Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems

 ,
,  ,
,  ,
,  ,
,  , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Plant Secondary Metabolites
2.1. Ecological Role of Biogenic Volatile Organic Compounds
2.2. Plant Volatiles Functions
2.2.1. Allelopathy
2.2.2. Adaptation to Abiotic Stresses
2.2.3. Intra-Plant Signaling
2.2.4. Inter-Plant Signaling
2.2.5. Direct Defense against Herbivores and Pathogens
2.2.6. Indirect Defense
2.3. Glandular Trichomes as Secreting Organs
2.3.1. Biosynthesis of Plant Volatiles in Glandular Trichomes
2.3.2. Ecological Roles of Glandular Trichomes
3. Mechanisms of Essential Oil Cytotoxicity
3.1. Molecular And Biochemical Targets in Prokaryotic Cells
3.2. Molecular and Biochemical Targets in Eukaryotic Cells
3.2.1. Anti-Cancer Activity
3.2.2. Antifungal Activity
3.2.3. Antiparasitic Activity
4. Essential Oil in Traditional Medicines: Case Studies
4.1. Traditional Iranian Medicine
4.1.1. Preclinical Studies
4.1.2. Clinical Studies
4.2. Ayurvedic Medicine
4.2.1. Preclinical Studies
Diabetes and Related Complications
Cancer and Radiotherapy
Neurodegenerative Diseases
4.2.2. Clinical Studies
4.3. Traditional Korean Medicine
Preclinical Studies
4.4. European and American Pharmacopoeias
4.4.1. Preclinical Studies
Eucalyptus Species Essential Oils
Mentha Species Essential Oils
Citrus Species Essential Oils
4.4.2. Clinical Studies
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Iriti, M.; Faoro, F. Chemical diversity and defence metabolism: How plants cope with pathogens and ozone pollution. Int. J. Mol. Sci. 2009, 10, 3371–3399. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Colnaghi, G.; Chemat, F.; Smadja, J.; Faoro, F.; Visinoni, F.A. Histo-cytochemistry and scanning electron microscopy of Lavender glandular trichomes following conventional and microwave-assisted hydrodistillation of essential oils: A comparative study. Flavour Fragr. J. 2006, 21, 704–712. [Google Scholar] [CrossRef]
- Fraenkel, G.S. The raison d’etre of secondary plant substances. Science 1959, 129, 1466–1470. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evol. Psychol. 1964, 586–608. [Google Scholar] [CrossRef]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Guillet, J.T.A.G.; Durst, T. Phytochemical diversity of insect defenses in tropical and temperate plant families. In Advances in Insect Chemical Ecology; Cambridge University Press: Cambridge, UK, 2004; pp. 56–77. [Google Scholar]
- Berenbaum, M.R.; Zangerl, A.R. Phytochemical diversity. In Phytochemical Diversity and Redundancy in Ecological Interactions; Springer: Berlin, Germany, 1996; pp. 1–24. [Google Scholar]
- Firn, R.D.; Jones, C.G. An explanation of secondary product “redundancy”. In Phytochemical Diversity and Redundancy in Ecological Interactions; Springer: Berlin, Germany, 1996; pp. 295–312. [Google Scholar]
- Harborne, J.B. Twenty-five years of chemical ecology. Nat. Prod. Rep. 2001, 18, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Firn, R.D.; Jones, C.G. Natural products—A simple model to explain chemical diversity. Nat. Prod. Rep. 2003, 20, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, E.; Gijzen, M. Phytochemical diversity: The sounds of silent metabolism. Plant Sci. 2009, 176, 161–169. [Google Scholar] [CrossRef]
- Firn, R.D.; Jones, C.G. The evolution of secondary metabolism—A unifying model. Mol. Microbiol. 2000, 37, 989–994. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Vocabulary of Natural Materials: ISO/D1S9235.2. 2013. Available online: http://www.iso.org/iso/iso_catalogue/catalogue_tc/catalogue_detail.htm?csnumber= 51017 (accessed on 6 June 2016).
- War, A.R.; Sharma, H.C.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Herbivore induced plant volatiles: Their role in plant defense for pest management. Plant Signal. Behav. 2011, 6, 1973–1978. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Blande, J.D. Where do herbivore-induced plant volatiles go? Front. Plant Sci. 2013, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Glas, J.J.; Schimmel, B.C.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Yoshimura, H. Monoterpenes of Salvia leucophylla. Curr. Bioact. Compd. 2012, 8, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Hare, J.D. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Annu. Rev. Entomol. 2011, 56, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Markus Lange, B.; Turner, G.W. Terpenoid biosynthesis in trichomes—Current status and future opportunities. Plant Biotechnol. J. 2013, 11, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yuan, Y.; Liu, Z.; Zhu, J. Development and structure of internal glands and external glandular trichomes in pogostemon cablin. PLoS ONE 2013, 8, e77862. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-S.; Kirchoff, B.K.; Liao, J.-P. The capitate and peltate glandular trichomes of Lavandula pinnata L. (lamiaceae): Histochemistry, ultrastructure, and secretion. J. Torrey Bot. Soc. 2008, 135, 155–167. [Google Scholar] [CrossRef]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Kapteyn, J.; Gang, D.R. A systems biology investigation of the MEP/terpenoid and shikimate/phenylpropanoid pathways points to multiple levels of metabolic control in sweet basil glandular trichomes. Plant J. 2008, 54, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M. The evolution of plant secretory structures and emergence of terpenoid chemical diversity. Annu. Rev. Plant Biol. 2015, 66, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.; Wang, E.; Shepherd, R. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.; Peiffer, M.; Luthe, D.S.; Felton, G.W. Trichomes as sensors: Detecting activity on the leaf surface. Plant Signal. Behav. 2010, 5, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Zitzelsberger, C.; Buchbauer, G. Essential oils as “a cry for help”. A review. Nat. Prod. Commun. 2015, 10, 1127–1138. [Google Scholar] [PubMed]
- Freires, I.A.; Denny, C.; Benso, B.; de Alencar, S.M.; Rosalen, P.L. Antibacterial activity of essential oils and their isolated constituents against cariogenic bacteria: A systematic review. Molecules 2015, 20, 7329–7358. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Exploitation of cytotoxicity of some essential oils for translation in cancer therapy. Evid. Based Complement. Alternat. Med. 2015, 2015, 397821. [Google Scholar] [CrossRef] [PubMed]
- Bayala, B.; Bassole, I.H.; Scifo, R.; Gnoula, C.; Morel, L.; Lobaccaro, J.M.; Simpore, J. Anticancer activity of essential oils and their chemical components—A review. Am. J. Cancer Res. 2014, 19, 591–607. [Google Scholar]
- Ben Arfa, A.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier, P. Antimicrobial activity of carvacrol related to its chemical structure. Lett. Appl. Microbiol. 2006, 43, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.; Tirogo, S.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Composition and antimicrobial activities of Lippia Multiflora moldenke, Mentha piperita L. and Ocimum basilicum L. Essential oils and their major monoterpene alcohols alone and in combination. Molecules 2010, 15, 7825–7839. [Google Scholar] [CrossRef] [PubMed]
- Dorman, H.; Deans, S. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, C.A.; Pendleton, S.J.; Crandall, P.G.; Ricke, S.C. Potential of plant essential oils and their components in animal agriculture—In vitro studies on antibacterial mode of action. Front. Vet. Sci. 2015, 2, 35. [Google Scholar] [CrossRef] [PubMed]
- Juliani, H.; Koroch, A.; Simon, J. Chemical diversity of essential oils of Ocimum species and their associated antioxidant and antimicrobial activity. In Essential Oils and Aromas: Green Extractions and Applications; Har Krishan Bhalla & Sons: Uttrakhand, India, 2009. [Google Scholar]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Inhibitory effects of selected plant essential oils on the growth of four pathogenic bacteria: E. Coli O157: H7, Salmonella typhimurium, staphylococcus aureus and listeria monocytogenes. Food Control 2007, 18, 414–420. [Google Scholar] [CrossRef]
- Hazzit, M.; Baaliouamer, A.; Veríssimo, A.; Faleiro, M.; Miguel, M. Chemical composition and biological activities of Algerian Thymus oils. Food Chem. 2009, 116, 714–721. [Google Scholar] [CrossRef]
- Ultee, A.; Bennik, M.; Moezelaar, R. The phenolic hydroxyl group of carvacrol is essential for action against the food-borne pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Nguefack, J.; Budde, B.B.; Jakobsen, M. Five essential oils from aromatic plants of Cameroon: Their antibacterial activity and ability to permeabilize the cytoplasmic membrane of Listeria innocua examined by flow cytometry. Lett. Appl. Microbiol. 2004, 39, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Ferreira, S.; Queiroz, J.A.; Domingues, F.C. Coriander (Coriandrum sativum L.) essential oil: Its antibacterial activity and mode of action evaluated by flow cytometry. J. Med. Microbiol. 2011, 60, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Ohmizo, C.; Yata, M.; Katsu, T. Bacterial cytoplasmic membrane permeability assay using ion-selective electrodes. J. Microbiol. Methods 2004, 59, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; McLean, R.J. Quorum signal inhibitors and their potential use against fish diseases. J. Aquat. Anim. Health 2016, 28, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Regina de Vasconcelos Goulart, F.; de Almeida Rodrigues, I.; Abreu, F.; Lins, U.; Alves, P.B.; Blank, A.F.; Valoni, É.; Sebastián, G.V.; Alviano, D.S. Antimicrobial action and anti-corrosion effect against sulfate reducing bacteria by lemongrass (Cymbopogon citratus) essential oil and its major component, the citral. AMB Express 2013, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-N.; Khan, I.; Kang, S.C. Chemical composition, mechanism of antibacterial action and antioxidant activity of leaf essential oil of Forsythia koreana deciduous shrub. Asian Pac. J. Trop. Med. 2015, 8, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, É.Y.; Baptista, E.B.; Resende Do Carmo, A.M.; Chaves, M.; Afonso, M.D.G.; Chicourel, E.L.; Barbosa Raposo, N.R. Potential of the essential oil from Pimenta pseudocaryophyllus as an antimicrobial agent. Acta Pharm. 2014, 64, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Z.-W.; Yin, Z.-Q.; Wei, Q.; Jia, R.-Y.; Zhou, L.-J.; Xu, J.; Song, X.; Zhou, Y.; Du, Y.-H. Antibacterial activity of leaf essential oil and its constituents from Cinnamomum longepaniculatum. Int. J. Clin. Exp. Med. 2014, 7, 1721. [Google Scholar] [PubMed]
- Cox, S.; Mann, C.; Markham, J.; Bell, H.; Gustafson, J.; Warmington, J.; Wyllie, S. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Appl. Microbiol. 2000, 88, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Bouhdid, S.; Abrini, J.; Zhiri, A.; Espuny, M.; Manresa, A. Investigation of functional and morphological changes in Pseudomonas aeruginosa and staphylococcus aureus cells induced by Origanum compactum essential oil. J. Appl. Microbiol. 2009, 106, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Bouhdid, S.; Abrini, J.; Amensour, M.; Zhiri, A.; Espuny, M.; Manresa, A. Functional and ultrastructural changes in Pseudomonas aeruginosa and Staphylococcus aureus cells induced by Cinnamomum verum essential oil. J. Appl. Microbiol. 2010, 109, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; Sharma, A.; Baek, K.-H. Antibacterial mode of action of ginkgo biloba leaf essential oil: Effect on morphology and membrane permeability. Bangladesh J. Pharmacol. 2015, 10, 337–350. [Google Scholar] [CrossRef]
