
Multivalent Carbohydrate-Lectin Interactions: How Synthetic Chemistry Enables Insights into Nanometric Recognition



Abstract
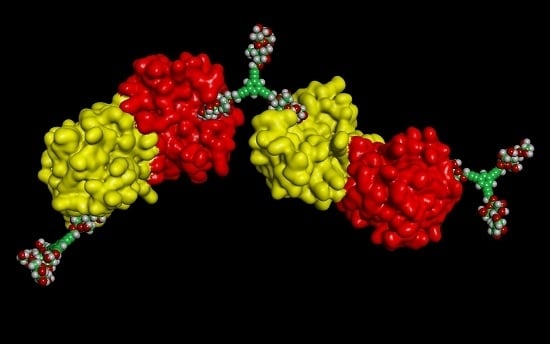
:
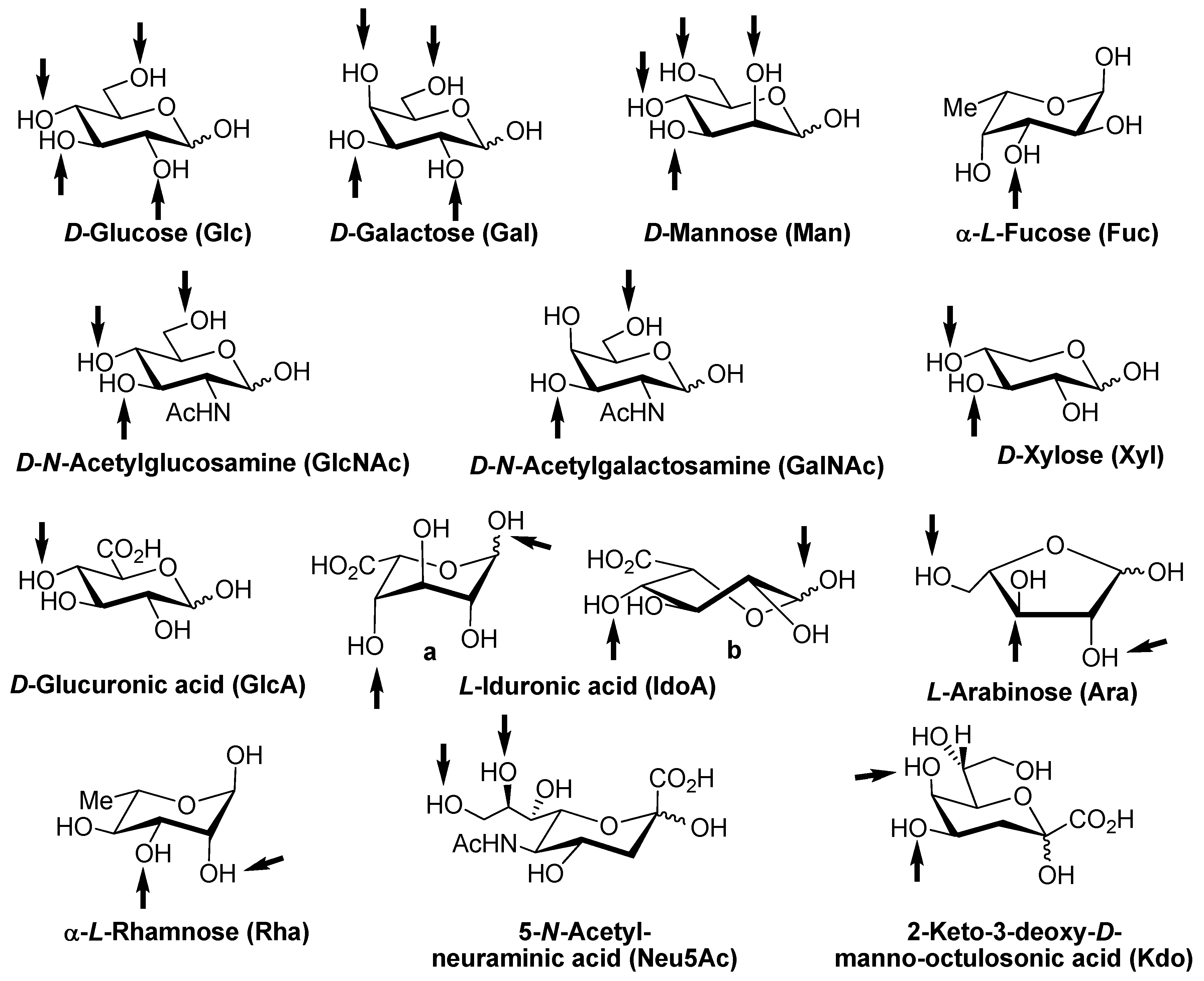
1. The Third Alphabet of Life
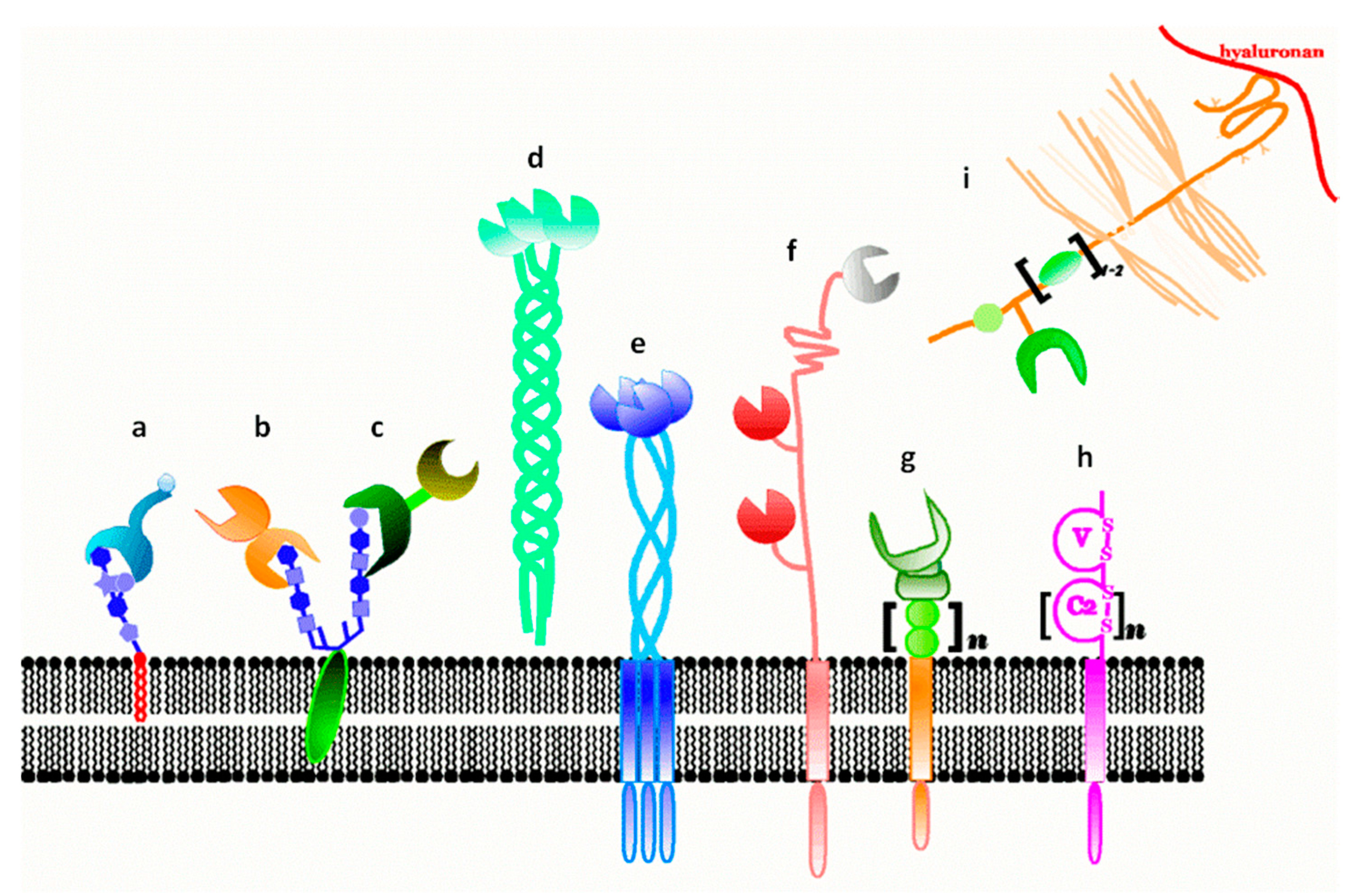
2. Lectins: Translators of Sugar-Encoded Information
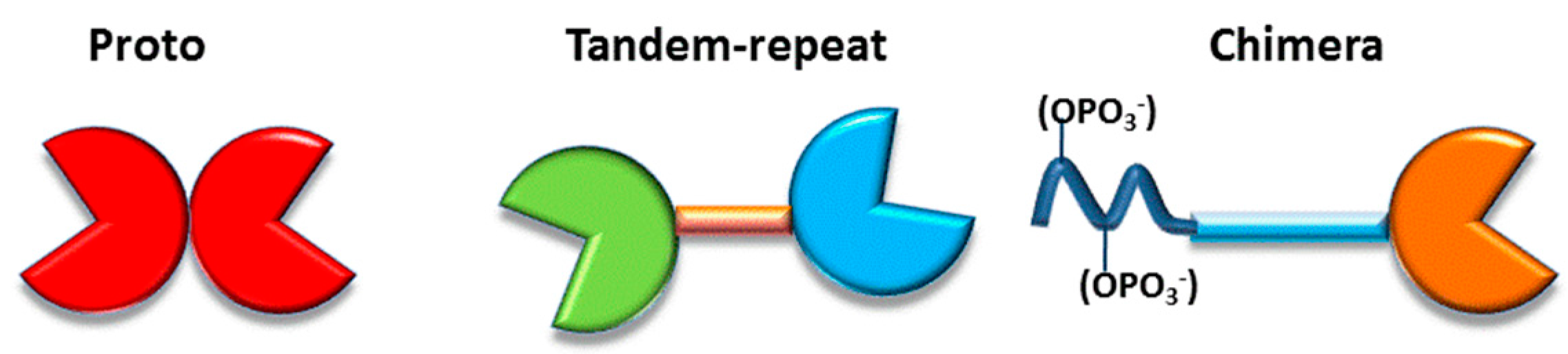
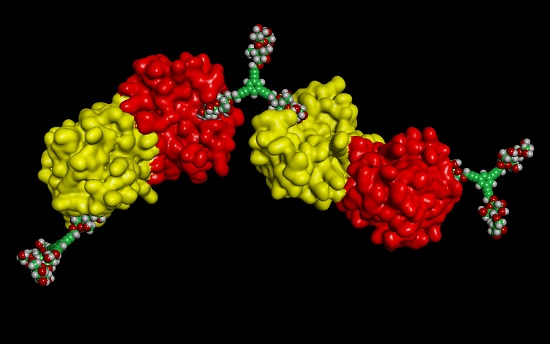
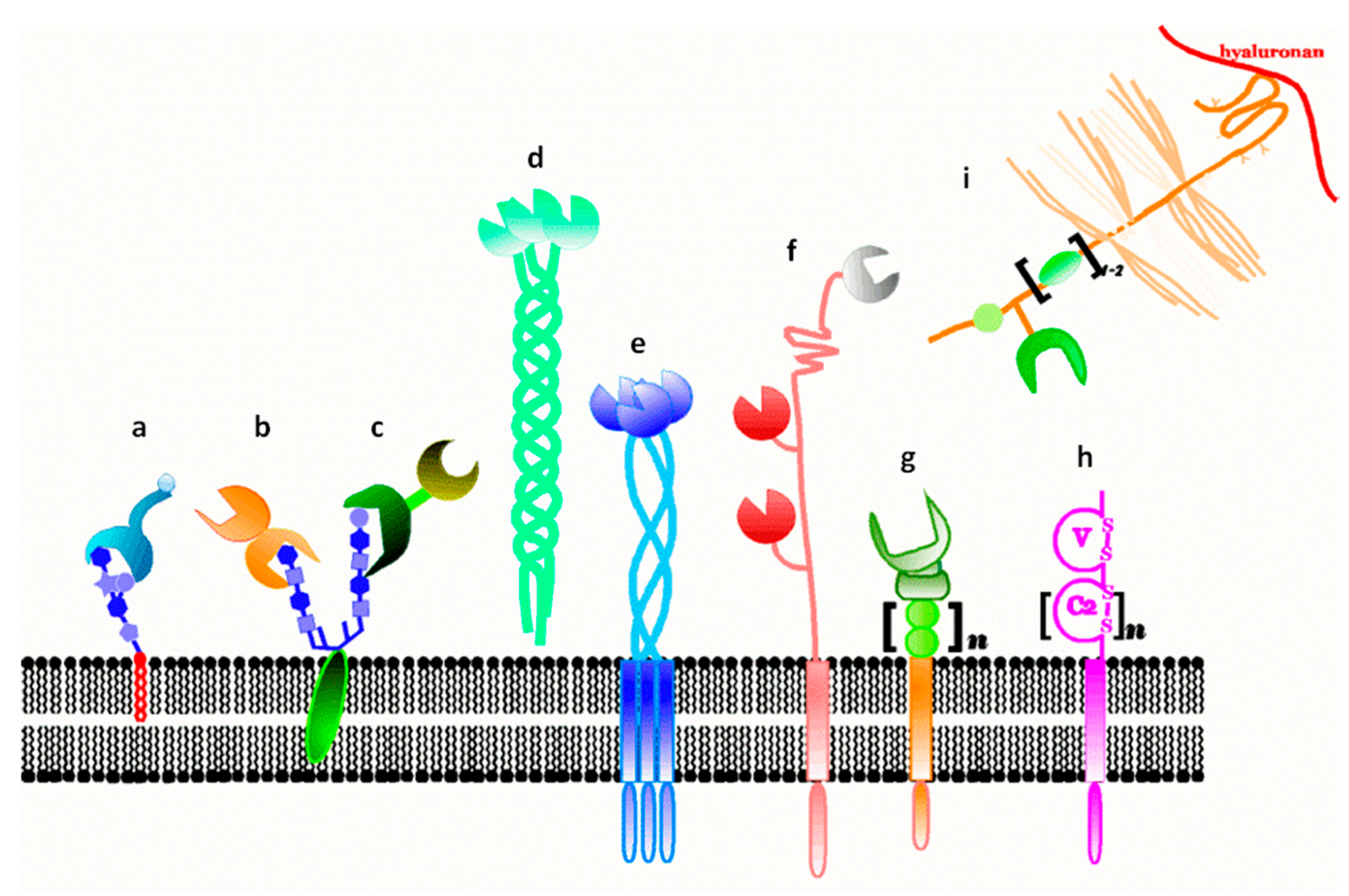
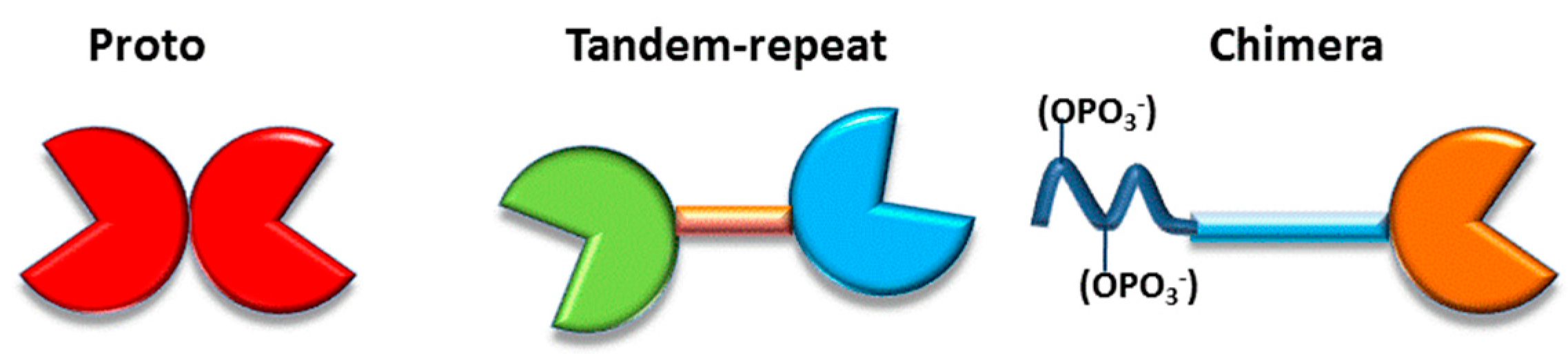
3. Galectins: Definition and Structures
- (i)
- to have a CRD with a β-sandwich fold, first described in this field for the leguminous lectin concanavalin A and illustrated in the Gallery of Lectins [27], is a compact structural platform consisting—in the case of human galectin-1—of two antiparallel β-sheets of five to six strands, respectively,
- (ii)
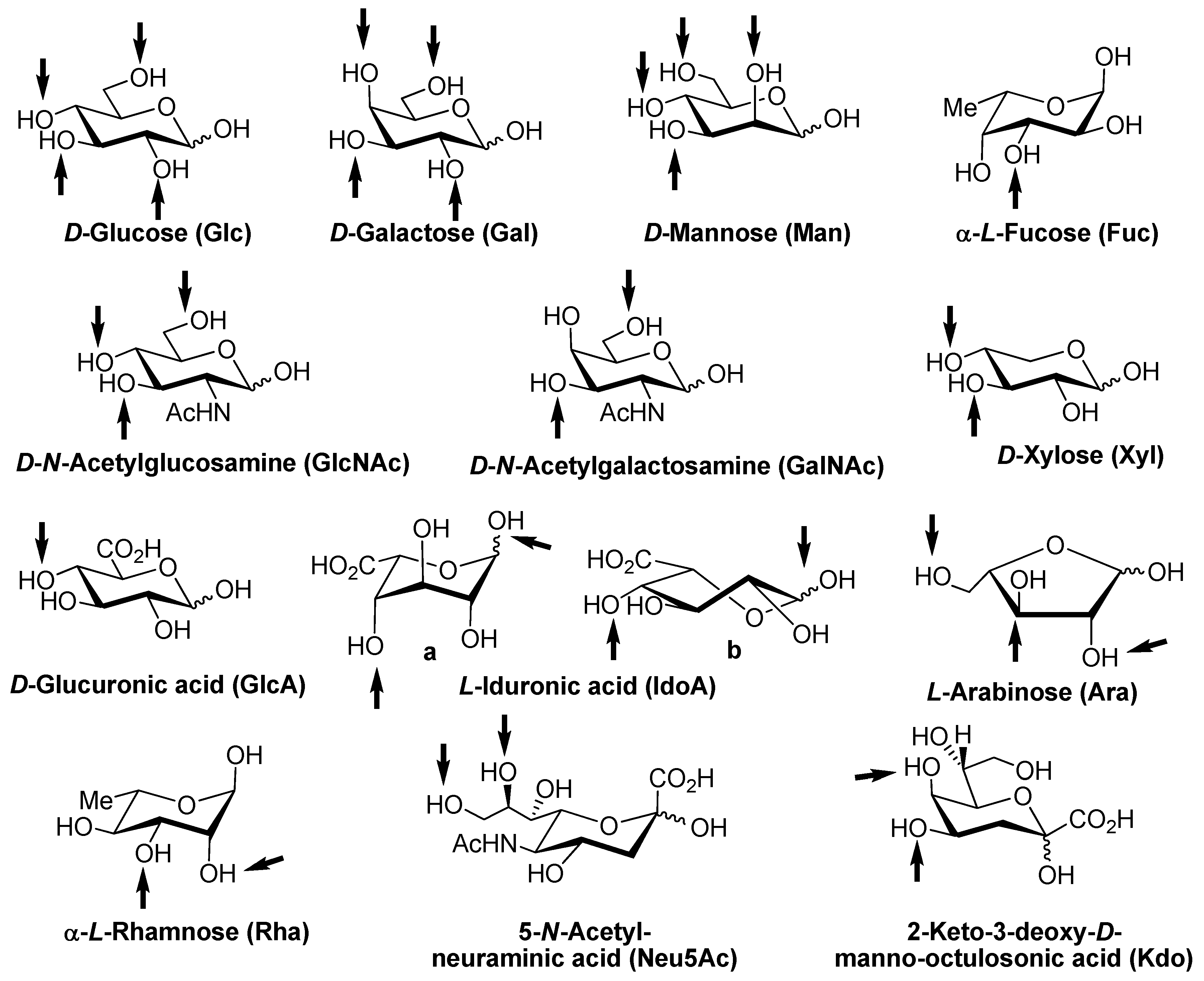
- to bind compounds from the group of β-galactosides and
- (iii)
4. Tailoring Monovalent Ligands for Selectivity
4.1. Modifications of the Aglycone
4.2. Modifications at the Glycosidic Linkage
4.3. Modification at Bioinspired Positions
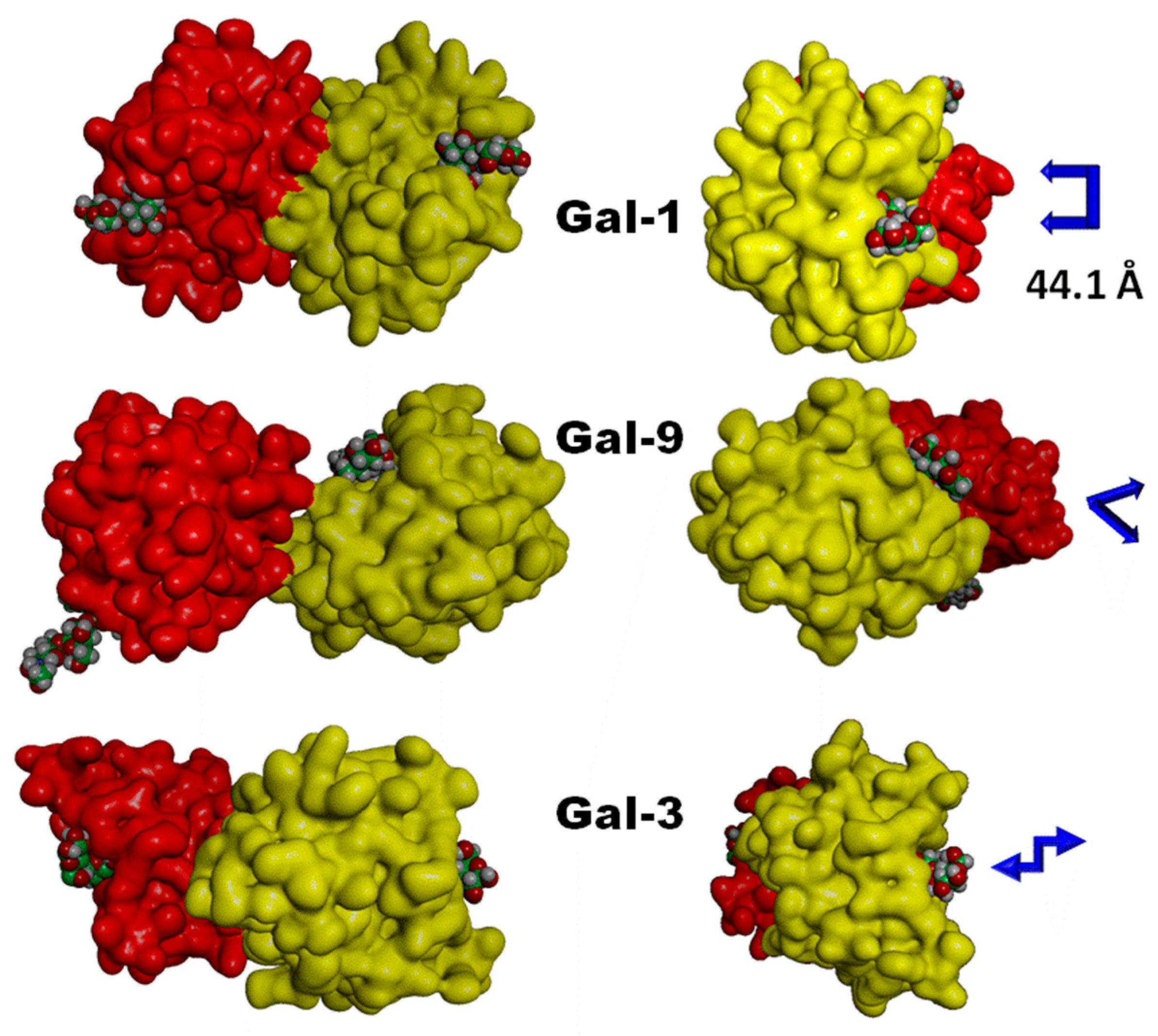
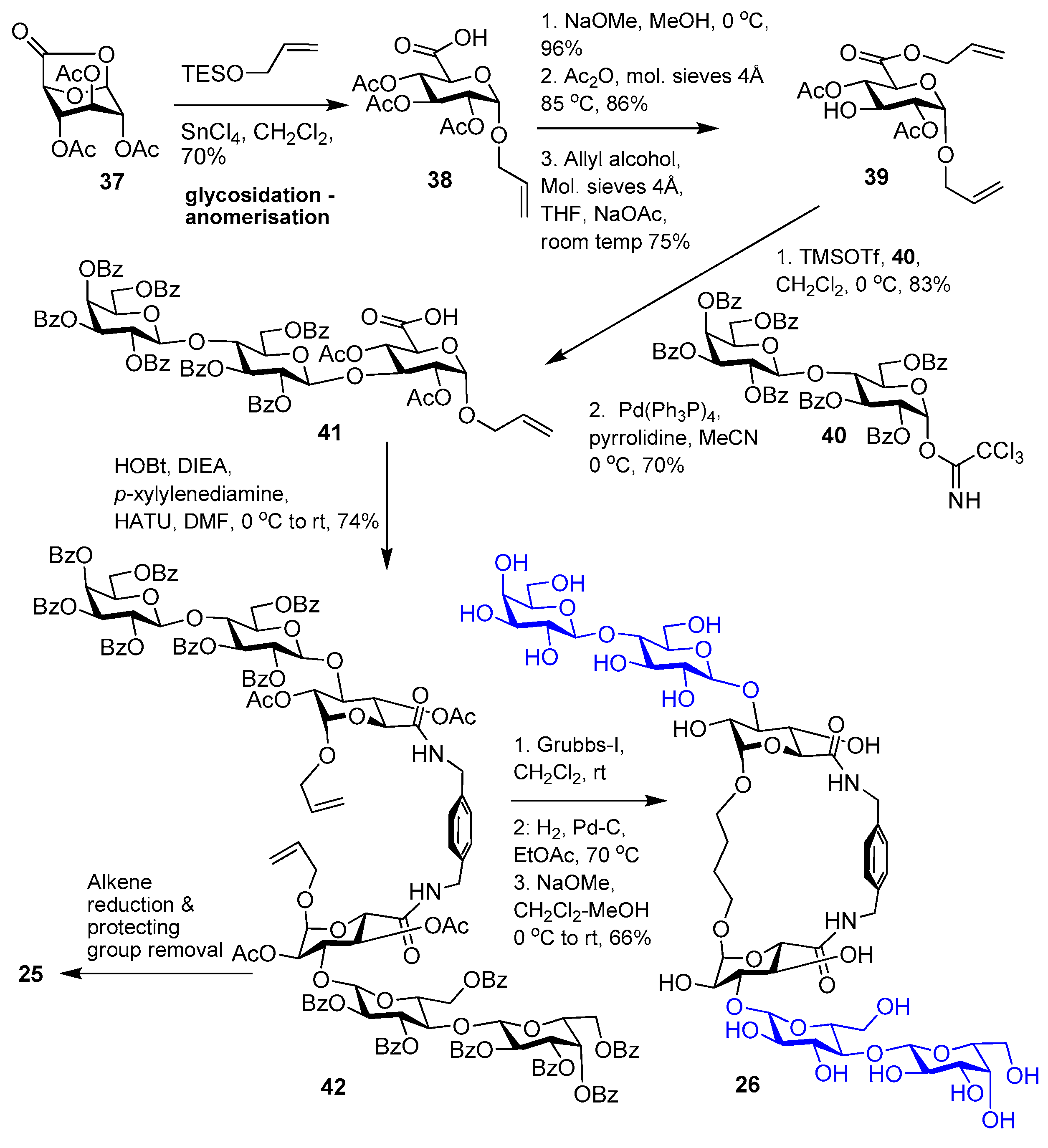
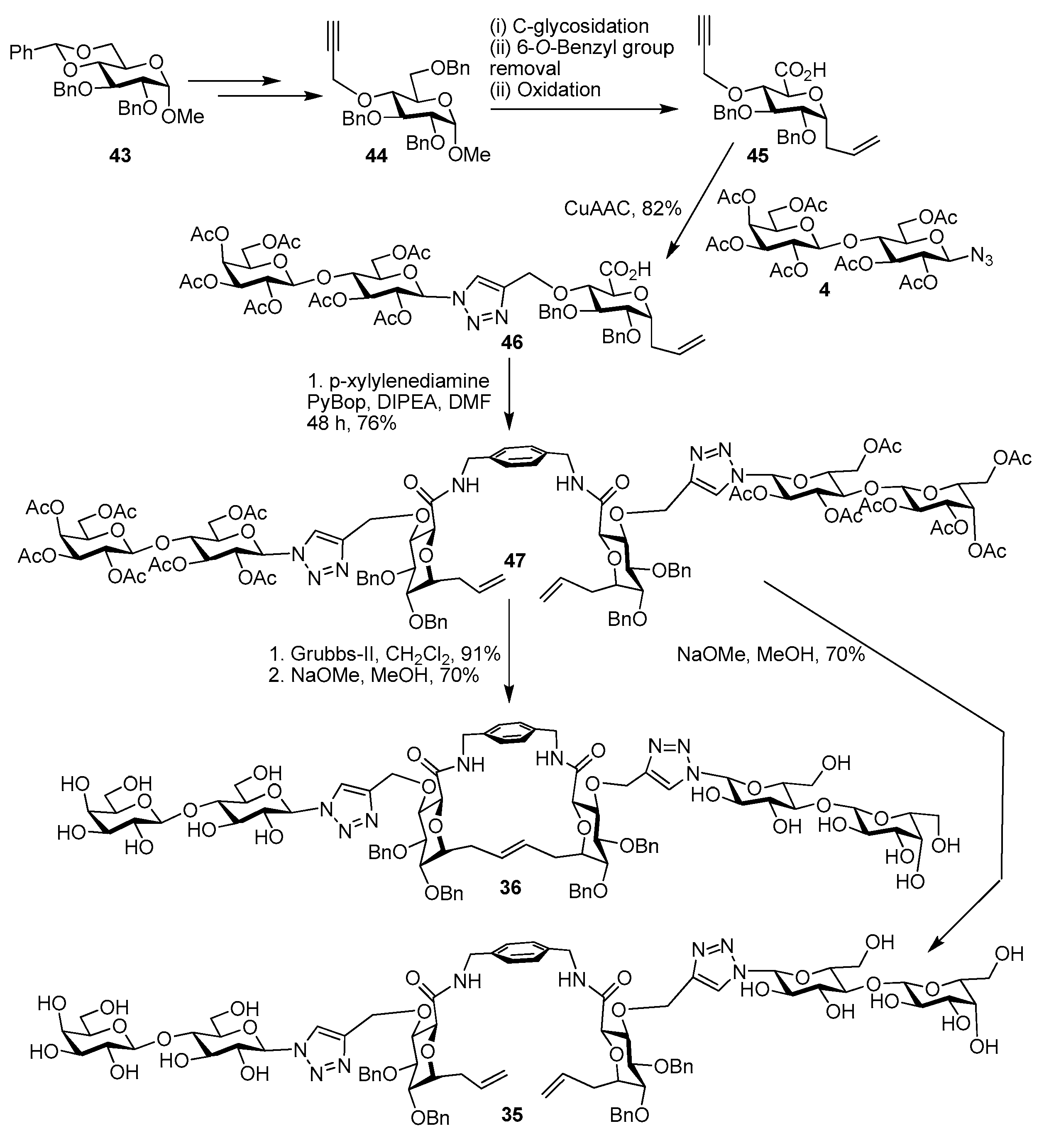
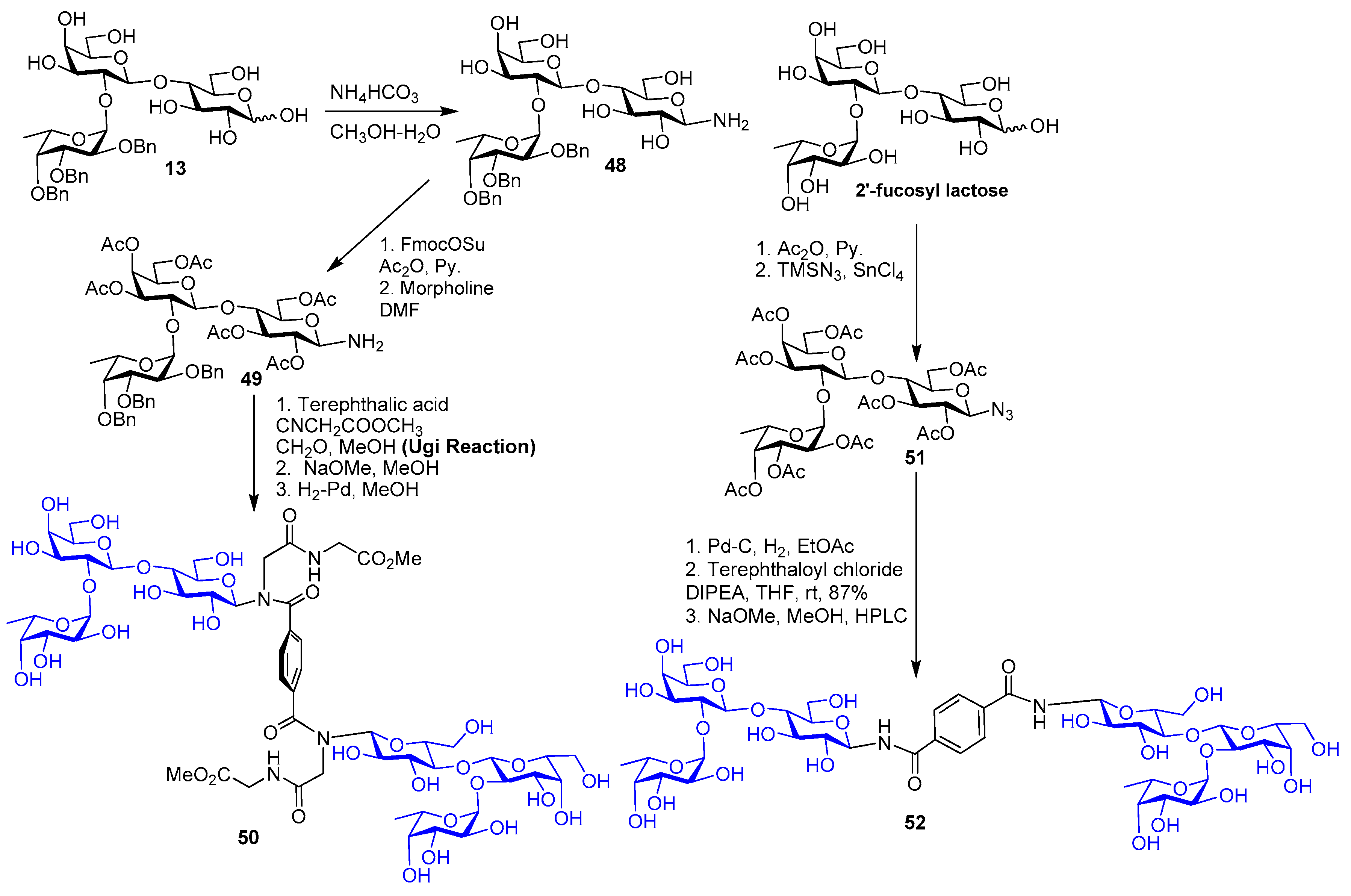
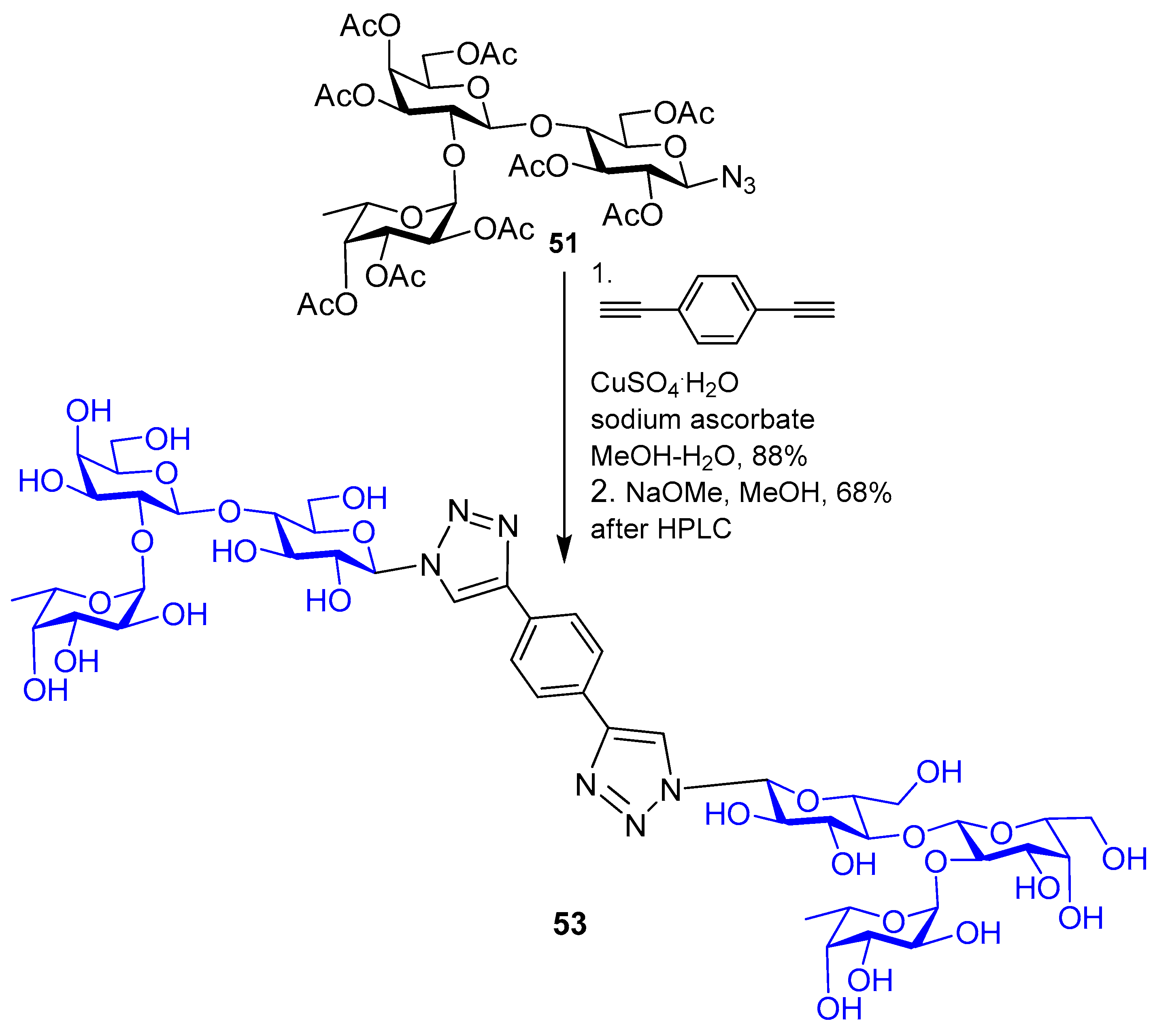
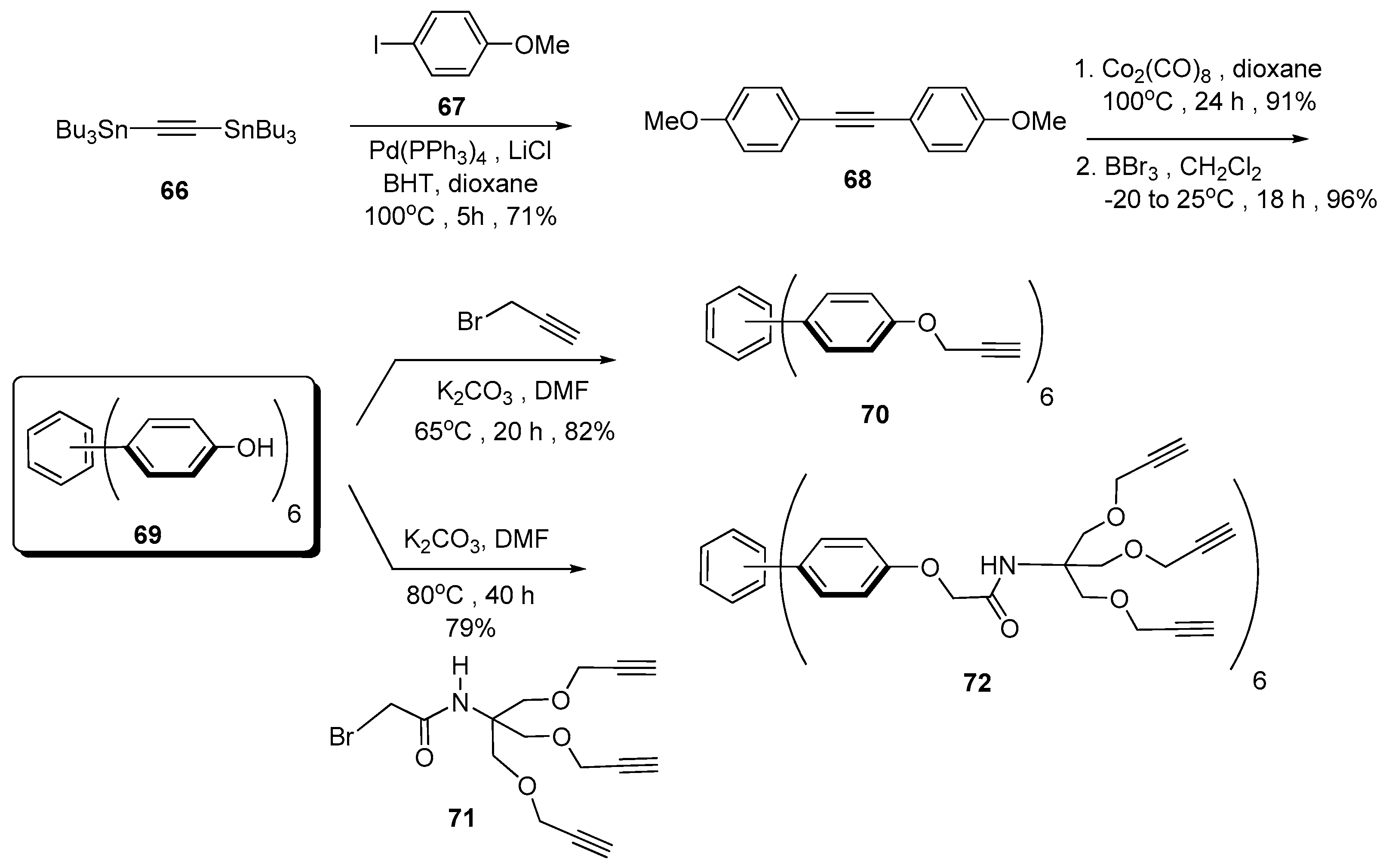
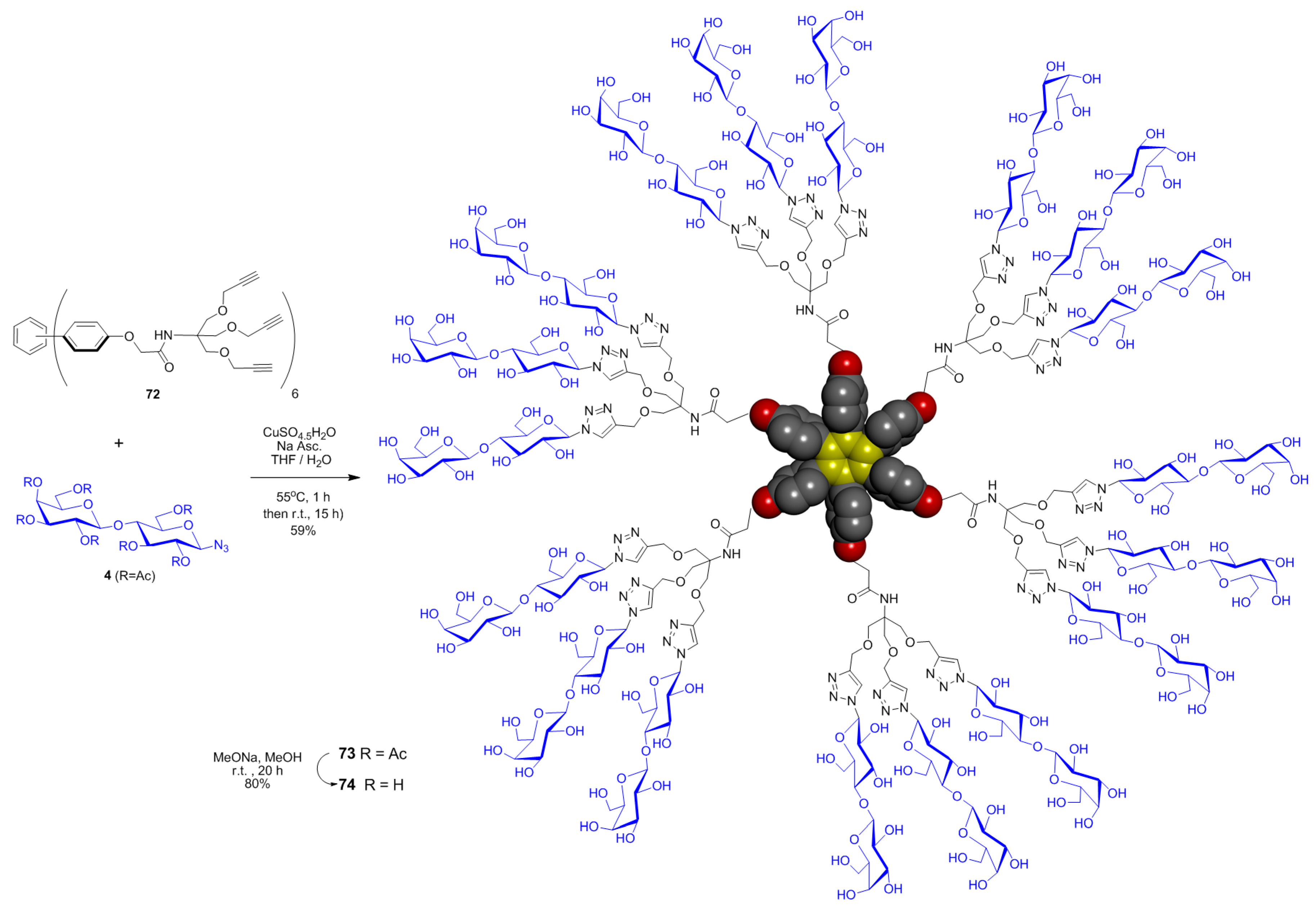
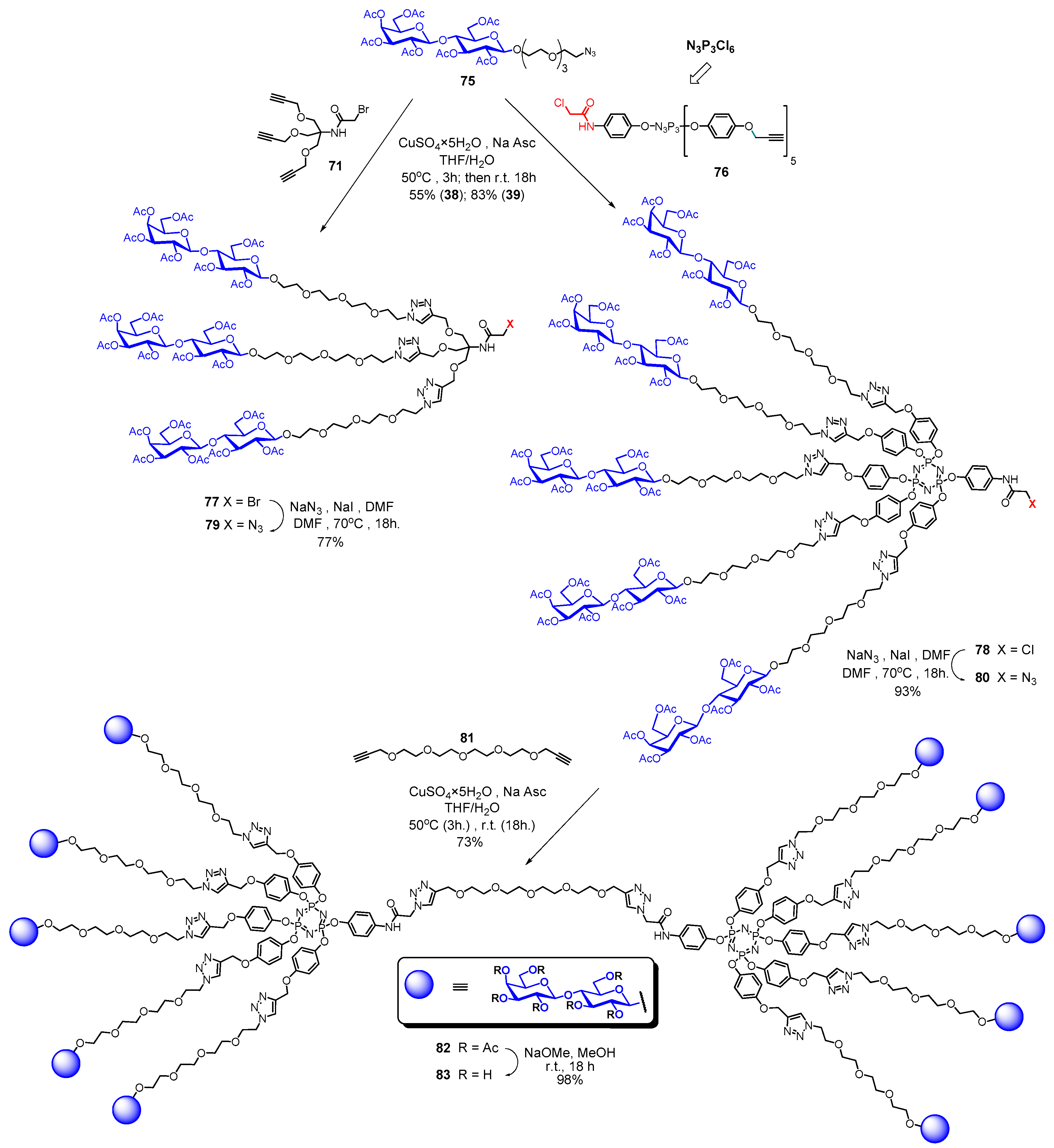
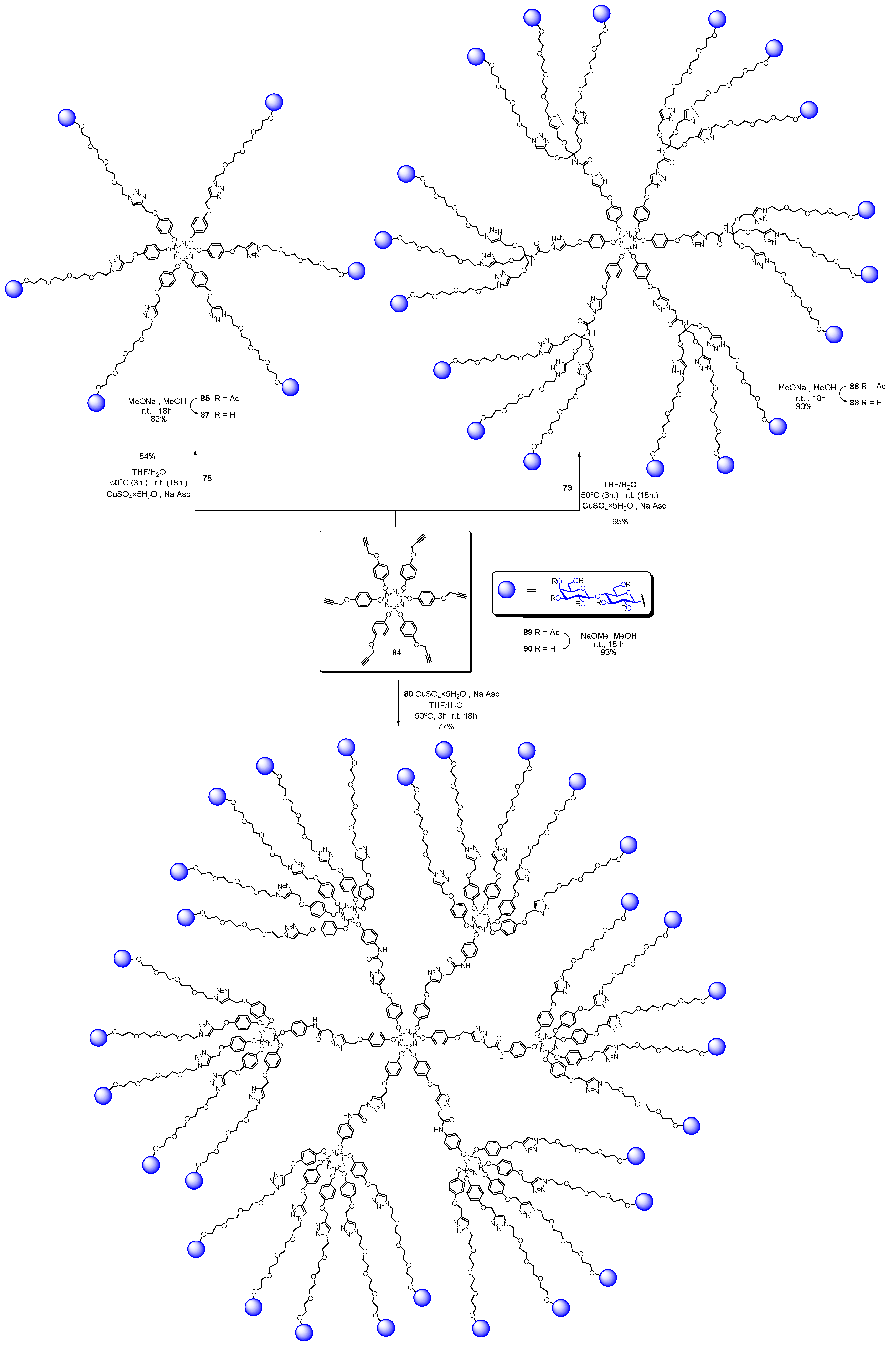
5. Tailoring Spatial Aspects for Selectivity
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sharon, N. Complex Carbohydrates. Their Chemistry, Biosynthesis, and Functions; Addison-Wesley Publ. Co.: Reading, MA, USA, 1975. [Google Scholar]
- Reuter, G.; Gabius, H.-J. Eukaryotic glycosylation: Whim of nature or multipurpose tool? Cell. Mol. Life Sci. 1999, 55, 368–422. [Google Scholar] [CrossRef] [PubMed]
- Zuber, C.; Roth, J. N-Glycosylation. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 87–110. [Google Scholar]
- Patsos, G.; Corfield, A. O-Glycosylation: Structural diversity and function. In The Sugar Code. Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 111–137. [Google Scholar]
- Kopitz, J. Glycolipids. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 177–198. [Google Scholar]
- Buddecke, E. Proteoglycans. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 199–216. [Google Scholar]
- Merzendorfer, H. Chitin: Structure, function and metabolism. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 217–229. [Google Scholar]
- Corfield, A.P.; Berry, M. Glycan variation and evolution in the eukaryotes. Trends Biochem. Sci. 2015, 40, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.-J. The magic of the sugar code. Trends Biochem. Sci. 2015, 40. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W.; Wu, G. The multi-tasked life of GM1 ganglioside, a true factotum of nature. Trends Biochem. Sci. 2015, 40, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Schengrund, C.-L. Gangliosides: Glycosphingolipids essential for normal neural development and function. Trends Biochem. Sci. 2015, 40, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Tavares, E.Q.; Buckeridge, M.S. Do plant cell walls have a code? Plant Sci. 2015, 241, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.Y.; Tang, C.M.; Exley, R.M. Sugar coating: Bacterial protein glycosylation and host-microbe interactions. Trends Biochem. Sci. 2015, 40, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Winterburn, P.J.; Phelps, C.F. The significance of glycosylated proteins. Nature 1972, 236, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Rüdiger, H.; Gabius, H.-J. The biochemical basis and coding capacity of the sugar code. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 3–13. [Google Scholar]
- Nishimura, S.-I. Toward automated glycan analysis. Adv. Carbohydr. Chem. Biochem. 2011, 65, 219–271. [Google Scholar] [PubMed]
- Montreuil, J. The history of glycoprotein research, a personal view. In Glycoproteins; Montreuil, J., Vliegenthart, J.F.G., Schachter, H., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; pp. 1–12. [Google Scholar]
- Pérez, S.; Imberty, A.; Carver, J.P. Molecular modeling: An essential component in the structure determination of oligosaccharides and polysaccharides. Adv. Comput. Biol. 1994, 1, 147–202. [Google Scholar]
- Von der Lieth, C.-W.; Siebert, H.-C.; Kozár, T.; Burchert, M.; Frank, M.; Gilleron, M.; Kaltner, H.; Kayser, G.; Tajkhorshid, E.; Bovin, N.V.; et al. Lectin ligands: New insights into their conformations and their dynamic behavior and the discovery of conformer selection by lectins. Acta Anat. 1998, 161, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Bush, C.A.; Martin-Pastor, M.; Imberty, A. Structure and conformation of complex carbohydrates of glycoproteins, glycolipids, and bacterial polysaccharides. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 269–293. [Google Scholar] [CrossRef] [PubMed]
