
Hyperforin Exhibits Antigenotoxic Activity on Human and Bacterial Cells

 ,
,
Abstract
:1. Introduction
2. Results
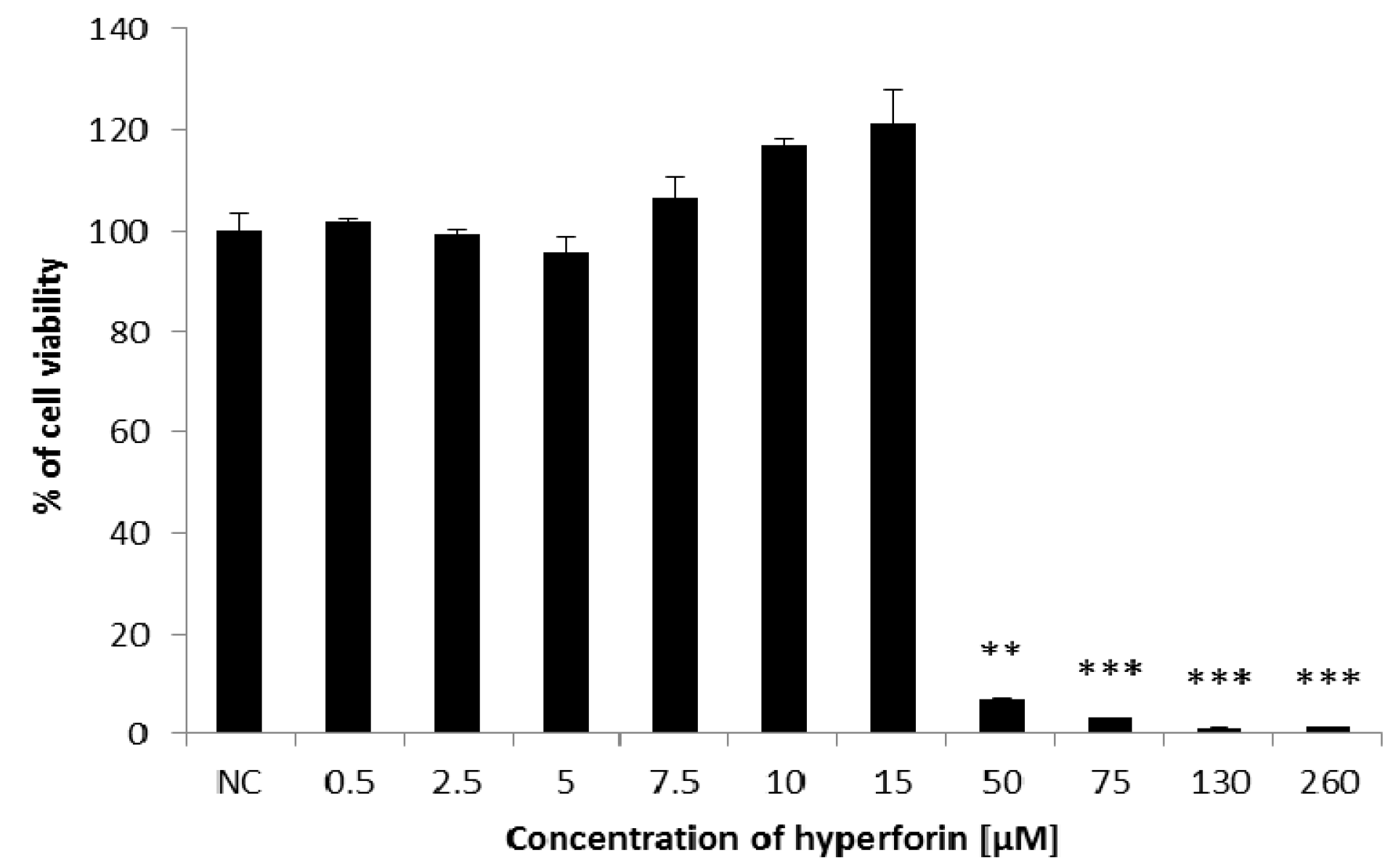
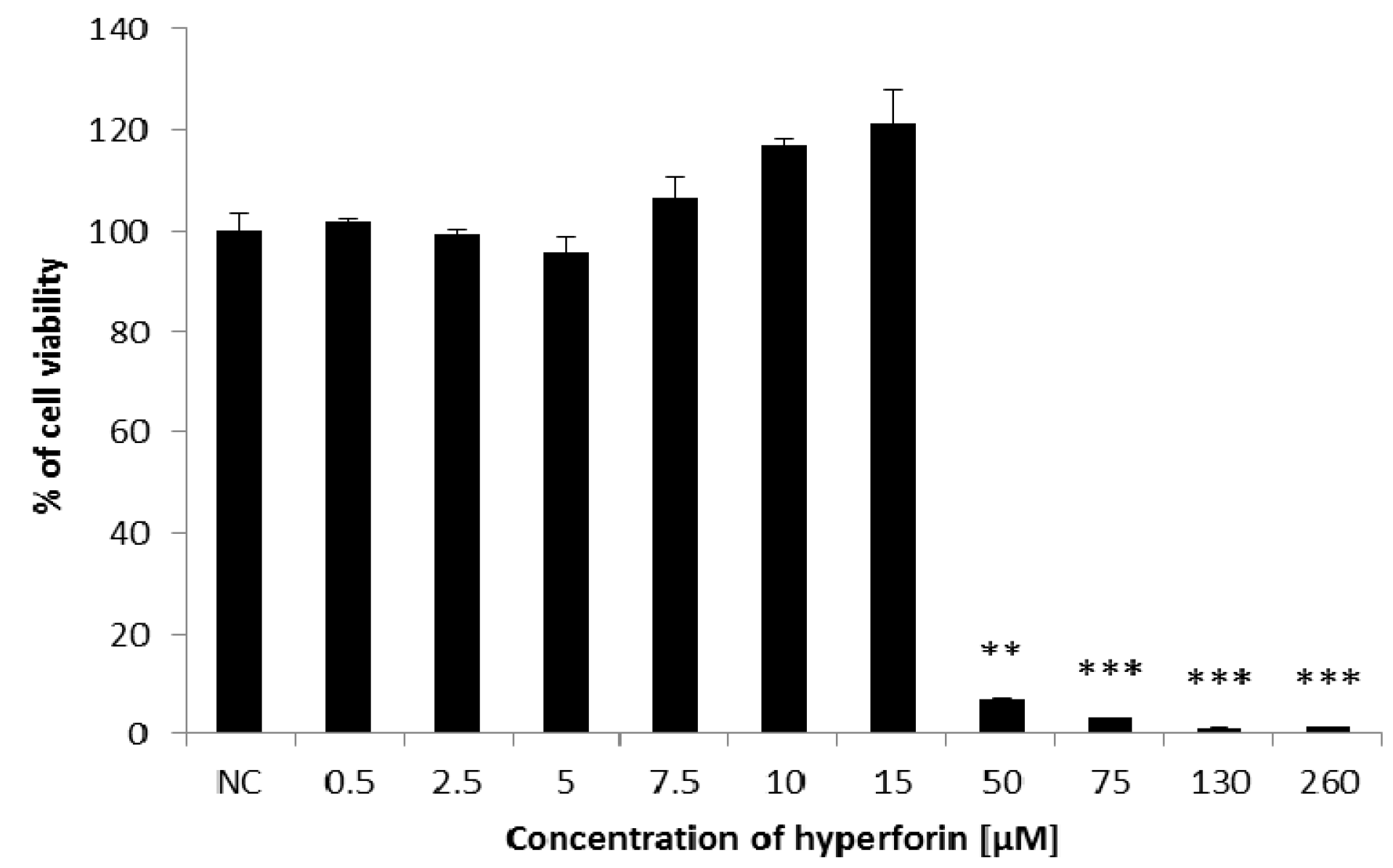
2.1. Cell Viability
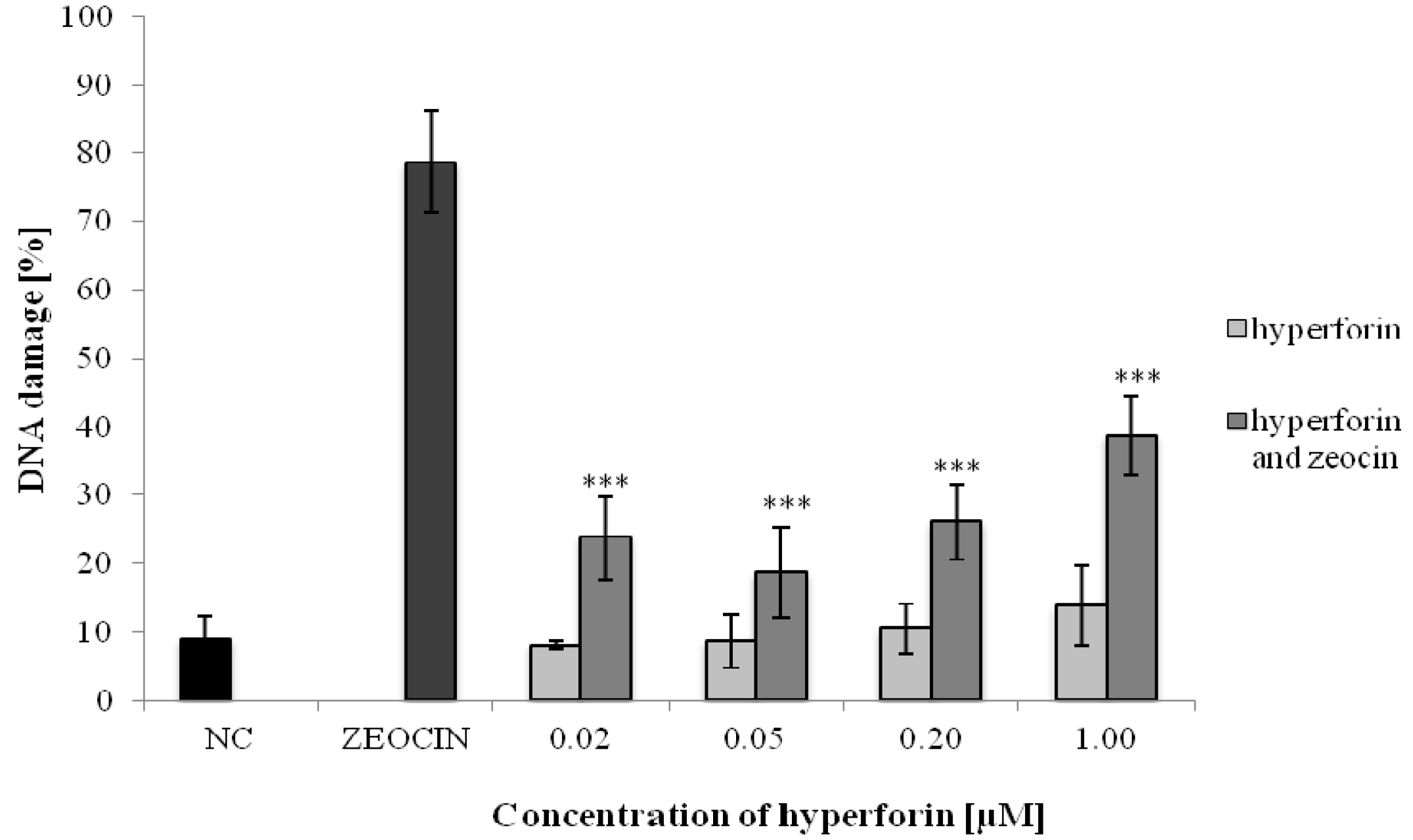
2.2. The Alkaline Comet Assay
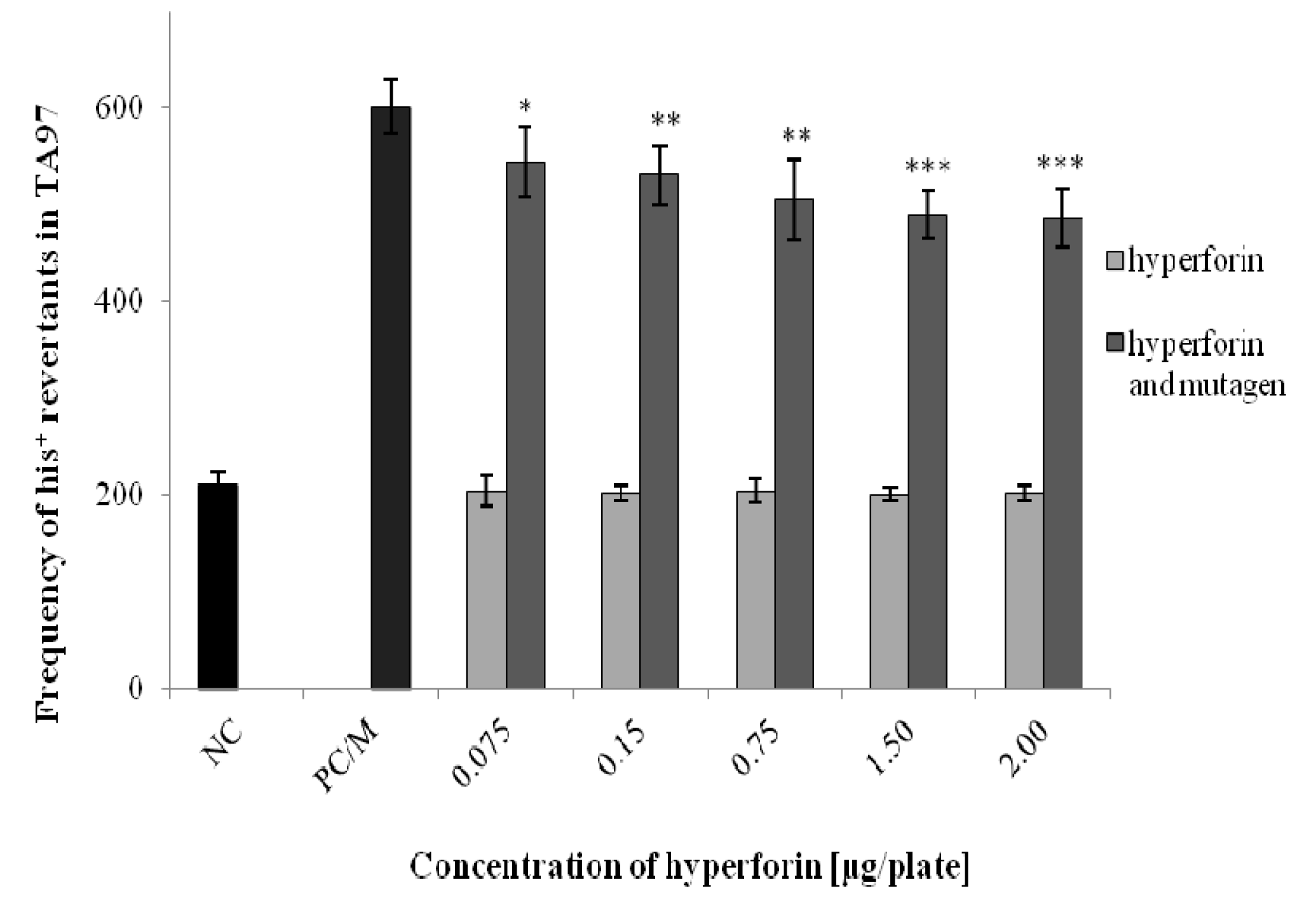
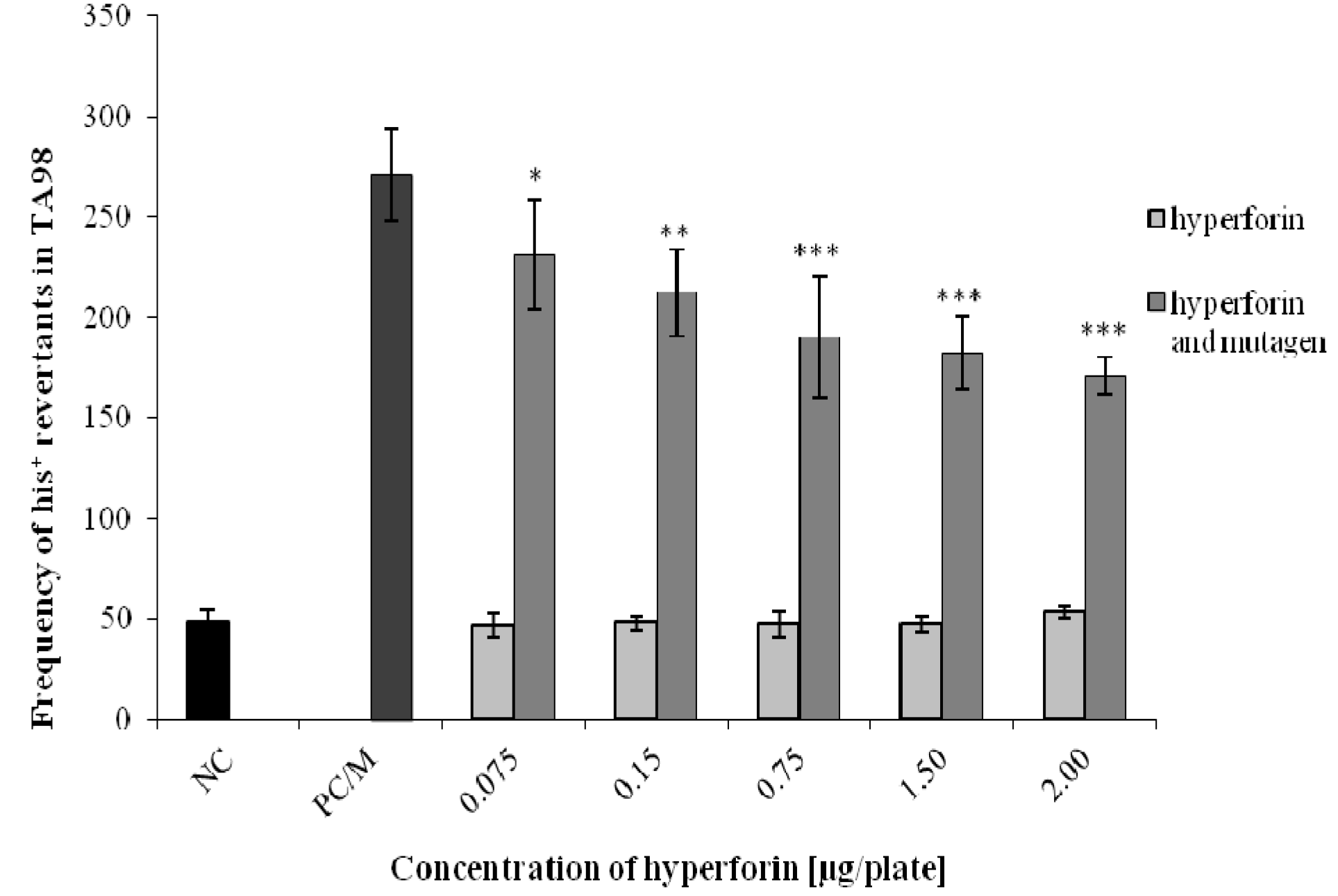
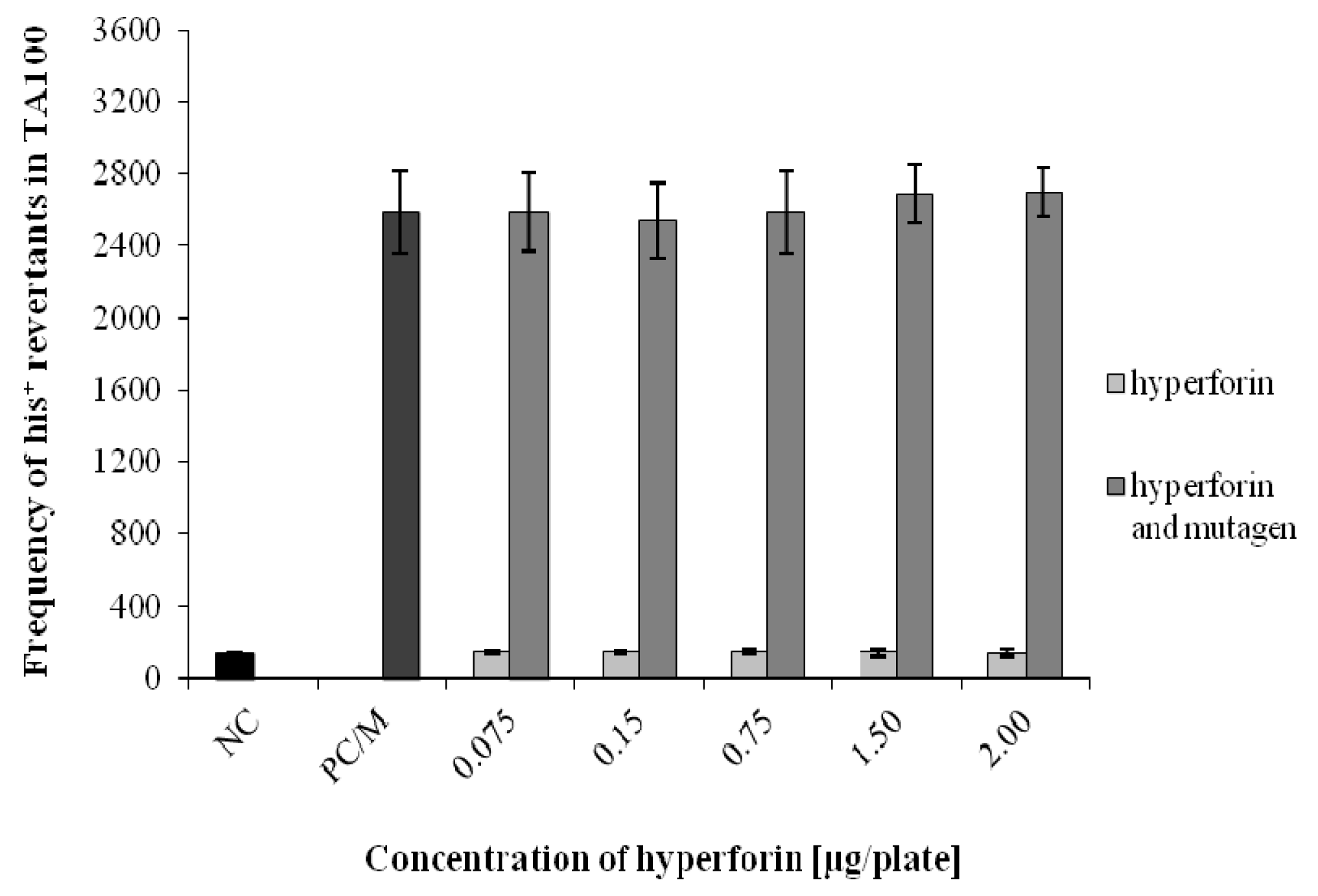
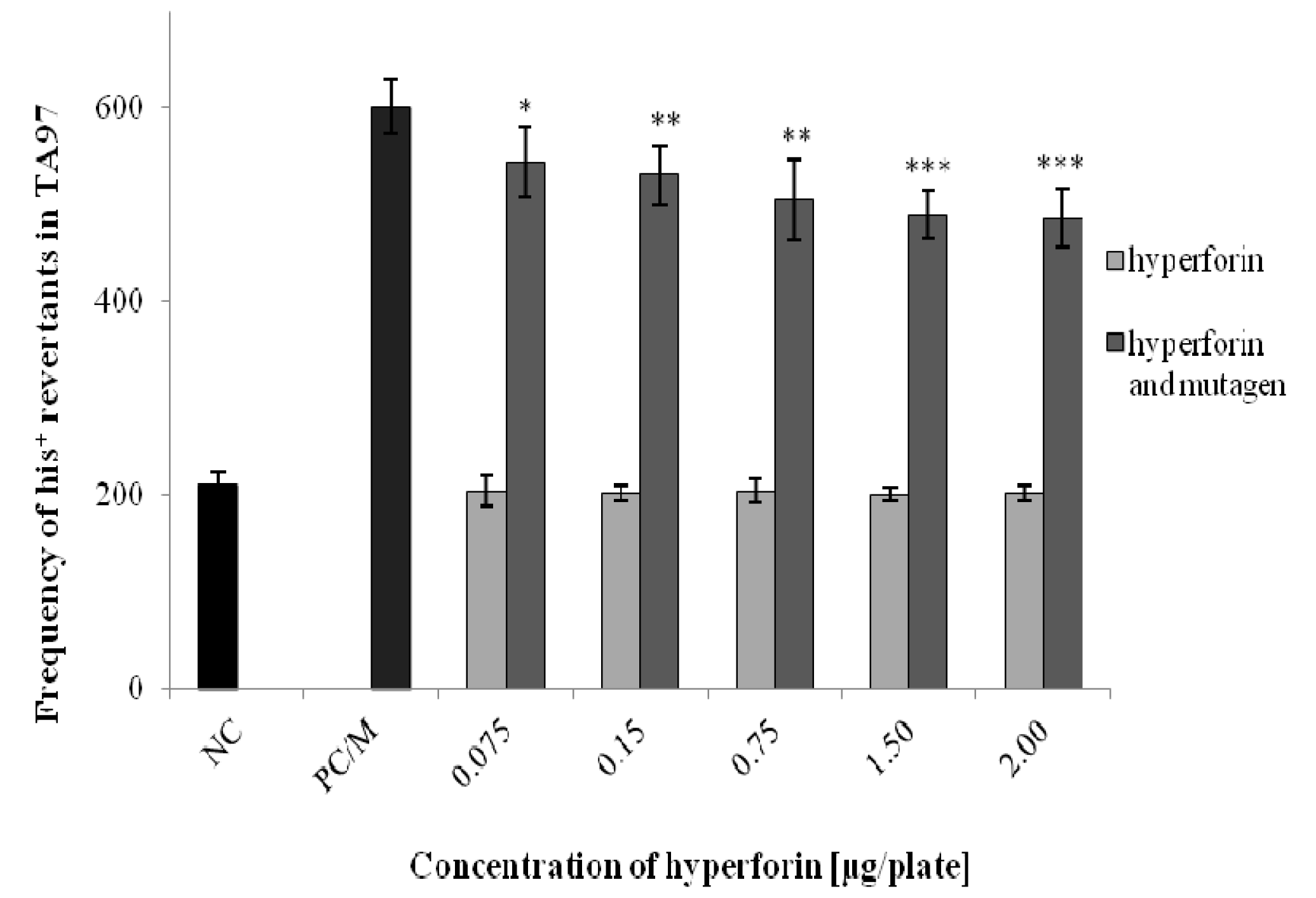
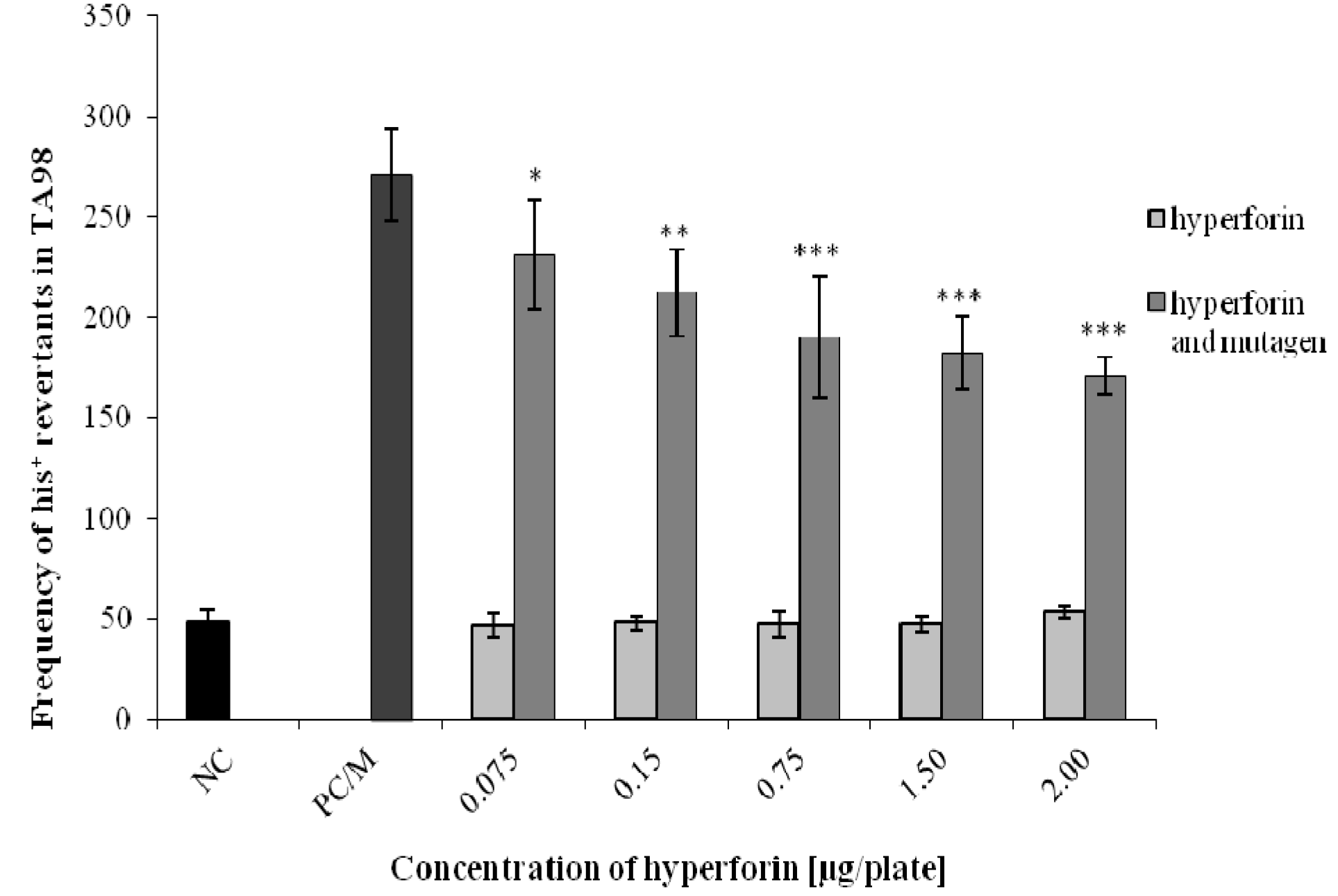
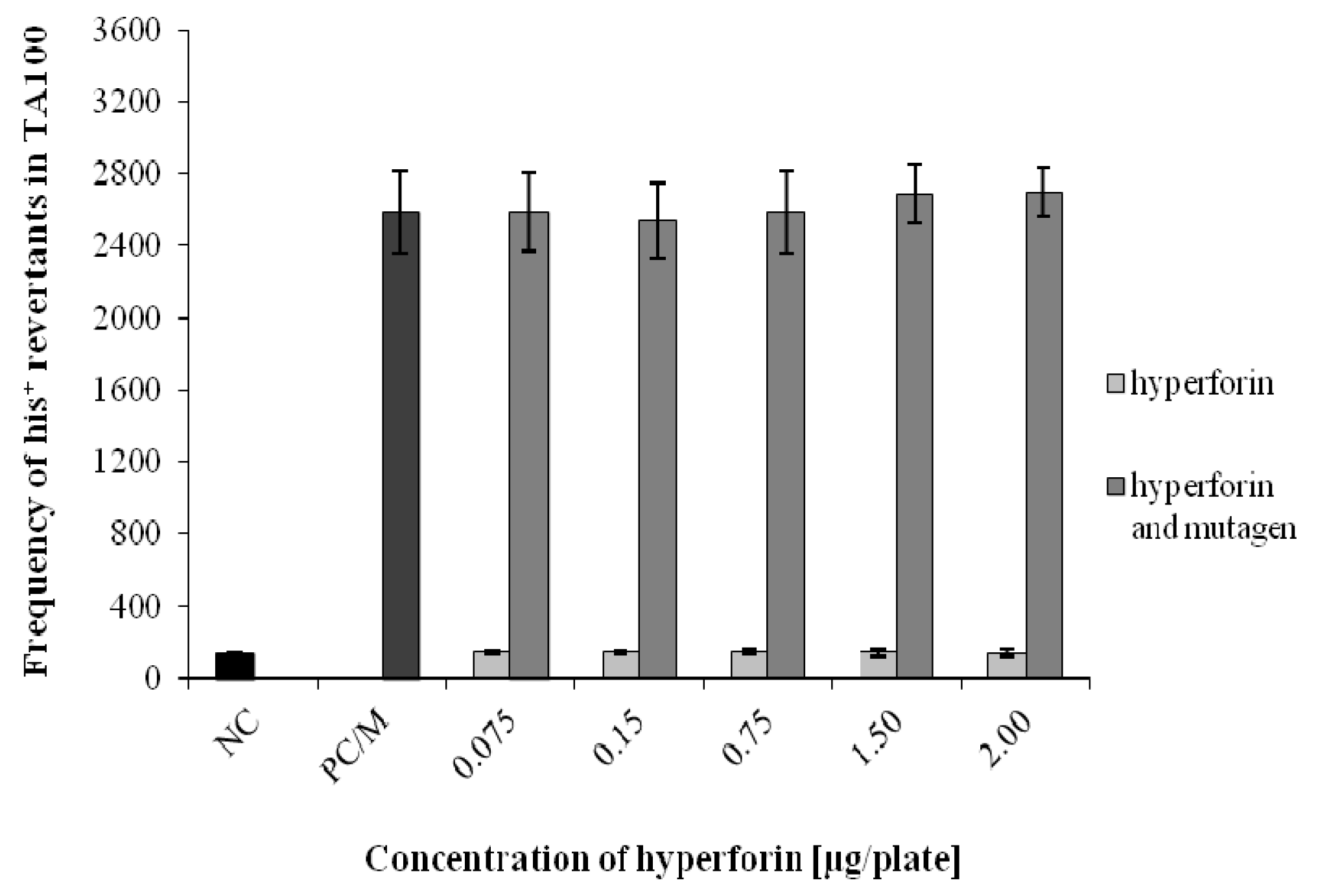
2.3. The Ames/S. typhimurium Test
2.4. In Vitro Mammalian Chromosome Aberration Test
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Experimental Cells
4.2.1. Human Lymphocytes
4.2.2. Bacterial Strains of S. typhimurium
4.2.3. Human Cell Line HepG2
4.2.4. Human Cell Line VH10
4.3. Cell Viability (MTT Test)
4.4. Comet Assay
4.5. Ames/S. typhimurium Test
4.6. In Vitro Mammalian Chromosome Aberration Test
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miadokova, E.; Nadova, S.; Vlckova, V.; Duhova, V.; Kopaskova, M.; Cipak, L.; Rauko, P.; Mucaji, P.; Grancai, D. Antigenotoxic Effect of Extract from Cynara cardunculus L. Phytother. Res. 2008, 22, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Fedel-Miyasato, L.E.; Formagio, A.S.; Auharek, S.A.; Kassuya, C.A.; Navarro, S.D.; Cunha-Laura, A.L.; Monreal, A.C.; Vieira, M.C.; Oliveira, R.J. Antigenotoxic and antimutagenic effects of Schinus terebinthifolius Raddi in Allium cepa and Swiss mice: A comparative study. Genet. Mol. Res. 2014, 13, 3411–3425. [Google Scholar] [CrossRef] [PubMed]
- Gafrikova, M.; Galova, E.; Sevcovivova, A.; Imreova, P.; Mucaji, P.; Miadokova, E. Extract from Armoracia rusticana and Its Flavonoid Components Protect Human Lymphocytes against Oxidative Damage Induced by Hydrogen Peroxide. Molecules 2014, 19, 3160–3172. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Husain, G.M.; Singh, P.N.; Kumar, V. Antiaggressive activity of hyperforin: A preclinical study. Drug Discov. Ther. 2009, 3, 162–167. [Google Scholar] [PubMed]