- Patra, J.K.; Baek, K.-H. Antibacterial activity and action mechanism of the essential oil from Enteromorpha linza L. Against foodborne pathogenic bacteria. Molecules 2016, 21, 388. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, R.; Ghaderi, L.; Rafati, H.; Aliahmadi, A.; McClements, D.J. Superior antibacterial activity of nanoemulsion of thymus daenensis essential oil against E. coli. Food Chem. 2016, 194, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.; Phillips, C. The mechanism of action of a citrus oil blend against Enterococcus faecium and Enterococcus faecalis. J. Appl. Microbiol. 2009, 106, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Zeidán-Chuliá, F.; Keskin, M.; Könönen, E.; Uitto, V.-J.; Söderling, E.; Moreira, J.C.F.; Gürsoy, U.K. Antibacterial and antigelatinolytic effects of Satureja hortensis L. essential oil on epithelial cells exposed to Fusobacterium nucleatum. J. Med. Food. 2015, 18, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish Oregano, Chinese Cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli o157: H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.A.; Zahin, M.; Hasan, S.; Husain, F.M.; Ahmad, I. Inhibition of quorum sensing regulated bacterial functions by plant essential oils with special reference to clove oil. Lett. Appl. Microbiol. 2009, 49, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Szabó, M.Á.; Varga, G.Z.; Hohmann, J.; Schelz, Z.; Szegedi, E.; Amaral, L.; Molnár, J. Inhibition of quorum-sensing signals by essential oils. Phytother. Res. 2010, 24, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.V.; Ortega-Ramirez, L.A.; Gutierrez-Pacheco, M.M.; Bernal-Mercado, A.T.; Rodriguez-Garcia, I.; Gonzalez-Aguilar, G.A.; Ponce, A.; Moreira, M.D.R.; Roura, S.I.; Ayala-Zavala, J.F. Oregano essential oil-pectin edible films as anti-quorum sensing and food antimicrobial agents. Front. Microbiol. 2014, 5, 699. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I.; Khan, M.S.; Ahmad, E.; Tahseen, Q.; Khan, M.S.; Alshabib, N.A. Sub-MICs of Mentha piperita essential oil and menthol inhibits AHL mediated quorum sensing and biofilm of Gram-negative bacteria. Front. Microbiol. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Mith, H.; Clinquart, A.; Zhiri, A.; Daube, G.; Delcenserie, V. The impact of oregano (Origanum heracleoticum) essential oil and carvacrol on virulence gene transcription by Escherichia coli O157: H7. FEMS Microbiol Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sepahi, E.; Tarighi, S.; Ahmadi, F.S.; Bagheri, A. Inhibition of quorum sensing in Pseudomonas aeruginosa by two herbal essential oils from apiaceae family. J. Microbiol. 2015, 53, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.; Krishnan, T.; Yiap, B.; Hu, C.; Chan, K.G.; Lim, S. Membrane disruption and anti-quorum sensing effects of synergistic interaction between Lavandula angustifolia (lavender oil) in combination with antibiotic against plasmid-conferred multi-drug-resistant Escherichia coli. J. Appl. Microbiol. 2014, 116, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.S.X.; Krishnan, T.; Chan, K.-G.; Lim, S. Antibacterial mode of action of Cinnamomum verum bark essential oil, alone and in combination with piperacillin, against a multi-drug-resistant Escherichia coli strain. J. Microbiol. Biotechnol. 2015, 25, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Guinoiseau, E.; Lorenzi, V.; Luciani, A.; Tomi, F.; Casanova, J.; Berti, L. Susceptibility of the multi-drug resistant strain of Enterobacter aerogenesea 289 to the terpene alcohols from Cistus ladaniferus essential oil. Nat. Prod. Commun. 2011, 6, 1159–1162. [Google Scholar] [PubMed]
- Aleksic, V.; Mimica-Dukic, N.; Simin, N.; Nedeljkovic, N.S.; Knezevic, P. Synergistic effect of Myrtus communis L. Essential oils and conventional antibiotics against multi-drug resistant Acinetobacter baumannii wound isolates. Phytomedicine 2014, 21, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, P.; Aleksic, V.; Simin, N.; Svircev, E.; Petrovic, A.; Mimica-Dukic, N. Antimicrobial activity of Eucalyptus camaldulensis essential oils and their interactions with conventional antimicrobial agents against multi-drug resistant Acinetobacter baumannii. J. Ethnopharmacol. 2016, 178, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Wang, Z.; Pu, X.; Zhou, S. Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicol. Appl. Pharmacol. 2011, 254, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Larghi, P.; Rimoldi, M.; Totaro, M.G.; Allavena, P.; Mantovani, A.; Sica, A. Cellular and molecular pathways linking inflammation and cancer. Immunobiology 2009, 214, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Storz, P. Reactive oxygen species in tumor progression. Front. Biosci. 2005, 10, 1881–1896. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. Apoptosis-targeted therapies for cancer. Cancer Cell 2003, 3, 17–22. [Google Scholar] [CrossRef]
- Gautam, N.; Mantha, A.K.; Mittal, S. Essential oils and their constituents as anticancer agents: A mechanistic view. BioMed Res. Int. 2014, 2014, 154106. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R.; Menichini, F.; Saab, A.M.; Statti, G.A.; Menichini, F. Cytotoxic activity of essential oils from Labiatae and Lauraceae families against in vitro human tumor models. Anticancer Res. 2007, 27, 3293–3299. [Google Scholar] [PubMed]
- Cha, J.-D.; Kim, Y.-H.; Kim, J.-Y. Essential oil and 1, 8-cineole from Artemisia lavandulaefolia induces apoptosis in KB cells via mitochondrial stress and caspase activation. Food Sci. Biotechnol. 2010, 19, 185–191. [Google Scholar] [CrossRef]
- Russo, A.; Formisano, C.; Rigano, D.; Senatore, F.; Delfine, S.; Cardile, V.; Rosselli, S.; Bruno, M. Chemical composition and anticancer activity of essential oils of mediterranean sage (Salvia officinalis L.) grown in different environmental conditions. Food Chem. Toxicol. 2013, 55, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Aljada, A.; Rizvi, S.A.; Nasr, A.; Alaskar, A.S.; Williams, J.D. In vitro cytotoxicity of Artemisia vulgaris L. essential oil is mediated by a mitochondria-dependent apoptosis in HL-60 leukemic cell line. BMC Complement. Altern. Med. 2014, 14, 226. [Google Scholar] [CrossRef] [PubMed]
- Legault, J.; Dahl, W.; Debiton, E.; Pichette, A.; Madelmont, J.-C. Antitumor activity of balsam fir oil: Production of reactive oxygen species induced by α-humulene as possible mechanism of action. Planta Med. 2003, 69, 402–407. [Google Scholar] [PubMed]
- Paik, S.-Y.; Koh, K.-H.; Beak, S.-M.; Paek, S.-H.; Kim, J.-A. The essential oils from Zanthoxylum schinifolium pericarp induce apoptosis of HepG2 human hepatoma cells through increased production of reactive oxygen species. Biol. Pharm. Bull. 2005, 28, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Sœur, J.; Marrot, L.; Perez, P.; Iraqui, I.; Kienda, G.; Dardalhon, M.; Meunier, J.-R.; Averbeck, D.; Huang, M.-E. Selective cytotoxicity of Aniba rosaeodora essential oil towards epidermoid cancer cells through induction of apoptosis. Mutat. Res. 2011, 718, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.-R.; Park, J.S.; Park, Y.-K.; Chae, Y.Z.; Lee, G.-H.; Park, G.-Y.; Jang, B.-C. Pinus densiflora leaf essential oil induces apoptosis via ROS generation and activation of caspases in YD-8 human oral cancer cells. Int. J. Oncol. 2012, 40, 1238–1245. [Google Scholar] [PubMed]
- Queiroz, R.M.d.; Takiya, C.M.; Guimarães, L.P.T.P.; Rocha, G.D.G.; Alviano, D.S.; Blank, A.F.; Alviano, C.S.; Gattass, C.R. Apoptosis-inducing effects of Melissa officinalis L. essential oil in Glioblastoma multiforme cells. Cancer Investig. 2014, 32, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Navarra, M.; Ferlazzo, N.; Cirmi, S.; Trapasso, E.; Bramanti, P.; Lombardo, G.E.; Minciullo, P.L.; Calapai, G.; Gangemi, S. Effects of bergamot essential oil and its extractive fractions on SH-SY5Y human neuroblastoma cell growth. J. Pharm. Pharmacol. 2015, 67, 1042–1053. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Hsieh, F.; Song, H.; Lin, J. Elevated phosphorylation and activation of PDK-1/AKT pathway in human breast cancer. Br. J. Cancer. 2005, 93, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- Cicenas, J.; Urban, P.; Vuaroqueaux, V.; Labuhn, M.; Küng, W.; Wight, E.; Mayhew, M.; Eppenberger, U.; Eppenberger-Castori, S. Increased level of phosphorylated akt measured by chemiluminescence-linked immunosorbent assay is a predictor of poor prognosis in primary breast cancer overexpressing ErbB-2. Breast Cancer Res. 2005, 7, R394–R401. [Google Scholar] [CrossRef] [PubMed]
- Suhail, M.M.; Wu, W.; Cao, A.; Mondalek, F.G.; Fung, K.-M.; Shih, P.-T.; Fang, Y.-T.; Woolley, C.; Young, G.; Lin, H.-K. Boswellia sacra essential oil induces tumor cell-specific apoptosis and suppresses tumor aggressiveness in cultured human breast cancer cells. BMC Complement Altern. Med. 2011, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Suhail, M.M.; Yang, Q.; Cao, A.; Fung, K.-M.; Postier, R.G.; Woolley, C.; Young, G.; Zhang, J.; Lin, H.-K. Frankincense essential oil prepared from hydrodistillation of Boswellia sacra gum resins induces human pancreatic cancer cell death in cultures and in a xenograft murine model. BMC Complement. Altern. Med. 2012, 12, 253. [Google Scholar] [CrossRef] [PubMed]
- Seal, S.; Chatterjee, P.; Bhattacharya, S.; Pal, D.; Dasgupta, S.; Kundu, R.; Mukherjee, S.; Bhattacharya, S.; Bhuyan, M.; Bhattacharyya, P.R. Vapor of volatile oils from Litsea cubeba seed induces apoptosis and causes cell cycle arrest in lung cancer cells. PLoS ONE 2012, 7, e47014. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Chen, Y.; Hsi, Y.-T.; Chang, C.-S.; Huang, L.-F.; Ho, C.-T.; Way, T.-D.; Kao, J.-Y. Chemical constituents and anticancer activity of Curcuma zedoaria roscoe essential oil against non-small cell lung carcinoma cells in vitro and in vivo. J. Agric. Food Chem. 2013, 61, 11418–11427. [Google Scholar] [CrossRef] [PubMed]
- Pathania, A.S.; Guru, S.K.; Verma, M.; Sharma, C.; Abdullah, S.T.; Malik, F.; Chandra, S.; Katoch, M.; Bhushan, S. Disruption of the PI3K/AKT/mTOR signaling cascade and induction of apoptosis in HL-60 cells by an essential oil from monarda citriodora. Food Chem. Toxicol. 2013, 62, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-M.; Lee, E.-O.; Kim, S.-H.; Lee, H.-J. Essential oil of Pinus koraiensis inhibits cell proliferation and migration via inhibition of p21-activated kinase 1 pathway in HCT116 colorectal cancer cells. BMC Complement. Altern. Med. 2014, 14, 275. [Google Scholar] [CrossRef] [PubMed]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.D.; Moon, S.E.; Kim, H.Y.; Cha, I.H.; Lee, K.Y. Essential oil of Artemisia capillaris induces apoptosis in KB cells via mitochondrial stress and caspase activation mediated by MAPK-stimulated signaling pathway. J. Food Sci. 2009, 74, T75–T81. [Google Scholar] [CrossRef] [PubMed]
- Sertel, S.; Eichhorn, T.; Plinkert, P.K.; Efferth, T. Cytotoxicity of Thymus vulgaris essential oil towards human oral cavity squamous cell carcinoma. Anticancer Res. 2011, 31, 81–87. [Google Scholar] [PubMed]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.-W. NF-kappaB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Pikarsky, E.; Ben-Neriah, Y. NF-kappaB inhibition: A double-edged sword in cancer? Eur. J. Cancer 2006, 42, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Malik, F.; Bhushan, S.; Sethi, V.K.; Shahi, A.K.; Taneja, S.C.; Qazi, G.N.; Singh, J. An essential oil and its major constituent isointermedeol induce apoptosis by increased expression of mitochondrial cytochrome c and apical death receptors in human leukaemia HL-60 cells. Chem. Biol. Interact. 2008, 171, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Choi, J.; Lou, Z.; Jiang, X.; Lee, S.-H. Patchouli alcohol, an essential oil of pogostemon cablin, exhibits anti-tumorigenic activity in human colorectal cancer cells. Int. Immunopharmacol. 2013, 16, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.-B.; Ky, N.; Ng, H.-M.; Hamza, M.S.; Zhao, Y. Curcuma wenyujin extract induces apoptosis and inhibits proliferation of human cervical cancer cells in vitro and in vivo. Integr. Cancer Ther. 2010, 9, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-K.; Hwang, J.-S.; So, J.-S.; Lee, C.-G.; Sahoo, A.; Ryu, J.-H.; Jeon, W.K.; Ko, B.S.; Im, C.-R.; Lee, S.H. Cinnamon extract induces tumor cell death through inhibition of NFκB and AP1. BMC Cancer 2010, 10, 392. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.-D.; Kim, J.-Y. Essential oil from Cryptomeria japonica induces apoptosis in human oral epidermoid carcinoma cells via mitochondrial stress and activation of caspases. Molecules 2012, 17, 3890–3901. [Google Scholar] [CrossRef] [PubMed]
