
Inhibitory Effects of Viscum coloratum Extract on IgE/Antigen-Activated Mast Cells and Mast Cell-Derived Inflammatory Mediator-Activated Chondrocytes


Abstract
:
1. Introduction
2. Results
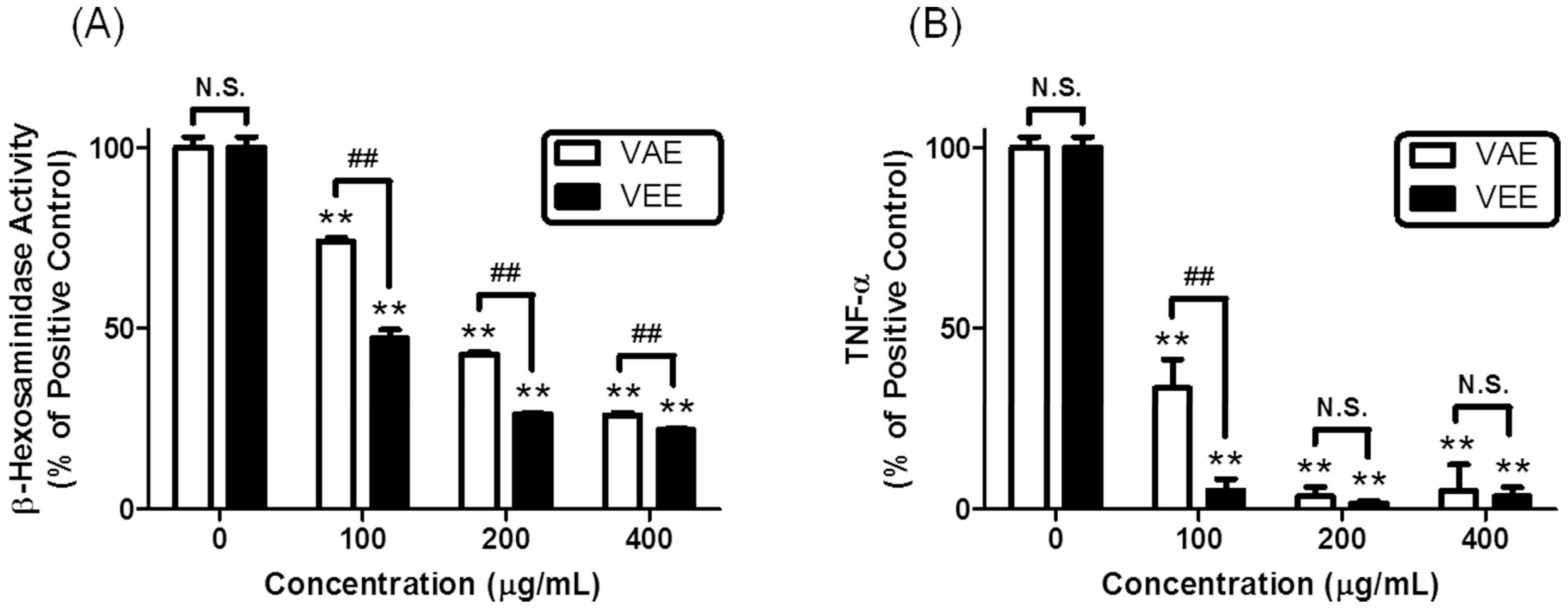
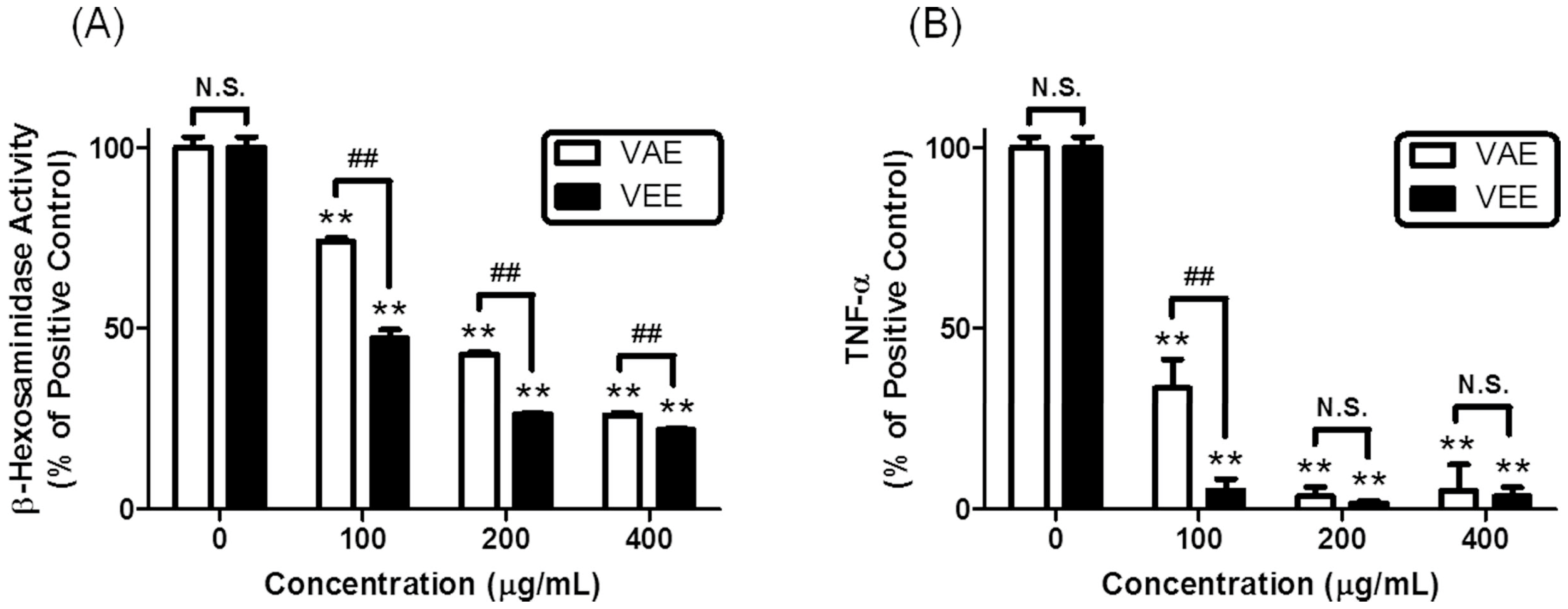
2.1. Inhibitory Effects of VAE or VEE on the IgE/Ag-Mediated Allergic Response in RBL-2H3 Cells
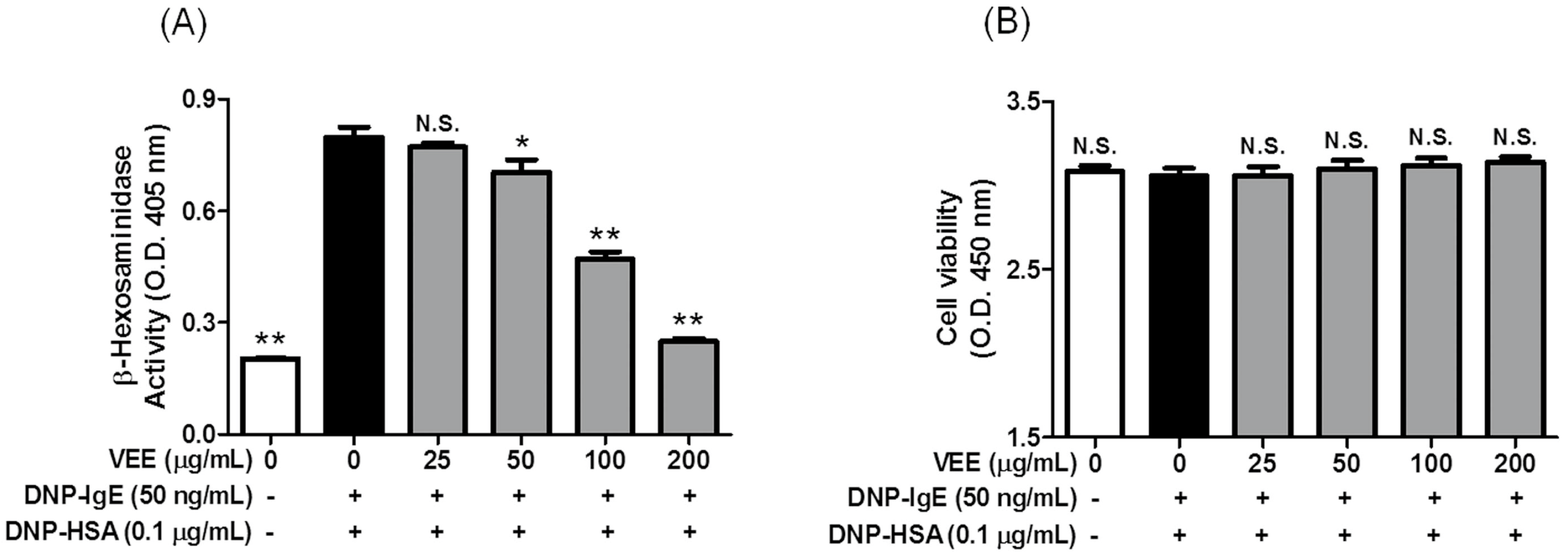
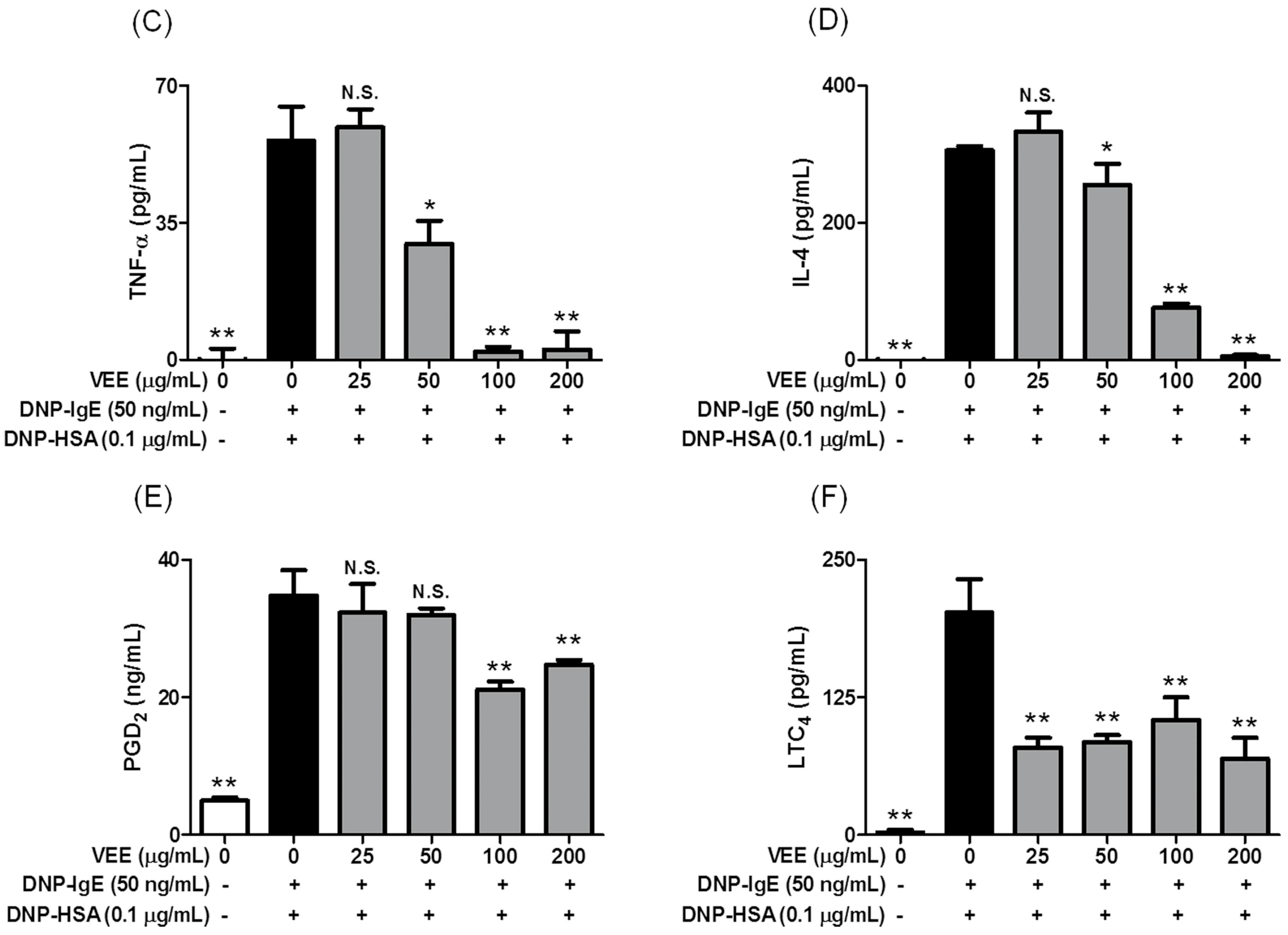
2.2. Inhibitory Effects of VEE on the Formation of Inflammatory Mediators
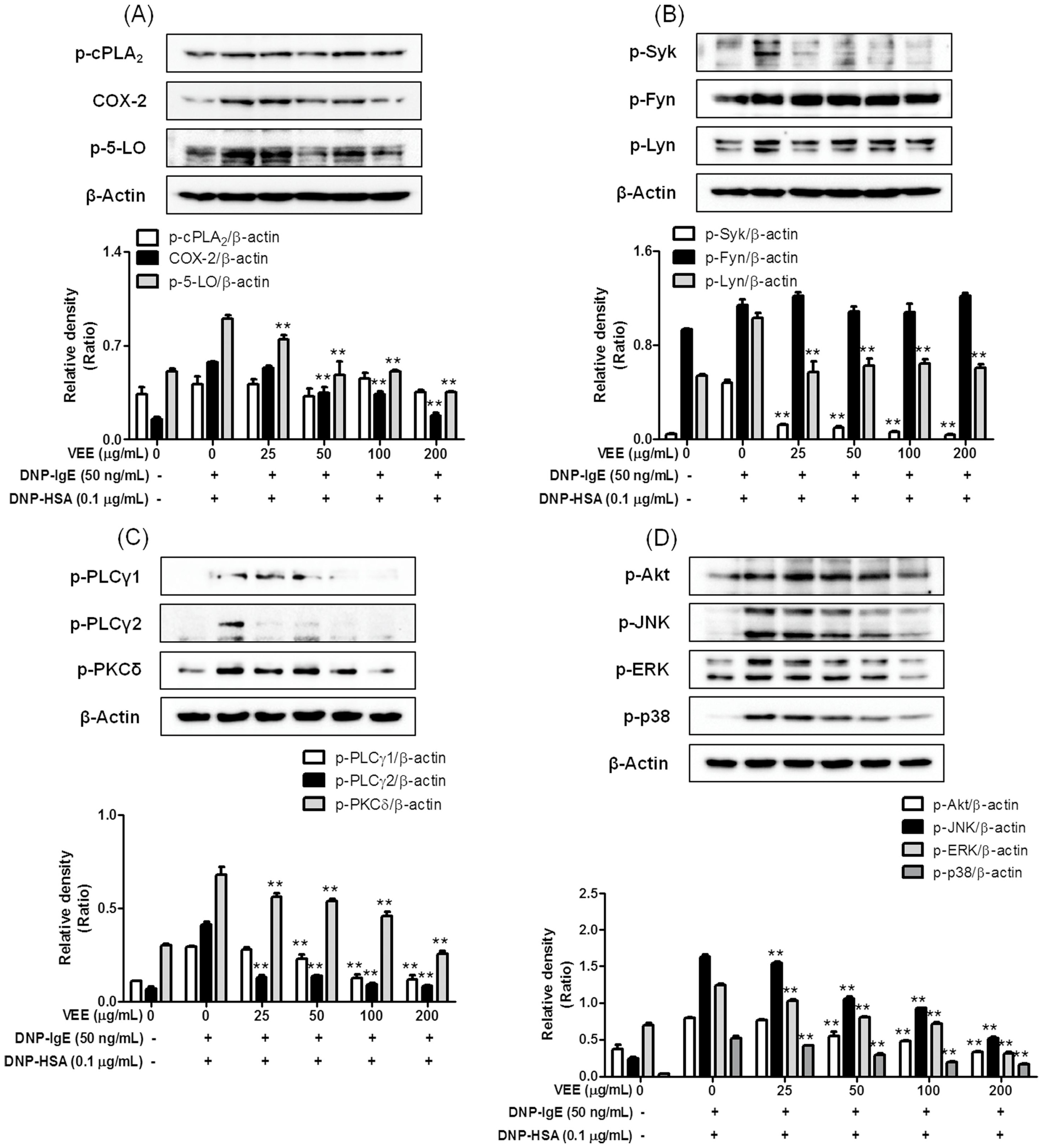
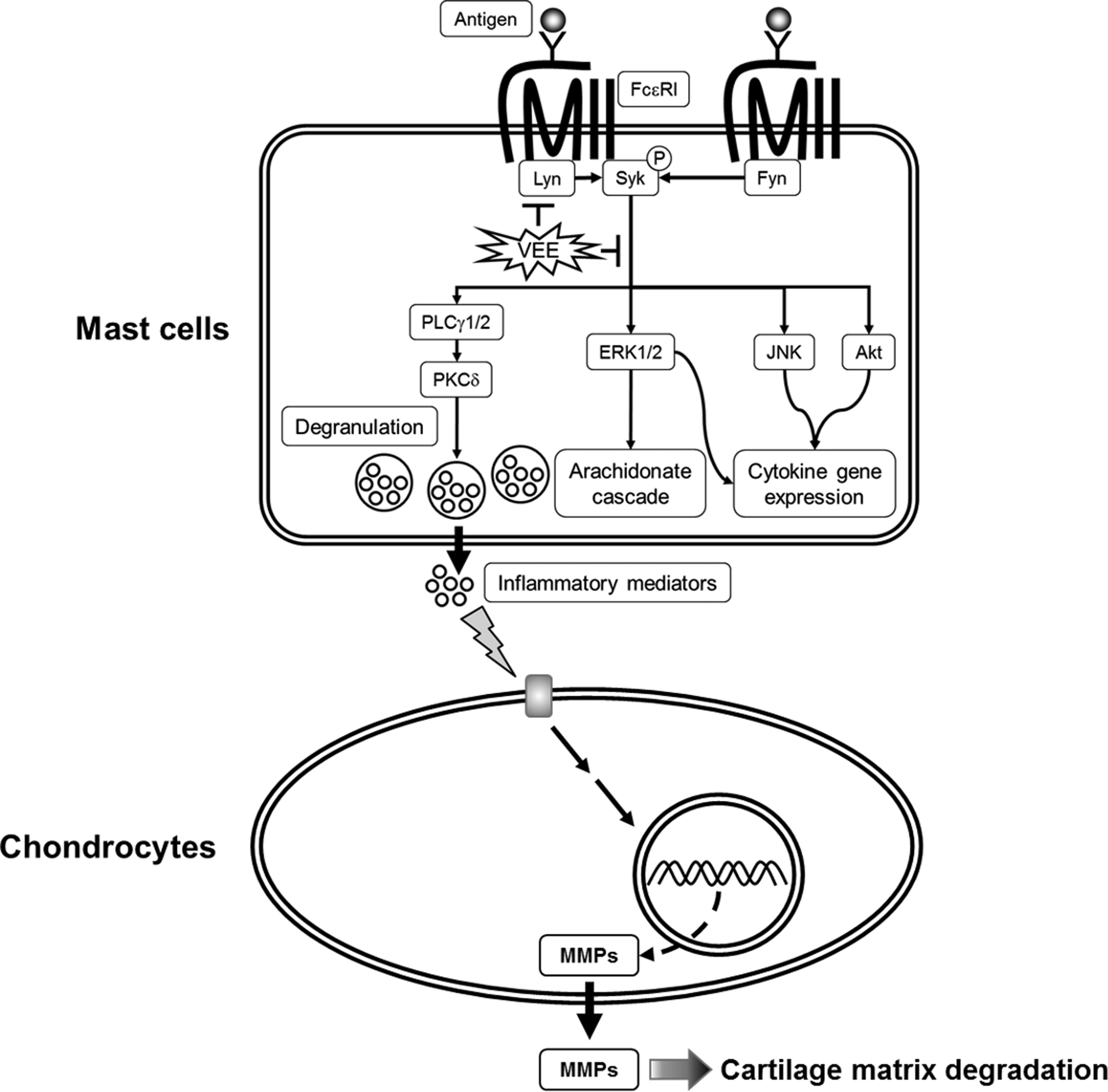
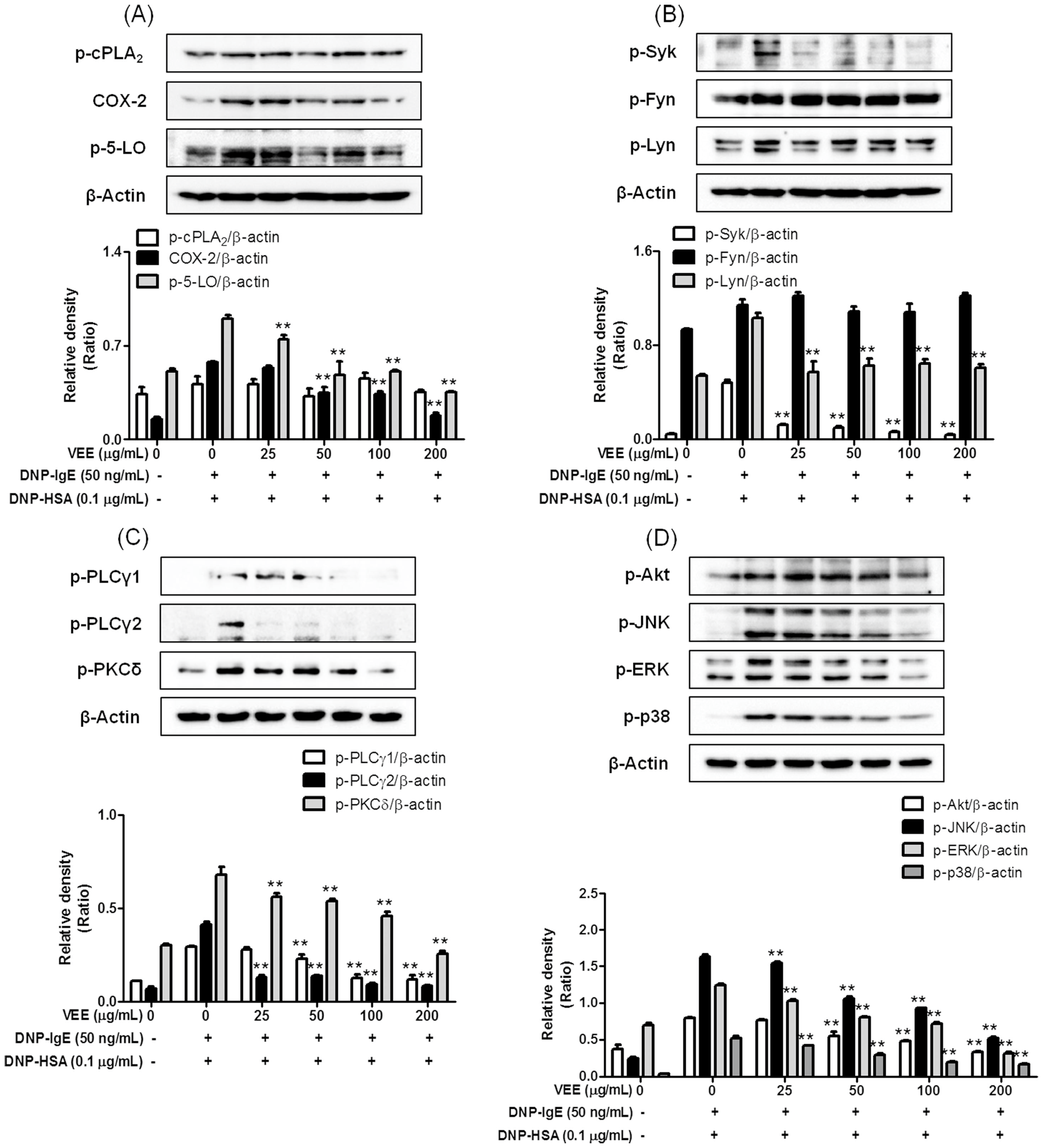
2.3. Effects of VEE on the Activation of FcεRI and Arachidonate Cascades
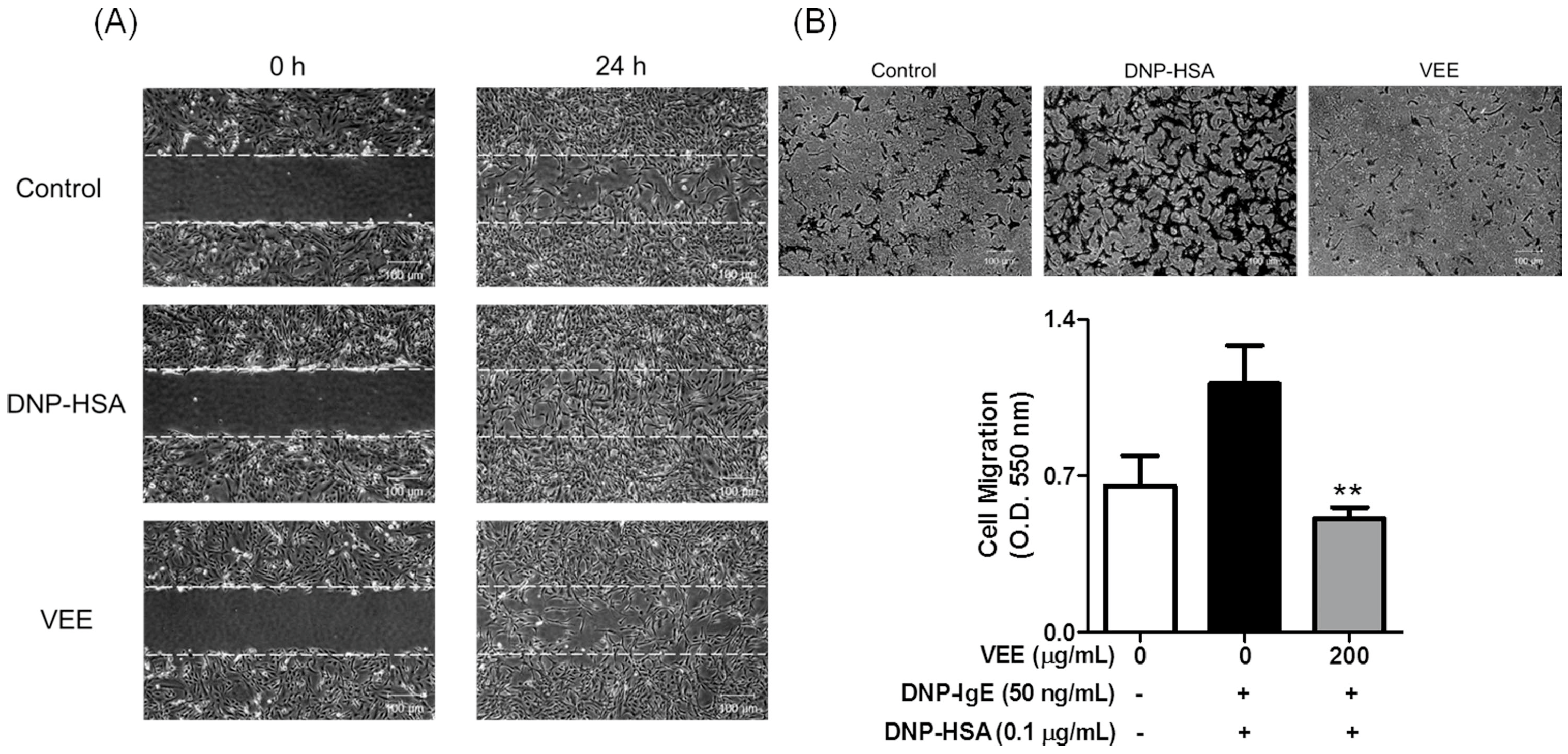
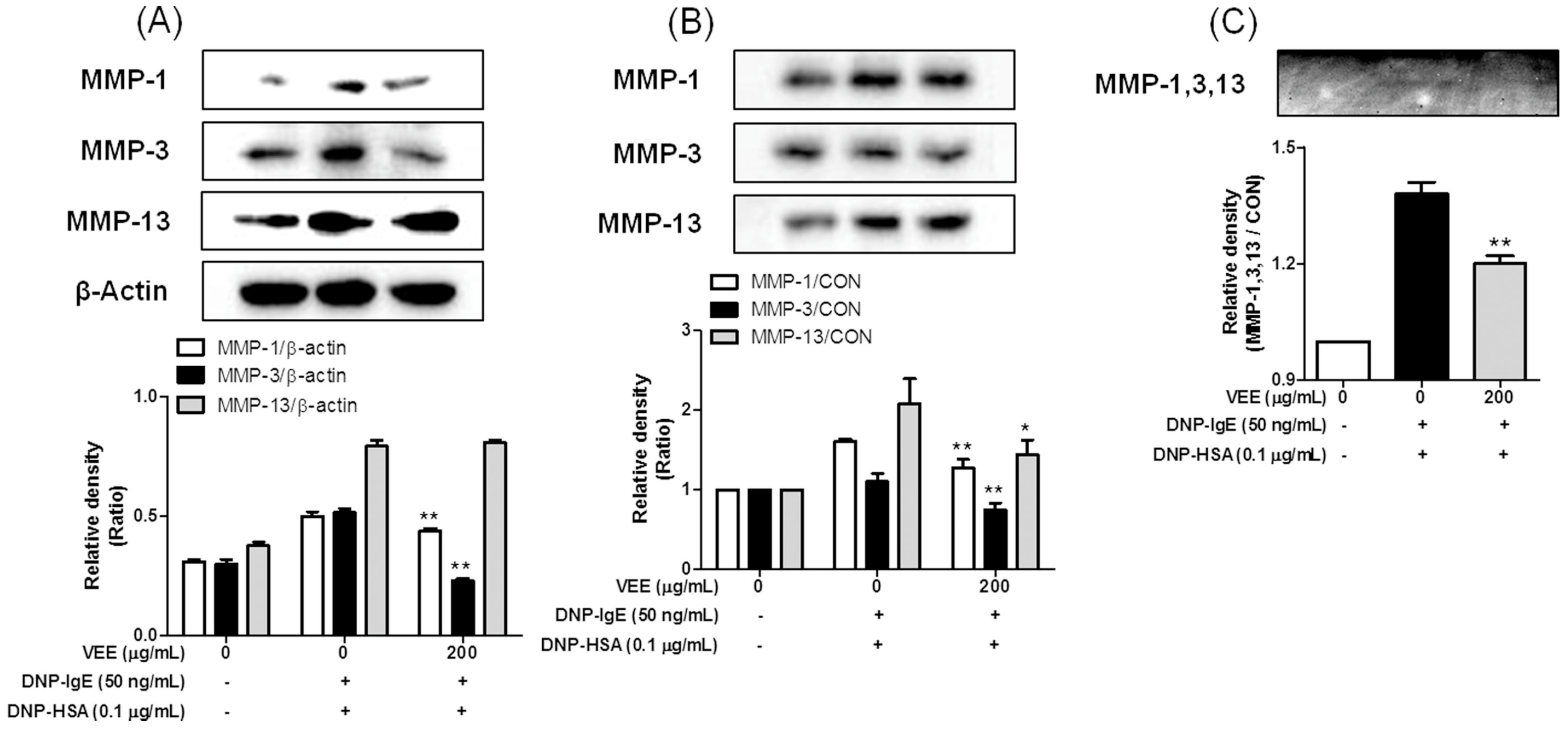
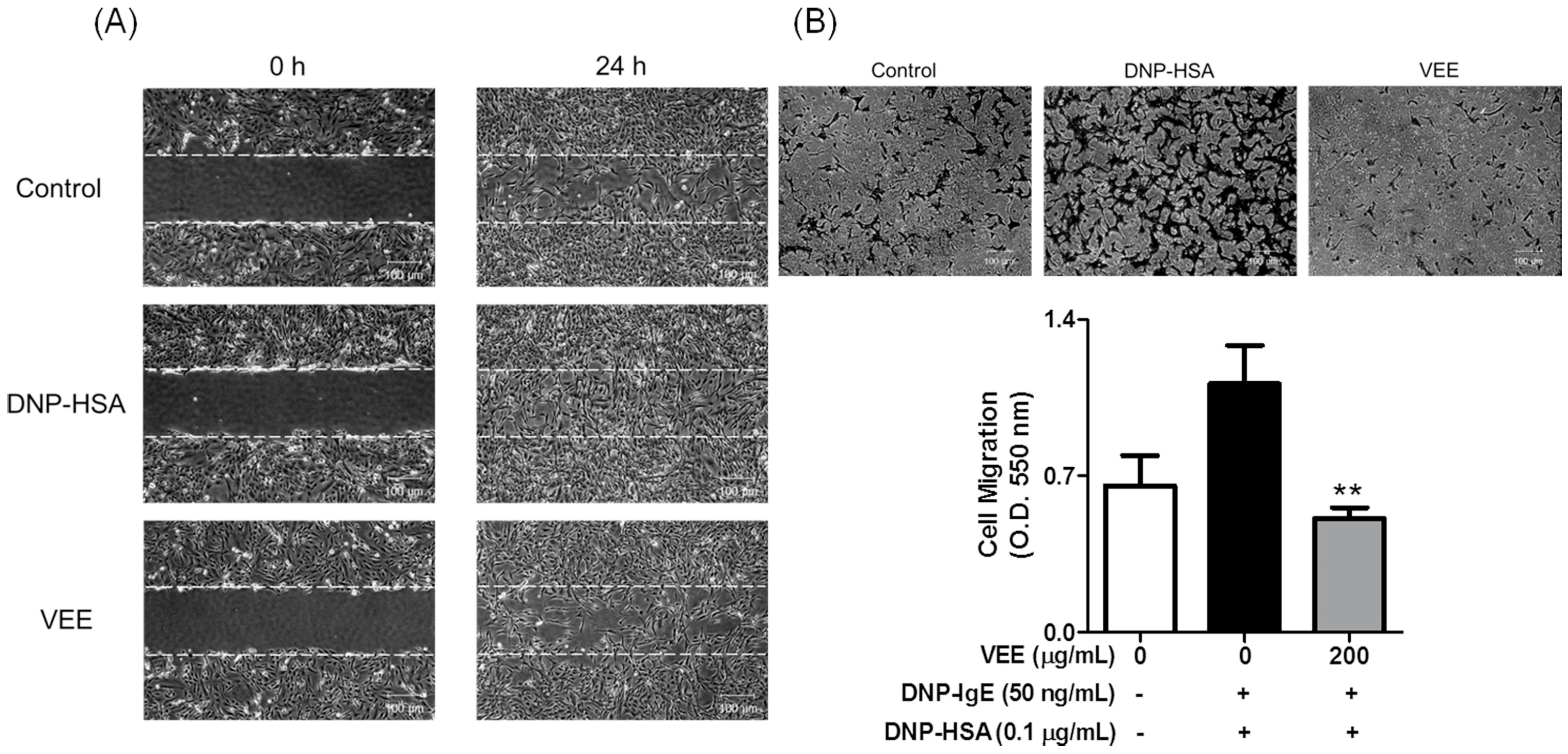
2.4. Effects of VEE on Cell Migration, and the Expression, Secretion, and/or Activity of MMPs in MDIM-Activated SW1353 Cells
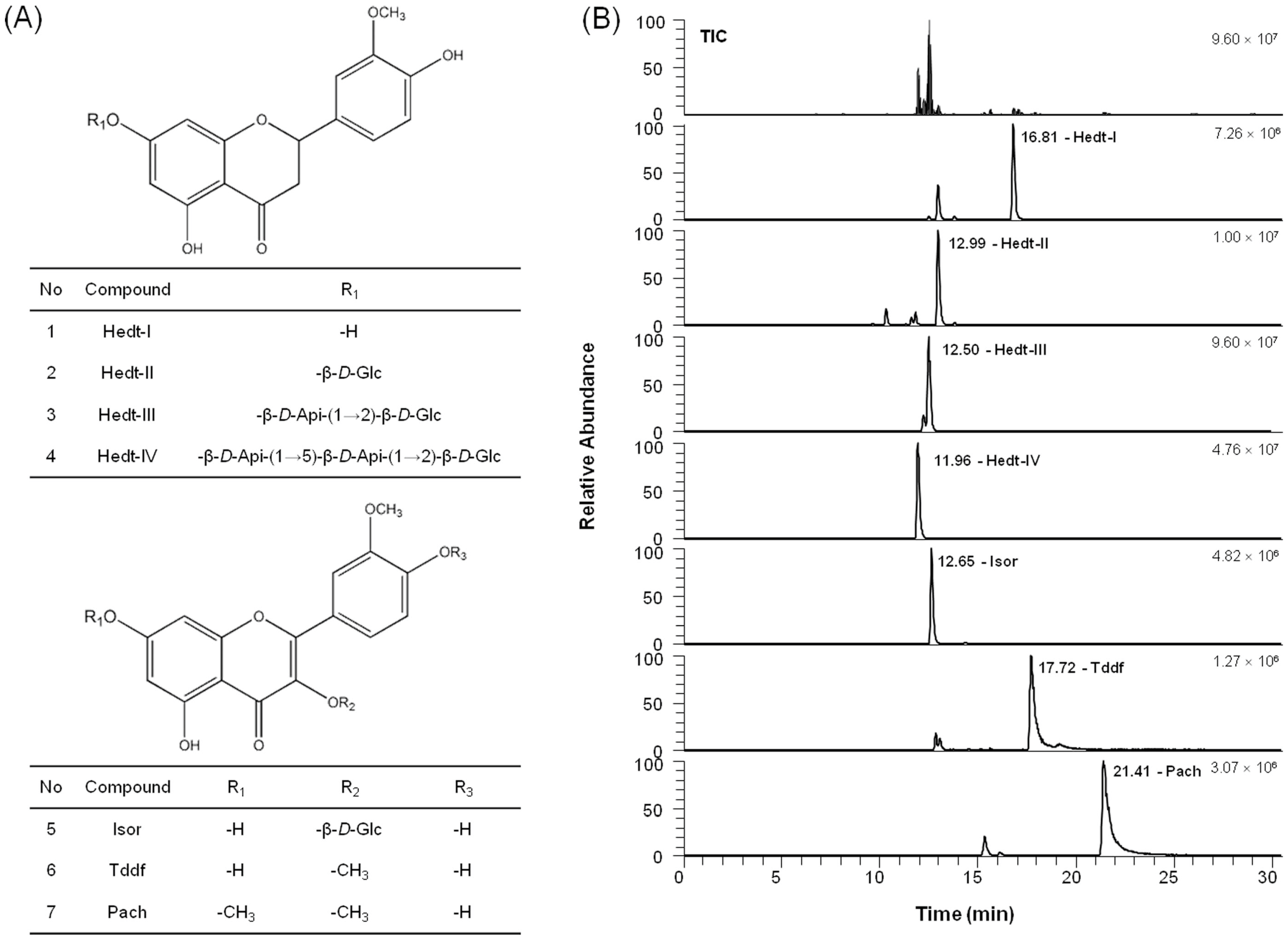
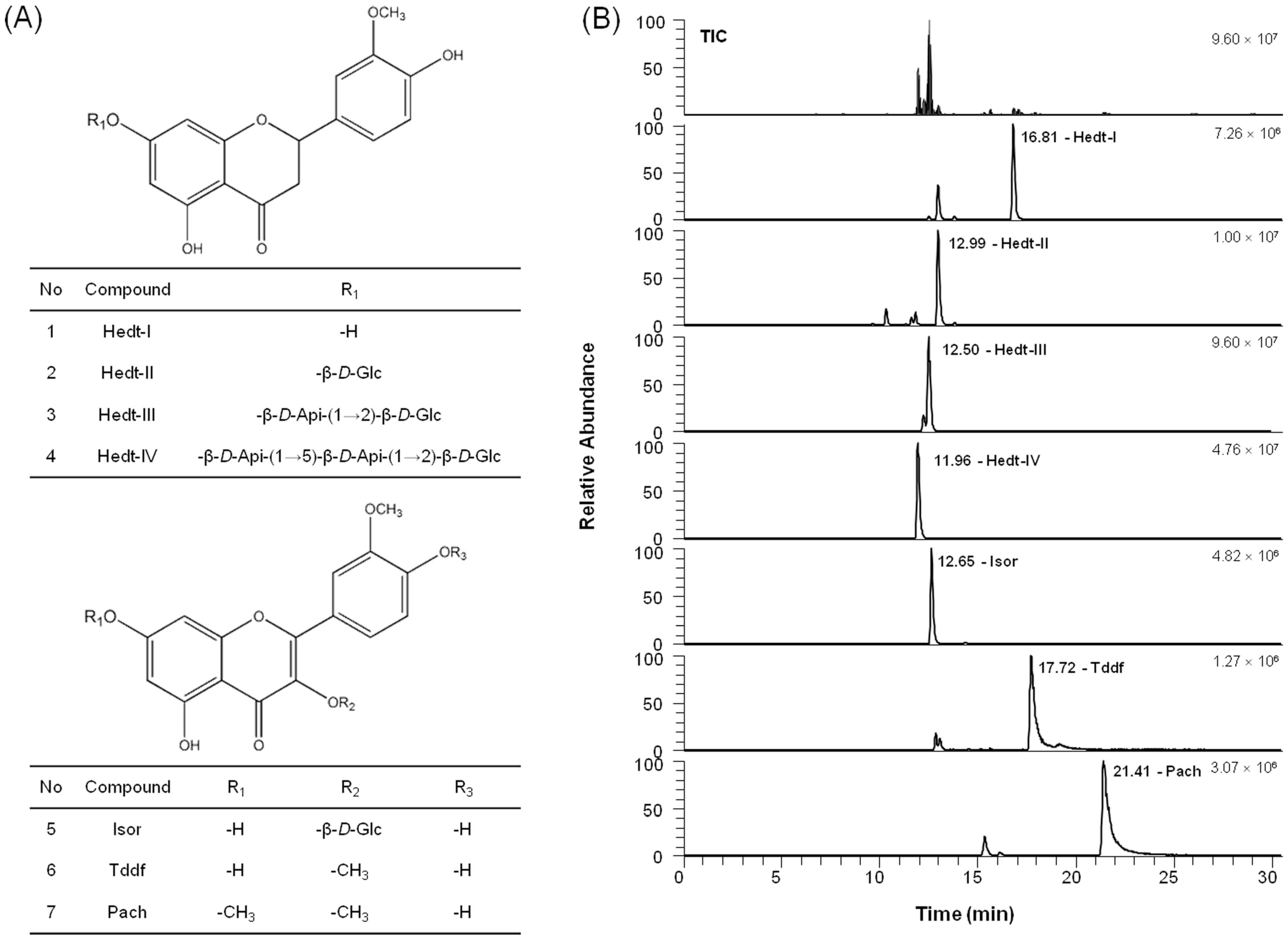
2.5. Chemical Profile of the Active Compounds in VEE
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Viscum Coloratum Extracts
4.3. Determination of Total Phenolic and Flavonoid Compounds
4.4. Analytical Methods
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. β-Hexosaminidase Activity Assay