- Greeson, J.M.; Sanford, B.; Monti, D.A. St. John’s wort (Hypericum perforatum): A review of the current pharmacological, toxicological, and clinical literature. Psychopharmacology 2001, 153, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Hashida, C.; Tanaka, N.; Kashiwada, Y.; Ogawa, M.; Takaishi, Y. Prenylated phloroglucinol derivatives from Hypericum perforatum var. angustifolium. Chem. Pharm. Bull. 2008, 56, 1164–1167. [Google Scholar] [CrossRef] [PubMed]
- Ivetic, V.; Trivic, S.; Pogancev, M.K.; Popovic, M.; Zlinská, J. Effects of St John’s wort (Hypericum perforatum L.) extracts on epileptogenesis. Molecules 2011, 16, 8062–8075. [Google Scholar] [CrossRef] [PubMed]
- Božin, B.; Kladar, N.; Grujić, N.; Anačkov, G.; Samojlik, I.; Gavarić, N.; Conić, B.S. Impact of origin and biological source on chemical composition, anticholinesterase and antioxidant properties of some St. John’s wort species (Hypericum spp., Hypericaceae) from the Central Balkans. Molecules 2013, 18, 11733–11750. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.A.; Martínez-Poveda, B.; Amores-Sánchez, M.I.; Quesada, A.R. Hyperforin: More than an antidepressant bioactive compound? Life Sci. 2006, 79, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Schempp, C.M.; Pelz, K.; Wittmer, A.; Schopf, E.; Simon, J.C. Antibacterial activity of hyperforin from St John’s wort, against multiresistant Staphylococcus aureus and Gram-positive bacteria. Lancet 1999, 353, 2129. [Google Scholar] [CrossRef]
- Moore, L.B.; Goodwin, B.; Jones, S.A.; Wisely, G.B.; Serabjit-Singh, C.J.; Willson, T.M.; Collins, J.L.; Kliewer, S.A. St. John’s wort induces hepatic drug metabolism through activation of the pregnane X receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 7500–7502. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Kisselev, P.; Roots, I. St. John’s wort extracts and some of their constituents potently inhibit ultimate carcinogen formation from benzo[a]pyrene-7,8-dihydrodiol by human CYP1A1. Cancer Res. 2003, 63, 8062–8068. [Google Scholar] [PubMed]