- Moirangthem, D.S.; Laishram, S.; Rana, V.S.; Borah, J.C.; Talukdar, N.C. Essential oil of Cephalotaxus griffithii needle inhibits proliferation and migration of human cervical cancer cells: Involvement of mitochondria-initiated and death receptor-mediated apoptosis pathways. Nat. Prod. Res. 2015, 29, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, S.; Kulshrestha, A. Fungal infections in intensive care unit: Challenges in diagnosis and management. Ann. Med. Health Sci. Res. 2013, 3, 238. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz Cabral, L.; Pinto, V.F.; Patriarca, A. Application of plant derived compounds to control fungal spoilage and mycotoxin production in foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Kurt, Ş.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Vale-Silva, L.; Gonçalves, M.; Cavaleiro, C.; Vaz, S.; Canhoto, J.; Pinto, E.; Salgueiro, L. Antifungal activity of phenolic-rich Lavandula multifida L. essential oil. Eur. J. Clin. Microbial. Infect. Dis. 2012, 31, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.D.; Mossini, S.A.G.; Ferreira, F.M.D.; Arrotéia, C.C.; da Costa, C.L.; Nakamura, C.V.; Machinski Junior, M. The inhibitory effects of Curcuma longa L. essential oil and curcumin on Aspergillus flavus link growth and morphology. Sci. World J. 2013, 2013, 343804. [Google Scholar] [CrossRef] [PubMed]
- Tolouee, M.; Alinezhad, S.; Saberi, R.; Eslamifar, A.; Zad, S.J.; Jaimand, K.; Taeb, J.; Rezaee, M.-B.; Kawachi, M.; Shams-Ghahfarokhi, M. Effect of Matricaria chamomilla L. flower essential oil on the growth and ultrastructure of Aspergillus niger van Tieghem. Int. J. Food Microbiol. 2010, 139, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Huang, B.; Luo, X.; Zeng, H.; Ban, X.; He, J.; Wang, Y. The control of Aspergillus flavus with Cinnamomum jensenianum Hand.-Mazz essential oil and its potential use as a food preservative. Food Chem. 2012, 130, 520–527. [Google Scholar] [CrossRef]
- Iwaki, T.; Iefuji, H.; Hiraga, Y.; Hosomi, A.; Morita, T.; Giga-Hama, Y.; Takegawa, K. Multiple functions of ergosterol in the fission yeast Schizosaccharomyces pombe. Microbiology 2008, 154, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, A.; Kumar, P.; Bhatt, R.; Manzoor, N. Antifungal activity of Coriaria nepalensis essential oil by disrupting ergosterol biosynthesis and membrane integrity against Candida. Yeast 2011, 28, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Ocimum sanctum essential oil and its active principles exert their antifungal activity by disrupting ergosterol biosynthesis and membrane integrity. Res. Microbial. 2010, 161, 816–823. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Freires, I.; Murata, R.M.; Furletti, V.F.; Sartoratto, A.; de Alencar, S.M.; Figueira, G.M.; de Oliveira Rodrigues, J.A.; Duarte, M.C.T.; Rosalen, P.L. Coriandrum sativum L. (coriander) essential oil: Antifungal activity and mode of action on Candida spp., and molecular targets affected in human whole-genome expression. PLoS ONE 2014, 9, e99086. [Google Scholar]
- Samber, N.; Khan, A.; Varma, A.; Manzoor, N. Synergistic anti-candidal activity and mode of action of Mentha piperita essential oil and its major components. Pharm. Boil. 2015, 53, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zeng, H.; Tian, J.; Ban, X.; Ma, B.; Wang, Y. Dill (Anethum graveolens L.) seed essential oil induces Candida albicans apoptosis in a metacaspase-dependent manner. Fungal Boil. 2014, 118, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.; Cardoso, T.; Ferreira, F.; Fernandes-Ferreira, M.; Piper, P.; Sousa, M.J. Mentha piperita essential oil induces apoptosis in yeast associated with both cytosolic and mitochondrial ROS-mediated damage. FEMS Yeast Res. 2014, 14, 1006–1014. [Google Scholar] [PubMed]
- Bagheri-Gavkosh, S.; Bigdeli, M.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Inhibitory effects of ephedra major host on Aspergillus parasiticus growth and aflatoxin production. Mycopathologia 2009, 168, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Mishra, A.K.; Dubey, N.; Tripathi, Y. Evaluation of Chenopodium ambrosioides oil as a potential source of antifungal, antiaflatoxigenic and antioxidant activity. Int. J. Food Microbiol. 2007, 115, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Rasooli, I.; Owlia, P. Chemoprevention by thyme oils of Aspergillus parasiticus growth and aflatoxin production. Phytochemistry 2005, 66, 2851–2856. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Shukla, R.; Singh, P.; Kumar, A.; Mishra, P.K.; Dubey, N.K. Efficacy of chemically characterized Piper betle L. essential oil against fungal and aflatoxin contamination of some edible commodities and its antioxidant activity. Int. J. Food Microbiol. 2010, 142, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M. Effect of Carum copticum essential oil on growth and aflatoxin formation by Aspergillus strains. Nat. Prod. Res. 2015, 29, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Sonker, N.; Singh, P. Efficacy of some essential oils against Aspergillus flavus with special reference to Lippia alba oil an inhibitor of fungal proliferation and aflatoxin B1 production in green gram seeds during storage. J. Food Sci. 2016, 81, M928. [Google Scholar] [CrossRef] [PubMed]
- Yahyaraeyat, R.; Khosravi, A.; Shahbazzadeh, D.; Khalaj, V. The potential effects of Zataria multiflora Boiss essential oil on growth, aflatoxin production and transcription of aflatoxin biosynthesis pathway genes of toxigenic Aspergillus parasiticus. Braz. J. Microbiol. 2013, 44, 649–655. [Google Scholar]
- Singh, N. Drug resistance mechanisms in clinical isolates of Leishmania donovani. Indian J. Med. Res. 2006, 123, 411. [Google Scholar] [PubMed]
- Antony, H.A.; Parija, S.C. Antimalarial drug resistance: An overview. Trop. Parasitol. 2016, 6, 30. [Google Scholar] [PubMed]
- Ellse, L.; Wall, R. The use of essential oils in veterinary ectoparasite control: A review. Med. Vet. Entomol. 2014, 28, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.; Wilkinson, J.M.; Cavanagh, H.M. Antiparasitic activity of two Lavandula essential oils against Giardia duodenalis, Trichomonas vaginalis and Hexamita inflata. Parasitol. Res. 2006, 99, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L.; Nance, M.R.; Garcia, M.; Scull, R.; Setzer, W.N. Comparative chemical, cytotoxicity and antileishmanial properties of essential oils from Chenopodium ambrosioides. Nat. Prod. Commun. 2011, 6, 281–286. [Google Scholar] [PubMed]
- Monzote, L.; García, M.; Pastor, J.; Gil, L.; Scull, R.; Maes, L.; Cos, P.; Gille, L. Essential oil from Chenopodium ambrosioides and main components: Activity against Leishmania, their mitochondria and other microorganisms. Exp. Parasitol. 2014, 136, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Esperandim, V.R.; da Silva Ferreira, D.; Rezende, K.C.S.; Magalhães, L.G.; Souza, J.M.; Pauletti, P.M.; Januario, A.H.; de Laurentz, R.D.S.; Bastos, J.K.; Símaro, G.V. In vitro antiparasitic activity and chemical composition of the essential oil obtained from the fruits of Piper cubeba. Planta Med. 2013, 79, 1653–1655. [Google Scholar] [CrossRef] [PubMed]
- Kpoviessi, S.; Bero, J.; Agbani, P.; Gbaguidi, F.; Kpadonou-Kpoviessi, B.; Sinsin, B.; Accrombessi, G.; Frédérich, M.; Moudachirou, M.; Quetin-Leclercq, J. Chemical composition, cytotoxicity and in vitro antitrypanosomal and antiplasmodial activity of the essential oils of four Cymbopogon species from Benin. J. Ethnopharmacol. 2014, 151, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Kpoviessi, B.G.K.; Kpoviessi, S.D.; Ladekan, E.Y.; Gbaguidi, F.; Frédérich, M.; Moudachirou, M.; Quetin-Leclercq, J.; Accrombessi, G.C.; Bero, J. In vitro antitrypanosomal and antiplasmodial activities of crude extracts and essential oils of Ocimum gratissimum linn from Benin and influence of vegetative stage. J. Ethnopharmacol. 2014, 155, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, D.; Tierney, M.; Sen, R.; Bergonzi, M.C.; Demirci, B.; Bilia, A.R.; Baser, K.H.C.; Brun, R.; Chatterjee, M. Antiprotozoal effect of Artemisia indica extracts and essential oil. Planta Med. 2015, 81, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Díaz, R.A.; Ibáñez-Escribano, A.; Burillo, J.; de las Heras, L.; del Prado, G.; Agulló-Ortuño, M.T.; Julio, L.F.; González-Coloma, A. Trypanocidal, trichomonacidal and cytotoxic components of cultivated Artemisia absinthium linnaeus (Asteraceae) essential oil. Mem. Inst. Oswaldo Cruz 2015, 110, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.A.; de Melo, N.I.; Aguiar, D.P.; de Oliveira, P.F.; Groppo, M.; da Silva Filho, A.A.; Rodrigues, V.; Cunha, W.R.; Tavares, D.C.; Magalhães, L.G. Anthelmintic effects of the essential oil of fennel (Foeniculum vulgare Mill., Apiaceae) against Schistosoma mansoni. Chem. Biodivers. 2015, 12, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.L.S.; dos Santos, J.C.; Bordignon, S.A.; Apel, M.A.; Henriques, A.T.; von Poser, G.L. Acaricidal properties of the essential oil from Hesperozygis ringens (Lamiaceae) on the cattle tick Riphicephalus (Boophilus) microplus. Bioresour. Technol. 2010, 101, 2506–2509. [Google Scholar] [CrossRef] [PubMed]
- De Souza Chagas, A.C.; de Sena Oliveira, M.C.; Giglioti, R.; Santana, R.C.M.; Bizzo, H.R.; Gama, P.E.; Chaves, F.C.M. Efficacy of 11 Brazilian essential oils on lethality of the cattle tick Rhipicephalus (Boophilus) microplus. Ticks Tick-Borne Dis. 2016, 7, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Li, A.Y.; Junior, L.M.C.; Castro-Arellano, I.; Liu, J. Evaluation of DEET and eight essential oils for repellency against nymphs of the lone star tick, Amblyomma americanum (Acari: Ixodidae). Exp. Appl. Acarol. 2016, 68, 241–249. [Google Scholar] [CrossRef] [PubMed]
- De Santana Campos, R.N.; Lima, C.B.N.; Oliveira, A.P.; Araújo, A.P.A.; Blank, A.F.; Alves, P.B.; Lima, R.N.; Araújo, V.A.; Santana, A.S.; Bacci, L. Acaricidal properties of vetiver essential oil from Chrysopogon zizanioides (Poaceae) against the tick species amblyomma cajennense and Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Vet. Parasitol. 2015, 212, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L.; Garcia, M.; Montalvo, A.; Linares, R.; Scull, R. Effect of oral treatment with the essential oil from Chenopodium ambrosioides against cutaneous leishmaniasis in BALB/c mice, caused by Leishmania amazonensis. Forsch. Komplementmed. 2009, 16, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Islamuddin, M.; Chouhan, G.; Want, M.Y.; Tyagi, M.; Abdin, M.Z.; Sahal, D.; Afrin, F. Leishmanicidal activities of Artemisia annua leaf essential oil against visceral Leishmaniasis. Front. Microbial. 2014, 5, 626. [Google Scholar] [CrossRef] [PubMed]
- Islamuddin, M.; Sahal, D.; Afrin, F. Apoptosis-like death in Leishmania donovani promastigotes induced by eugenol-rich oil of Syzygium aromaticum. J. Med. Microbial. 2014, 63, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Misra, P.; Kumar, A.; Khare, P.; Gupta, S.; Kumar, N.; Dube, A. Pro-apoptotic effect of the landrace Bangla Mahoba of Piper betle on Leishmania donovani may be due to the high content of eugenol. J. Med. Microbial. 2009, 58, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Dinis, A.; Santos-Rosa, M.; Alves, V.; Salgueiro, L.; Cavaleiro, C.; Sousa, M. Activity of Thymus capitellatus volatile extract, 1, 8-cineole and borneol against Leishmania species. Vet. Parasitol. 2014, 200, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Grbovic, S.; Orcic, D.; Couladis, M.; Jovin, E.; Bugarin, D.; Balog, K.; Mimica-Dukic, N. Variation of essential oil composition of Eucalyptus camaldulensis (Myrtaceae) from the Montengero coastline. Acta Period. Technol. 2010, 41, 151–158. [Google Scholar] [CrossRef]
- Barr, A.; Chapman, J.; Smith, N.; Wightman, G.; Knight, T.; Mills, L.; Andrews, M.; Alexander, V. Traditional Aboriginal Medicines in the Northern Territory of Australia by Aboriginal Communities of the Northern Territory; Conservation Commission of the Northern Territory of Australia: Darwin, Australia, 1993. [Google Scholar]