- Imberty, A.; Pérez, S. Structure, conformation, and dynamics of bioactive oligosaccharides: Theoretical approaches and experimental validations. Chem. Rev. 2000, 100, 4567–4588. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Kozár, T.; Kojima, S.; Unverzagt, C.; Gabius, H.-J. From structural to functional glycomics: Core substitutions as molecular switches for shape and lectin affinity of N-glycans. Biol. Chem. 2009, 390, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.V.; André, S.; Gabius, H.-J. The third dimension of reading the sugar code by lectins: Design of glycoclusters with cyclic scaffolds as tools with the aim to define correlations between spatial presentation and activity. Molecules 2013, 18, 4026–4053. [Google Scholar] [PubMed]
- André, S.; Kaltner, H.; Manning, J.C.; Murphy, P.V.; Gabius, H.-J. Lectins: Getting familiar with translators of the sugar code. Molecules 2015, 20, 1788–1823. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H. Bifunctional properties of lectins: Lectins redefined. Trends Biochem. Sci. 1988, 13, 480–482. [Google Scholar] [CrossRef]
- Solís, D.; Bovin, N.V.; Davis, A.P.; Jiménez-Barbero, J.; Romero, A.; Roy, R.; Smetana, K., Jr.; Gabius, H.-J. A guide into glycosciences: How chemistry, biochemistry and biology cooperate to crack the sugar code. Biochim. Biophys. Acta 2015, 1850, 186–235. [Google Scholar] [CrossRef] [PubMed]
- Quiocho, F.A. Carbohydrate-binding proteins: Tertiary structures and protein-sugar interactions. Annu. Rev. Biochem. 1986, 55, 287–315. [Google Scholar] [CrossRef] [PubMed]
- Rini, J.M. Lectin structure. Annu. Rev. Biophys. Biomol. Struct. 1995, 24, 551–577. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.-J.; André, S.; Jiménez-Barbero, J.; Romero, A.; Solís, D. From lectin structure to functional glycomics: Principles of the sugar code. Trends Biochem. Sci. 2011, 36, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.-J.; Kaltner, H.; Kopitz, J.; André, S. The glycobiology of the CD system: A dictionary for translating marker designations into glycan/lectin structure and function. Trends Biochem. Sci. 2015, 40, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W.; Wu, G.; André, S.; Bleich, D.; Huet, G.; Kaltner, H.; Kopitz, J.; Gabius, H.-J. Beyond glycoproteins as galectin counterreceptors: Tumor/effector T cell growth control via ganglioside GM1. Ann. N. Y. Acad. Sci. 2012, 1253, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.-J.; Manning, J.C.; Kopitz, J.; André, S.; Kaltner, H. Sweet complementarity: The functional pairing of glycans with lectins. Cell Mol. Life Sci. 2016, 73, 1989–2016. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Sanchez-Ruderisch, H.; Nakagawa, H.; Buchholz, M.; Kopitz, J.; Forberich, P.; Kemmner, W.; Böck, C.; Deguchi, K.; Detjen, K.M.; et al. Tumor suppressor p16INK4a: Modulator of glycomic profile and galectin-1 expression to increase susceptibility to carbohydrate-dependent induction of anoikis in pancreatic carcinoma cells. FEBS J. 2007, 274, 3233–3256. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Eriksson, H.; Manning, J.C.; Detjen, K.M.; André, S.; Nishimura, S.-I.; Lehtiö, J.; Gabius, H.-J. Tumor suppressor p16INK4a: Anoikis-favoring decrease in N/O-glycan/cell surface sialylation by downregulation of enzymes in sialic acid biosynthesis in tandem in a pancreatic carcinoma model. FEBS J. 2012, 279, 4062–4080. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ruderisch, H.; Fischer, C.; Detjen, K.M.; Welzel, M.; Wimmel, A.; Manning, J.C.; André, S.; Gabius, H.-J. Tumor suppressor p16INK4a: Downregulation of galectin-3, an endogenous competitor of the pro-anoikis effector galectin-1, in a pancreatic carcinoma model. FEBS J. 2010, 277, 3552–3563. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.K.; Oscarson, S.; Roy, R.; Das, S.K.; Page, D.; Macaluso, F.; Brewer, C.F. Thermodynamic, kinetic, and electron microscopy studies of concanavalin A and Dioclea grandiflora lectin cross-linked with synthetic divalent carbohydrates. J. Biol. Chem. 2005, 280, 8640–8646. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.K.; Gabius, H.-J.; André, S.; Kaltner, H.; Lensch, M.; Brewer, C.F. Galectins bind to the multivalent glycoprotein asialofetuin with enhanced affinities and a gradient of decreasing binding constants. Biochemistry 2005, 44, 12564–12571. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.-J. Animal and human lectins. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 317–328. [Google Scholar]
- Cooper, D.N.W. Galectinomics: Finding themes in complexity. Biochim. Biophys. Acta 2002, 1572, 209–231. [Google Scholar] [CrossRef]
- Gready, J.N.; Zelensky, A.N. Routes in lectin evolution: Case study on the C-type lectin-like domains. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 329–346. [Google Scholar]