4.8. Enzyme-Linked Immunosorbent Assay for TNF-α and IL-4
4.9. Enzyme Immunoassay Analysis for PGD2 and LTC4
4.10. Cell Migration
4.11. Precipitation of Secreted Proteins
4.12. Zymography Assay
4.13. Immunoblotting Analysis
4.14. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Xu, Y.; Chen, G. Mast cell and autoimmune diseases. Mediat. Inflamm. 2015, 2015, 246126. [Google Scholar] [CrossRef] [PubMed]
- Uzzaman, A.; Cho, S.H. Chapter 28: Classification of hypersensitivity reactions. Allergy Asthma Proc. 2012, 33 (Suppl. 1), S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Gilfillan, A.M.; Tkaczyk, C. Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 2006, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Mican, J.M.; Metcalfe, D.D. Arthritis and mast cell activation. J. Allergy Clin. Immunol. 1990, 86, 677–683. [Google Scholar] [CrossRef]
- Lohman, R.J.; Cotterell, A.J.; Barry, G.D.; Liu, L.; Suen, J.Y.; Vesey, D.A.; Fairlie, D.P. An antagonist of human protease activated receptor-2 attenuates PAR2 signaling, macrophage activation, mast cell degranulation, and collagen-induced arthritis in rats. FASEB J. 2012, 26, 2877–2887. [Google Scholar] [CrossRef] [PubMed]