- Hostanska, K.; Reichling, J.; Bommer, S.; Weber, M.; Saller, R. Hyperforin a constituent of St John’s wort (Hypericum perforatum L.) extract induces apoptosis by triggering activation of caspases and with hypericin synergistically exerts cytotoxicity towards human malignant cell lines. Eur. J. Pharm. Biopharm. 2003, 56, 121–132. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Billard, C.; Merhi, F.; Bauvois, B. Mechanistic insights into the antileukemic activity of hyperforin. Curr. Cancer Drug Targets 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ševčovičová, A.; Šemeláková, M.; Plšíková, J.; Loderer, D.; Imreová, P.; Gálová, E.; Kožurková, M.; Miadoková, E.; Fedoročko, P. DNA-protective activities of hyperforin and aristoforin. Toxicol. In Vitro 2015, 29, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Komoroski, B.J.; Zhang, S.; Cai, H.; Hutzler, J.M.; Frye, R.; Tracy, T.S.; Strom, S.C.; Lehmann, T.; Ang, C.Y.; Cui, Y.Y.; et al. Induction and inhibition of cytochromes P450 by the St. John’s wort constituent hyperforin in human hepatocyte cultures. Drug Metab. Dispos. 2004, 32, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Krol, K.; Brozda, I.; Skoneczny, M.; Bretner, M.; Skoneczna, A. A genomic screen revealing the importance of vesicular trafficking pathways in genome maintenance and protection against genotoxic stress in diploid Saccharomyces cerevisiae cells. PLoS ONE 2015, 10, e0120702. [Google Scholar] [CrossRef] [PubMed]
- Hudecova, A.; Hasplova, K.; Kellovska, L.; Ikreniova, M.; Miadokova, E.; Galova, E.; Horvathova, E.; Vaculcikova, D.; Gregan, F.; Dusinska, M. Gentiana asclepiadea and Armoracia rusticana can modulate the adaptive response induced by zeocin in human lymphocytes. Neoplasma 2012, 59, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Julias, J.G.; Ferris, A.L.; Hughes, S.H. Construction and characterization of a replication-completet retroviral shuttle vector plasmid. J. Virol. 2002, 76, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Oliva-Trastoy, M.; Defais, M.; Larminat, F. Resistance to the antibiotic Zeocin by stable expression of the Sh ble gene does not fully suppress Zeocin-induced DNA cleavage in human cells. Mutagenesis 2005, 20, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.A.; Jalal, J.A.; Ioanndes, C.; Moselhy, S.S. Assessment of the antimutagenic effect of Doash tea extract fractions. Toxicol. Ind. Health 2012, 28, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Thorne, D.; Kilford, J.; Hollings, M.; Dalrymple, A.; Ballantyne, M.; Meredith, C.; Dillon, D. The mutagenic assessment of mainstream cigarette smoke using the Ames assay: A multi-strain approach. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 782, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Oyaga, S.C.; Valdés, J.C.; Paez, S.B.; Marquez, K.H. DFT description of intermolecular forces between 9-aminoacridines and DNA base pairs. J. Theory. Chem. 2013, 2013, 526569. [Google Scholar]
- Miao, Z.H.; Rao, V.A.; Agama, K.; Antony, S.; Kohn, K.W.; Pommier, Y. 4-nitroquinoline-1-oxide induces the formation of cellular topoisomerase I-DNA cleavage complexes. Cancer Res. 2006, 66, 6540–6545. [Google Scholar] [CrossRef] [PubMed]
- Al-Qurainy, F.; Khan, S. Mutagenic effects of sodium azide and its application in crop improvement. World Appl. Sci. J. 2009, 6, 1589–1601. [Google Scholar]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Nebert, D.W. The Ah locus: Genetic differences in toxicity, cancer, mutation and birth defects. Crit. Rev. Toxicol. 1989, 20, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.P.; Ramos, K.S. Impact of cellular metabolism on the biological effects of benzo[a]pyrene and related hydrocarbons. Drug Metab. Rev. 2001, 33, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Denissenko, M.F.; Pao, A.; Rang, M.; Pfeifer, G.P. Preferential formation of benzo(a)pyrene adducts at lung cancer mutational hotspots in P53. Science 1996, 274, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhang, Q.Y. The Role of Small-Intestinal P450 Enzymes in Protection against Systemic Exposure of Orally Administered Benzo[a]Pyrene. J. Pharmacol. Exp. Ther. 2010, 334, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic activation of polycyclic aromatic hydrocarbons to carcinogens by cytochromes P450 1A1 and 1B1. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Oda, Y.; Gillam, E.M.; Guengerich, F.P.; Inoue, K. Metabolic activatiom of polycyclic aromatic hydrocarbons and other procarcinogens by cytochromes P450 1A1 and P450 1B1 allelic variants and other human cytochromes P450 in Salemonella typhimurium NM2009. Drug Metab. Dispos. 2001, 29, 1176–1182. [Google Scholar] [PubMed]