- Iscan, G.; KIrimer, N.; Kürkcüoglu, M.N.; Baser, H.C.; Demirci, F. Antimicrobial screening of Mentha piperita essential oils. J. Agric. Food Chem. 2002, 50, 3943–3946. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Kahla-Nakbi, A.B.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): A short review. Phytother. Res. 2007, 21, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and Dermatophyte species. J. Med. Microbial. 2009, 58, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Parle, M.; Dhingra, D.; Kulkarni, S. Improvement of mouse memory by Myristica fragrans seeds. J. Med. Food 2004, 7, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Olajide, O.A.; Ajayi, F.F.; Ekhelar, A.I.; Awe, S.O.; Makinde, J.M.; Alada, A. Biological effects of Myristica fragrans (nutmeg) extract. Phytother. Res. 1999, 13, 344–345. [Google Scholar] [CrossRef]
- Sonavane, G.; Sarveiya, V.; Kasture, V.; Kasture, S. Anxiogenic activity of Myristica fragrans seeds. Pharmacol. Biochem. Behav. 2002, 71, 239–244. [Google Scholar] [CrossRef]
- Wiesehöfer, J. Ancient Persia; St Martin’s Press: New York, NY, USA, 2006. [Google Scholar]
- Dadegi, F. 9th Century AD/2006. Bondahesh; Tous Press: Tehran, Iran, 2006. (In Persian) [Google Scholar]
- Biruni, A. 10th Century AD/2004. Al Seydaneh fi al Teb; University Press: Tehran, Iran, 2004. (In Persian) [Google Scholar]
- Mikaili, P.; Shayegh, J.; Sarahroodi, S.; Sharifi, M. Pharmacological properties of herbal oil extracts used in Iranian traditional medicine. Adv. Environ. Biol. 2012, 6, 153–158. [Google Scholar]
- Heravi, M.G. Qarabadin-e-Salehi; Dar-ol-Khalafeh: Tehran, Iran, 1765. (In Persian) [Google Scholar]
- Avicenna. 1025/1988. Canon of Medicine; Soroosh Press: Tehran, Iran, 1988; Volume 5. (In Persian) [Google Scholar]
- Tonekaboni, H. 1670/2007. Tohfat ol Momenin; Nashre Shahr Press: Tehran, Iran, 2007. (In Persian) [Google Scholar]
- Campbell, K.; Glatz, C.E. Mechanisms of aqueous extraction of soybean oil. J. Agric. Food Chem. 2009, 57, 10904–10912. [Google Scholar] [CrossRef] [PubMed]
- Yucel, I.; Guzin, G. Topical henna for capecitabine induced hand-foot syndrome. Investig. New Drugs 2008, 26, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Daniel, A.N.; Sartoretto, S.M.; Schmidt, G.; Caparroz-Assef, S.M.; Bersani-Amado, C.A.; Cuman, R.K.N. Anti-inflammatory and antinociceptive activities a of eugenol essential oil in experimental animal models. Rev. Bras. Farmacogn. 2009, 19, 212–217. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Ghannadi, A.; Hajiloo, M. Analgesic and anti-inflammatory effects of Rosa damascena hydroalcoholic extract and its essential oil in animal models. Iran. J. Pharm. Res. 2010, 163–168. [Google Scholar]
- Harries, N.; James, K.; Pugh, W. Antifoaming and carminative actions of volatile oils. J. Clin. Pharm. Ther. 1977, 2, 171–177. [Google Scholar] [CrossRef]
- Carvalho-Freitas, M.I.R.; Costa, M. Anxiolytic and sedative effects of extracts and essential oil from Citrus aurantium L. Biol. Pharm. Bull. 2002, 25, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Giner-Larza, E.M.; Máñez, S.; Giner, R.M.; Recio, M.C.; Prieto, J.M.; Cerdá-Nicolás, M.; Ríos, J. Anti-inflammatory triterpenes from Pistacia terebinthus galls. Planta Med. 2002, 68, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.E.; Senol, F.S.; Gulpinar, A.R.; Sekeroglu, N.; Kartal, M.; Sener, B. Neuroprotective potential of some terebinth coffee brands and the unprocessed fruits of Pistacia terebinthus L. and their fatty and essential oil analyses. Food Chem. 2012, 130, 882–888. [Google Scholar] [CrossRef]
- Kanter, M.; Coskun, O.; Kalayc, M.; Buyukbas, S.; Cagavi, F. Neuroprotective effects of Nigella sativa on experimental spinal cord injury in rats. Hum. Exp. Toxicol. 2006, 25, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.O.; Blumberg, J.B. In vitro activity of almond skin polyphenols for scavenging free radicals and inducing quinone reductase. J. Agric. Food Chem 2008, 56, 4427–4434. [Google Scholar] [CrossRef] [PubMed]
- Pourgholami, M.; Kamalinejad, M.; Javadi, M.; Majzoob, S.; Sayyah, M. Evaluation of the anticonvulsant activity of the essential oil of Eugenia caryophyllata in male mice. J. Ethnopharmacol. 1999, 64, 167–171. [Google Scholar] [CrossRef]
- Pourgholami, M.; Majzoob, S.; Javadi, M.; Kamalinejad, M.; Fanaee, G.; Sayyah, M. The fruit essential oil of Pimpinella anisum exerts anticonvulsant effects in mice. J. Ethnopharmacol. 1999, 66, 211–215. [Google Scholar] [CrossRef]
- Sayyah, M.; Saroukhani, G.; Peirovi, A.; Kamalinejad, M. Analgesic and anti-inflammatory activity of the leaf essential oil of Laurus nobilis Linn. Phytother. Res. 2003, 17, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hungju, F. Studies on the essential oils of clove buds and clove leaves. Zhong Caoyao 1981, 12, 339–342. [Google Scholar]
- Dallmeier, K.; Carlini, E. Anesthetic, hypothermic, myorelaxant and anticonvulsant effects of synthetic eugenol derivatives and natural analogues. Pharmacology 1981, 22, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Firuzi, O.; Asadollahi, M.; Gholami, M.; Javidnia, K. Composition and biological activities of essential oils from four Heracleum species. Food Chem. 2010, 122, 117–122. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Miri, A.; Hoseini-Alfatemi, S.M.; Sharifi-Rad, M.; Setzer, W.N.; Hadjiakhoondi, A. Chemical composition and biological activity of Pulicaria vulgaris essential oil from Iran. Nat. Prod. Commun. 2014, 9, 1633–1636. [Google Scholar] [PubMed]
- Sharifi-Rad, J.; Hoseini-Alfatemi, S.M.; Sharifi-Rad, M.; Sharifi-Rad, M.; Iriti, M.; Sharifi-Rad, M.; Sharifi-Rad, R.; Raeisi, S. Phytochemical compositions and biological activities of essential oil from Xanthium strumarium L. Molecules 2015, 20, 7034–7047. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Sharifi-Rad, M.; Hoseini-Alfatemi, S.M.; Iriti, M.; Sharifi-Rad, M.; Sharifi-Rad, M. Composition, cytotoxic and antimicrobial activities of Satureja intermedia CA Mey essential oil. Int. J. Mol. Sci. 2015, 16, 17812–17825. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Hoseini-Alfatemi, S.M.; Sharifi-Rad, M.; Setzer, W.N. Chemical composition, antifungal and antibacterial activities of essential oil from Lallemantia royleana (Benth. In Wall.) Benth. J. Food Saf. 2015, 35, 19–25. [Google Scholar] [CrossRef]
- Abdollahi, M.; Hamzehzarghani, H.; Saharkhiz, M.J. Effects of the essential oil of Zataria multiflora boiss, a thyme-like medicinal plant from Iran on the growth and sporulation of Aspergillus niger both in vitro and on lime fruits. J. Food Saf. 2011, 31, 424–432. [Google Scholar] [CrossRef]
- Razzaghi-Abyaneh, M.; Shams-Ghahfarokhi, M.; Yoshinari, T.; Rezaee, M.-B.; Jaimand, K.; Nagasawa, H.; Sakuda, S. Inhibitory effects of Satureja hortensis L. essential oil on growth and aflatoxin production by Aspergillus parasiticus. Int. J. Food Microbiol. 2008, 123, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Piri, K. Antifungal effects of two medicinal plant native to Iran. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 2712–2715. [Google Scholar]
- Shokri, H.; Sharifzadeh, A.; Tamai, I.A. Anti-Candida zeylanoides activity of some Iranian plants used in traditional medicine. J. Mycol. Méd./J. Med. Mycol. 2012, 22, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Naeini, A.; Khosravi, A.; Chitsaz, M.; Shokri, H.; Kamlnejad, M. Anti-Candida albicans activity of some Iranian plants used in traditional medicine. J. Mycol. Méd./J. Med. Mycol. 2009, 19, 168–172. [Google Scholar] [CrossRef]
- Sheikhan, F.; Jahdi, F.; Khoei, E.M.; Shamsalizadeh, N.; Sheikhan, M.; Haghani, H. Episiotomy pain relief: Use of lavender oil essence in primiparous Iranian women. Complement. Ther. Clin. Pract. 2012, 18, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Ghelardini, C.; Galeotti, N.; Salvatore, G.; Mazzanti, G. Local anaesthetic activity of the essential oil of Lavandula angustifolia. Planta Med. 1999, 65, 700–703. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, Y.; Hara, C.; Tamura, K.; Fujii, T.; Nakamura, K.-I.; Masujima, T.; Aoki, T. Sedative effect on humans of inhalation of essential oil of linalool: Sensory evaluation and physiological measurements using optically active linalools. Anal. Chim. Acta 1998, 365, 293–299. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Hart, S. Studies on the mode of action of the essential oil of Lavender (Lavandula angustifolia P. Miller). Phytother. Res. 1999, 13, 540–542. [Google Scholar] [CrossRef]
- Raisi, D.Z.; Hosseini, B.F.; Bekhradi, R. Effect of lavender inhalation on the symptoms of primary dysmenorrhea and the amount of menstrual bleeding: A randomized clinical trial. Complement. Ther. Med. 2014, 22, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Andersch, B.; Milsom, I. An epidemiologic study of young women with dysmenorrhea. Am. J. Obstet. Gynecol. 1982, 144, 655–660. [Google Scholar] [CrossRef]
- Bikmoradi, A.; Seifi, Z.; Poorolajal, J.; Araghchian, M.; Safiaryan, R.; Oshvandi, K. Effect of inhalation aromatherapy with lavender essential oil on stress and vital signs in patients undergoing coronary artery bypass surgery: A single-blinded randomized clinical trial. Complement. Ther. Med. 2015, 23, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Tully, P.J.; Bennetts, J.S.; Baker, R.A.; McGavigan, A.D.; Turnbull, D.A.; Winefield, H.R. Anxiety, depression, and stress as risk factors for atrial fibrillation after cardiac surgery. Heart Lung J. Acute Crit. Care 2011, 40, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Bakhsha, F.; Mazandarani, M.; Aryaei, M.; Jafari, S.Y.; Bayate, H. Phytochemical and anti-oxidant activity of Lavandula angustifolia Mill. Essential oil on preoperative anxiety in patients undergoing diagnostic curettage. Int. J. Women’s Health Reprod. Sci. 2014, 2, 268–271. [Google Scholar] [CrossRef]
- Cavanagh, H.; Wilkinson, J. Biological activities of lavender essential oil. Phytother. Res. 2002, 16, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Re, L.; Barocci, S.; Sonnino, S.; Mencarelli, A.; Vivani, C.; Paolucci, G.; Scarpantonio, A.; Rinaldi, L.; Mosca, E. Linalool modifies the nicotinic receptor—Ion channel kinetics at the mouse neuromuscular junction. Pharmacol. Res. 2000, 42, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Dash, B.; Kashyap, L. Basic Principles of Ayurveda; Concept Publishing Company: Delhi, India, 1980. [Google Scholar]
- Nutional Institue of Health, National Center for Complementary and Integrative Health. Ayurvedic Medicine: In Depth. Available online: https://nccih.nih.gov/health/ayurveda/introduction.htm (accessed on 15 June 2016).
- Mayaud, L.; Carricajo, A.; Zhiri, A.; Aubert, G. Comparison of bacteriostatic and bactericidal activity of 13 essential oils against strains with varying sensitivity to antibiotics. Lett. Appl. Microbial. 2008, 47, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.K.; Valdramidis, V.P.; O’Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P. Application of natural antimicrobials for food preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef] [PubMed]
- Ademiluyi, A.O.; Oyeleye, S.I.; Oboh, G. Biological activities, antioxidant properties and phytoconstituents of essential oil from sweet basil (Ocimum basilicum L.) leaves. Comp. Clin. Pathol. 2016, 25, 169–176. [Google Scholar] [CrossRef]
- Oboh, G.; Ogunsuyi, O.B.; Ogunbadejo, M.D.; Adefegha, S.A. Influence of gallic acid on α-amylase and α-glucosidase inhibitory properties of acarbose. J. Food Drug Anal. 2016, 24, 627–634. [Google Scholar] [CrossRef]
- Dehghan, H.; Sarrafi, Y.; Salehi, P. Antioxidant and antidiabetic activities of 11 herbal plants from Hyrcania region, Iran. J. Food Drug Anal. 2016, 24, 179–188. [Google Scholar] [CrossRef]
- Ohkuwa, T.; Sato, Y.; Naoi, M. Hydroxyl radical formation in diabetic rats induced by streptozotocin. Life Sci. 1995, 56, 1789–1798. [Google Scholar] [CrossRef]
- Oboh, G.; Akinbola, I.A.; Ademosun, A.O.; Sanni, D.M.; Odubanjo, O.V.; Olasehinde, T.A.; Oyeleye, S.I. Essential oil from clove bud (Eugenia aromatica Kuntze) inhibit key enzymes relevant to the management of type-2 diabetes and some pro-oxidant induced lipid peroxidation in rats pancreas in vitro. J. Oleo Sci. 2015, 64, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Olasehinde, T.A.; Ademosun, A.O. Essential oil from lemon peels inhibit key enzymes linked to neurodegenerative conditions and pro-oxidant induced lipid peroxidation. J. Oleo Sci. 2014, 63, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Formisano, C.; Rigano, D.; Cardile, V.; Arnold, N.A.; Senatore, F. Comparative phytochemical profile and antiproliferative activity on human melanoma cells of essential oils of three Lebanese salvia species. Ind. Crops Prod. 2016, 83, 492–499. [Google Scholar] [CrossRef]