- Kakizoe, E.; Li, S.H.; Kobayashi, Y.; Nishikori, Y.; Dekio, S.; Okunishi, H. Increases in mast cells and chymase in fibroproliferative paws of collagen-induced arthritic mice. Inflamm. Res. 1999, 48, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.C.; Rajabalaya, R.; Lee, S.H.; Tennakoon, K.U.; Le, Q.V.; Idris, A.; Zulkipli, I.N.; Keasberry, N.; David, S.R. Parasitic Mistletoes of the Genera Scurrula and Viscum: From Bench to Bedside. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.; Kim, J.; Choi, Y.; Choj, K.; Park, K. One of the Korean mistletoe species, Loranthus yadoriki Sieb. exhibited potent inhibitory activities against monoamine oxidases. Planta Med. 2011, 77. [Google Scholar] [CrossRef]
- Gorter, R.W.; van Wely, M.; Stoss, M.; Wollina, U. Subcutaneous infiltrates induced by injection of mistletoe extracts (Iscador). Am. J. Ther. 1998, 5, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Hyun, C.K.; Shin, H.K. Cytotoxic effects of the components in heat-treated mistletoe (Viscum album). Cancer Lett. 1999, 139, 207–213. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, J.K.; Kim, H.Y.; Park, S.M.; Lee, S.M. Immunomodulating effects of Korean mistletoe lectin in vitro and in vivo. Int. Immunopharmacol. 2009, 9, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Z.; Fan, R.; Gao, X.; Yu, M.; Li, H.; Wei, H.; Bi, K. Simultaneous determination of ten flavonoids from Viscum coloratum grown on different host species and different sources by LC-MS. Chem. Pharm. Bull. 2011, 59, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Yang, E.J.; Son, Y.K.; Yeo, J.H.; Song, K.S. Enhanced anti-oxidative effect of fermented Korean mistletoe is originated from an increase in the contents of caffeic acid and lyoniresinol. Food Funct. 2016, 7, 2270–2277. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Anti-allergic action of aged black garlic extract in RBL-2H3 cells and passive cutaneous anaphylaxis reaction in mice. J. Med. Food 2014, 17, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Kim, N.Y.; Seo, J.M.; Kim, S.J.; Lee, S.Y.; Kim, S.K.; Kim, H.D.; Lee, S.W.; Kim, M.R. Inhibitory effects of mulberry fruit extract in combination with naringinase on the allergic response in IgE-activated RBL-2H3 cells. Int. J. Mol. Med. 2014, 33, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Yang, J.H.; Yang, H.J.; Cho, W.K.; Ma, J.Y. Inhibitory effect of fermented Arctium lappa fruit extract on the IgE-mediated allergic response in RBL2H3 cells. Int. J. Mol. Med. 2016, 37, 501–508. [Google Scholar] [PubMed]
- Sun, H.B. Mechanical loading, cartilage degradation, and arthritis. Ann. N. Y. Acad. Sci. 2010, 1211, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Malemud, C.J. Matrix metalloproteinases: Role in skeletal development and growth plate disorders. Front. Biosci. J. Virtual Libr. 2006, 11, 1702–1715. [Google Scholar] [CrossRef]