- Cao, H.; Crocker, P.R. Evolution of CD33-related siglecs: Regulating host immune functions and escaping pathogen exploitation? Immunology 2011, 132, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mitra, N.; Cruz, P.; Deng, L.; Varki, N.; Angata, T.; Green, E.D.; Mullikin, J.; Hayakawa, T.; Varki, A. Evolution of siglec-11 and siglec-16 genes in hominins. Mol. Biol. Evol. 2012, 29, 2073–2086. [Google Scholar] [CrossRef] [PubMed]
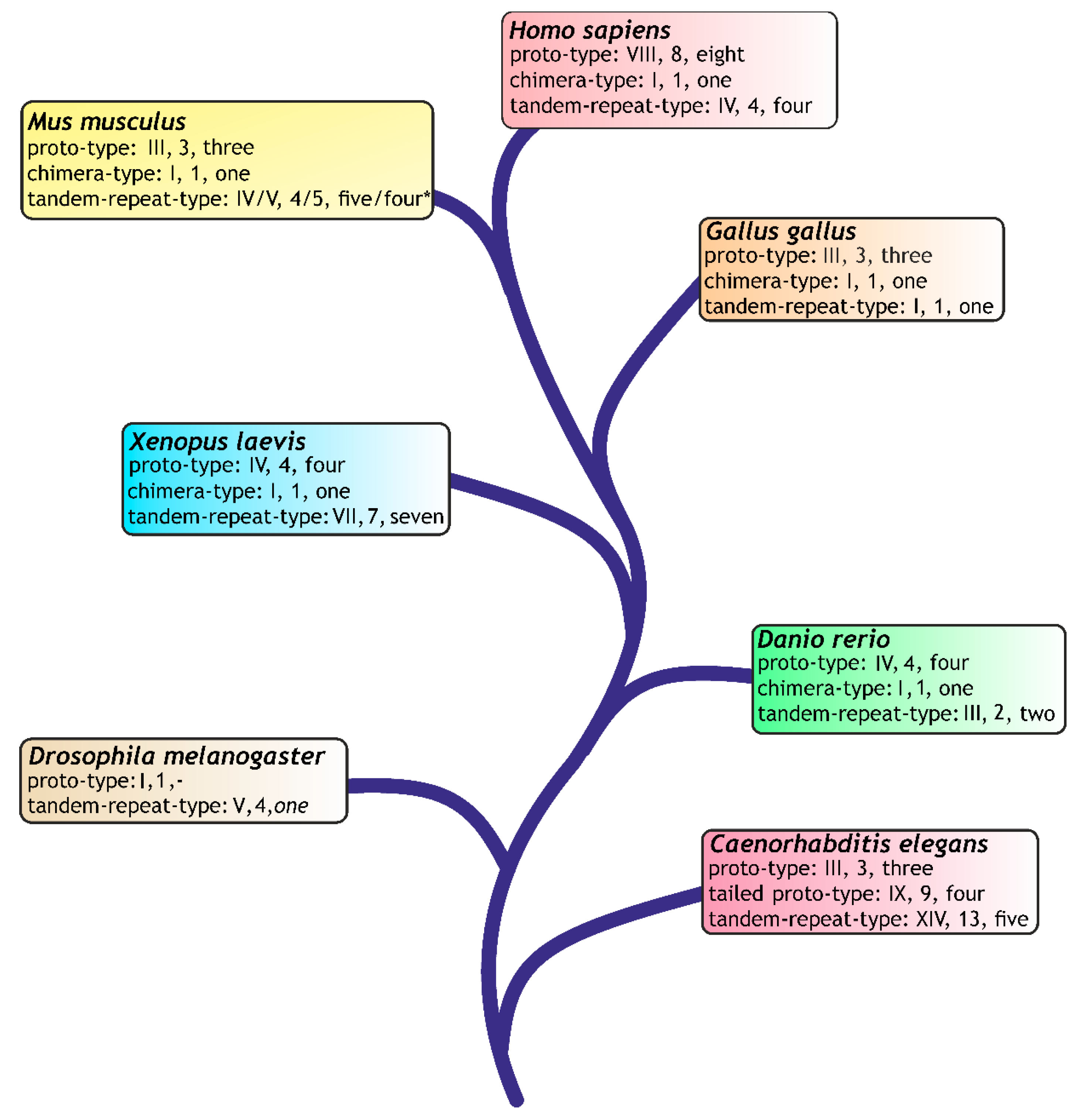
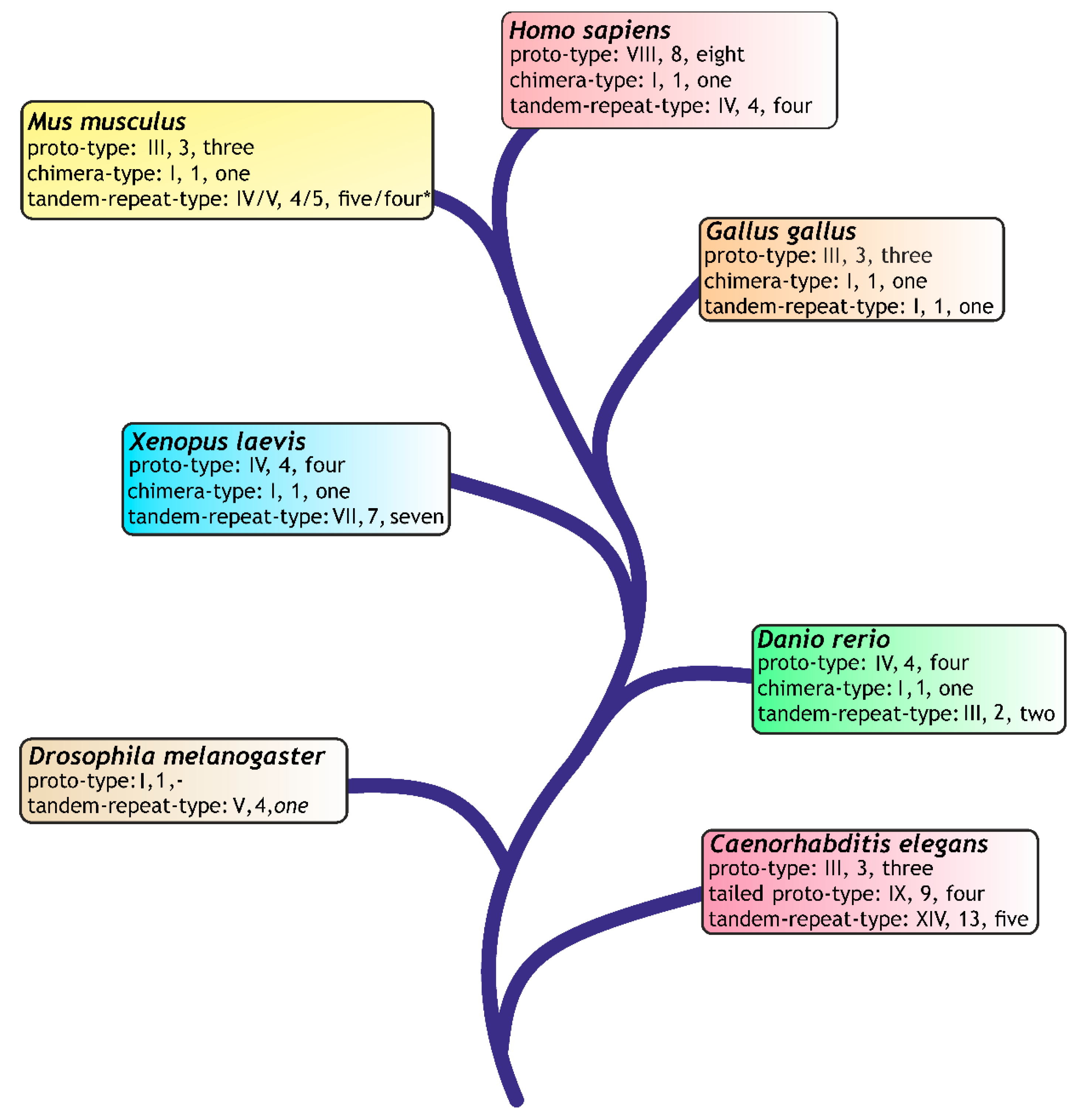
- Kaltner, H.; Gabius, H.-J. A toolbox of lectins for translating the sugar code: The galectin network in phylogenesis and tumors. Histol. Histopathol. 2012, 27, 397–416. [Google Scholar] [PubMed]
- Kaltner, H.; Raschta, A.-S.; Manning, J.C.; Gabius, H.-J. Copy-number variation of functional galectin genes: Studying animal galectin-7 (p53-induced gene 1 in man) and tandem-repeat-type galectins-4 and -9. Glycobiology 2013, 23, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H. Soluble lectins: A new class of extracellular proteins. Science 1984, 223, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Kasai, K.-I.; Hirabayashi, J. Galectins: A family of animal lectins that decipher glycocodes. J. Biochem. 1996, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barondes, S.H. Galectins: A personal review. Trends Glycosci. Glycotechnol. 1997, 9, 1–7. [Google Scholar] [CrossRef]
- Dawson, H.; André, S.; Karamitopoulou, E.; Zlobec, I.; Gabius, H.-J. The growing galectin network in colon cancer and clinical relevance of cytoplasmic galectin-3 reactivity. Anticancer Res. 2013, 33, 3053–3059. [Google Scholar] [PubMed]
- Katzenmaier, E.-M.; André, S.; Kopitz, J.; Gabius, H.-J. Impact of sodium butyrate on the network of adhesion/growth-regulatory galectins in human colon cancer in vitro. Anticancer Res. 2014, 34, 5429–5438. [Google Scholar] [PubMed]
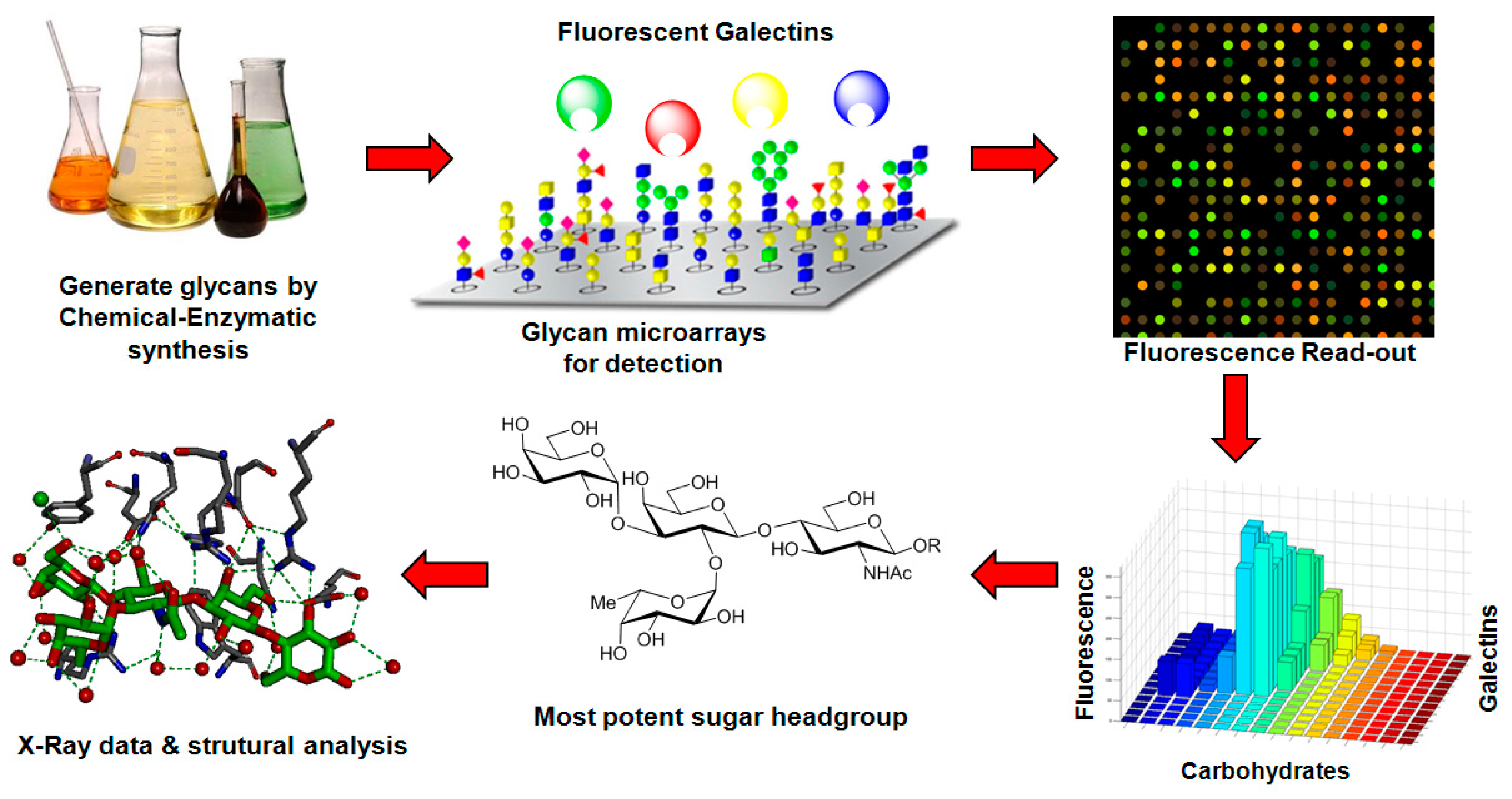
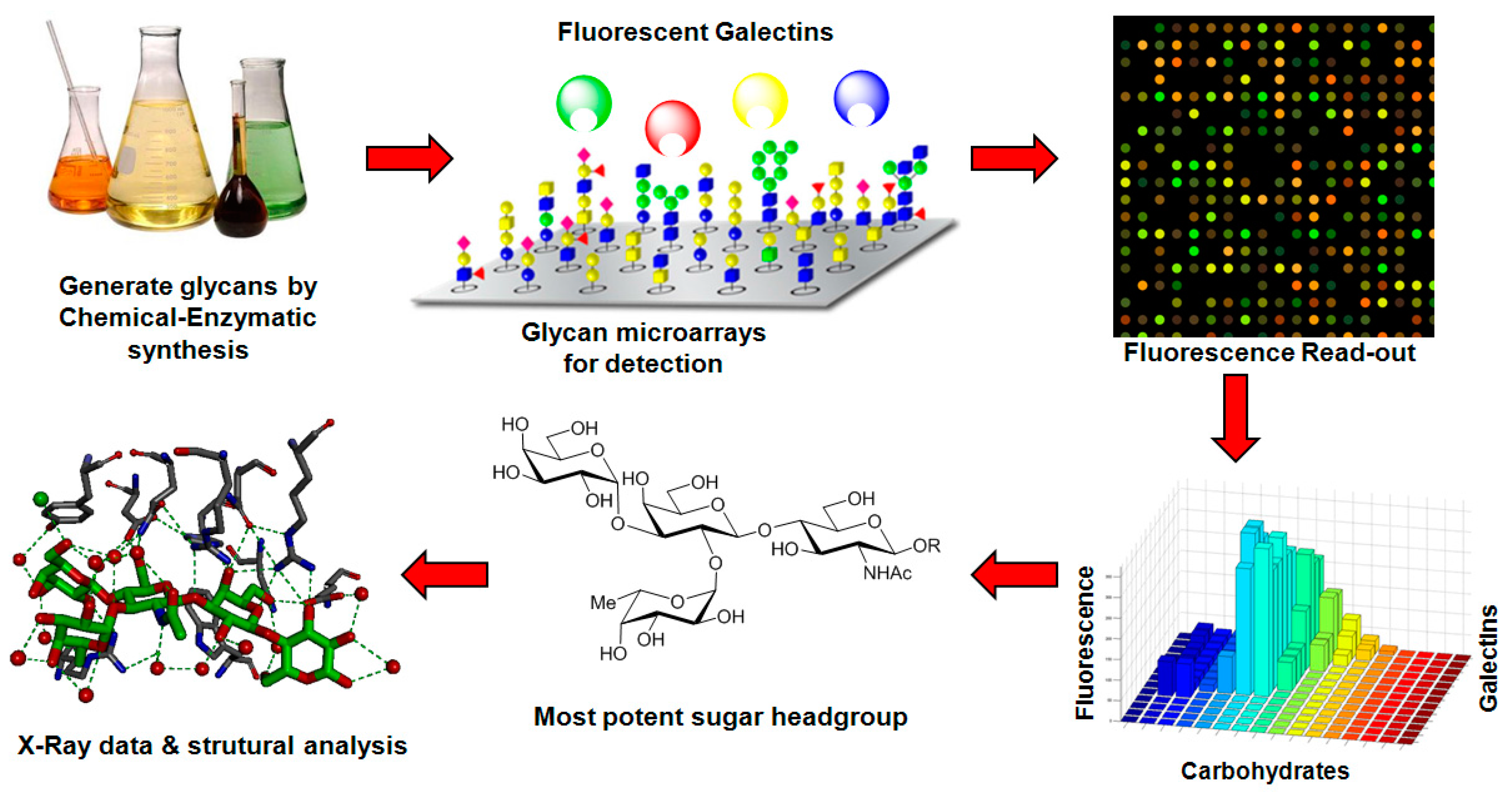
- Gupta, G.; Surolia, A.; Sampathkumar, S.G. Lectin microarrays for glycomic analysis. Omics 2010, 14, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Donczo, B.; Kerekgyarto, J.; Szurmai, Z.; Guttman, A. Glycan microarrays: New angles and new strategies. Analyst 2014, 139, 2650–2657. [Google Scholar] [CrossRef] [PubMed]
- Geissner, A.; Anish, C.; Seeberger, P.H. Glycan arrays as tools for infectious disease research. Curr. Opin. Chem. Biol. 2014, 18, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Heimburg-Molinaro, J.; Cummings, R.D.; Smith, D.F. Chemistry of natural glycan microarrays. Curr. Opin. Chem. Biol. 2014, 18, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Heimburg-Molinaro, J.; Smith, D.F.; Cummings, R.D. Glycan microarrays of fluorescently-tagged natural glycans. Glycoconj. J. 2015, 32, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Teichberg, V.I.; Silman, I.; Beitsch, D.D.; Resheff, G. A β-d-galactoside binding protein from electric organ tissue of Electrophorus electricus. Proc. Natl. Acad. Sci. USA 1975, 72, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, C.; Leffler, H.; Barondes, S.H. Multiple soluble b-galactoside-binding lectins from human lung. J. Biol. Chem. 1987, 262, 7383–7390. [Google Scholar] [PubMed]
- Ahmed, H.; Allen, H.J.; Sharma, A.; Matta, K.L. Human splenic galaptin: Carbohydrate-binding specificity and characterization of the combining site. Biochemistry 1990, 29, 5315–5319. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.T.; Ichikawa, Y.; Allen, H.J.; Lee, Y.C. Binding characteristics of galactoside-binding lectin (galaptin) from human spleen. J. Biol. Chem. 1990, 265, 7864–7871. [Google Scholar] [PubMed]
- Knibbs, R.N.; Agrwal, N.; Wang, J.L.; Goldstein, I.J. Carbohydrate-binding protein 35. II. Analysis of the interaction of the recombinant polypeptide with saccharides. J. Biol. Chem. 1993, 268, 14940–14947. [Google Scholar] [PubMed]
- Ahmad, N.; Gabius, H.-J.; Kaltner, H.; André, S.; Kuwabara, I.; Liu, F.-T.; Oscarson, S.; Norberg, T.; Brewer, C.F. Thermodynamic binding studies of cell surface carbohydrate epitopes to galectins-1, -3 and -7. Evidence for differential binding specificities. Can. J. Chem. 2002, 80, 1096–1104. [Google Scholar] [CrossRef]
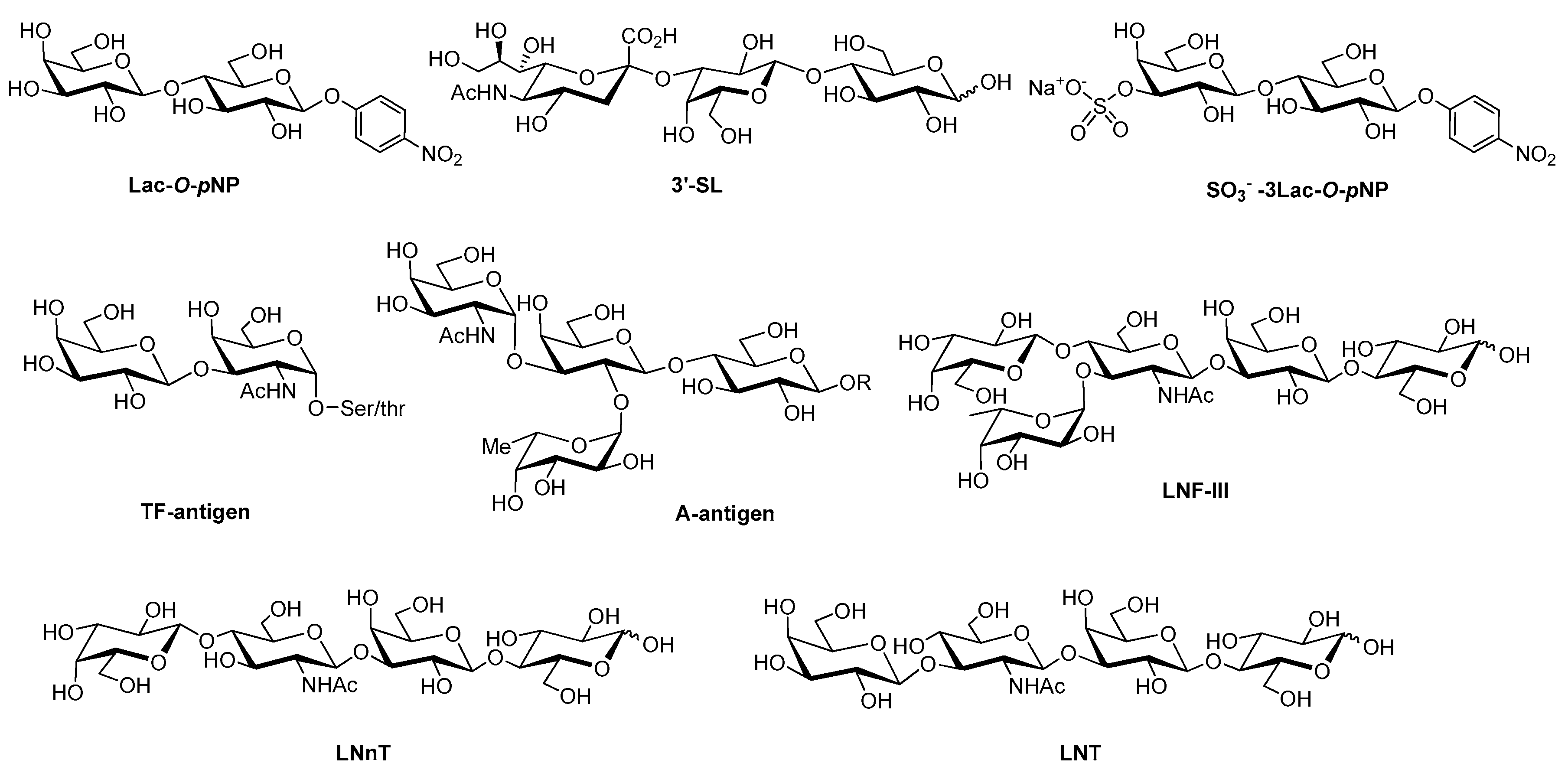
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Müller, W.E.G.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Wu, A.M.; Wu, J.H.; Liu, J.-H.; Singh, T.; André, S.; Kaltner, H.; Gabius, H.-J. Effects of polyvalency of glycotopes and natural modifications of human blood group ABH/Lewis sugars at the Galb1-terminated core saccharides on the binding of domain-I of recombinant tandem-repeat-type galectin-4 from rat gastrointestinal tract (G4-N). Biochimie 2004, 86, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Mehta, P.; Slanina, K.A.; Blixt, O.; Leffler, H.; Smith, D.F.; Cummings, R.D. Galectin-1, -2, and -3 exhibit differential recognition of sialylated glycans and blood group antigens. J. Biol. Chem. 2008, 283, 10109–10123. [Google Scholar] [CrossRef] [PubMed]