- Kamatou, G.; Makunga, N.; Ramogola, W.; Viljoen, A. South African Salvia species: A review of biological activities and phytochemistry. J. Ethnopharmacol. 2008, 119, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Baliga, M.S.; Rao, S.; Rai, M.P.; D’souza, P. Radio protective effects of the ayurvedic medicinal plant Ocimum sanctum Linn. (Holy Basil): A memoir. J. Cancer Res. Ther. 2016, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Mabro, M.; Faivre, S.; Raymond, E. A risk-benefit assessment of amifostine in cytoprotection. Drug Saf. 1999, 21, 367–387. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Gupta, D.; Chawla, R.; Sagar, R.; Sharma, A.; Kumar, R.; Prasad, J.; Singh, S.; Samanta, N.; Sharma, R.K. Radioprotection by plant products: Present status and future prospects. Phytother. Res. 2005, 19, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Hosseinimehr, S.J. Trends in the development of radioprotective agents. Drug Discov. Today 2007, 12, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Prakash, P.; Gupta, N. Therapeutic uses of Ocimum sanctum Linn (Tulsi) with a note on eugenol and its pharmacological actions: A short review. Indian J. Physiol. Pharmacol. 2005, 49, 125. [Google Scholar] [PubMed]
- Singh, E.; Sharma, S.; Dwivedi, J.; Sharma, S. Diversified potentials of Ocimum sanctum Linn (Tulsi): An exhaustive survey. J. Nat. Prod. Plant Resour. 2012, 2, 39–48. [Google Scholar]
- Ganasoundari, A.; Devi, P.U.; Rao, B. Enhancement of bone marrow radioprotection and reduction of WR-2721 toxicity by Ocimum sanctum. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1998, 397, 303–312. [Google Scholar] [CrossRef]
- Martin, A.; Oesterheld, J.; Konradi, C.; Clein, P.; Heckers, S. Pediatric psychopharmacology, I: Pharmacokinetic and pharmacodynamic principles. Child Adolesc. Psychiatry Compr. Textb. 2002, 939–950. [Google Scholar]
- Nieoullon, A. Neurodegenerative diseases and neuroprotection: Current views and prospects. J. Appl. Biomed. 2011, 9, 173–183. [Google Scholar] [CrossRef]
- Brown, R.H. Amyotrophic lateral sclerosis and the inherited motor neuron diseases. In Molecular Neurology; Martin, J.B., Ed.; Scientific American: New York, NY, USA, 1998; pp. 223–238. [Google Scholar]
- Centonze, D.; Rossi, S.; Prosperetti, C.; Tscherter, A.; Bernardi, G.; Maccarrone, M.; Calabresi, P. Abnormal sensitivity to cannabinoid receptor stimulation might contribute to altered gamma-aminobutyric acid transmission in the striatum of R6/2 Huntington’s disease mice. Biol. Psychiatry 2005, 57, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Cicchetti, F.; Drouin-Ouellet, J.; Gross, R.E. Environmental toxins and Parkinson’s disease: What have we learned from pesticide-induced animal models? Trends Pharmacol. Sci. 2009, 30, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Moisan, F.; Spinosi, J.; Dupupet, J.L.; Delabre, L.; Mazurie, J.L.; Goldberg, M.; Imbernon, E.; Tzourio, C.; Elbaz, A. The relation between type of farming and prevalence of Parkinson’s disease among agricultural workers in five french districts. Mov. Disord. 2011, 26, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Adefeghaa, S.; Oboha, G.; Odubanjoa, T.; Ogunsuyia, O. A comparative study on the antioxidative activities, anticholinesterase properties and essential oil composition of clove (Syzygium aromaticum) bud and Ethiopian pepper (Xylopia aethiopica). Riv. Ital. Sost. Gr. 2015, 92, 257–268. [Google Scholar]
- Ademosun, A.O.; Oboh, G.; Olupona, A.J.; Oyeleye, S.I.; Adewuni, T.M.; Nwanna, E.E. Comparative study of chemical composition, in vitro inhibition of cholinergic and monoaminergic enzymes, and antioxidant potentials of essential oil from peels and seeds of sweet orange (Citrus sinensis [L.] Osbeck) fruits. J. Food Biochem. 2016, 40, 53–60. [Google Scholar] [CrossRef]
- Oboh, G.; Bello, F.O.; Ademosun, A.O.; Akinyemi, A.J.; Adewuni, T.M. Antioxidant, hypolipidemic, and anti-angiotensin-1-converting enzyme properties of lemon (Citrus limon) and lime (Citrus aurantifolia) juices. Comp. Clin. Pathol. 2015, 24, 1395–1406. [Google Scholar] [CrossRef]
- Perry, R.; Terry, R.; Watson, L.; Ernst, E. Is lavender an anxiolytic drug? A systematic review of randomised clinical trials. Phytomedicine 2012, 19, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.; Hughes, B.M. Expectancies, not aroma, explain impact of lavender aromatherapy on psychophysiological indices of relaxation in young healthy women. Br. J. Health Psychol. 2008, 13, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Braden, R.; Reichow, S.; Halm, M.A. The use of the essential oil lavandin to reduce preoperative anxiety in surgical patients. J. Perianesthesia Nurs. 2009, 24, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, A.K.; Yılmaz, E.; Çeçen, D. Effects of aroma inhalation on examination anxiety. Teach. Learn. Nurs. 2008, 3, 125–130. [Google Scholar] [CrossRef]
- Motomura, N.; Sakurai, A.; Yotsuya, Y. Reduction of mental stress with lavender odorant. Percept. Mot. Skills 2001, 93, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Muzzarelli, L.; Force, M.; Sebold, M. Aromatherapy and reducing preprocedural anxiety: A controlled prospective study. Gastroenterol. Nurs. 2006, 29, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Sgoutas-Emch, S.; Fox, T.; Preston, M.; Brooks, C.; Serber, E. Stress management: Aromatherapy as an alternative. Sci. Rev. Altern. Med. 2001, 5, 90–95. [Google Scholar]
- Toda, M.; Morimoto, K. Effect of lavender aroma on salivary endocrinological stress markers. Arch. Oral Boil. 2008, 53, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Dunn, C.; Sleep, J.; Collett, D. Sensing an improvement: An experimental study to evaluate the use of aromatherapy, massage and periods of rest in an intensive care unit. J. Adv. Nurs. 1995, 21, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Soden, K.; Vincent, K.; Craske, S.; Lucas, C.; Ashley, S. A randomized controlled trial of aromatherapy massage in a hospice setting. Palliat. Med. 2004, 18, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Uebaba, K.; Ogawa, H.; Tatsuse, T.; Wang, B.-H.; Hisajima, T.; Venkatraman, S. Pharmaco-physio-psychologic effect of Ayurvedic oil-dripping treatment using an essential oil from Lavendula angustifolia. J. Altern. Complement. Med. 2008, 14, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Morris, N. The effects of lavender (Lavendula angustifolium) baths on psychological well-being: Two exploratory randomised control trials. Complement. Ther. Med. 2002, 10, 223–228. [Google Scholar] [CrossRef]
- Bradley, B.F.; Brown, S.L.; Chu, S.; Lea, R.W. Effects of orally administered lavender essential oil on responses to anxiety-provoking film clips. Hum. Psychopharmacol. Clin. Exp. 2009, 24, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Kasper, S.; Gastpar, M.; Müller, W.E.; Volz, H.-P.; Möller, H.-J.; Dienel, A.; Schläfke, S. Silexan, an orally administered Lavandula oil preparation, is effective in the treatment of ‘subsyndromal’ anxiety disorder: A randomized, double-blind, placebo controlled trial. Int. Clin. Psychopharmacol. 2010, 25, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Woelk, H.; Schläfke, S. A multi-center, double-blind, randomised study of the Lavender oil preparation Silexan in comparison to Lorazepam for generalized anxiety disorder. Phytomedicine 2010, 17, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Moon, G. Brief review of cancer treatment focused on JIJU. J. Korean Tradit. Oncol. 2008, 11, 1–11. [Google Scholar] [CrossRef]
- Kim, J.Y.; Pham, D.D. Sasang constitutional medicine as a holistic tailored medicine. Evid.-Based Complement. Altern. Med. 2009, 6, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.; Lee, E.; Kim, C.; Lee, J.; Lixing, L. Sasang constitutional medicine and traditional chinese medicine: A comparative overview. Evid.-Based Complement. Altern. Med. 2011, 2012, 980807. [Google Scholar] [CrossRef] [PubMed]
- Macwan, C.P.; Patel, M.A.; Patel, A.J. Review: Plant sources as potential anti-cancer agents. Int. J. Preclin. Pharm. Res. 2014, 5, 4–11. [Google Scholar]
- Ahmed, H.H.; Abd-Rabou, A.A.; Hassan, A.Z.; Kotob, S.E. Phytochemical analysis and anti-cancer investigation of Bswellia serrata bioactive constituents in vitro. Asian Pac. J. Cancer Prev. 2014, 16, 7179–7188. [Google Scholar] [CrossRef]
- Cha, J.-D.; Jeong, M.-R.; Kim, H.-Y.; Lee, J.-C.; Lee, K.-Y. Mapk activation is necessary to the apoptotic death of kb cells induced by the essential oil isolated from artemisia iwayomogi. J. Ethnopharmacol. 2009, 123, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lv, X.; Shao, L.; Ma, Y.; Cheng, W.; Gao, H. Essential oil from Artemisia lavandulaefolia induces apoptosis and necrosis of HeLa cells. Zhong Yao Cai 2013, 36, 1988–1992. [Google Scholar] [PubMed]
- Son, Y.-O.; Kim, J.; Lim, J.-C.; Chung, Y.; Chung, G.-H.; Lee, J.-C. Ripe fruits of Solanum nigrum L. inhibits cell growth and induces apoptosis in MCF-7 cells. Food Chem. Toxicol. 2003, 41, 1421–1428. [Google Scholar] [CrossRef]
- Keawsa-Ard, S.; Liawruangrath, B.; Liawruangrath, S.; Teerawutgulrag, A.; Pyne, S.G. Essential oil of Solanum spirale fruits and its biological activities. Chiang Mai J. Sci. 2016, 43, 546–554. [Google Scholar]
- Keawsa-Ard, S.; Liawruangrath, B.; Liawruangrath, S.; Teerawutgulrag, A.; Pyne, S.G. Chemical constituents and antioxidant and biological activities of the essential oil from leaves of Solanum spirale. Nat. Prod. Commun. 2012, 7, 955. [Google Scholar] [PubMed]
- Osorio, A.; Silva, T.M.; Duarte, L.P.; Ferraz, V.P.; Pereira, M.T.; Mercadante-Simões, M.O.; Evangelista, F.C.; Sabino, A.P.; Alcântara, A.F. Essential oil from flowers of Solanum stipulaceum: Composition, effects of γ-radiation, and antileukemic activity. J. Braz. Chem. Soc. 2015, 26, 2233–2240. [Google Scholar]
- Essien, E.; Ogunwande, I.; Setzer, W.; Ekundayo, O. Chemical composition, antimicrobial, and cytotoxicity studies on S. erianthum and S. macranthum essential oils. Pharm. Boil. 2012, 50, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hou, T.-T.; Hai-Liang, X.; Qiao-Yan, Z. Estrogen-like activity of volatile components from Vitex rotundifolia L. Indian J. Med. Res. 2007, 126, 68–72. [Google Scholar] [PubMed]
- Jayaprakasha, G.K.; Jena, B.S.; Negi, P.S.; Sakariah, K.K. Evaluation of antioxidant activities and antimutagenicity of turmeric oil: A byproduct from curcumin production. Z. Naturforsch. C Bio. Sci. 2002, 57, 828–835. [Google Scholar] [CrossRef]
- Liju, V.B.; Jeena, K.; Kuttan, R. An evaluation of antioxidant, anti-inflammatory, and antinociceptive activities of essential oil from Curcuma longa. L. Indian J. Pharmacol. 2011, 43, 526–531. [Google Scholar] [PubMed]
- Liju, V.B.; Jeena, K.; Kuttan, R. Chemopreventive activity of turmeric essential oil and possible mechanisms of action. Asian Pac. J. Cancer Prev. 2013, 15, 6575–6580. [Google Scholar] [CrossRef]
- He, J. Clinic application of oil of zedoary in paediatrics. Xiandai Zhong Xi Yi Jie He Za Zhi 2006, 15, 501. [Google Scholar]
- Chen, W.; Lu, Y.; Gao, M.; Wu, J.; Wang, A.; Shi, R. Anti-angiogenesis effect of essential oil from Curcuma zedoaria in vitro and in vivo. J. Ethnopharmacol. 2011, 133, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.Y.; Xu, Q.; Shi, L.C.; Zhang, W.B. Inhibitory effects of curcuma aromatica oil on proliferation of hepatoma in mice. World J. Gastroenterol. 2000, 6, 216–219. [Google Scholar] [PubMed]
- Ding, Y.; Xu, A. Effects of oil of zedoary and its valid component against tumor. Zhong Yao Cai 2005, 28, 152–156. [Google Scholar]