- Anzenbacher, P.; Anzenbacherová, E. Cytochromes P450 and metabolism of xenobiotics. Cell. Mol. Life Sci. 2001, 58, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Paine, M.F.; Hart, H.L.; Ludington, S.S.; Haining, R.L.; Rettie, A.E.; Zeldin, D.C. The human intestinal cytochrome P450 “pie”. Drug Metab. Dispos. 2006, 34, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, S.; Ikemoto, K.; Takahashi, J.; Miki, H.; Namba, M.; Asahi, S. Establishment of the transformants expressing human cytochrome P450 subtypes in HepG2, and their applications on drug metabolism and toxicology. Toxicol. In Vitro 2001, 15, 245–256. [Google Scholar] [CrossRef]
- Rueff, J.; Chiapella, C.; Chipman, J.K.; Darroudi, F.; Silva, I.D.; Duverger-van Bogaert, M.; Fonti, E.; Glatt, H.R.; Isern, P.; Laires, A.; et al. Development and validation of alternative metabolic systems for mutagenicity testing in short-term assays. Mutat. Res. 1996, 353, 151–176. [Google Scholar] [CrossRef]
- Obach, R.S. Inhibition of human cytochrome P450 enzymes by constituents of St. John’s Wort, an herbal preparation used in the treatment of depression. J. Pharmacol. Exp. Ther. 2000, 294, 88–95. [Google Scholar] [PubMed]
- Fantini, M.; Gianni, L.; Santelmo, C.; Drudi, F.; Castellani, C.; Affatato, A.; Nicolini, M.; Ravaioli, A. Lipoplatin treatment in lung and breast cancer. Chemother. Res. Pract. 2011, 2001, 125192. [Google Scholar] [CrossRef] [PubMed]
- Hunakova, L.; Gronesova, P.; Horvathova, E.; Chalupa, I.; Cholujova, D.; Duraj, J.; Sedlak, J. Modulation of cisplatin sensitivity in human ovarian carcinoma A2780 and SKOV3 cell lines by sulforaphane. Toxicol. Lett. 2014, 230, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Jendželovský, R.; Jendželovská, Z.; Hiľovská, L.; Kovaľ, J.; Mikeš, J.; Fedoročko, P. Proadifen sensitizes resistant ovarian adenocarcinoma cells to cisplatin. Toxicol. Lett. 2016, 243, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Koyabu, N.; Morimoto, S.; Shoyama, Y.; Ohtani, H.; Sawada, Y. Functional induction and de-induction of P-glycoprotein by St. John’s wort and its ingredients in a human colon adenocarcinoma cell line. Drug Metab. Dispos. 