- Krzeminski, M.; Singh, T.; André, S.; Lensch, M.; Wu, A.M.; Bonvin, A.M.; Gabius, H.-J. Human galectin-3 (Mac-2 antigen): Defining molecular switches of affinity to natural glycoproteins, structural and dynamic aspects of glycan binding by flexible ligand docking and putative regulatory sequences in the proximal promoter region. Biochim. Biophys. Acta 2011, 1810, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, E.M.; Matveeva, V.K.; Kaltner, H.; André, S.; Vokhmyanina, O.A.; Pazynina, G.V.; Severov, V.V.; Ryzhov, I.M.; Korchagina, E.Y.; Belyanchikov, I.M.; et al. Comparative lectinology: Delineating glycan-specificity profiles of the chicken galectins using neoglycoconjugates in a cell assay. Glycobiology 2015, 25, 726–734. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rauthu, S.R.; Shiao, T.C.; André, S.; Miller, M.C.; Madej, E.; Mayo, K.H.; Gabius, H.-J.; Roy, R. Defining the potential of aglycone modifications for affinity/selectivity enhancement against medically relevant lectins: Synthesis, activity screening, and HSQC-based NMR analysis. ChemBioChem 2015, 16, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Merkle, R.K.; Cummings, R.D. Asparagine-linked oligosaccharides containing poly-N-acetyllactosamine chains are preferentially bound by immobilized calf heart agglutinin. J. Biol. Chem. 1988, 263, 16143–16149. [Google Scholar] [PubMed]
- Ideo, H.; Seko, A.; Ishizuka, I.; Yamashita, K. The N-terminal carbohydrate recognition domain of galectin-8 recognizes specific glycosphingolipids with high affinity. Glycobiology 2003, 13, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, S.; Oberg, C.T.; Carlsson, M.C.; Sundin, A.; Nilsson, U.J.; Smith, D.; Cummings, R.D.; Almkvist, J.; Karlsson, A.; Leffler, H. Affinity of galectin-8 and its carbohydrate recognition domains for ligands in solution and at the cell surface. Glycobiology 2007, 17, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Slanina, K.A.; Horton, J.R.; Smith, D.F.; Cummings, R.D. Dimeric Galectin-8 induces phosphatidylserine exposure in leukocytes through polylactosamine recognition by the C-terminal domain. J. Biol. Chem. 2008, 283, 20547–20559. [Google Scholar] [CrossRef] [PubMed]
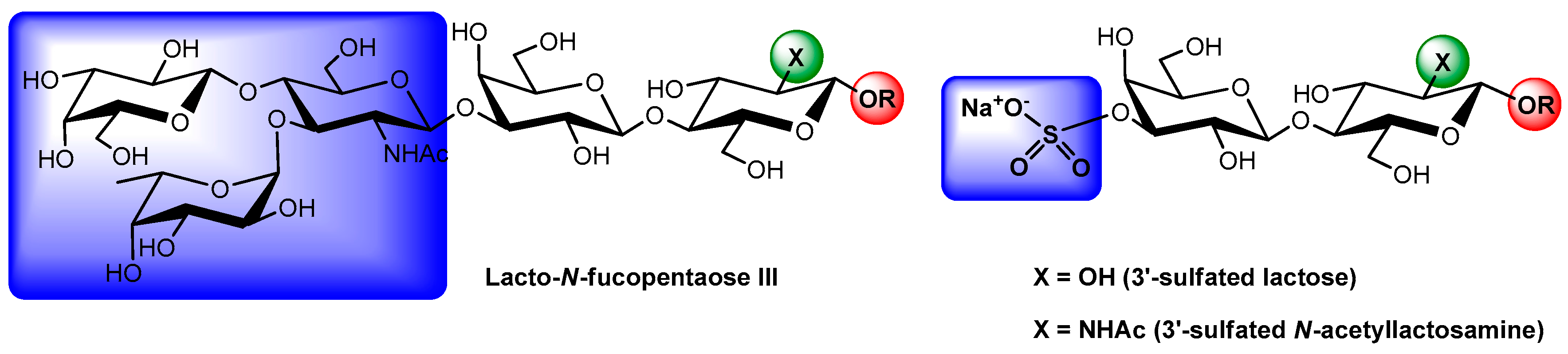
- Ideo, H.; Matsuzaka, T.; Nonaka, T.; Seko, A.; Yamashita, K. Galectin-8-N-domain recognition mechanism for sialylated and sulfated glycans. J. Biol. Chem. 2011, 286, 11346–11355. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.M.; Scholz, B.A.; Buzamet, E.; Kopitz, J.; André, S.; Menendez, M.; Romero, A.; Solís, D.; Gabius, H.-J. Natural single amino acid polymorphism (F19Y) in human galectin-8: Detection of structural alterations and increased growth-regulatory activity on tumor cells. FEBS J. 2014, 281, 1446–1464. [Google Scholar] [CrossRef] [PubMed]
- Vokhmyanina, O.A.; Rapoport, E.M.; André, S.; Severov, V.V.; Ryzhov, I.; Pazynina, G.V.; Korchagina, E.; Gabius, H.-J.; Bovin, N.V. Comparative study of the glycan specificities of cell-bound human tandem-repeat-type galectin-4, -8 and -9. Glycobiology 2012, 22, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.M.; Gilles, U.; Lindner, I.; André, S.; Romero, A.; Reusch, D.; Gabius, H.-J. Combining crystallography and hydrogen-deuterium exchange to study galectin-ligand complexes. Chem. Eur. J. 2015, 21, 13558–13568. [Google Scholar] [CrossRef] [PubMed]
- Arya, P.; Kutterer, K.M.; Qin, H.; Roby, J.; Barnes, M.L.; Kim, J.M.; Roy, R. Diversity of C-linked neoglycopeptides for the exploration of subsite-assisted carbohydrate binding interactions. Bioorg. Med. Chem. Lett. 1998, 8, 1127–1132. [Google Scholar] [CrossRef]
- André, S.; Giguère, D.; Dam, T.K.; Brewer, C.F.; Gabius, H.-J.; Roy, R. Synthesis and screening of a small glycomimetic library for inhibitory activity on medically relevant galactoside-specific lectins in assays of increasing biorelevance. New J. Chem. 2010, 34, 2229–2240. [Google Scholar] [CrossRef]
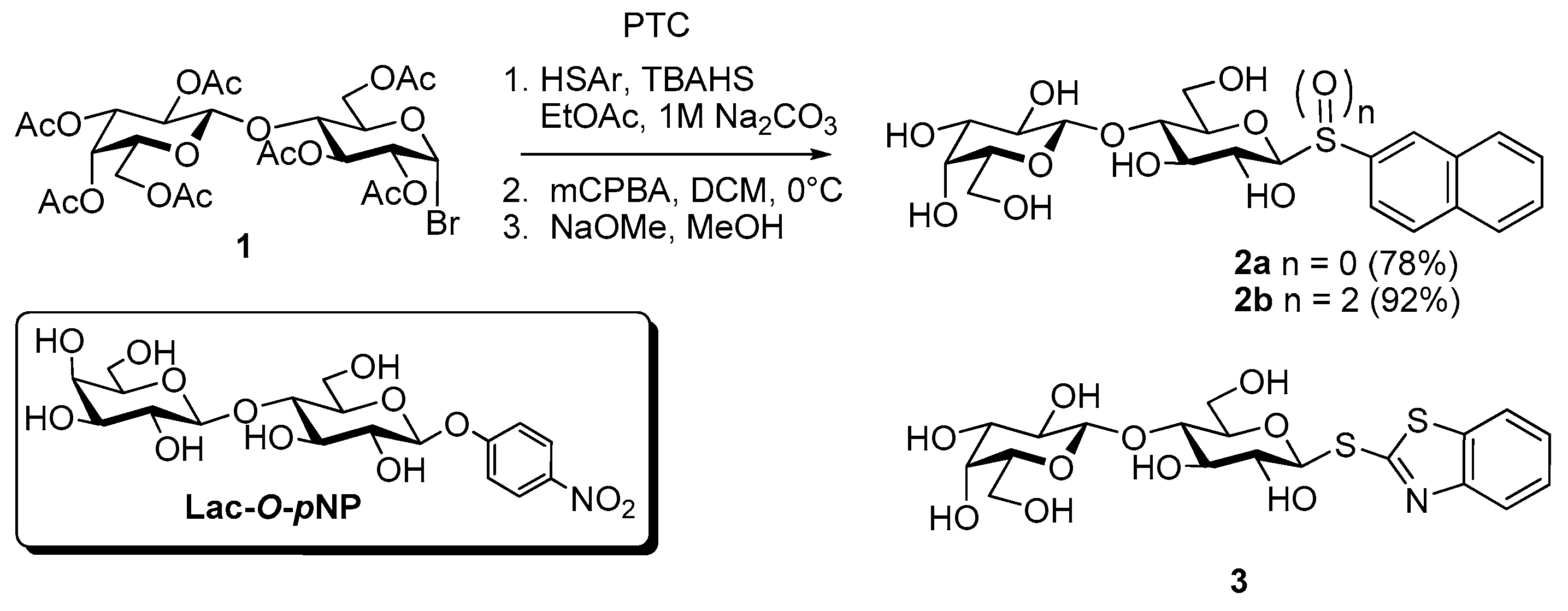
- Giguère, D.; Sato, S.; St-Pierre, C.; Sirois, S.; Roy, R. Aryl O- and S-galactosides and lactosides as specific inhibitors of human galectins-1 and -3: Role of electrostatic potential at O-3. Bioorg. Med. Chem. Lett. 2006, 16, 1668–1672. [Google Scholar] [CrossRef] [PubMed]
- Giguère, D.; Bonin, M.A.; Cloutier, P.; Patnam, R.; St-Pierre, C.; Sato, S.; Roy, R. Synthesis of stable and selective inhibitors of human galectins-1 and -3. Bioorg. Med. Chem. 2008, 16, 7811–7823. [Google Scholar] [CrossRef] [PubMed]
- Solís, D.; Romero, A.; Kaltner, H.; Gabius, H.-J.; Díaz-Mauriño, T. Different architecture of the combining sites of two chicken galectins revealed by chemical-mapping studies with synthetic ligand derivatives. J. Biol. Chem. 1996, 271, 12744–12748. [Google Scholar] [PubMed]