- Shi, H.; Tan, B.; Ji, G.; Lu, L.; Cao, A.; Shi, S.; Xie, J. Zedoary oil (Ezhu You) inhibits proliferation of AGS cells. Chin. Med. 2013, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Perchellet, J.; Perchellet, E.; Abney, N.; Zirnstein, J.; Belman, S. Effects of garlic and onion oils on glutathione peroxidase activity, the ratio of reduced/oxidized glutathione and ornithine decarboxylase induction in isolated mouse epidermal cells treated with tumor promoters. Cancer Biochem. Biophys. 1986, 8, 299–312. [Google Scholar] [PubMed]
- Winking, M.; Sarikaya, S.; Rahmanian, A.; Jödicke, A.; Böker, D.-K. Boswellic acids inhibit glioma growth: A new treatment option? J. Neuro-Oncol. 2000, 46, 97–103. [Google Scholar] [CrossRef]
- Park, K.-R.; Nam, D.; Yun, H.-M.; Lee, S.-G.; Jang, H.-J.; Sethi, G.; Cho, S.K.; Ahn, K.S. B-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/Akt/mTOR/S6K1 pathways and ROS-mediated mapks activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Ho, C.-T.; Chin, C.-K.; Badmaev, V.; Ma, W.; Huang, M.-T. Inhibitory activity of boswellic acids from Boswellia serrata against human leukemia HL-60 cells in culture. Planta Med. 1998, 64, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.-N.; Hong, S.-H.; Jeong, H.-J.; Lee, E.-H.; Kim, N.-G.; Choi, S.-D.; Ra, K.-W.; Kim, K.-S.; Kang, B.-K.; Kim, J.-J. Inhibitory effect of Artemisia capillaris on ethanol-induced cytokines (TNF-α, IL-1α) secretion in hep G2 cells. Immunopharmacol. Immunotoxicol. 2002, 24, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.-D.; Jeong, M.-R.; Jeong, S.-I.; Moon, S.-E.; Kim, J.-Y.; Kil, B.-S.; Song, Y.-H. Chemical composition and antimicrobial activity of the essential oils of Artemisia scoparia and A. capillaris. Planta Med. 2005, 71, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-H.; Kim, Y.-H.; Kil, B.-S.; Kim, K.-J.; Jeong, S.-I.; You, Y.-O. Chemical composition and antibacterial activity of essential oil of Artemisia iwayomogi. Planta Med. 2003, 69, 1159–1162. [Google Scholar] [PubMed]
- Cha, J.-D.; Jeong, M.-R.; Choi, H.-J.; Jeong, S.-I.; Moon, S.-E.; Yun, S.-I.; Kim, Y.-H.; Kil, B.-S.; Song, Y.-H. Chemical composition and antimicrobial activity of the essential oil of Artemisia lavandulaefolia. Planta Med. 2005, 71, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, H.J.; Jeong, S.J.; Lee, M.H.; Kim, S.H. Essential oil of Pinus koraiensis leaves exerts antihyperlipidemic effects via up-regulation of low-density lipoprotein receptor and inhibition of acyl-coenzyme A: Cholesterol acyltransferase. Phytother. Res. 2012, 26, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Sung, B.; Yadav, V.R.; Cho, S.G.; Liu, M.; Aggarwal, B.B. Acetyl-11-keto-β-boswellic acid suppresses invasion of pancreatic cancer cells through the downregulation of CXCR4 chemokine receptor expression. Int. J. Cancer 2011, 129, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R.; Statti, G.A.; Menichini, F. Jacaranone: A cytotoxic constituent fromsenecio ambiguus subsp. Ambiguus (Biv.) DC. against renal adenocarcinoma achn and prostate carcinoma LNCaP cells. Arch. Pharm. Res. 2007, 30, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.-H.; Ko, E.; Oh, B.-G.; Kim, S.-H.; Kim, Y.; Shin, M.; Hong, M.; Bae, H. Inhibition effects of Vitex rotundifolia on inflammatory gene expression in A549 human epithelial cells. Ann. Allergy Asthma Immunol. 2009, 103, 152–159. [Google Scholar] [CrossRef]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and p388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Baik, K.-U.; Jung, S.-H.; Ahn, B.-Z. Recognition of pharmacophore of ar-turmerone for its anticancer activity. Arch. Pharm. Res. 1993, 16, 254–256. [Google Scholar] [CrossRef]
- Li, Y.; Wo, J.M.; Liu, Q.; Li, X.; Martin, R.C. Chemoprotective effects of Curcuma aromatica on esophageal carcinogenesis. Ann. Surg. Oncol. 2009, 16, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yang, F.-Q.; Li, S.-P.; Hu, G.; Lee, S.; Wang, Y.-T. Essential oil of Curcuma wenyujin induces apoptosis in human hepatoma cells. World J. Gastroenterol. 2008, 14, 4309–4318. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-S.; Li, T.; Wu, G.-S.; Dang, Y.-Y.; Hao, W.-H.; Chen, X.-P.; Lu, J.-J.; Wang, Y.-T. Effects of furanodiene on 95-d lung cancer cells: Apoptosis, autophagy and G1 phase cell cycle arrest. Am. J. Chin. Med. 2014, 42, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.; Wang, X.; Li, Y.; Sun, X.; Tai, W.; Li, T.; Guo, T. Induction of apoptosis by furanodiene in HL60 leukemia cells through activation of TNFR1 signaling pathway. Cancer Lett. 2008, 271, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Bachir, R.G.; Benali, M. Antibacterial activity of the essential oils from the leaves of Eucalyptus globulus against Escherichia coli and Staphylococcus aureus. Asian Pac. J. Trop. Biomed. 2012, 2, 739–742. [Google Scholar] [CrossRef]
- Döll-Boscardin, P.M.; Sartoratto, A.; Sales Maia, B.H.L.D.N.; Padilha de Paula, J.; Nakashima, T.; Farago, P.V.; Kanunfre, C.C. In vitro cytotoxic potential of essential oils of Eucalyptus benthamii and its related terpenes on tumor cell lines. Evid.-Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Elaissi, A.; Rouis, Z.; Salem, N.A.B.; Mabrouk, S.; Ben Salem, Y.; Salah, K.B.H.; Aouni, M.; Farhat, F.; Chemli, R.; Harzallah-Skhiri, F. Chemical composition of 8 Eucalyptus species’ essential oils and the evaluation of their antibacterial, antifungal and antiviral activities. BMC Complement. Altern. Med. 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- European Pharmacopeia Commision. European Pharmacopeia, 4th ed.; Council of Europe: Strasbourg-Cedex, France, 2002; Volume 2.8.12, pp. 183–184. [Google Scholar]
- Bugarin, D.; Grbović, S.; Orčič, D.; Mitić-Ćulafić, D.; Knežević-Vukčević, J.; Mimica-Dukić, N. Essential oil of Eucalyptus gunnii Hook. as a novel source of antioxidant, antimutagenic and antibacterial agents. Molecules 2014, 19, 19007–19020. [Google Scholar] [CrossRef] [PubMed]
- Sebei, K.; Sakouhi, F.; Herchi, W.; Khouja, M.L.; Boukhchina, S. Chemical composition and antibacterial activities of seven Eucalyptus species essential oils leaves. Biol. Res. 2015, 48, 7. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, A.; Yitayew, B.; Tesema, A.; Taddese, S. In vitro antimicrobial activity of essential oil of Thymus schimperi, Matricaria chamomilla, Eucalyptus globulus, and Rosmarinus officinalis. Int. J. Microbial. 2016, 2016, 9545693. [Google Scholar]
- Baptista, E.B.; Zimmermann-Franco, D.C.; Lataliza, A.A.B.; Raposo, N.R.B. Chemical composition and antifungal activity of essential oil from Eucalyptus smithii against dermatophytes. Rev. Soc. Bras. Med. Trop. 2015, 48, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Ho, Y.-C.; Lim, J.-M.; Chang, T.-Y.; Ho, C.-L.; Chang, T.-M. Investigation of the anti-melanogenic and antioxidant characteristics of Eucalyptus camaldulensis flower essential oil and determination of its chemical composition. Int. J. Mol. Sci. 2015, 16, 10470–10490. [Google Scholar] [CrossRef] [PubMed]
- Gbenou, J.D.; Ahounou, J.F.; Akakpo, H.B.; Laleye, A.; Yayi, E.; Gbaguidi, F.; Baba-Moussa, L.; Darboux, R.; Dansou, P.; Moudachirou, M. Phytochemical composition of Cymbopogon citratus and Eucalyptus citriodora essential oils and their anti-inflammatory and analgesic properties on wistar rats. Mol. Boil. Rep. 2013, 40, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Saharkhiz, M.J.; Motamedi, M.; Zomorodian, K.; Pakshir, K.; Miri, R.; Hemyari, K. Chemical composition, antifungal and antibiofilm activities of the essential oil of Mentha piperita L. ISRN Pharm. 2012, 2012, 718645. [Google Scholar] [PubMed]
- Sun, Z.; Wang, H.; Wang, J.; Zhou, L.; Yang, P. Chemical composition and anti-inflammatory, cytotoxic and antioxidant activities of essential oil from leaves of Mentha piperita grown in China. PLoS ONE 2014, 9, e114767. [Google Scholar] [CrossRef] [PubMed]
- Furneri, P.M.; Mondello, L.; Mandalari, G.; Paolino, D.; Dugo, P.; Garozzo, A.; Bisignano, G. In vitro antimycoplasmal activity of Citrus bergamia essential oil and its major components. Eur. J. Med. Chem. 2012, 52, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Adukwu, E.; Allen, S.C.; Phillips, C.A. The anti-biofilm activity of lemongrass (Cymbopogon flexuosus) and grapefruit (Citrus paradisi) essential oils against five strains of Staphylococcus aureus. J. Appl. Microbial. 2012, 113, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Ellouze, I.; Abderrabba, M.; Sabaou, N.; Mathieu, F.; Lebrihi, A.; Bouajila, J. Season’s variation impact on Citrus aurantium leaves essential oil: Chemical composition and biological activities. J. Food Sci. 2012, 77, T173–T180. [Google Scholar] [CrossRef] [PubMed]
- Hsouna, A.B.; Hamdi, N.; Halima, N.B.; Abdelkafi, S. Characterization of essential oil from Citrus aurantium L. Flowers: Antimicrobial and antioxidant activities. J. Oleo Sci. 2013, 62, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.H.; Bouajila, J.; Lebrihi, A.; Mathieu, F.; Romdhane, M.; Zagrouba, F. Chemical composition and in vitro antimicrobial and antioxidant activities of Citrus aurantium L. flowers essential oil (Neroli oil). Pak. J. Biol. Sci. 2012, 15, 1034. [Google Scholar] [CrossRef] [PubMed]
- Metoui, N.; Gargouri, S.; Amri, I.; Fezzani, T.; Jamoussi, B.; Hamrouni, L. Activity antifungal of the essential oils; aqueous and ethanol extracts from Citrus aurantium L. Nat. Prod. Res. 2015, 29, 2238–2241. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, D.; Ashour, M.L.; Mulyaningsih, S.; El-Shazly, A.; Wink, M. Chemical composition of the essential oils of variegated pink-fleshed lemon (Citrus x Limon L. Burm. F.) and their anti-inflammatory and antimicrobial activities. Zeitschrift Naturforschung C 2013, 68, 275–284. [Google Scholar] [CrossRef]
- Settanni, L.; Randazzo, W.; Palazzolo, E.; Moschetti, M.; Aleo, A.; Guarrasi, V.; Mammina, C.; San Biagio, P.; Marra, F.; Moschetti, G. Seasonal variations of antimicrobial activity and chemical composition of essential oils extracted from three Citrus limon L. Burm. Cultivars. Nat. Prod. Res. 2014, 28, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Mitoshi, M.; Kuriyama, I.; Nakayama, H.; Miyazato, H.; Sugimoto, K.; Kobayashi, Y.; Jippo, T.; Kuramochi, K.; Yoshida, H.; Mizushina, Y. Suppression of allergic and inflammatory responses by essential oils derived from herbal plants and citrus fruits. Int. J. Mol. Med. 2014, 33, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Polo, C.; Moraes, T.; Pellizzon, C.; Marques, M.; Rocha, L.; Hiruma-Lima, C.A. Gastric ulcers in middle-aged rats: The healing effect of essential oil from Citrus aurantium L. (Rutaceae). Evid.-Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Bonamin, F.; Moraes, T.M.; dos Santos, R.C.; Kushima, H.; Faria, F.M.; Silva, M.A.; Junior, I.V.; Nogueira, L.; Bauab, T.M.; Brito, A.R.S. The effect of a minor constituent of essential oil from Citrus aurantium: The role of β-myrcene in preventing peptic ulcer disease. Chem. Boil. Interact. 2014, 212, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, M.; Luini, A.; Bombelli, R.; Corasaniti, M.T.; Bagetta, G.; Marino, F. The essential oil of bergamot stimulates reactive oxygen species production in human polymorphonuclear leukocytes. Phytother. Res. 2014, 28, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Tundis, R.; Bonesi, M.; Sanzo, G.D.; Verardi, A.; Lopresto, C.G.; Pugliese, A.; Menichini, F.; Balducchi, R.; Calabrò, V. Chemical profile and antioxidant properties of extracts and essential oils from Citrus × limon (L.) burm. Cv. Femminello comune. Chem. Biodivers. 2016, 13, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Celia, C.; Trapasso, E.; Locatelli, M.; Navarra, M.; Ventura, C.A.; Wolfram, J.; Carafa, M.; Morittu, V.M.; Britti, D.; Di Marzio, L. Anticancer activity of liposomal bergamot essential oil (BEO) on human neuroblastoma cells. Colloids Surf. B Biointerfaces 2013, 112, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Ciociaro, A.; Berliocchi, L.; Cassiano, M.G.V.; Rombolà, L.; Ragusa, S.; Bagetta, G.; Blandini, F.; Corasaniti, M.T. Implication of limonene and linalyl acetate in cytotoxicity induced by bergamot essential oil in human neuroblastoma cells. Fitoterapia 2013, 89, 48–57. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Institutes of Health. ClinicalTrials.gov. Available online: http://clinicaltrial.gov/ (accessed on 23 June 2016).