- Paul, W.E. Interleukin-4 production by Fc epsilon R+ cells. Skin Pharmacol. 1991, 4 (Suppl. 1), 8–14. [Google Scholar] [CrossRef] [PubMed]
- Russo, C.; Polosa, R. TNF-alpha as a promising therapeutic target in chronic asthma: A lesson from rheumatoid arthritis. Clin. Sci. 2005, 109, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Park, E.S.; Kim, M.R.; Sok, D.E. Inhibitory effect of N-Acyl dopamines on IgE-mediated allergic response in RBL-2H3 cells. Lipids 2013, 48, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Sok, D.E.; Kim, M.R. Effect of endocannabinoids on IgE-mediated allergic response in RBL-2H3 cells. Int. Immunopharmacol. 2013, 17, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Yang, J.H.; Kim, Y.S.; Cho, W.K.; Ma, J.Y. Inhibitory Effect of Loranthus parasiticus on IgE-Mediated Allergic Responses in RBL-2H3 Cells. Mediat. Inflamm. 2016, 2016, 8742562. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M.; Tsuchimochi, K.; Ijiri, K.; Li, Y. Defining the roles of inflammatory and anabolic cytokines in cartilage metabolism. Ann. Rheum. Dis. 2008, 67 (Suppl. 3), iii75–iii82. [Google Scholar] [CrossRef] [PubMed]
- Burrage, P.S.; Mix, K.S.; Brinckerhoff, C.E. Matrix metalloproteinases: Role in arthritis. Front. Biosci. J. Virtual Libr. 2006, 11, 529–543. [Google Scholar] [CrossRef]