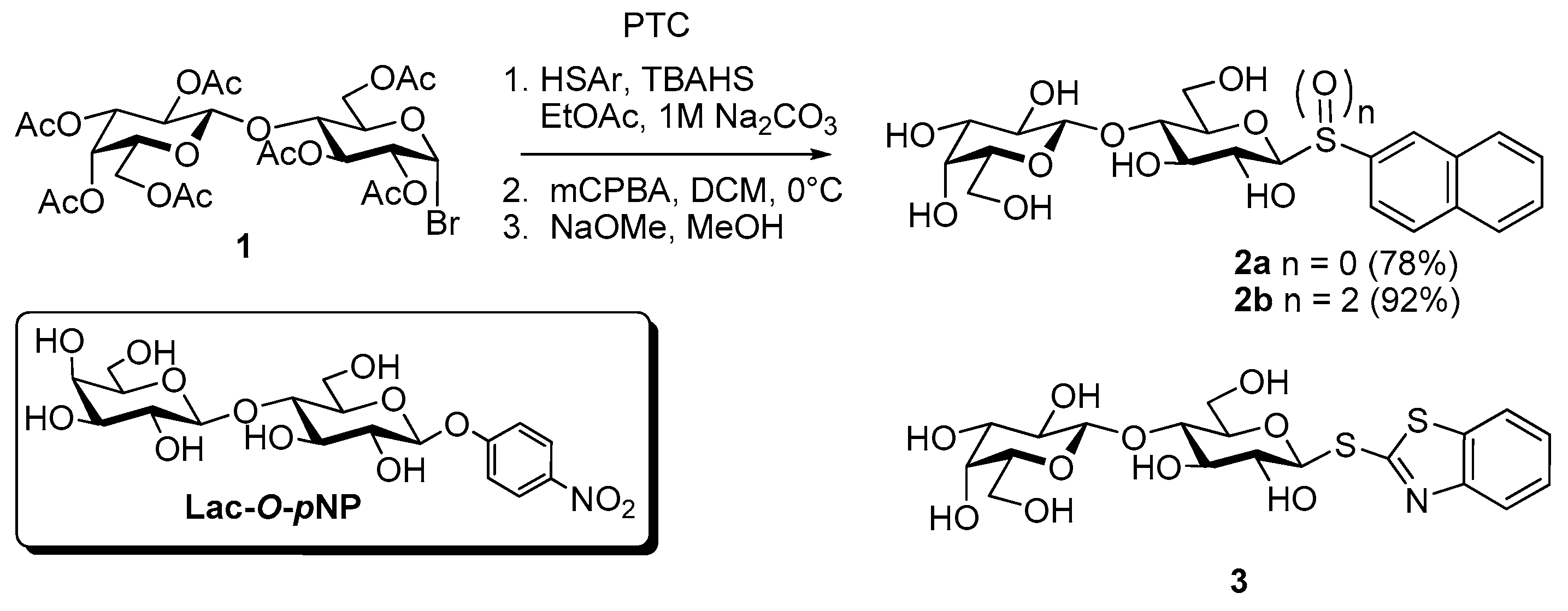
- Roy, R.; Tropper, F.D.; Cao, S.; Kim, J.M. Anomeric group transformations under PTC. ACS Symp. Ser. 1997, 659, 163–180. [Google Scholar]
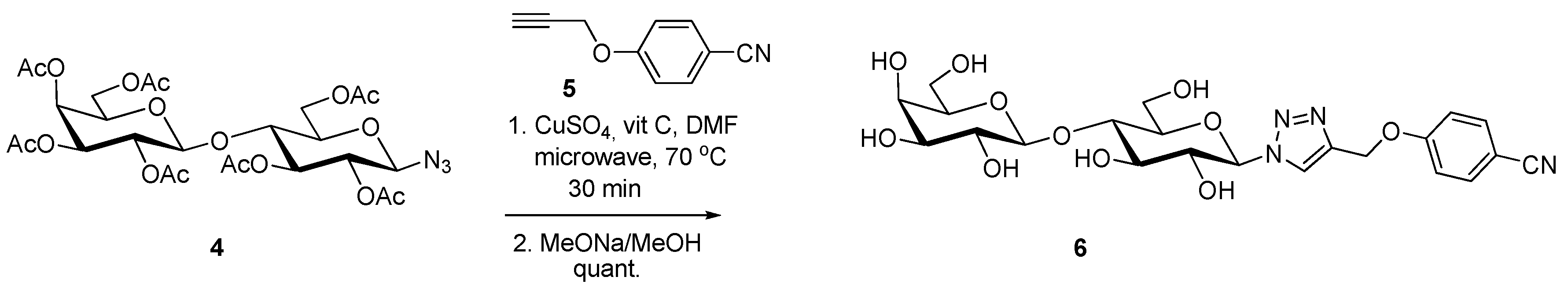
- Giguère, D.; Patnam, R.; Bellefleur, M.A.; St-Pierre, C.; Sato, S.; Roy, R. Carbohydrate triazoles and isoxazoles as inhibitors of galectins-1 and -3. Chem. Commun. 2006. [Google Scholar] [CrossRef]
- Rodrigue, K.; Ganne, G.; Blanchard, B.; Saucier, C.; Giguère, D.; Shiao, T.C.; Varrot, A.; Imberty, A.; Roy, R. Aromatic thioglycoside inhibitors against the virulence factor LecA from Pseudomonas aeruginosa. Org. Biomol. Chem. 2013, 11, 6906–6918. [Google Scholar] [CrossRef] [PubMed]
- López-Lucendo, M.F.; Solís, D.; André, S.; Hirabayashi, J.; Kasai, K.-I.-I.; Kaltner, H.; Gabius, H.-J.; Romero, A. Growth-regulatory human galectin-1: Crystallographic characterisation of the structural changes induced by single-site mutations and their impact on the thermodynamics of ligand binding. J. Mol. Biol. 2004, 343, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Asensio, J.L.; Espinosa, J.F.; Dietrich, H.; Cañada, F.J.; Schmidt, R.R.; Martín-Lomas, M.; André, S.; Gabius, H.-J.; Jiménez-Barbero, J. Bovine heart galectin-1 selects a distinct (syn) conformation of C-lactose, a flexible lactose analogue. J. Am. Chem. Soc. 1999, 121, 8995–9000. [Google Scholar] [CrossRef]
- García-Aparicio, V.; Sollogoub, M.; Blériot, Y.; Colliou, V.; Andrè, S.; Asensio, J.L.; Cañada, F.J.; Gabius, H.-J.; Sinay, P.; Jiménez-Barbero, J. The conformation of the C-glycosyl analogue of N-acetyllactosamine in the free state and bound to a toxic plant agglutinin and human adhesion/growth-regulatory galectin-1. Carbohydr. Res. 2007, 342, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Strino, F.; Lii, J.H.; Koppisetty, C.A.; Nyholm, P.G.; Gabius, H.-J. Selenoglycosides in silico: Ab initio-derived reparameterization of MM4, conformational analysis using histo-blood group ABH antigens and lectin docking as indication for potential of bioactivity. J. Comput. Aided Mol. Des. 2010, 24, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
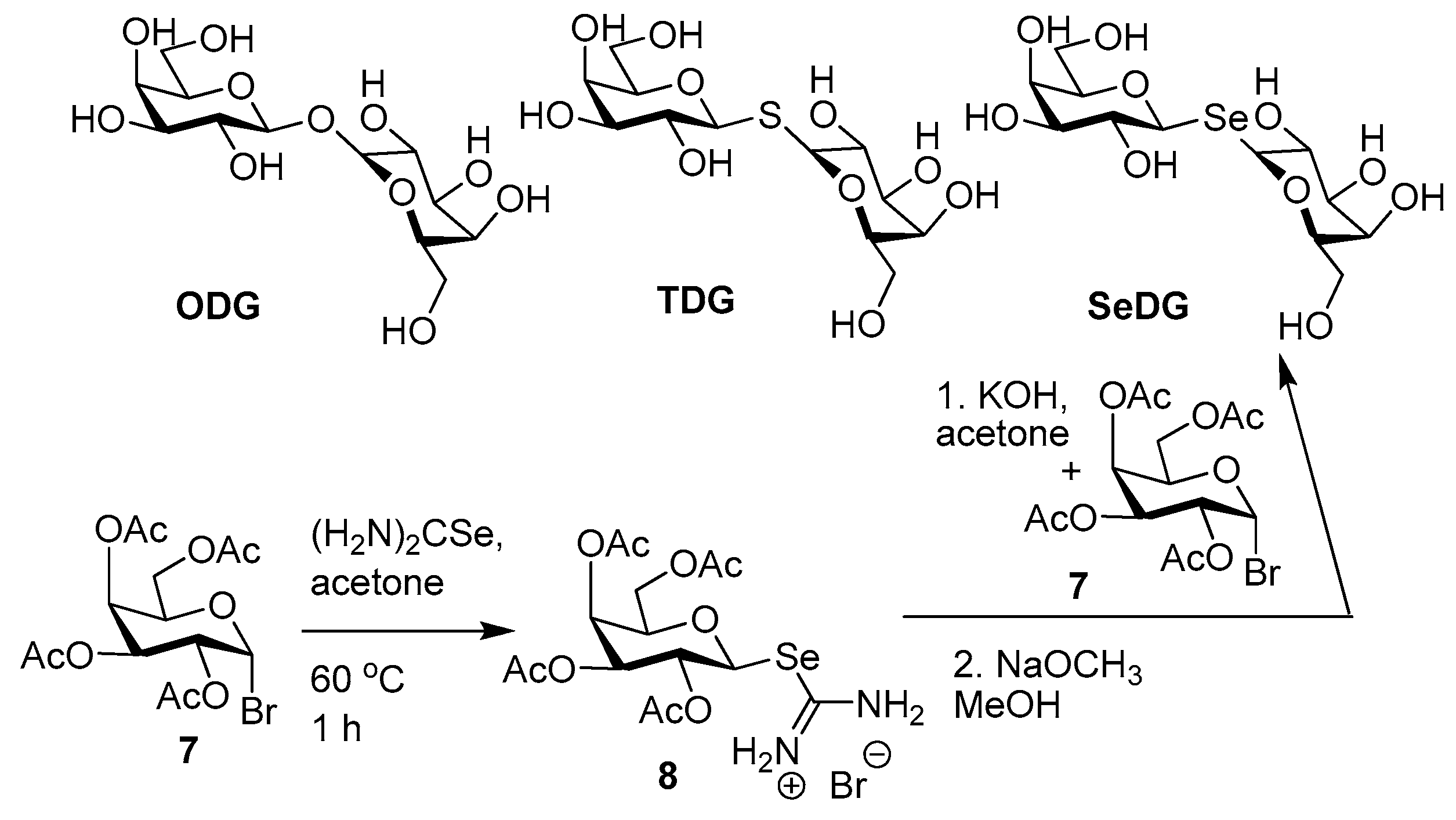
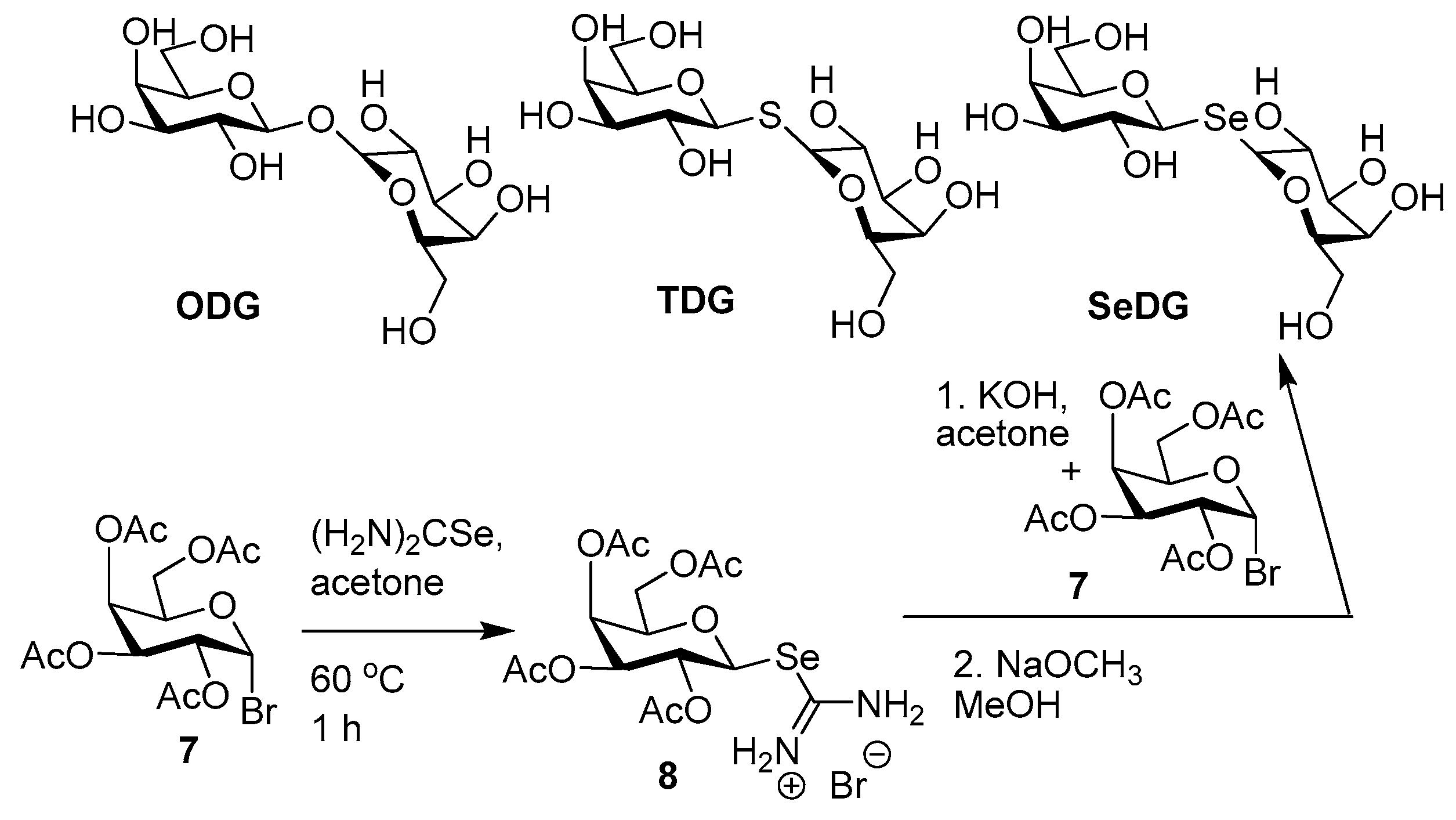
- André, S.; Kover, K.E.; Gabius, H.-J.; Szilagyi, L. Thio- and selenoglycosides as ligands for biomedically relevant lectins: Valency-activity correlations for benzene-based dithiogalactoside clusters and first assessment for (di)selenodigalactosides. Bioorg. Med. Chem. Lett. 2015, 25, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Oscarson, S. The chemist’s way to synthesize glycosides. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 31–51. [Google Scholar]
- Wang, G.-N.; Andre, S.; Gabius, H.-J.; Murphy, P.V. Bi- to tetravalent glycoclusters: Synthesis, structure-activity profiles as lectin inhibitors and impact of combining both valency and headgroup tailoring on selectivity. Org. Biomol. Chem. 2012, 10, 6893–6907. [Google Scholar] [CrossRef] [PubMed]
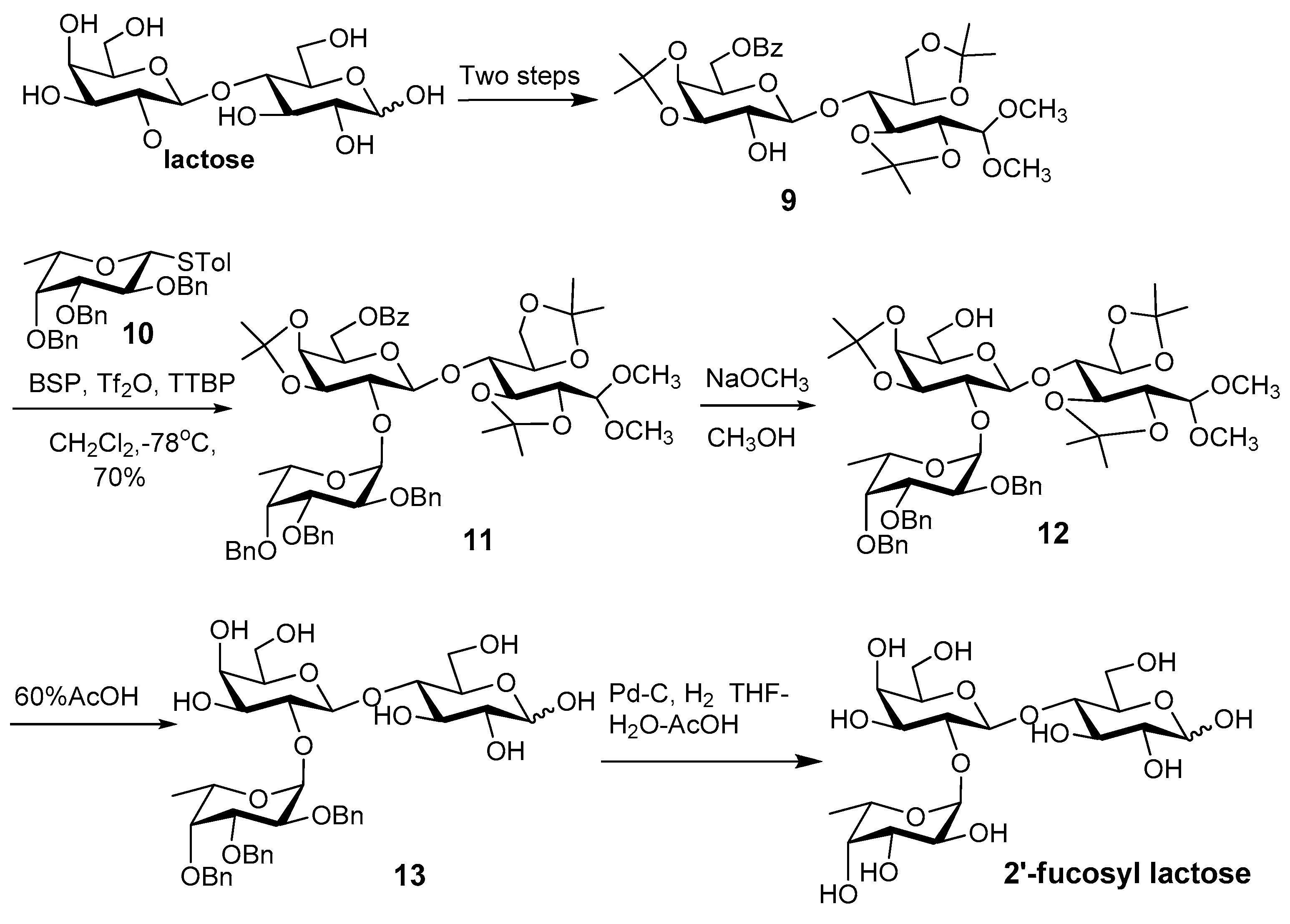
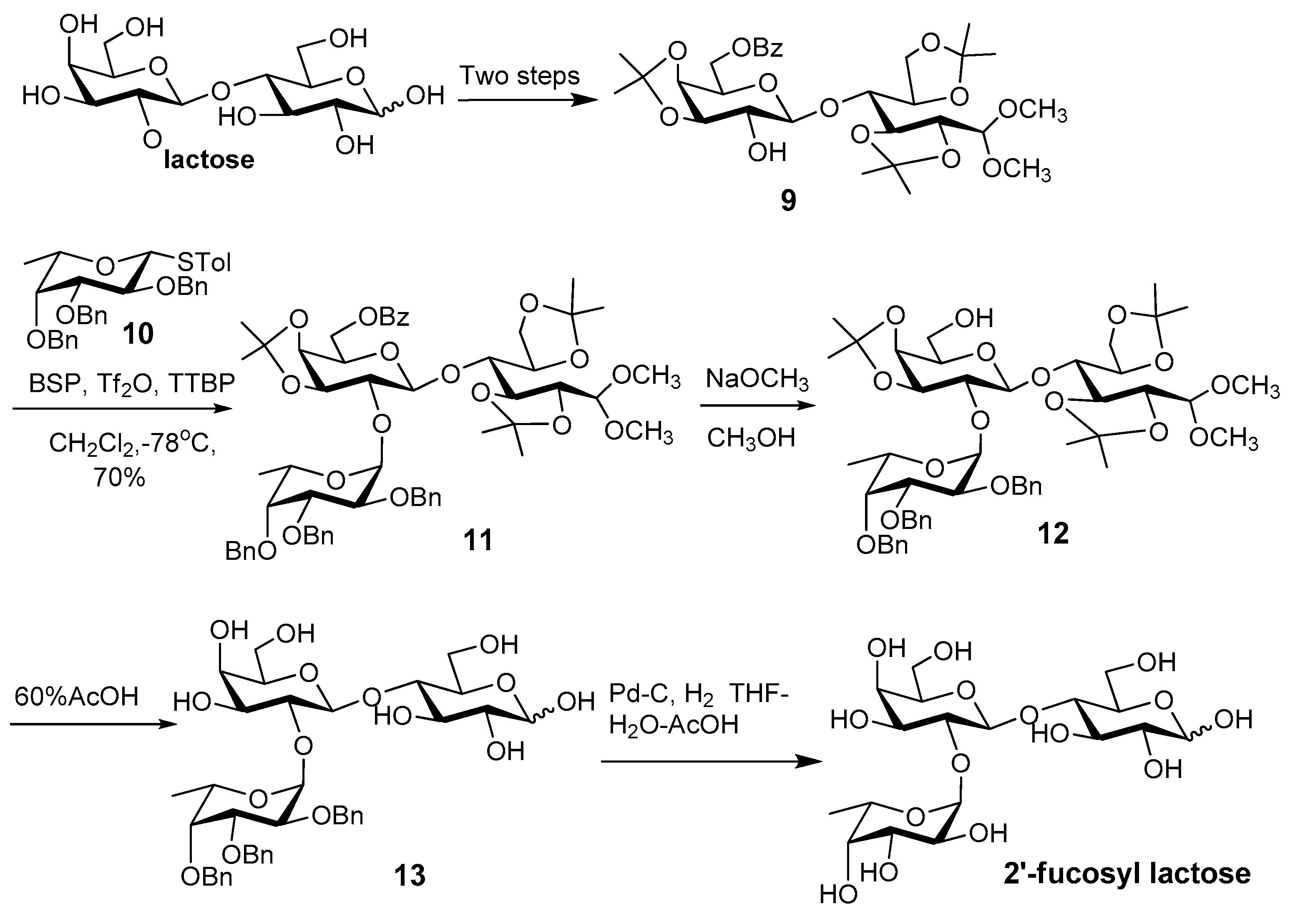
- Abbas, S.A.; Barlow, J.J.; Matta, K.L. Synthetic studies in carbohydrates. 14. Synthesis of O-α-l-fucopyranosyl-(1–2)-O-β-d-galactopyranosyl-(1–4)-d-glucopyranose (2′-O-α-l-fucopyranosyl-lactose). Carbohydr. Res. 1981, 88, 51–60. [Google Scholar] [CrossRef]
- Crich, D.; Smith, M. 1-Benzenesulfinyl piperidine/trifluoromethanesulfonic anhydride: A potent combination of shelf-stable reagents for the low-temperature conversion of thioglycosides to glycosyl triflates and for the formation of diverse glycosidic linkages. J. Am. Chem. Soc. 2001, 123, 9015–9020. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.V. Lewis acid promoted anomerisation: Recent developments and applications. Carbohydr. Chem. 2016, 41, 90–123. [Google Scholar]
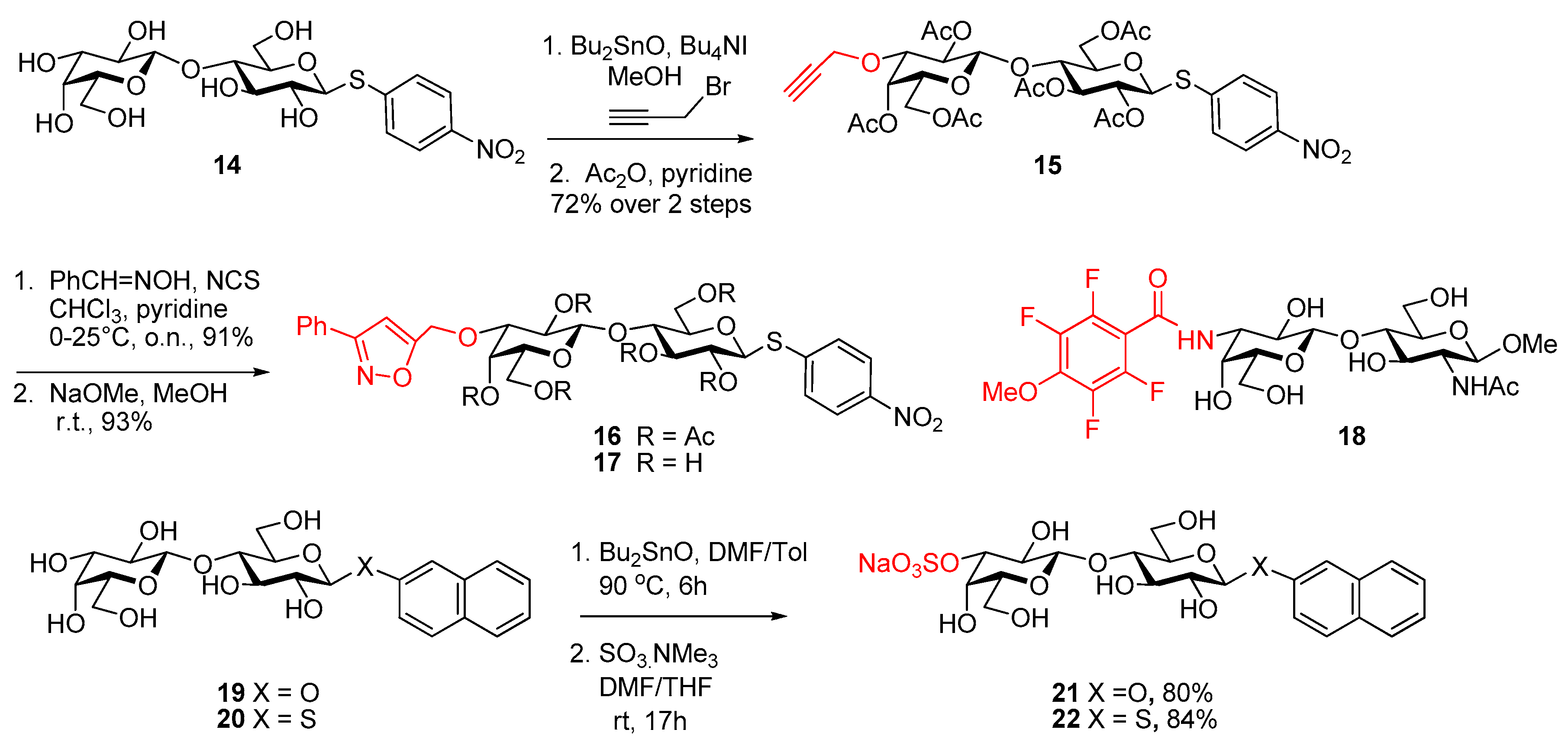
- Giguère, D.; André, S.; Bonin, M.A.; Bellefleur, M.A.; Provencal, A.; Cloutier, P.; Pucci, B.; Roy, R.; Gabius, H.-J. Inhibitory potential of chemical substitutions at bioinspired sites of α-d-galactopyranose on neoglycoprotein/cell surface binding of two classes of medically relevant lectins. Bioorg. Med. Chem. 2011, 19, 3280–3287. [Google Scholar] [CrossRef] [PubMed]
- Sörme, P.; Arnoux, P.; Kahl-Knutsson, B.; Leffler, H.; Rini, J.M.; Nilsson, U.J. Structural and thermodynamic studies on cation-p interactions in lectin-ligand complexes: High-affinity galectin-3 inhibitors through fine-tuning of an arginine-arene interaction. J. Am. Chem. Soc. 2005, 127, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Lee, R.T.; Rice, K.G.; Ichikawa, Y.; Wong, T.C. Topography of binding sites of animal lectins: Ligands’ view. Pure Appl. Chem. 1991, 63, 499–506. [Google Scholar] [CrossRef]