2004, 33, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Izzo, A.A. Herb-drug interactions with St John’s wort (Hypericum perforatum): An update on clinical observations. AAPS J. 2009, 11, 710–727. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.L. P-glycoprotein Inhibition for Optimal Drug Delivery. Drug Target Insights 2013, 7, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gibalová, L.; Sereš, M.; Rusnák, A.; Ditte, P.; Labudová, M.; Uhrík, B.; Pastorek, J.; Sedlák, J.; Breier, A.; Sulová, Z. P-glycoprotein depresses cisplatin sensitivity in L1210 cells by inhibiting cisplatin-induced caspase-3 activation. Toxicol. In Vitro 2012, 26, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Al-Abd, A.M.; El-Dine, R.S.; El-Halawany, A.M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: A review. J. Adv. Res. 2015, 6, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Fattah, F.; Lee, E.H.; Weisensel, N.; Wang, Y.; Lichter, N.; Hendrickson, E.A. Ku regulates the non-homologous end joining pathway choice of DNA double-strand break repair in human somatic cells. PLoS Genet. 2010, 6, e1000855. [Google Scholar] [CrossRef] [PubMed]
- Moggs, J.G.; Yarema, K.J.; Essigmann, J.M.; Wood, R.D. Analysis of incision sites produced by human cell extracts and purified proteins during nucleotide excision repair of a 1,3-intrastrand d(GpTpG)-cisplatin adduct. J. Biol. Chem. 1996, 271, 7177–7186. [Google Scholar] [PubMed]
- Wu, Q.; Christensen, L.A.; Legerski, R.J.; Vasquez, K.M. Mismatch repair participates in error-free processing of DNA interstrand crosslinks in human cells. EMBO Rep. 2005, 6, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Miadokova, E.; Chalupa, I.; Vlckova, V.; Sevcovicova, A.; Nadova, S.; Kopaskova, M.; Hercegova, A.; Gasperova, P.; Alfoldiova, L.; Komjatiova, M.; et al. Genotoxicity and antigenotoxicity evaluation of non-photoactivated hypericin. Phytother. Res. 2010, 24, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Kolman, A.; Kleman, M.; Bohušová, T.; Gustafsson, J.A.; Övervik, E. Mutagenic and cytotoxic action of heated pork meat extracts in human diploid