- Nigrovic, P.A.; Lee, D.M. Mast cells in inflammatory arthritis. Arthritis Res. Ther. 2005, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, Y.; Shimoi, K.; Kumazawa, S.; Yamamoto, K.; Kinae, N.; Osawa, T. Identification and antioxidant activity of flavonoid metabolites in plasma and urine of eriocitrin-treated rats. J. Agric. Food Chem. 2000, 48, 3217–3224. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lim, H.J.; Ryu, J.H. In vitro anti-inflammatory activity of 3-O-methyl-flavones isolated from Siegesbeckia glabrescens. Bioorg. Med. Chem. Lett. 2008, 18, 1511–1514. [Google Scholar] [CrossRef] [PubMed]
- Ishitsuka, H.; Ohsawa, C.; Ohiwa, T.; Umeda, I.; Suhara, Y. Antipicornavirus flavone Ro 09-0179. Antimicrob. Agents Chemother. 1982, 22, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Fanning, L.B.; Boyce, J.A. Lipid mediators and allergic diseases. Ann. Allergy Asthma Immunol. 2013, 111, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Siraganian, R.P. Inhibition of IgE-mediated histamine release from rat basophilic leukemia cells and rat mast cells by inhibitors of transmethylation. J. Immunol. 1981, 127, 1339–1344. [Google Scholar] [PubMed]
- Ouyang, P. An in vitro model to study mesenchymal-epithelial transformation. Biochem. Biophys. Res. Commun. 