- Azzimonti, B.; Cochis, A.; Beyrouthy, M.E.; Iriti, M.; Uberti, F.; Sorrentino, R.; Landini, M.M.; Rimondini, L.; Varoni, E.M. Essential oil from berries of lebanese Juniperus excelsa M. Bieb displays similar antibacterial activity to chlorhexidine but higher cytocompatibility with human oral primary cells. Molecules 2015, 20, 9344–9357. [Google Scholar] [CrossRef] [PubMed]
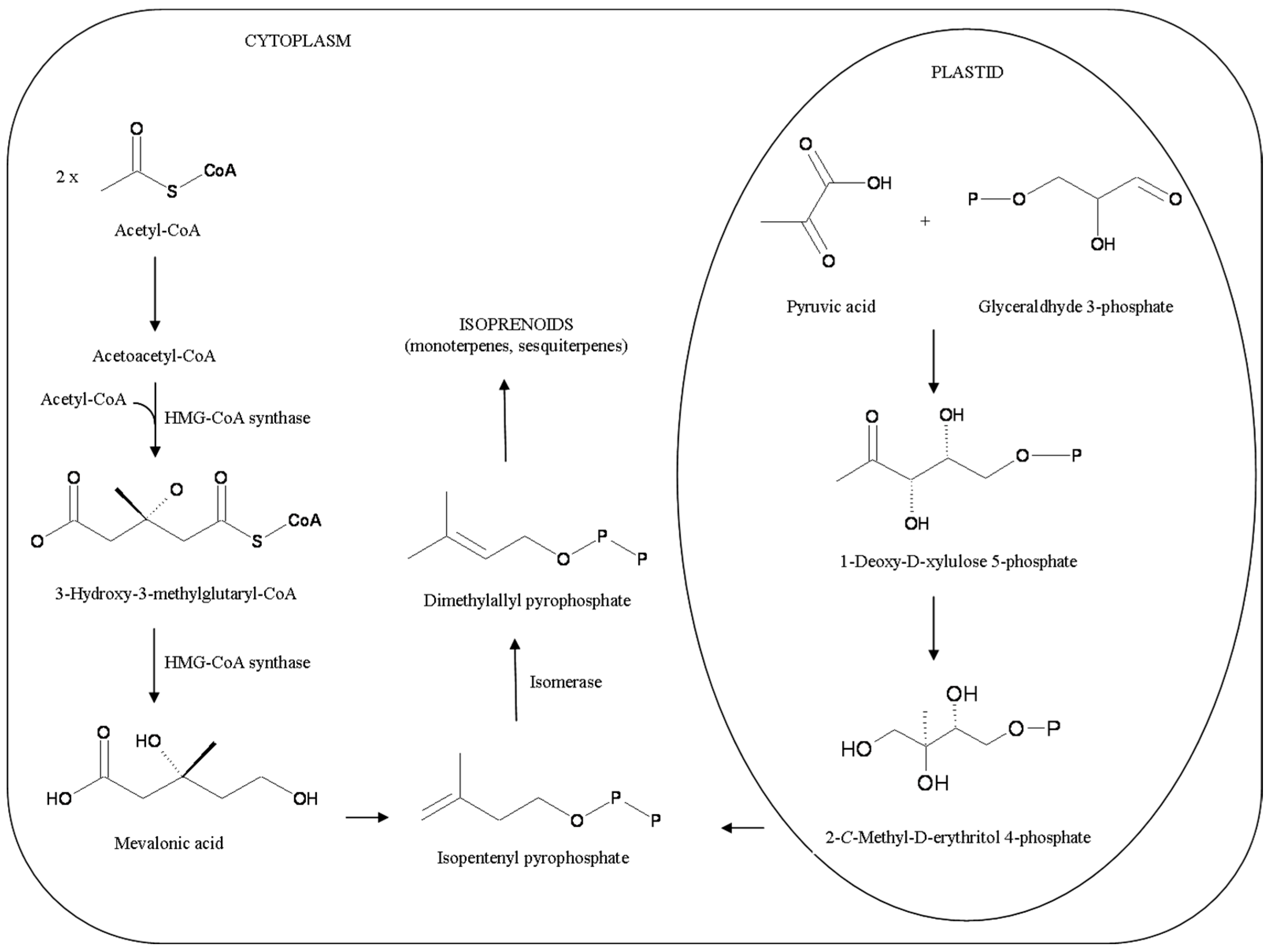

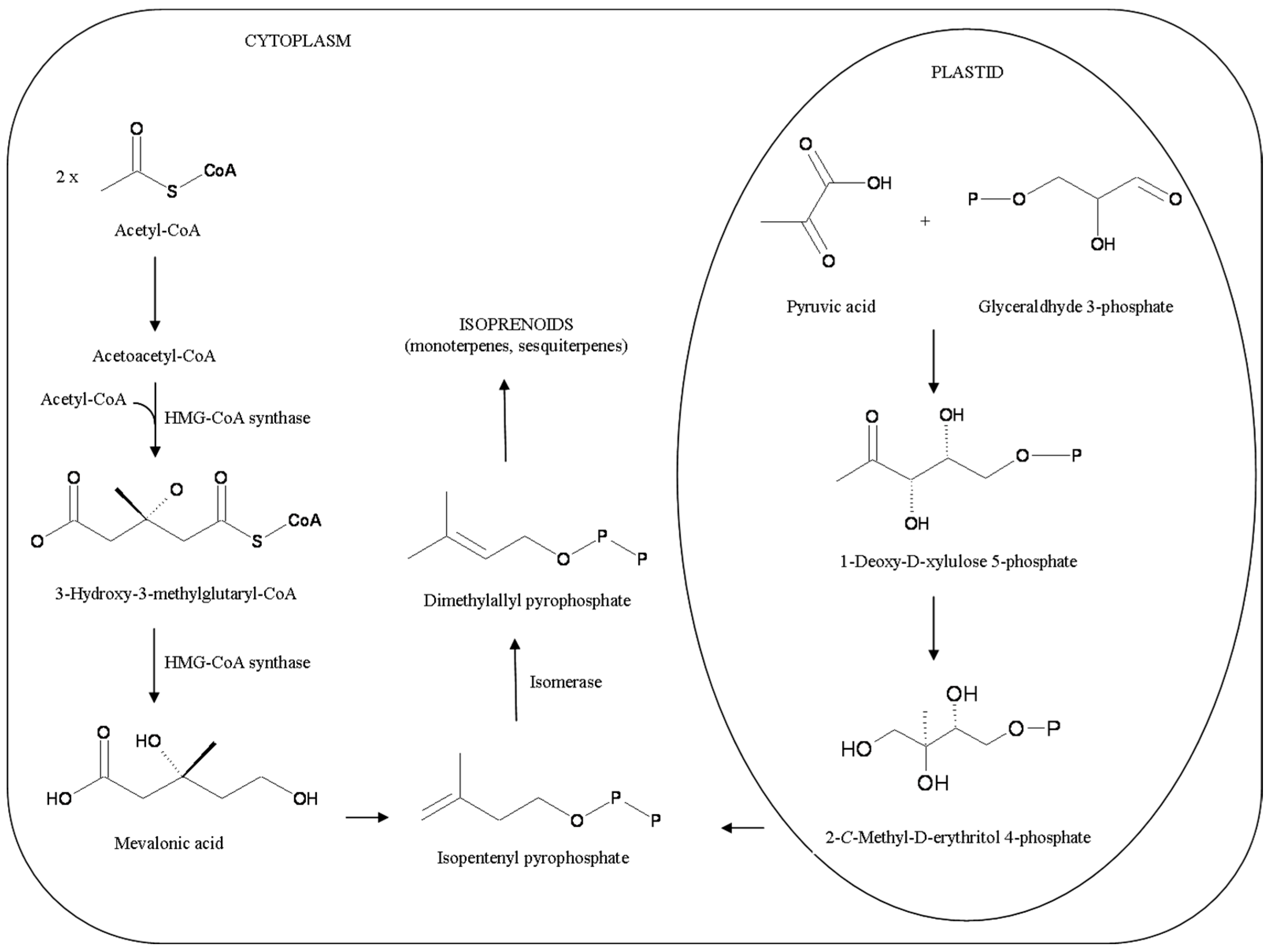{kind=link}
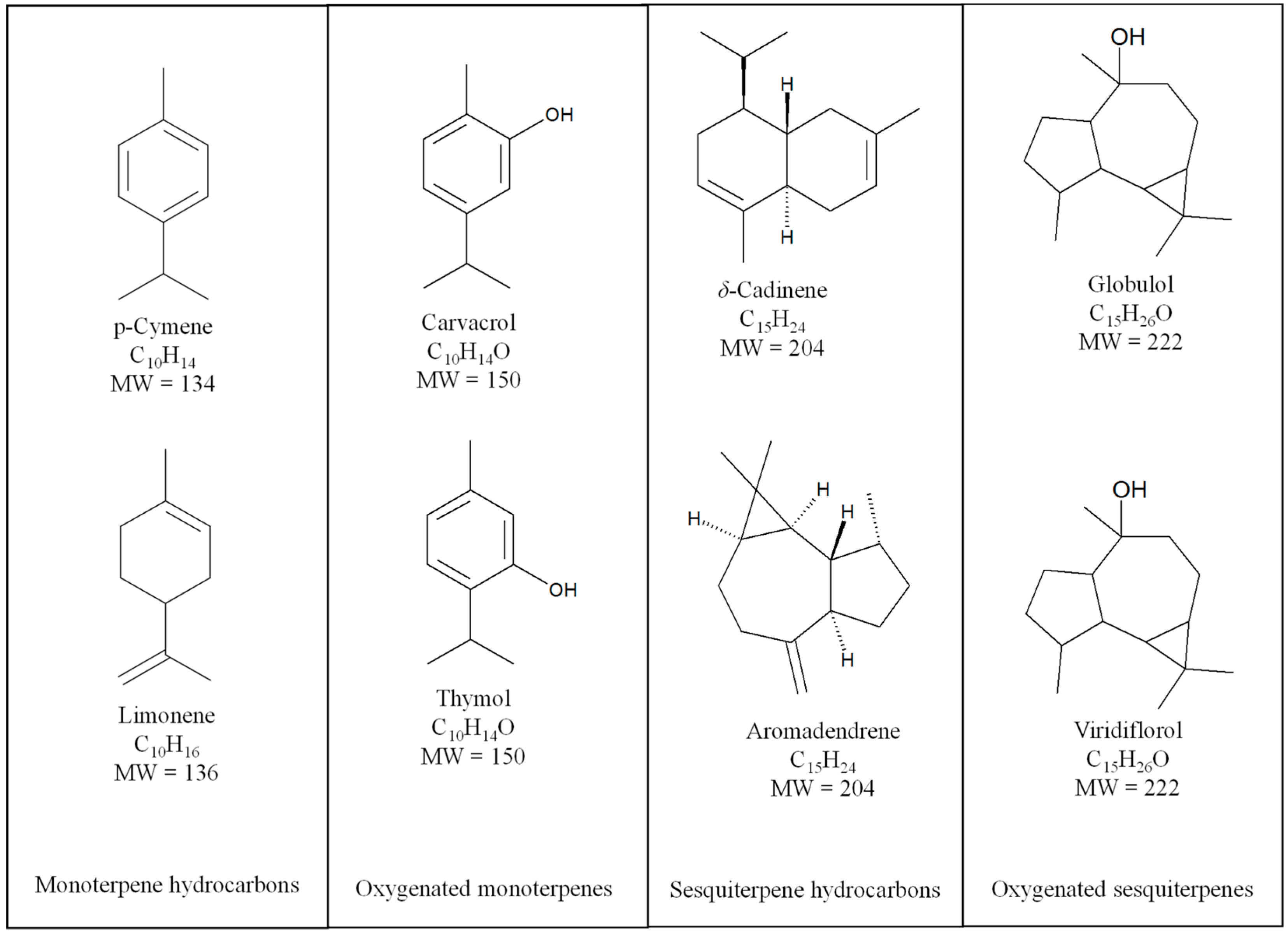
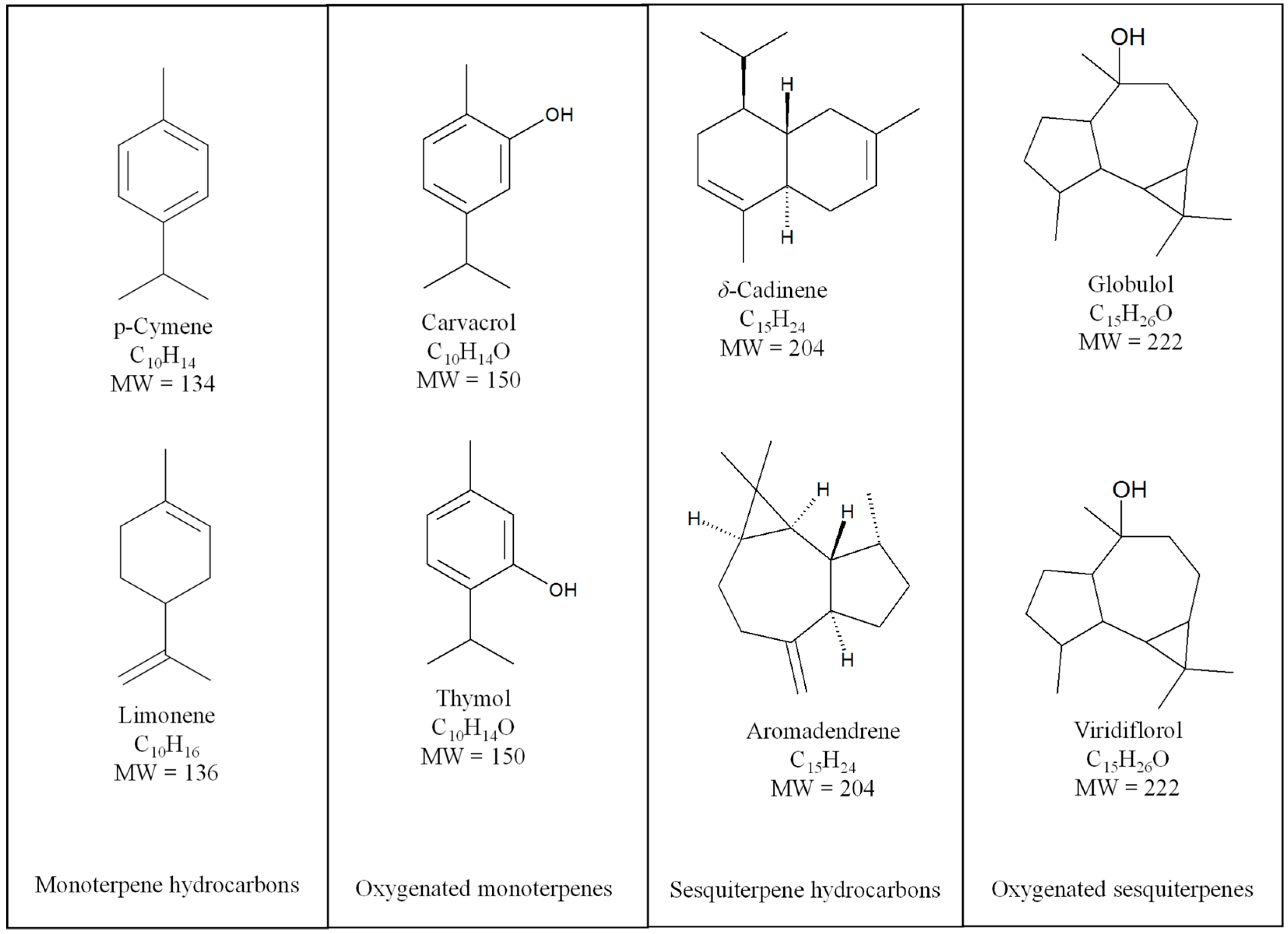{kind=link}
| Family | Scientific Name | Tumor (Cancer Cell Lines) | References |
|---|---|---|---|
| Amaryllidaceae | Allium sativum L. | Human breast, colon, lung, skin and liver cancers | [246] |
| Burseraceae | Boswellia serrata Roxb. ex Colebr. | Human colon carcinoma (HCT116) | [247] |
| Compositae | Artemisia capillaris Thunb. | Human oral epidermal carcinoma (KB) | [95] |
| Artemisia iwayomogi Kitam. | Human oral epidermal carcinoma (KB) | [248] | |
| Artemisia lavandulaefolia Nakai | Human cervix adenocarcinoma (HeLa) | [249] | |
| Pinaceae | Pinus koraiensis Siebold & Zucc. | Human colon carcinoma (HCT116) | [93] |
| Solanaceae | Solanum nigrum L. | Liver, mammary, uterine cervix and gastric cancers | [250] |
| Solanum spirale Roxb. | Human breast (MCF-7) and small cell lung (NCI-H187) cancers, human oral epidermal carcinoma (KB) | [251,252] | |
| Solanum stipulaceum Roem. & Schult | Human acute promyelocytic leukemia (HL-60) and acute monocytic leukemia (THP-1) | [253] | |
| Solanum erianthum D. Don | Human breast carcinoma (Hs 578T) and prostate carcinoma (PC-3) | [254] | |
| Solanum macranthum Dunal | Human breast carcinoma (Hs 578T) and prostate carcinoma (PC-3) | [254] | |
| Verbenaceae | Vitex rotundifolia L.f. | Breast cancer (MCF-7) | [255] |
| Zingiberaceae | Curcuma longa L. | Human oral epidermal carcinoma (KB), murine leukemia (P388), leukemia (HL-60, K-562 and L1210) | [256,257,258,259] |
| Curcuma zedoaria Roxb. | Mouse melanoma (B16BL6), human hepatoma (SMMC-7721), non-small cell lung carcinoma (NSCLC) | [91,260] | |
| Curcuma wenyujin Y.H.Chen & C.Ling | Cervical, liver (HepG2), lung and gastric (AGS and MGC 803) cancers | [261,262,263] |
| Essential Oil Name | Botanical Name | Plant Part Used | European Pharmacopoeias | American Pharmacopoeias |
|---|---|---|---|---|
| Anise oil | Pimpinella anisum L. | ripe fruit | X | X |
| Bitter-fennel fruit oil | Foeniculum vulgare Mill., spp. vulgare var. vulgare | ripe fruit | X | X |
| Bitter-fennel herb oil | Foeniculum vulgare Mill., spp. vulgare var. vulgare | aerial parts | X | |
| Caraway oil | Carum carvi L. | dry fruit | X | X |
| Cardamom oil | Elettaria cardamomum L. | seeds | X | |
| Cassia oil | Cinnamomum aromaticum Nees | leave and young branches | X | |
| Cinnamon bark oil, Ceylon | Cinnamomum zeylanicum Nees | bark of the shoot | X | |
| Cinnamon leaf oil, Ceylon | Cinnamomum zeylanicum Nees | leaves | X | |
| Citronella oil | Cymbopogon winterianus Jowitt ex Bor | fresh or partially dried aerial parts | X | |
| Clary sage oil | Salvia sclarea L. | fresh or dried flowering stem | X | |
| Clove oil | Syzygium aromaticum (L.) Merr. & L.M.Perry | dried flower buds | X | X |
| Coriander oil | Coriandrum sativum L. | fruits | X | X |
| Dwarf pine oil | Pinus mugo Turra | fresh leaves and twigs | X | |
| Eucalyptus oil | genus Eucalyptus | fresh leaves or fresh terminal branches of various species of Eucalyptus rich in 1,8-cineole | X | |
| Juniper oil | Juniperus communis L. | ripe, non-fermented berry cones | X | |
| Lavender oil | Lavandula angustifolia Mill. | flowering tops | X | |
| Lemon oil | Citrus limon L. | fresh peel | X | X |
| Mandarin oil | Citrus reticulata Blanco | fresh peel | X | |
| Matricaria oil | Matricaria recutita L. | fresh or dried flower-heads or flowering tops | X | |
| Mint oil (partly dementholised by cristallization) | Mentha canadensis L. | fresh, flowering aerial parts | X | |
| Neroli oil | Citrus aurantium L. | fresh flowers | X | |
| Nutmeg oil | Myristica fragrans Houtt. | dried and crushed kernels | X | |
| Peppermint oil | Mentha piperita L. | fresh aerial parts of the flowering plant | X | X |
| Pine sylvestris oil | Pinus sylvestris L. | fresh leaves and branches | X | |
| Rose oil | Rosa gallica L., Rosa damascene Miller, Rosa alba L., Rosa centifolia L. | fresh flowers | X | |
| Rosemary oil | Rosmarinus officinalis L. | flowering aerial parts | X | |
| Spanish sage oil | Salvia lavandulifolia Vahl | flowering aerial parts | X | |
| Spike lavender oil | Lavandula latifolia Medik | flowering tops | X | |
| Star anise oil | Illicium verum Hook | dry ripe fruits | X | |
| Sweet orange oil | Citrus sinensis (L.) Osbeck | fresh peel | X | X |
| Tea tree oil | genus Melaleuca | foliage and terminal branches | X | |
| Thyme oil | Thymus vulgaris L. or Thymus zygis L. | fresh flowering aerial parts | X | |
| Turpentine oil, Pinus pinaster type | Pinus pinaster Aiton | oleoresin obtained by tapping | X |
| “Eucalyptus Essential Oil” Clinical Trials | |
|---|---|
| Study Title | Study to Investigate Effects of Different Therapies for the Treatment of Hypertension |
| Condition | Hypertension |
| Status | Completed |
| Start and Completion dates | March 2014–July 2015 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Crossover Assignment | |
| Masking: Open Label | |
| Primary Purpose: Treatment | |
| Primary Outcome | Blood pressure |
| Condition | Hypertension |
| Status | Completed |
| Start and Completion dates | August 2013–April 2014 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Intervention Model: Crossover Assignment | |
| Masking: Single Blind (Subject) | |
| Primary Outcome | Blood pressure response to Inhalation of Essential Eucalyptus Oil |
| Study Title | Treatment of Acute Rhino-Sinusitis With Essential Oils of Aromatic Plants |
| Condition | Rhino-Sinusitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a relief in the nasal obstruction within the first 20 min after first administration of treatment with the spray |
| Study Title | Treatment of Acute Pharyngo-Tonsillitis with Essential Oils of Aromatic Plants |
| Condition | Viral Pharyngitis—Viral Tonsillitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a throat pain relief within the first 20 min after first administration of treatment with the spray |
| Study Title | Treatment of Acute Tracheitis and Laryngitis with Essential Oils of Aromatic Plants |
| Condition | Viral Laryngitis—Viral Tracheitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a cough or hoarseness relief within the first 20 min after first administration of treatment with the spray |
| Study Title | Study on Hypertonic Saline Nasal Spray (PhytosunDecon) |
| Condition | Nasal Congestion |
| Status | Completed |
| Start and Completion dates | March 2014–October 2014 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Pharmacokinetics Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Caregiver) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Assessment of the speed of relief nasal congestion |
| “Mentha Essential Oil” Clinical Trials | |