fibroblasts. Mutagenesis 1992, 7, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, Y.; Costa, S.; Collins, A.R.; Azqueta, A. DNA Damage DNA repair and cytotoxicity: Hedgehogs are not always dead. Mutagenesis 2013, 28, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Horvathova, E.; Navarova, J.; Galova, E.; Sevcovicova, A.; Chodakova, L.; Snahnicanova, Z.; Melusova, M.; Kozics, K.; Slamenova, D. Assessment of antioxidative, chelating, and DNA-protective effects of selected essential oil components (eugenol, carvacrol, thymol, borneol, eucalyptol) of plants and intact Rosmarinus officinalis oil. J. Agric. Food Chem. 2014, 62, 6632–6639. [Google Scholar] [CrossRef] [PubMed]
- Einset, J.; Collins, A.R. DNA repair after X-irradiation: Lessons from plants. Mutagenesis 2015, 30, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.; Bischoff, R.; Stegmüller, S.; Cartus, A.; Schrenk, D. Comparative investigation of the mutagenicity of propenylic and allylic asarone isomers in the Ames fluctuation assay. Mutagenesis 2016, 31, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are not available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HF/B(a)P Concentration (µM) | Number of Aberrant Metaphases | Number of Chromosome Aberrations | Total Number of CA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chromatid | Isochromatid | Exchange | |||||||||
| g | b/f | g | b/f | ring | dic | qr | tr | dmin | |||
| HepG2 | |||||||||||
| IC | 4 | 1 | 3 | - | - | - | - | - | - | 1 | 4 |
| SC | 3 | 1 | 3 | - | - | - | - | - | - | - | 3 |
| HF 0.75 | 2 | 1 | 2 | - | - | - | - | - | - | - | 2 |
| HF 1.5 | 2 | - | 2 | - | - | - | - | - | - | - | 2 |
| PC B(a)P 1.7 | 35 *** | - | 33 | 1 | 5 | - | 1 | - | - | - | 39 *** |
| HF 0.75 + B(a)P 1.7 | 8 ### | 1 | 7 | - | 1 | - | - | - | - | - | 8 ### |
| HF 1.5 + B(a)P 1.7 | 8 ### | 1 | 8 | - | - | - | 1 | - | - | - | 9 ### |
| HF/cisPt Concentration (µM) | Number of Aberrant Metaphases | Number of Chromosome Aberrations | Total Number of CA | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chromatid | Isochromatid | Exchange | |||||||||
| g | b/f | g | b/f | ring | dic | qr | tr | dmin | |||
| HepG2 | |||||||||||