1998, 246, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Chang, Y.; Jung, J.C. Staurosporine induces ganglion cell differentiation in part by stimulating urokinase-type plasminogen activator expression and activation in the developing chick retina. Biochem. Biophys. Res. Commun. 2012, 423, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Yim, N.H.; Im, M.; Jung, Y.P.; Kim, T.; Ma, J.Y. Suppression of the invasive potential of highly malignant tumor cells by KIOM-C, a novel herbal medicine, via inhibition of NF-kappaB activation and MMP-9 expression. Oncol. Rep. 2014, 31, 287–297. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | tR (min) | Formula | [M − H]− Calculated (m/z) | [M − H]− Detected (m/z) | Delta (ppm) | MS/MS (m/z) | Area Ratio (VEE/VAE) | |
|---|---|---|---|---|---|---|---|---|---|
| Ratio (%) | SD | ||||||||
| 1 | Hedt-I | 16.81 | C16H14O6 | 301.07176 | 301.07108 | −0.043 | 151.00218 | 235.71 | 16.09 |
| 2 | Hedt-II | 12.99 | C22H24O11 | 463.12458 | 463.12326 | 0.159 | 301.07114, 151.00224 | 96.24 | 5.25 |
| 3 | Hedt-III | 12.50 | C27H32O15 | 595.16684 | 595.16547 | −0.781 | 301.07104, 151.00211 | 126.39 | 6.01 |
| 4 | Hedt-IV | 11.96 | C32H40O19 | 727.20910 | 727.20770 | −0.747 | 301.07101 | 128.20 | 9.00 |
| 5 | Isor | 12.65 | C22H22O12 | 477.10385 | 477.10291 | −0.442 | 357.06079, 314.04269, 271.02429 | 162.03 | 8.45 |
| 6 | Tddf | 17.72 | C17H14O7 | 329.06668 | 329.06619 | 0.281 | 314.04269, 299.0191 | 671.04 | 56.65 |