- Lee, R.T.; Lee, Y.C. Enhanced biochemical affinities of multivalent neoglycoconjugates. In Neoglycoconjugates, Preparation and Applications; Lee, Y.C., Lee, R.T., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 23–50. [Google Scholar]
- Roy, R. A decade of glycodendrimer chemistry. Trends Glycosci. Glycotechnol. 2003, 15, 291–310. [Google Scholar] [CrossRef]
- Chabre, Y.M.; Roy, R. The chemist’s way to prepare multivalency. In The Sugar Code: Fundamentals of Glycosciences; Gabius, H.-J., Ed.; Wiley-VCH: Weinheim, Germany, 2009; pp. 53–70. [Google Scholar]
- Chabre, Y.M.; Roy, R. Design and creativity in synthesis of multivalent neoglycoconjugates. Adv. Carbohydr. Chem. Biochem. 2010, 63, 165–393. [Google Scholar] [PubMed]
- Chabre, Y.M.; Roy, R. Multivalent glycoconjugate syntheses and applications using aromatic scaffolds. Chem. Soc. Rev. 2013, 42, 4657–4708. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Shiao, T.C. Glyconanosynthons as powerful scaffolds and building blocks for the rapid construction of multifaceted, dense and chiral dendrimers. Chem. Soc. Rev. 2015, 44, 3924–3941. [Google Scholar] [CrossRef] [PubMed]
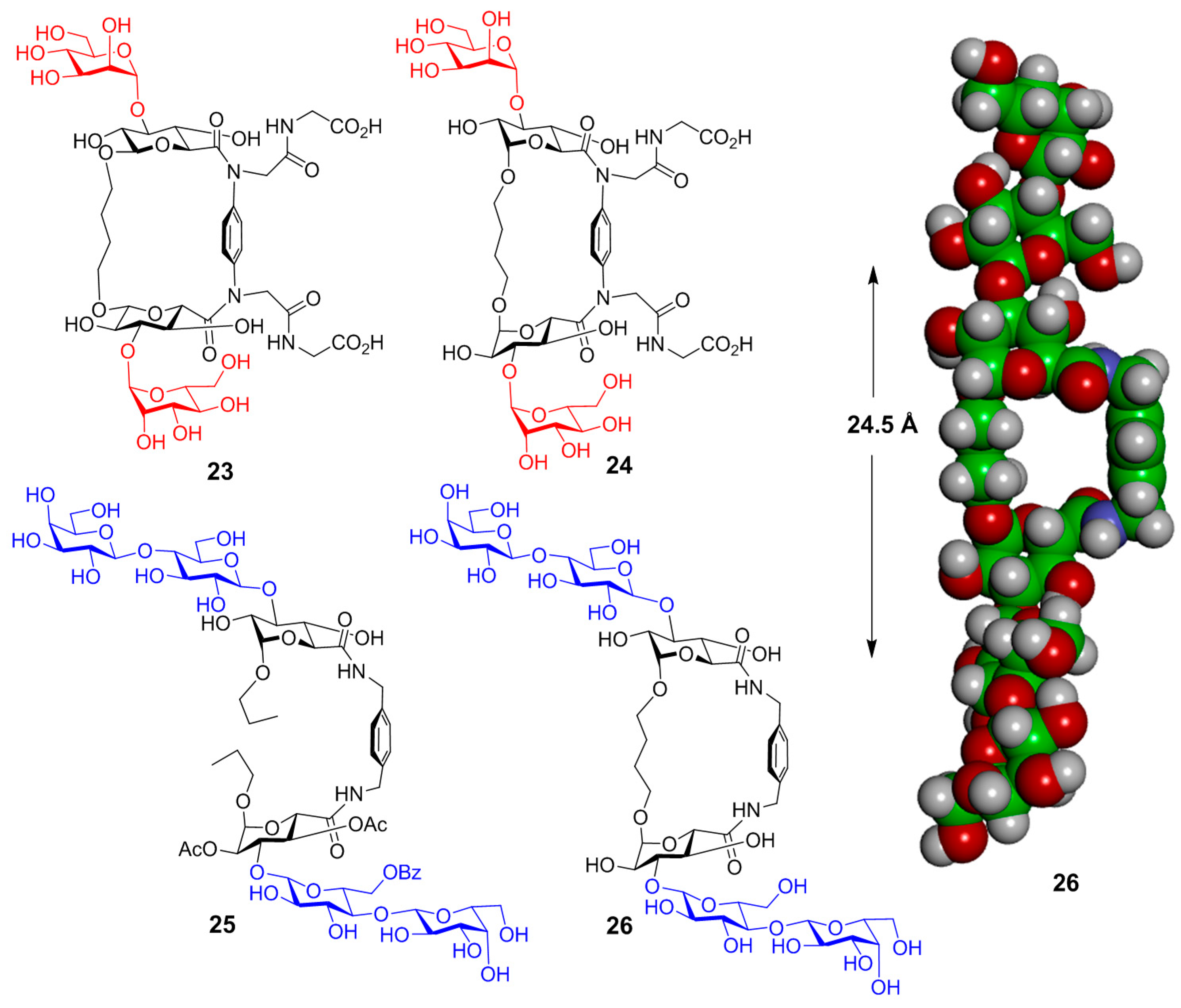
- Velasco-Torrijos, T.; Murphy, P.V. Metathesis of structurally preorganized bivalent carbohydrates. Synthesis of macrocyclic and oligomeric scaffolds. Org. Lett. 2004, 6, 3961–3964. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Torrijos, T.; Murphy, P.V. Synthesis and conformational analysis of novel water soluble macrocycles incorporating carbohydrates, including a beta-cyclodextrin mimic. Tetrahedron Asymmetry 2005, 16, 261–272. [Google Scholar] [CrossRef]
- André, S.; Velasco-Torrijos, T.; Leyden, R.; Gouin, S.; Tosin, M.; Murphy, P.V.; Gabius, H.-J. Phenylenediamine-based bivalent glycocyclophanes: Synthesis and analysis of the influence of scaffold rigidity and ligand spacing on lectin binding in cell systems with different glycomic profiles. Org. Biomol. Chem. 2009, 7, 4715–4725. [Google Scholar] [CrossRef] [PubMed]
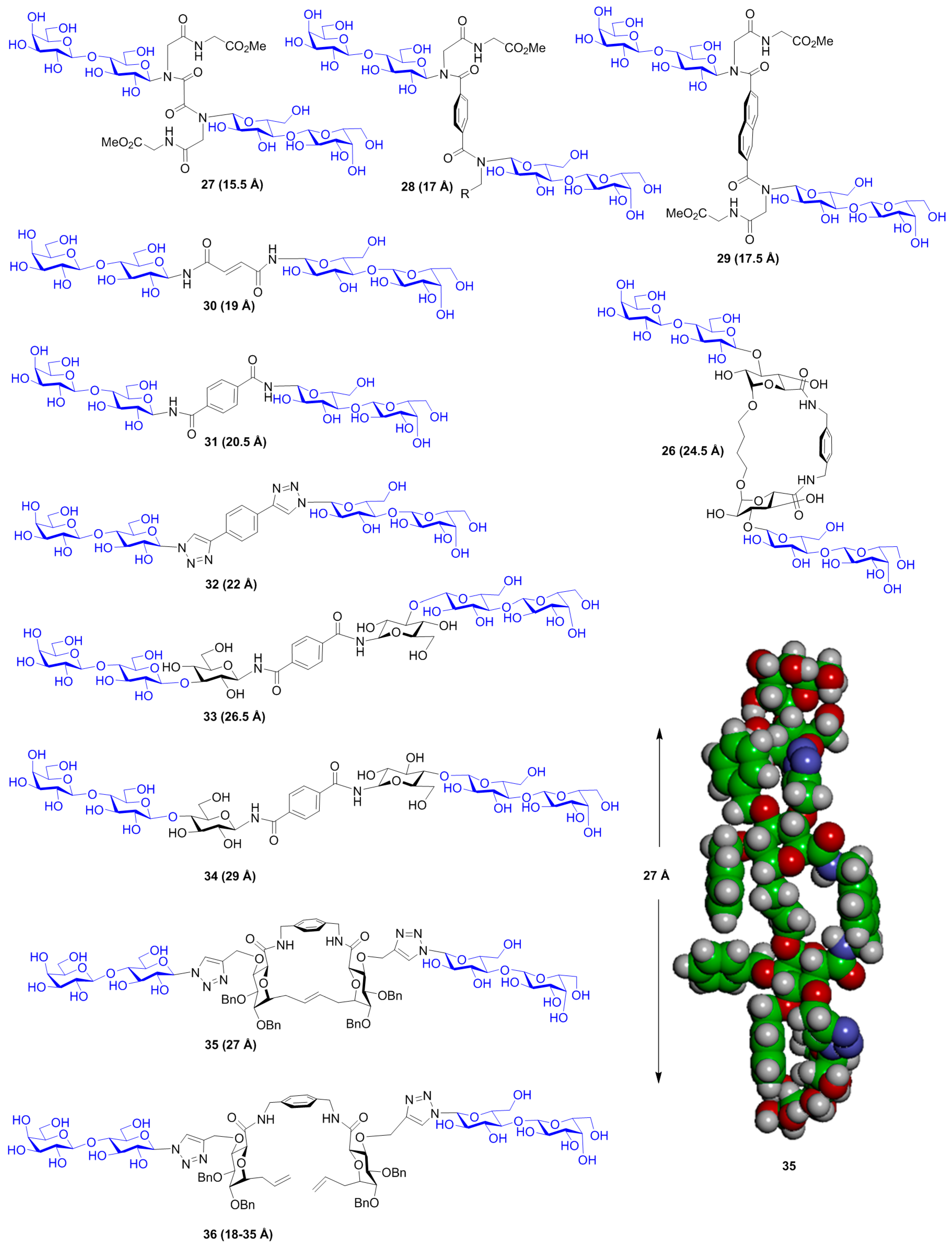
- Leyden, R.; Velasco-Torrijos, T.; André, S.; Gouin, S.; Gabius, H.-J.; Murphy, P.V. Synthesis of bivalent lactosides based on terephthalamide, N,N′-diglucosylterephthalamide, and glycophane scaffolds and assessment of their inhibitory capacity on medically relevant lectins. J. Org. Chem. 2009, 74, 9010–9026. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Jarikote, D.V.; Yan, D.; Vincenz, L.; Wang, G.N.; Kaltner, H.; Murphy, P.V.; Gabius, H.-J. Synthesis of bivalent lactosides and their activity as sensors for differences between lectins in inter- and intrafamily comparisons. Bioorg. Med. Chem. Lett. 2012, 22, 313–318. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Wang, G.N.; Gabius, H.-J.; Murphy, P.V. Combining glycocluster synthesis with protein engineering: An approach to probe into the significance of linker length in a tandem-repeat-type lectin (galectin-4). Carbohydr. Res. 2014, 389, 25–38. [Google Scholar] [CrossRef] [PubMed]
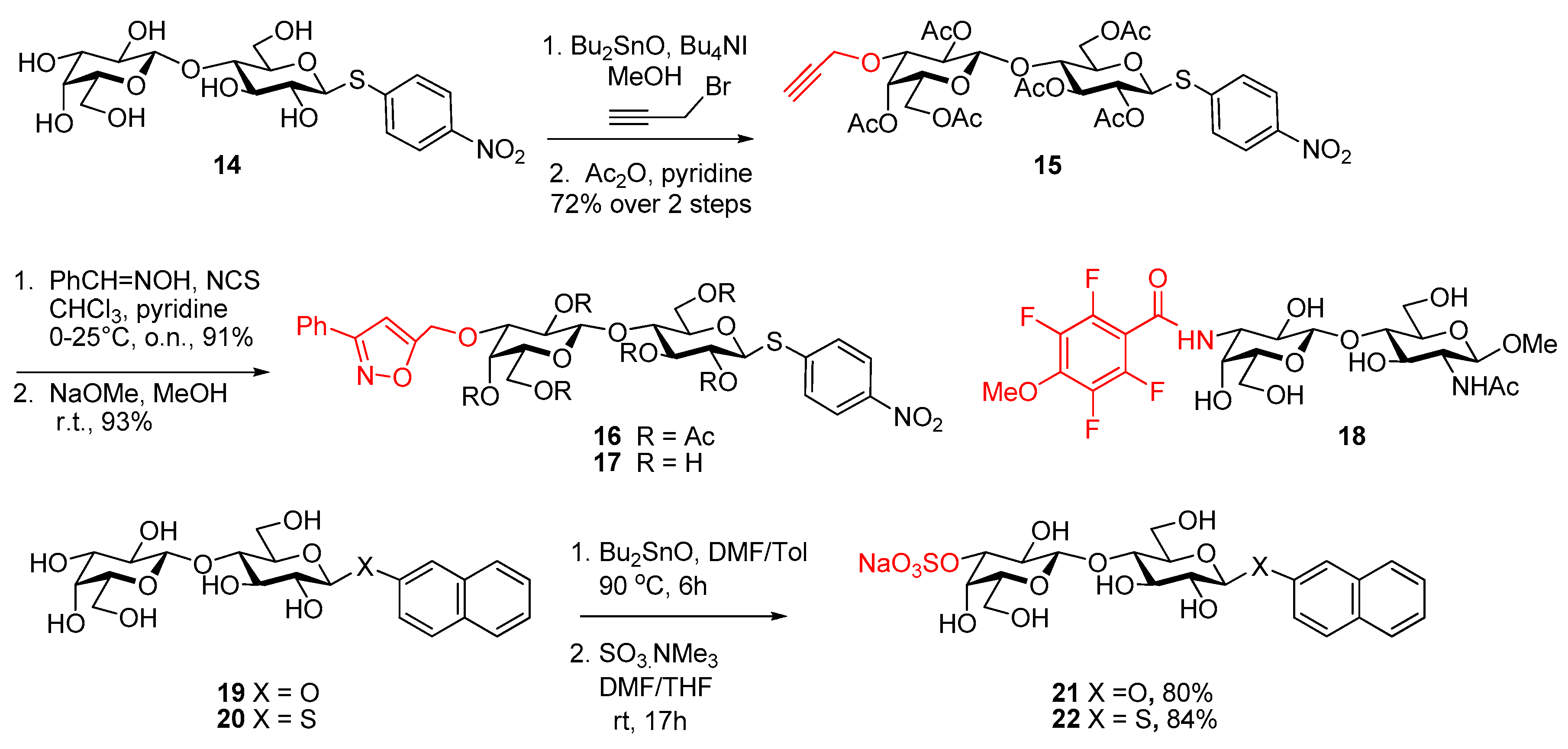
- Poláková, M.; Pitt, N.; Tosin, M.; Murphy, P.V. Glycosidation reactions of silyl ethers with conformationally inverted donors derived from glucuronic acid: Stereoselective synthesis of glycosides and 2-deoxyglycosides. Angew. Chem. Int. Ed. 2004, 43, 2518–2521. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.; Poláková, M.; Pitt, N.; Tosin, M.; Murphy, P.V. Glycosidation-anomerisation reactions of 6,1-anhydroglucopyranuronic acid and anomerisation of β-d-glucopyranosiduronic acids promoted by SnCl4. Chem. Eur. J. 2007, 13, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Jarikote, D.V.; O’Reilly, C.; Murphy, P.V. Ultrasound-assisted synthesis of C-glycosides. Tetrahedron Lett. 2010, 51, 6776–6778. [Google Scholar] [CrossRef]
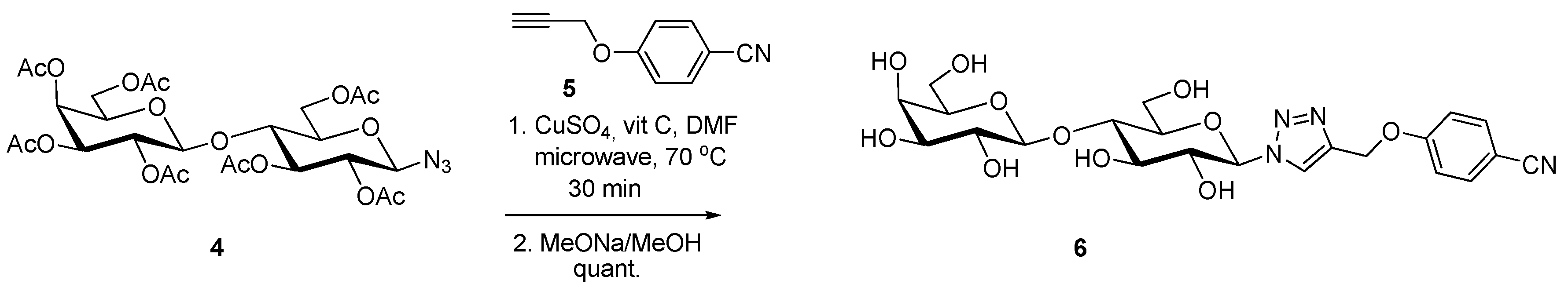
- André, S.; Liu, B.; Gabius, H.-J.; Roy, R. First demonstration of differential inhibition of lectin binding by synthetic tri-and tetravalent glycoclusters from cross-coupling of rigidified 2-propynyl lactoside. Org. Biomol. Chem. 2003, 1, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Yonemura, T.; Matsuura, K.; Kobayashi, K. Tris-bipyridine ruthenium complex-based glycoclusters: Amplified luminescence and enhanced lectin affinities. Tetrahedron Lett. 2001, 42, 3989–3992. [Google Scholar] [CrossRef]
- Roy, R.; Kim, J.M. Cu(II)-self-assembling bipyridyl-glycoclusters and dendrimers bearing the Tn-antigen cancer marker: Syntheses and lectin binding properties. Tetrahedron 2003, 59, 3881–3893. [Google Scholar] [CrossRef]
- Renaudet, O. Recent advances on cyclopeptide-based glycoclusters. Mini Rev. Org. Chem. 2008, 5, 274–286. [Google Scholar] [CrossRef]