| IC | 2 | 1 | 2 | 1 | - | - | - | - | - | - | 2 |
| SC | 3 | 1 | 2 | - | - | - | 1 | - | - | - | 3 |
| HF 0.75 | 1 | 1 | 1 | - | - | - | - | - | - | - | 1 |
| HF 1.5 | 3 | - | 2 | - | 1 | - | - | - | - | - | 3 |
| PC cisPt 0.5 | 38 *** | 2 | 36 | - | 8 | - | 2 | 1 | - | - | 47 *** |
| HF 0.75+ cisPt 0.5 | 10 *,### | 2 | 5 | - | 4 | - | 2 | - | - | - | 11 *,### |
| HF 1.5 + cisPt 0.5 | 14 **,### | 2 | 10 | - | 3 | - | 1 | - | - | - | 14 **,### |
| VH10 | |||||||||||
| IC | 2 | - | 2 | - | - | - | - | - | - | - | 2 |
| SC | 2 | 1 | 1 | 1 | - | - | 1 | - | - | - | 2 |
| HF 0.75 | 1 | - | 1 | - | - | - | - | - | - | - | 1 |
| HF 1.5 | 1 | - | - | - | - | - | 1 | - | - | - | 1 |
| PC cisPt 0.75 | 33 *** | 2 | 26 | - | 9 | - | - | 4 | 5 | - | 44 *** |
| HF 0.75 + cisPt 0.75 | 12 **,### | 1 | 8 | - | 3 | - | 1 | 1 | 1 | - | 14 ***,### |
| HF 1.5 + cisPt 0.75 | 11 **,### | 3 | 8 | - | - | - | 1 | - | 2 | - | 11 **,### |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imreova, P.; Feruszova, J.; Kyzek, S.; Bodnarova, K.; Zduriencikova, M.; Kozics, K.; Mucaji, P.; Galova, E.; Sevcovicova, A.; Miadokova, E.; et al. Hyperforin Exhibits Antigenotoxic Activity on Human and Bacterial Cells. Molecules 2017, 22, 167. https://doi.org/10.3390/molecules22010167
Imreova P, Feruszova J, Kyzek S, Bodnarova K, Zduriencikova M, Kozics K, Mucaji P, Galova E, Sevcovicova A, Miadokova E, et al. Hyperforin Exhibits Antigenotoxic Activity on Human and Bacterial Cells. Molecules. 2017; 22(1):167. https://doi.org/10.3390/molecules22010167
Chicago/Turabian StyleImreova, Petronela, Jana Feruszova, Stanislav Kyzek, Kristina Bodnarova, Martina Zduriencikova, Katarina Kozics, Pavel Mucaji, Eliska Galova, Andrea Sevcovicova, Eva Miadokova, and et al. 2017. "Hyperforin Exhibits Antigenotoxic Activity on Human and Bacterial Cells" Molecules 22, no. 1: 167. https://doi.org/10.3390/molecules22010167
APA StyleImreova, P., Feruszova, J., Kyzek, S., Bodnarova, K., Zduriencikova, M., Kozics, K., Mucaji, P., Galova, E., Sevcovicova, A., Miadokova, E., & Chalupa, I. (2017). Hyperforin Exhibits Antigenotoxic Activity on Human and Bacterial Cells. Molecules, 22(1), 167. https://doi.org/10.3390/molecules22010167