| 7 | Pach | 21.41 | C18H16O7 | 343.08233 | 343.08142 | −0.051 | 328.05823, 313.03488 | 994.33 | 78.50 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, J.-M.; Yang, J.-H.; Kim, Y.S.; Yang, H.J.; Cho, W.-K.; Ma, J.Y. Inhibitory Effects of Viscum coloratum Extract on IgE/Antigen-Activated Mast Cells and Mast Cell-Derived Inflammatory Mediator-Activated Chondrocytes. Molecules 2017, 22, 37. https://doi.org/10.3390/molecules22010037
Yoo J-M, Yang J-H, Kim YS, Yang HJ, Cho W-K, Ma JY. Inhibitory Effects of Viscum coloratum Extract on IgE/Antigen-Activated Mast Cells and Mast Cell-Derived Inflammatory Mediator-Activated Chondrocytes. Molecules. 2017; 22(1):37. https://doi.org/10.3390/molecules22010037
Chicago/Turabian StyleYoo, Jae-Myung, Ju-Hye Yang, Young Soo Kim, Hye Jin Yang, Won-Kyung Cho, and Jin Yeul Ma. 2017. "Inhibitory Effects of Viscum coloratum Extract on IgE/Antigen-Activated Mast Cells and Mast Cell-Derived Inflammatory Mediator-Activated Chondrocytes" Molecules 22, no. 1: 37. https://doi.org/10.3390/molecules22010037
APA StyleYoo, J.-M., Yang, J.-H., Kim, Y. S., Yang, H. J., Cho, W.-K., & Ma, J. Y. (2017). Inhibitory Effects of Viscum coloratum Extract on IgE/Antigen-Activated Mast Cells and Mast Cell-Derived Inflammatory Mediator-Activated Chondrocytes. Molecules, 22(1), 37. https://doi.org/10.3390/molecules22010037