- André, S.; Renaudet, O.; Bossu, I.; Dumy, P.; Gabius, H.-J. Cyclic neoglycodecapeptides: How to increase their inhibitory activity and selectivity on lectin/toxin binding to a glycoprotein and cells. J. Pept. Sci. 2011, 17, 427–437. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Sansone, F.; Kaltner, H.; Casnati, A.; Kopitz, J.; Gabius, H.-J.; Ungaro, R. Calix[n]arene-based glycoclusters: Bioactivity of thiourea-linked galactose/lactose moieties as inhibitors of binding of medically relevant lectins to a glycoprotein and cell-surface glycoconjugates and selectivity among human adhesion/growth-regulatory galectins. ChemBioChem 2008, 9, 1649–1661. [Google Scholar] [PubMed]
- André, S.; Grandjean, C.; Gautier, F.M.; Bernardi, S.; Sansone, F.; Gabius, H.-J.; Ungaro, R. Combining carbohydrate substitutions at bioinspired positions with multivalent presentation towards optimising lectin inhibitors: Case study with calixarenes. Chem. Commun. 2011, 47, 6126–6128. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Gabius, H.-J.; André, S.; Kaltner, H.; Sabesan, S.; Roy, R.; Liu, B.; Macaluso, F.; Brewer, C.F. Galectin-3 precipitates as a pentamer with synthetic multivalent carbohydrates and forms heterogeneous cross-linked complexes. J. Biol. Chem. 2004, 279, 10841–10847. [Google Scholar] [CrossRef] [PubMed]
- Kopitz, J.; Vértesy, S.; André, S.; Fiedler, S.; Schnölzer, M.; Gabius, H.-J. Human chimera-type galectin-3: Defining the critical tail length for high-affinity glycoprotein/cell surface binding and functional competition with galectin-1 in neuroblastoma cell growth regulation. Biochimie 2014, 104, 90–99. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Cejas Ortega, P.J.; Perez, M.A.; Roy, R.; Gabius, H.-J. Lactose-containing starburst dendrimers: Influence of dendrimer generation and binding-site orientation of receptors (plant/animal lectins and immunoglobulins) on binding properties. Glycobiology 1999, 9, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Chabre, Y.M.; Contino-Pepin, C.; Placide, V.; Shiao, T.C.; Roy, R. Expeditive synthesis of glycodendrimer scaffolds based on versatile TRIS and mannoside derivatives. J. Org. Chem. 2008, 73, 5602–5605. [Google Scholar] [CrossRef] [PubMed]
- Chabre, Y.M.; Brisebois, P.P.; Abbassi, L.; Kerr, S.C.; Fahy, J.V.; Marcotte, I.; Roy, R. Hexaphenylbenzene as a rigid template for the straightforward syntheses of “star-shaped” glycodendrimers. J. Org. Chem. 2011, 76, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Zhang, I.; Abbassi, L.; Rej, R.; Maysinger, D.; Roy, R. A fast track strategy toward highly functionalized dendrimers with different structural layers: An “onion peel approach”. Polym. Chem. 2015, 6, 1436–1444. [Google Scholar] [CrossRef]
- Sharma, R.; Naresh, K.; Chabre, Y.M.; Rej, R.; Saadeh, N.K.; Roy, R. “Onion peel” dendrimers: A straightforward synthetic approach towards highly diversified architectures. Polym. Chem. 2014, 5, 4321–4331. [Google Scholar] [CrossRef]
- Sharma, R.; Kottari, N.; Chabre, Y.M.; Abbassi, L.; Shiao, T.C.; Roy, R. A highly versatile convergent/divergent “onion peel” synthetic strategy toward potent multivalent glycodendrimers. Chem. Commun. 2014, 50, 13300–13303. [Google Scholar] [CrossRef] [PubMed]
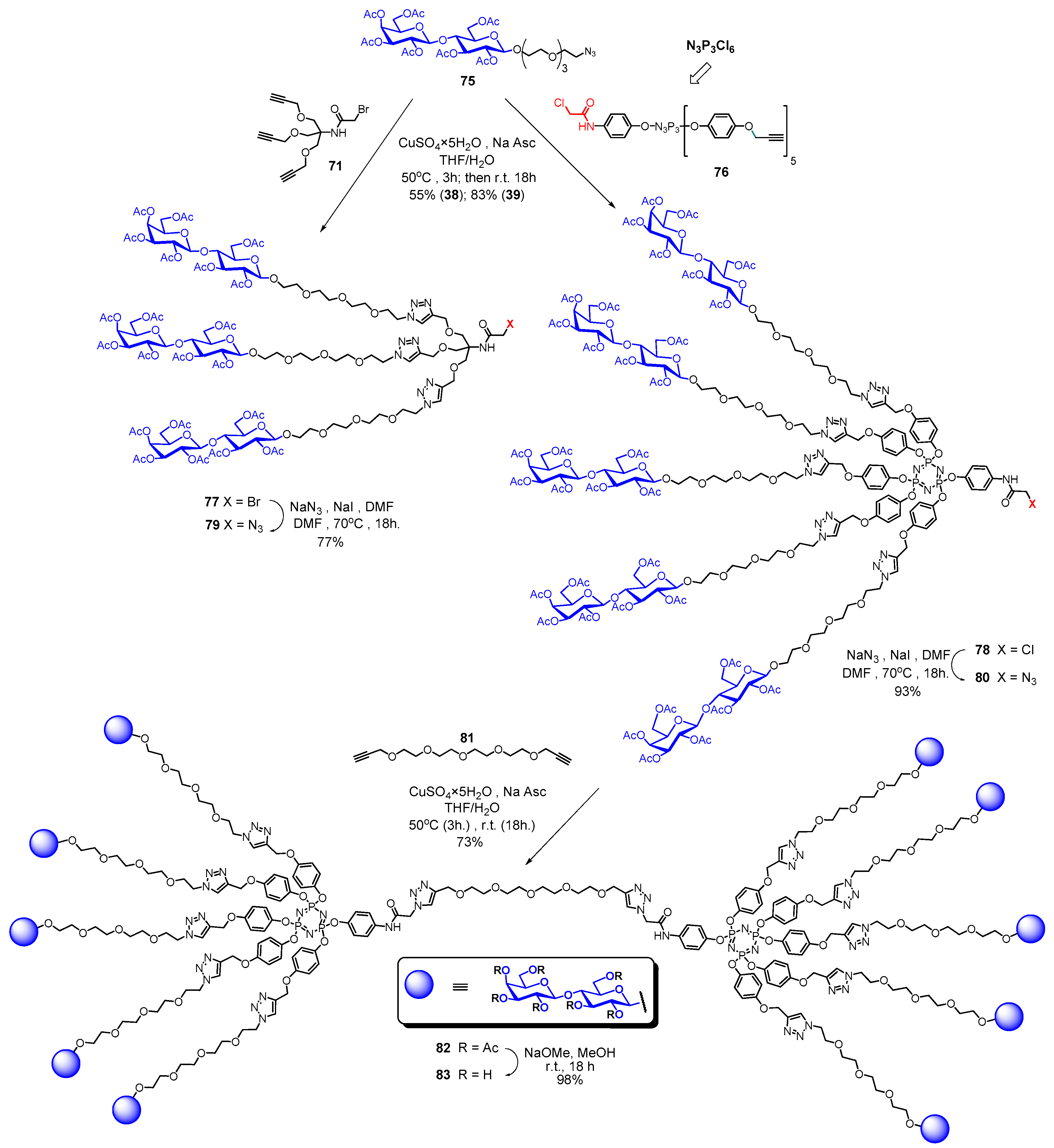
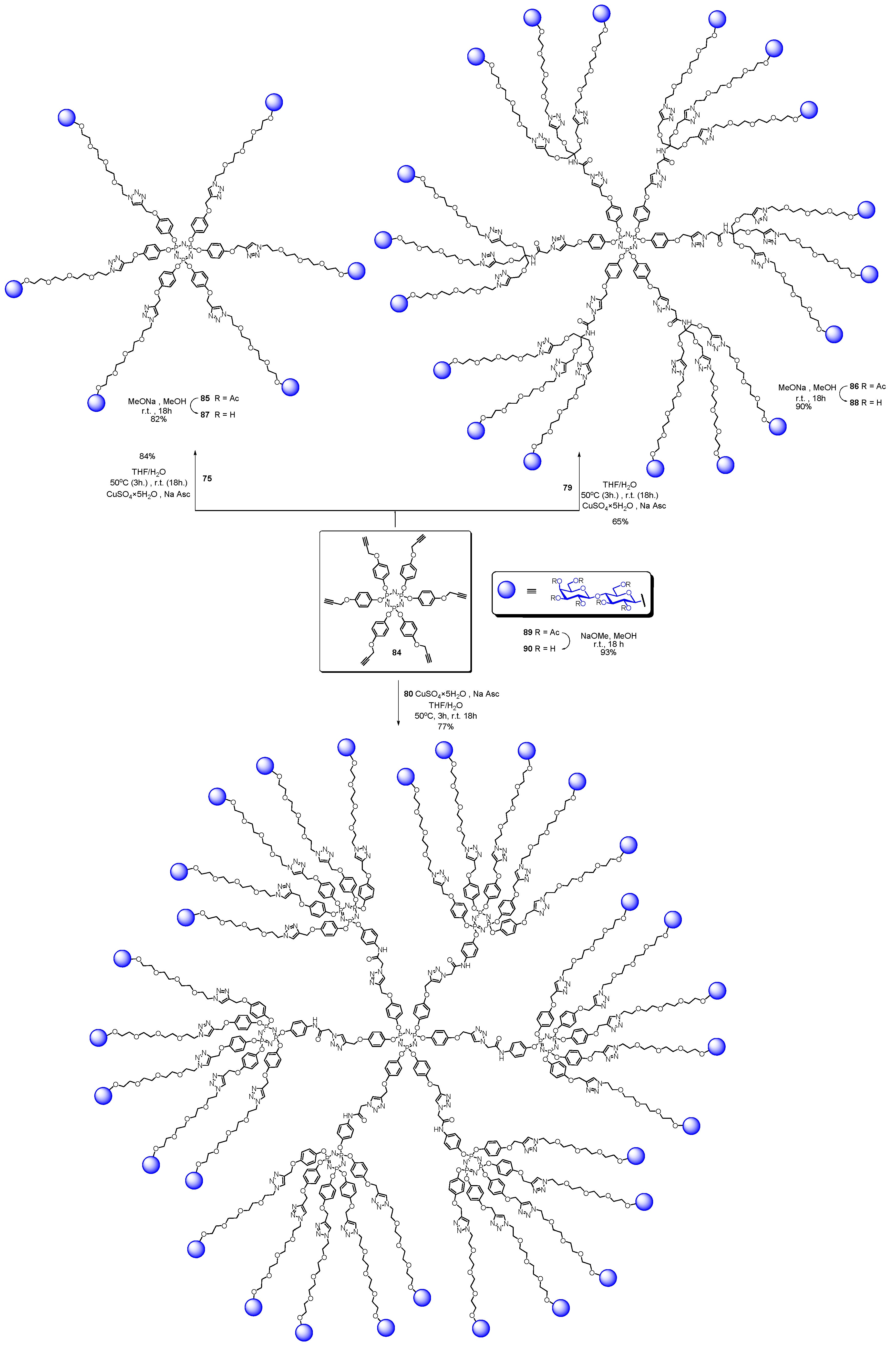
- Abbassi, L.; Chabre, Y.M.; Kottari, N.; Arnold, A.A.; André, S.; Josserand, J.; Gabius, H.-J.; Roy, R. Multifaceted glycodendrimers with programmable bioactivity through convergent, divergent, and accelerated approaches using polyfunctional cyclotriphosphazenes. Polym. Chem. 2015, 6, 7666–7683. [Google Scholar] [CrossRef]
- Percec, V.; Leowanawat, P.; Sun, H.J.; Kulikov, O.; Nusbaum, C.D.; Tran, T.M.; Bertin, A.; Wilson, D.A.; Peterca, M.; Zhang, S.; et al. Modular synthesis of amphiphilic Janus glycodendrimers and their self-assembly into glycodendrimersomes and other complex architectures with bioactivity to biomedically relevant lectins. J. Am. Chem. Soc. 2013, 135, 9055–9077. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Zhang, S.; Wang, Z.; Sherman, S.E.; Moussodia, R.O.; Peterca, M.; Muncan, A.; Williams, D.R.; Hammer, D.A.; Vértesy, S.; et al. Onion-like glycodendrimersomes from sequence-defined Janus glycodendrimers and influence of architecture on reactivity to a lectin. Proc. Natl. Acad. Sci. USA 2016, 113, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Moussodia, R.-O.; Murzeau, C.; Sun, H.J.; Klein, M.L.; Vértesy, S.; André, S.; Roy, R.; Gabius, H.-J.; Percec, V. Dissecting molecular aspects of cell interactions using glycodendrimersomes with programmable glycan presentation and engineered human lectins. Angew. Chem. Int. Ed. 2015, 54, 4036–4040. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xiao, Q.; Sherman, S.E.; Muncan, A.; Vicente, A.D.R.; Wang, Z.; Hammer, D.A.; Williams, D.; Chen, Y.; Pochan, D.J.; et al. Glycodendrimersomes from sequence-defined Janus glycodendrimers reveal high activity and sensor capacity for the agglutination by natural variants of human lectins. J. Am. Chem. Soc. 2015, 137, 13334–13344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Moussodia, R.-O.; Vértesy, S.; André, S.; Klein, M.L.; Gabius, H.-J.; Percec, V. Unraveling functional significance of natural variations of a human galectin by glycodendrimersomes with programmable glycan surface. Proc. Natl. Acad. Sci. USA 2015, 112, 5585–5590. [Google Scholar] [CrossRef] [PubMed]
