. https://doi.org/10.3390/molecules22101743
Gomes A, Teixeira C, Ferraz R, Prudêncio C, Gomes P. Wound-Healing Peptides for Treatment of Chronic Diabetic Foot Ulcers and Other Infected Skin Injuries. Molecules. 2017; 22(10):1743. https://doi.org/10.3390/molecules22101743
Chicago/Turabian StyleGomes, Ana, Cátia Teixeira, Ricardo Ferraz, Cristina Prudêncio, and Paula Gomes. 2017. "Wound-Healing Peptides for Treatment of Chronic Diabetic Foot Ulcers and Other Infected Skin Injuries" Molecules 22, no. 10: 1743. https://doi.org/10.3390/molecules22101743
APA StyleGomes, A., Teixeira, C., Ferraz, R., Prudêncio, C., & Gomes, P. (2017). Wound-Healing Peptides for Treatment of Chronic Diabetic Foot Ulcers and Other Infected Skin Injuries. Molecules, 22(10), 1743. https://doi.org/10.3390/molecules22101743