|---|---|
| Study Title | Study of an Essential Oil and a Delmopinol Mouthrinse Effect on Dental Plaque Accumulation Index, Gingivitis Index and on Streptococcus Mutans, Lactobacillus, Aerobic and Anaerobic Oral Bacteria Colony Counts |
| Condition | Gingivitis—Dental Plaque Accumulation |
| Status | Completed |
| Start and Completion dates | April 2007–April 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Open Label | |
| Primary Purpose: Prevention | |
| Primary Outcome | Loe & Silness Gingival Index (1963) |
| Quigley, Hein & Turesky Dental Plaque Index (1970) | |
| Study Title | Essential Oils Mouthrinse and Dental Floss, Comparison of Efficacy on Interproximal Gingivitis and Dental Plaque Accumulation |
| Condition | Gingivitis Dental—Plaque Accumulation |
| Status | Completed |
| Start and Completion dates | September 2007–January 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Open Label | |
| Primary Purpose: Prevention | |
| Primary Outcome | Lobene Gingival Index |
| Saxton & Ouderaa Bleeding Index | |
| Quigley, Hein & Turesky Dental Plaque Index | |
| Study Title | Treatment of Acute Rhinosinusitis with Essential Oils of Aromatic Plants |
| Condition | Rhinosinusitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a relief in the nasal obstruction within the first 20 min after first administration of treatment with the spray |
| Study Title | Treatment of Acute Pharyngo-Tonsillitis with Essential Oils of Aromatic Plants |
| Condition | Viral Pharyngitis—Viral Tonsillitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a throat pain relief within the first 20 min after first administration of treatment with the spray. |
| Study Title | Treatment of Acute Tracheitis and Laryngitis with Essential Oils of Aromatic Plants |
| Condition | Viral Laryngitis—Viral Tracheitis |
| Status | Completed |
| Start and Completion dates | January 2008–May 2008 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | To demonstrate a cough or hoarseness relief within the first 20 min after first administration of treatment with the spray |
| Study Title | Dental Implants and Mouth Rinse |
| Condition | Reduction in Bacterial Counts Through the Use of Mouthwash |
| Status | Recruiting |
| Start and Completion dates | November 2013–June 2016 (hypothesized) |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Open Label | |
| Primary Purpose: Supportive Care | |
| Primary Outcome | Oral Rinse Comparison |
| Study Title | A Clinical Trial to Test the Effect of Marketed Mouth Rinses on Decreasing Plaque and Gum Inflammation |
| Condition | Plaque—Gingivitis |
| Status | Completed |
| Start and Completion dates | August 2014–September 2014 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Safety/Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Single Blind (Investigator) | |
| Primary Outcome | Whole Mouth Mean Modified Gingival Index (MGI) at Week 3 |
| Whole Mouth Mean Plaque Index (PI) at Week 3 | |
| Study Title | Freeze Remedy to Alleviate Morning Sickness |
| Condition | Morning Sickness |
| Status | Recruiting |
| Start and Completion dates | August 2014–February 2015 (hypothesized) |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Caregiver) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Improvement of Nausea and Vomiting of Pregnancy (NVP) |
| “Citrus Essential Oil” Clinical Trials | |
|---|---|
| Study Title | The Effects of Smell on Mood and Physical Responses |
| Condition | Stress—Anxiety—Depression |
| Status | Completed |
| Start and Completion dates | August 2015–March 2016 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Safety/Efficacy Study | |
| Intervention Model: Crossover Assignment | |
| Masking: Double Blind (Subject, Investigator) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Cortisol and Catecholamine Production; Immune Function; Skin Barrier Repair; Delayed Hypersensitivity to Candida (DTH) |
| Study Title | Impact of AV2 Antiviral Drug on the Treatment of HPV-associated Lesions of the Uterine Cervix (KINVAV) |
| Condition | Uterine Cervical Dysplasia—Papillomavirus Infections |
| Status | Recruiting |
| Start and Completion dates | January 2015–June 2016 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Single Group Assignment | |
| Masking: Double Blind (Subject, Caregiver, Investigator, Outcomes Assessor) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Change of lesions (lesion size) |
| Study Title | Aromatherapy for Chemotherapy-induced Symptoms |
| Condition | Chemotherapy Symptoms |
| Status | Recruiting |
| Start and Completion dates | January 2016–July 2017 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Outcomes Assessor) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Mean change in severity of symptoms |
| Study Title | Limonene Study in Women with Breast Cancer |
| Condition | Breast Cancer |
| Status | Completed |
| Start and Completion dates | August 2009–March 2011 |
| Study Type and Design | Interventional |
| Endpoint Classification: Pharmacokinetics/Dynamics Study | |
| Intervention Model: Single Group Assignment | |
| Masking: Open Label | |
| Primary Purpose: Prevention | |
| Primary Outcome | Breast tissue limonene level |
| Study Title | Does Massage with or without Aromatherapy Reduce Infant’s Distress? (aromatherapy) |
| Condition | Distress |
| Status | Unknown |
| Start and Completion dates | January 2008–December 2009 |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Factorial Assignment | |
| Masking: Single Blind (Outcomes Assessor) | |
| Primary Purpose: Supportive Care | |
| Primary Outcome | Level of COMFORT with videotaped COMFORT behavior scale |
| Study Title | Freeze Remedy to Alleviate Morning Sickness |
| Condition | Morning Sickness |
| Status | Recruiting |
| Start and Completion dates | August 2012–February 2015 (hypothesized) |
| Study Type and Design | Interventional |
| Allocation: Randomized | |
| Endpoint Classification: Efficacy Study | |
| Intervention Model: Parallel Assignment | |
| Masking: Double Blind (Subject, Caregiver) | |
| Primary Purpose: Treatment | |
| Primary Outcome | Improvement of Nausea and Vomiting in Pregnancy |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 2017, 22, 70. https://doi.org/10.3390/molecules22010070
Sharifi-Rad J, Sureda A, Tenore GC, Daglia M, Sharifi-Rad M, Valussi M, Tundis R, Sharifi-Rad M, Loizzo MR, Ademiluyi AO, et al. Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules. 2017; 22(1):70. https://doi.org/10.3390/molecules22010070
Chicago/Turabian StyleSharifi-Rad, Javad, Antoni Sureda, Gian Carlo Tenore, Maria Daglia, Mehdi Sharifi-Rad, Marco Valussi, Rosa Tundis, Marzieh Sharifi-Rad, Monica R. Loizzo, Adedayo Oluwaseun Ademiluyi, and et al. 2017. "Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems" Molecules 22, no. 1: 70. https://doi.org/10.3390/molecules22010070
APA StyleSharifi-Rad, J., Sureda, A., Tenore, G. C., Daglia, M., Sharifi-Rad, M., Valussi, M., Tundis, R., Sharifi-Rad, M., Loizzo, M. R., Ademiluyi, A. O., Sharifi-Rad, R., Ayatollahi, S. A., & Iriti, M. (2017). Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules, 22(1), 70. https://doi.org/10.3390/molecules22010070