- Dettmann, W.; Grandbois, M.; André, S.; Benoit, M.; Wehle, A.K.; Kaltner, H.; Gabius, H.-J.; Gaub, H.E. Differences in zero-force and force-driven kinetics of ligand dissociation from β-galactoside-specific proteins (plant and animal lectins, immunoglobulin G) monitored by plasmon resonance and dynamic single molecule force microscopy. Arch. Biochem. Biophys. 2000, 383, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Pace, K.E.; Baum, L.G. Induction of T lymphocyte apoptosis: A novel function for galectin-1. Trends Glycosci. Glycotechnol. 1997, 9, 21–29. [Google Scholar] [CrossRef]
- Rorive, S.; Belot, N.; Decaestecker, C.; Lefranc, F.; Gordower, L.; Micik, S.; Maurage, C.-A.; Kaltner, H.; Ruchoux, M.-M.; Danguy, A.; et al. Galectin-1 is highly expressed in human gliomas with relevance for modulation of invasion of tumor astrocytes into the brain parenchyma. Glia 2001, 33, 241–255. [Google Scholar] [CrossRef]
- Sanchez-Ruderisch, H.; Detjen, K.M.; Welzel, M.; André, S.; Fischer, C.; Gabius, H.-J.; Rosewicz, S. Galectin-1 sensitizes carcinoma cells to anoikis via the fibronectin receptor α5β1-integrin. Cell Death Differ. 2011, 18, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-T.; Yang, R.Y.; Hsu, D.K. Galectins in acute and chronic inflammation. Ann. N. Y. Acad. Sci. 2012, 1253, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Toegel, S.; Bieder, D.; André, S.; Kayser, K.; Walzer, S.M.; Hobusch, G.; Windhager, R.; Gabius, H.-J. Human osteoarthritic knee cartilage: Fingerprinting of adhesion/growth-regulatory galectins in vitro and in situ indicates differential upregulation in severe degeneration. Histochem. Cell Biol. 2014, 142, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Thiemann, S.; Baum, L.G. Galectins and immune responses—Just how do they do those things they do? Annu. Rev. Immunol. 2016, 34, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Toegel, S.; Weinmann, D.; André, S.; Walzer, S.M.; Bilban, M.; Schmidt, S.; Chiari, C.; Windhager, R.; Krall, C.; Bennani-Baiti, I.M.; et al. Galectin-1 couples glycobiology to inflammation in osteoarthritis through the activation of an NF-κB-regulated gene network. J. Immunol. 2016, 196, 1910–1921. [Google Scholar] [CrossRef] [PubMed]
- Pál, Z.; Antal, P.; Srivastava, S.K.; Hullám, G.; Semsei, A.F.; Gál, J.; Svébis, M.; Soós, G.; Szalai, C.; André, S.; et al. Non-synonymous single nucleotide polymorphisms in genes for immunoregulatory galectins: Association of galectin-8 (F19Y) occurrence with autoimmune diseases in a Caucasian population. Biochim. Biophys. Acta 2012, 1820, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Yadavalli, S.S.; Zhang, S.; Sherman, S.E.; Fiorin, E.; da Silva, L.; Wilson, D.A.; Hammer, D.A.; André, S.; Gabius, H.-J.; et al. Bioactive cell-like hybrids co-assembled from (glyco)dendrimersomes with bacterial membranes. Proc. Natl. Acad. Sci. USA 2016, 113, E1134–E1141. [Google Scholar] [CrossRef] [PubMed]
- Majewski, J.; André, S.; Jones, E.; Chi, E.; Gabius, H.-J. X-ray reflectivity and grazing incidence diffraction studies of interaction between huma adhesion/growth-regulatory galectin-1 and DPPE:GM1 lipid monolayer at the air/water interface. Biochemistry 2015, 80, 943–956. [Google Scholar] [PubMed]
- Swanson, M.D.; Boudreaux, D.M.; Salmon, L.; Chugh, J.; Winter, H.C.; Meagher, J.L.; André, S.; Murphy, P.V.; Oscarson, S.; Roy, R.; et al. Engineering a therapeutic lectin by uncoupling mitogenicity from antiviral activity. Cell 2015, 163, 746–758. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Kaltner, H.; Kayser, K.; Murphy, P.V.; Gabius, H.-J. Merging carbohydrate chemistry with lectin histochemistry to study inhibition of lectin binding by glycoclusters in the natural tissue context. Histochem. Cell Biol. 2016, 145, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Leffler, H.; Nilsson, U. Low-molecular weight inhibitors of galectins. ACS Symp. Ser. 2012, 1115, 47–59. [Google Scholar]
- Oeberg, C.T.; Leffler, H.; Nilsson, U.J. Inhibition of galectins with small molecules. Chimia 2011, 65, 18–23. [Google Scholar] [CrossRef]
- Hockl, P.F.; Wolosiuk, A.; Pérez-Sáez, J.M.; Bordoni, A.V.; Croci, D.O.; Toum-Terrones, Y.; Soler-Illia, G.J.; Rabinovich, G.A. Glyco-nano-oncology: Novel therapeutic opportunities by combining small and sweet. Pharm. Res. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
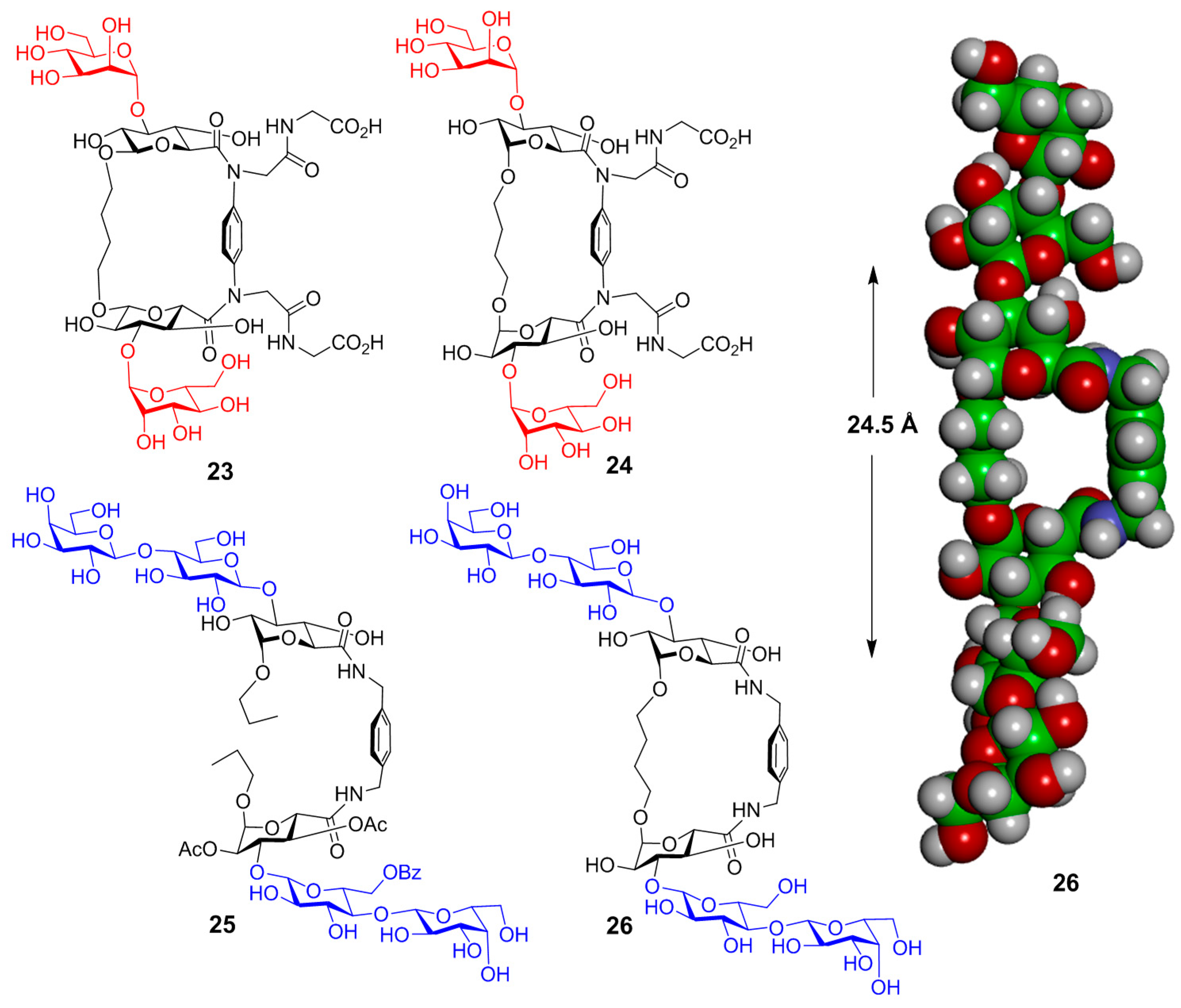
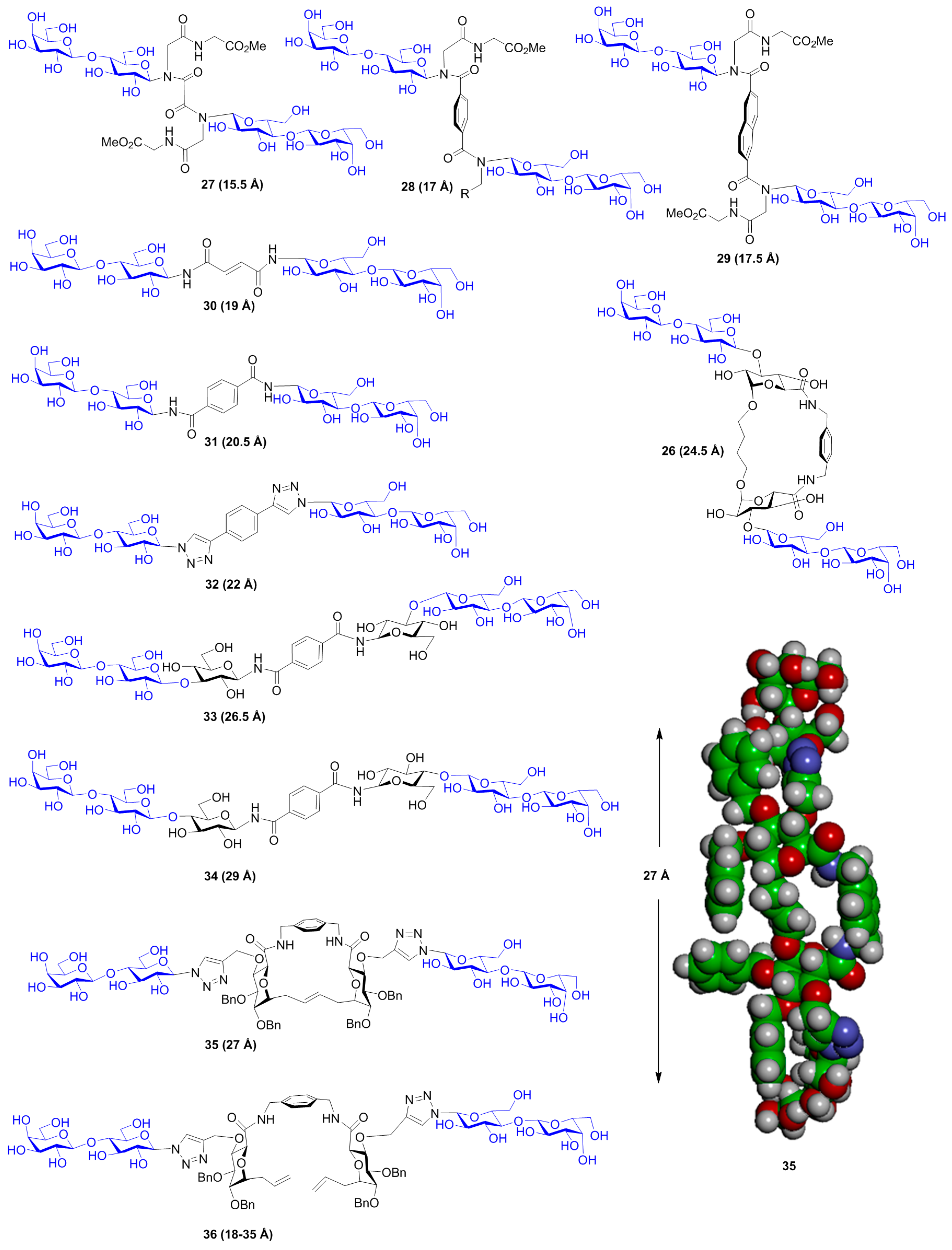
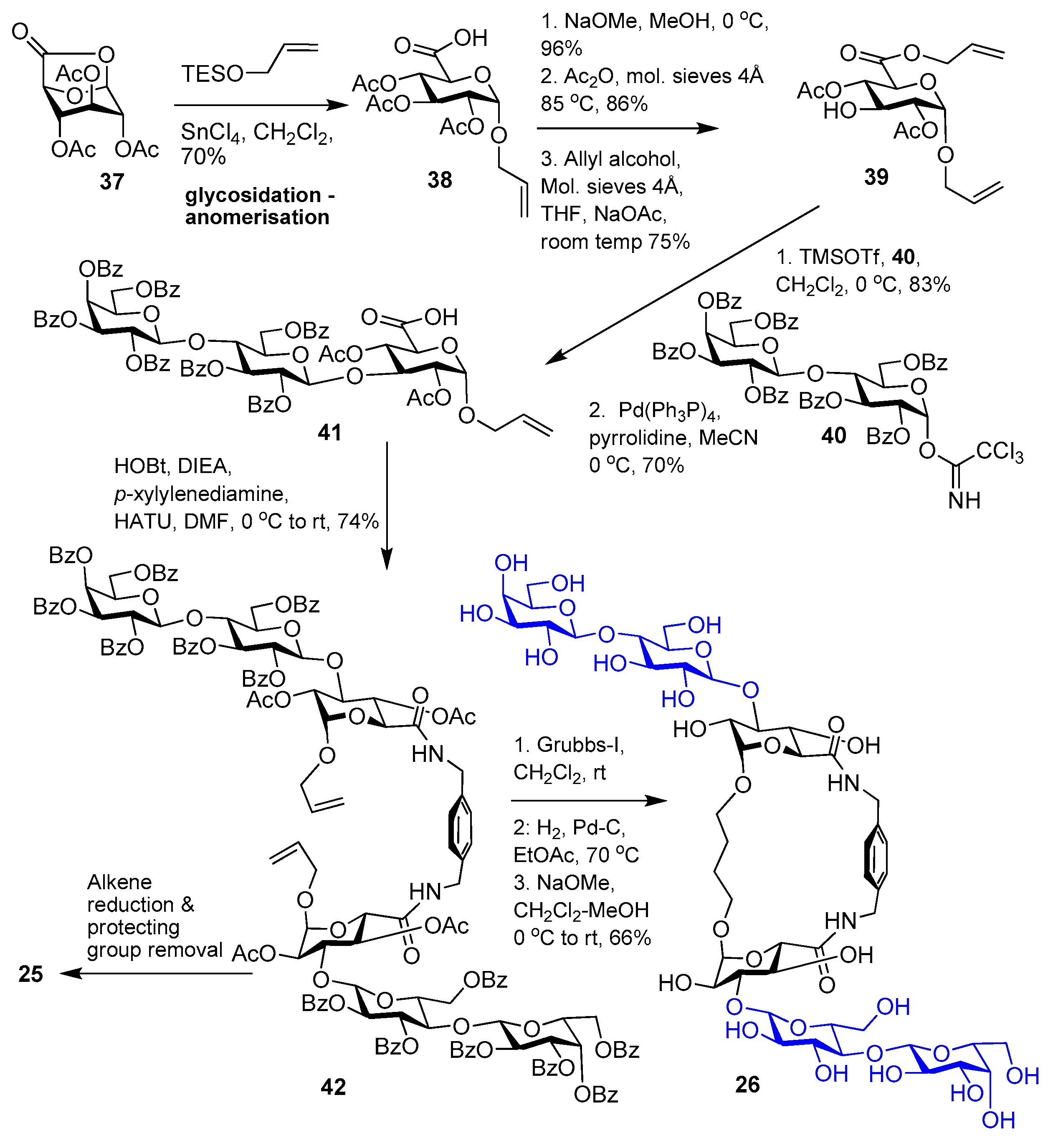
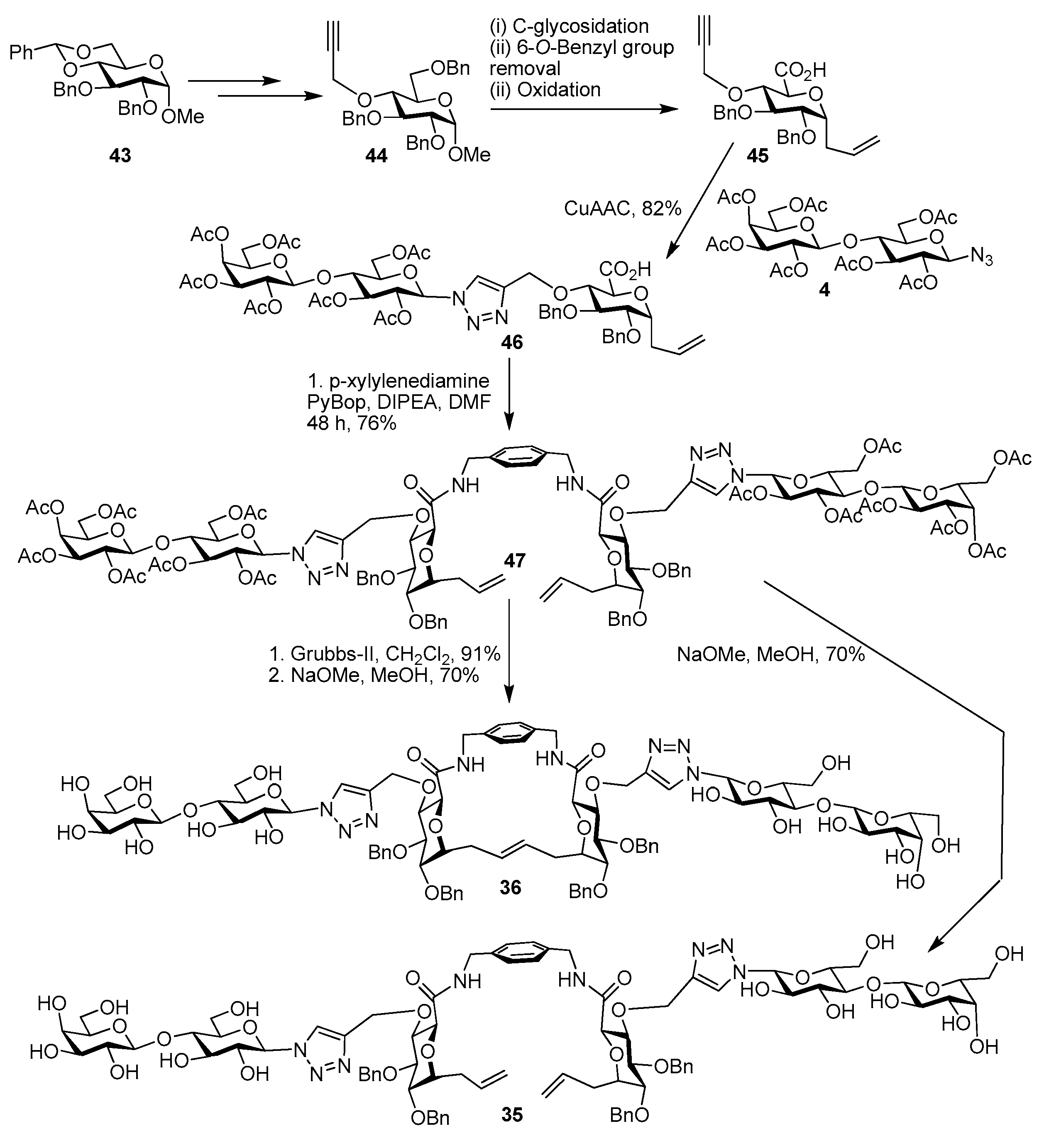
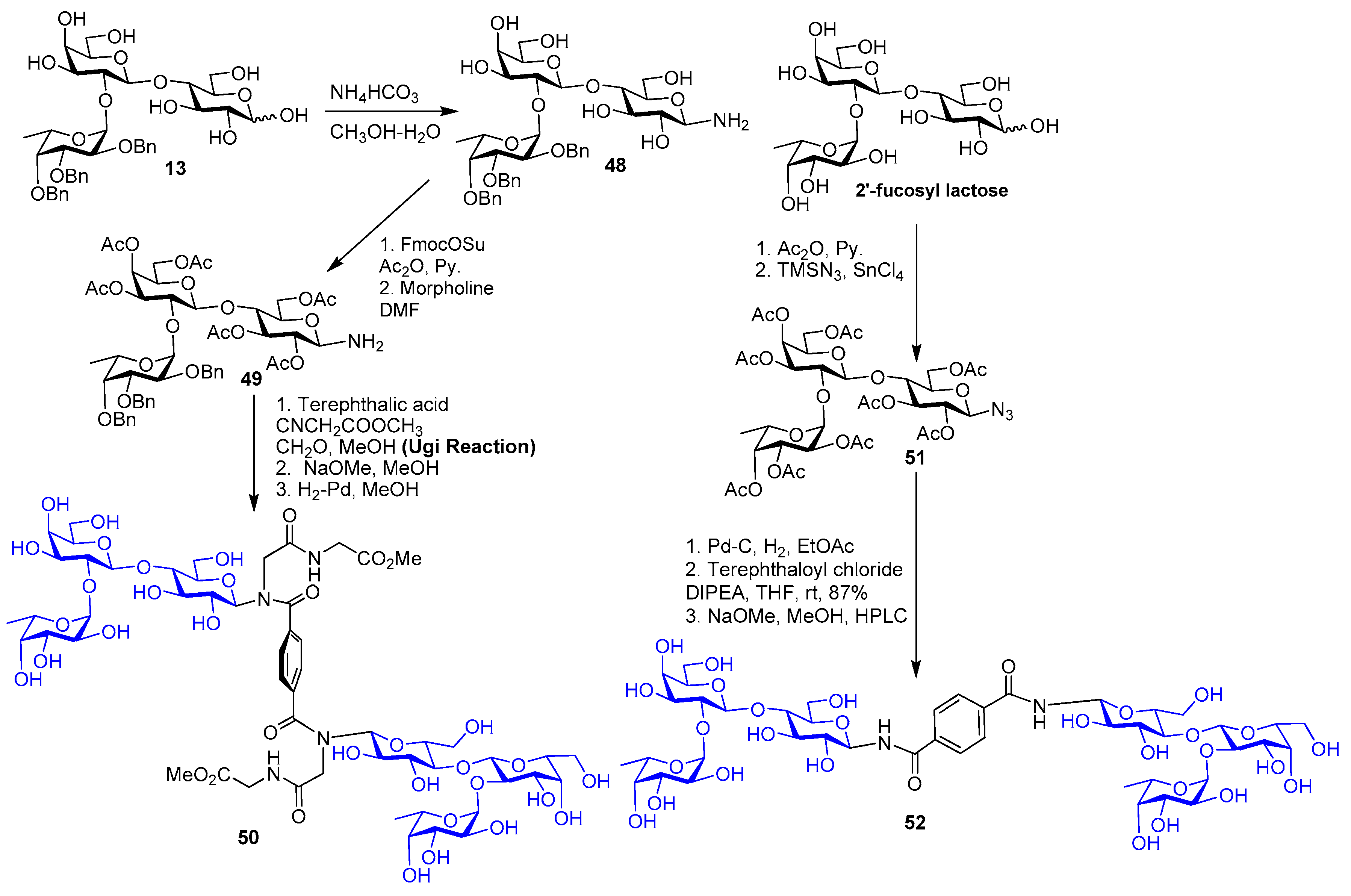
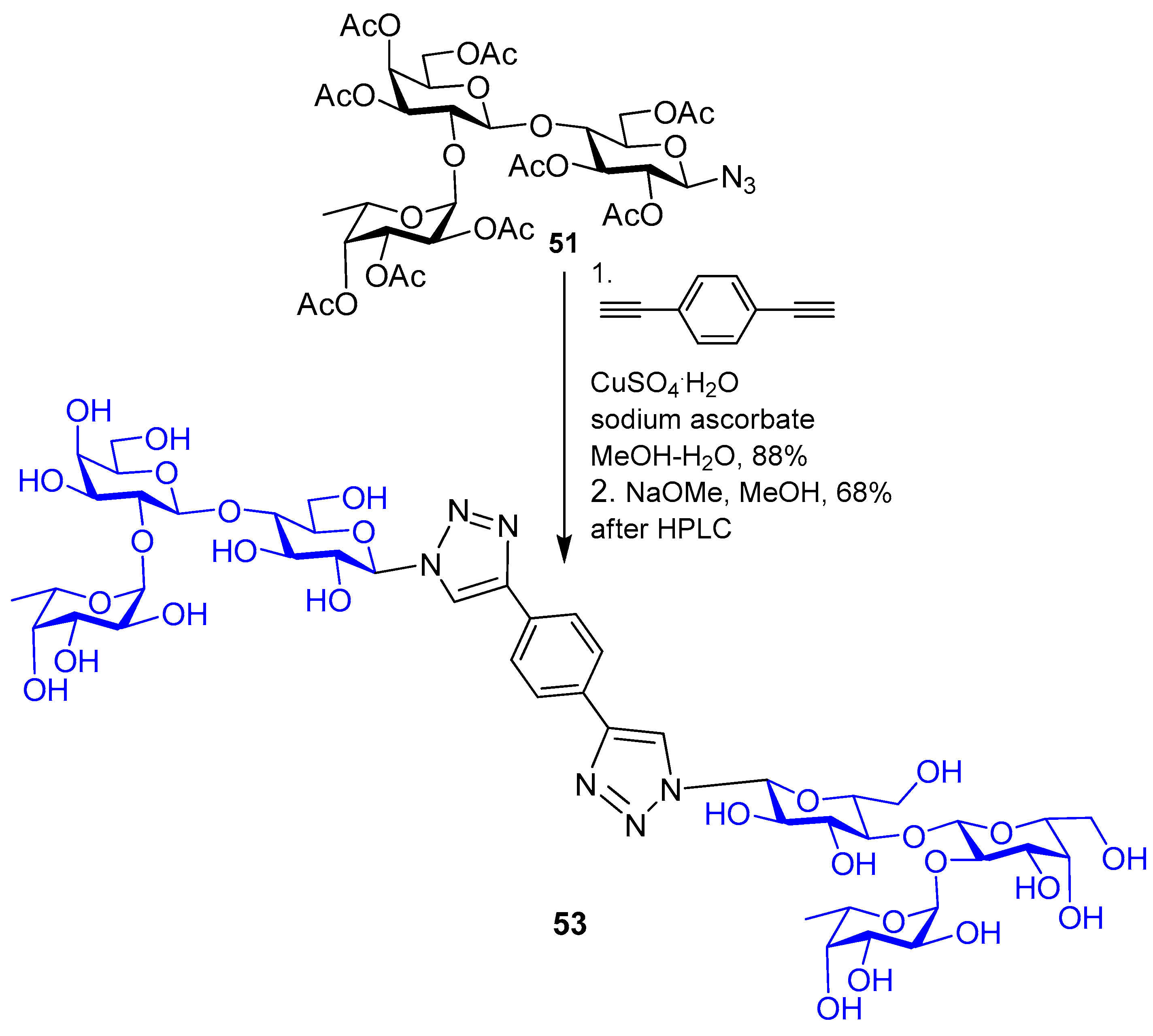
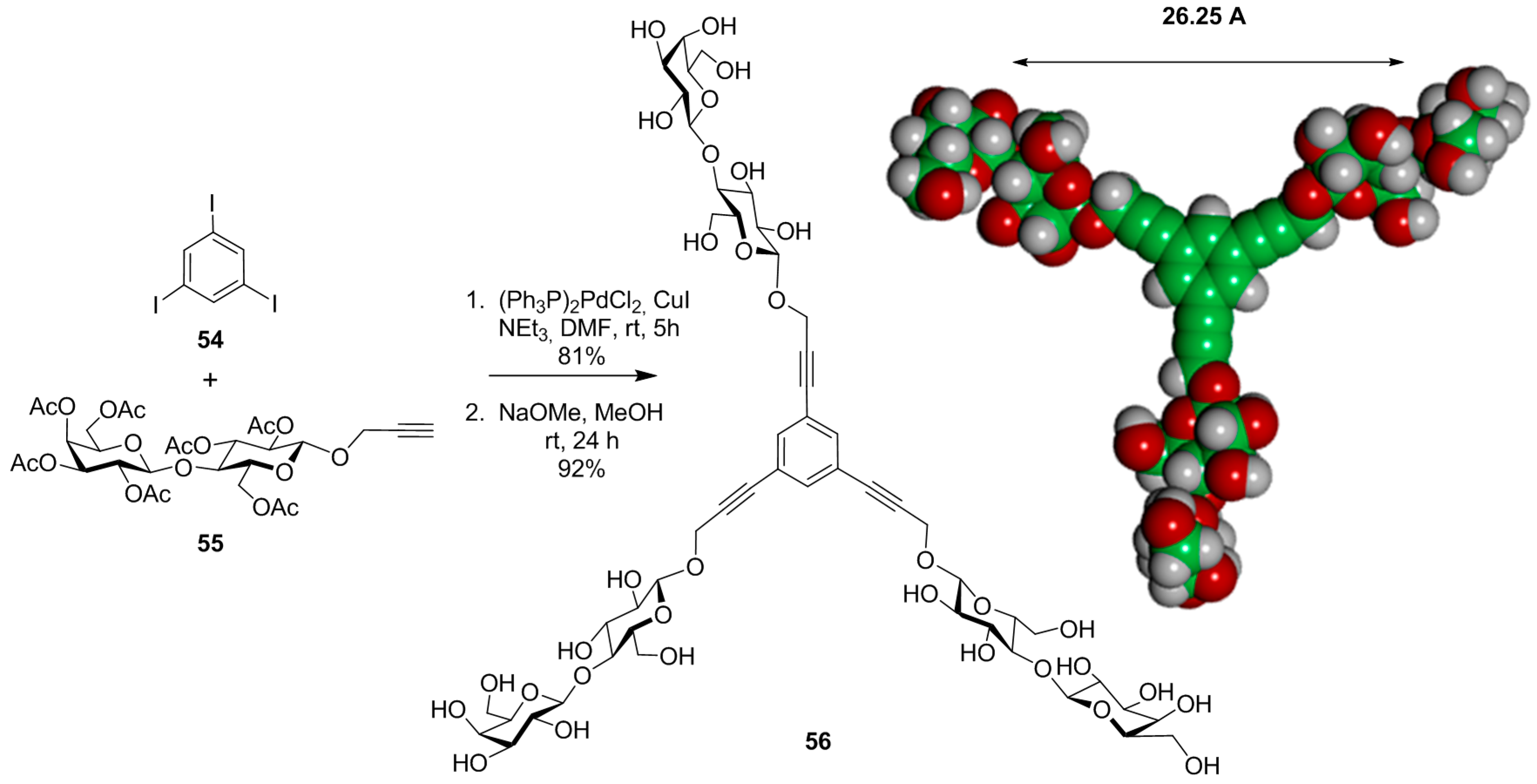
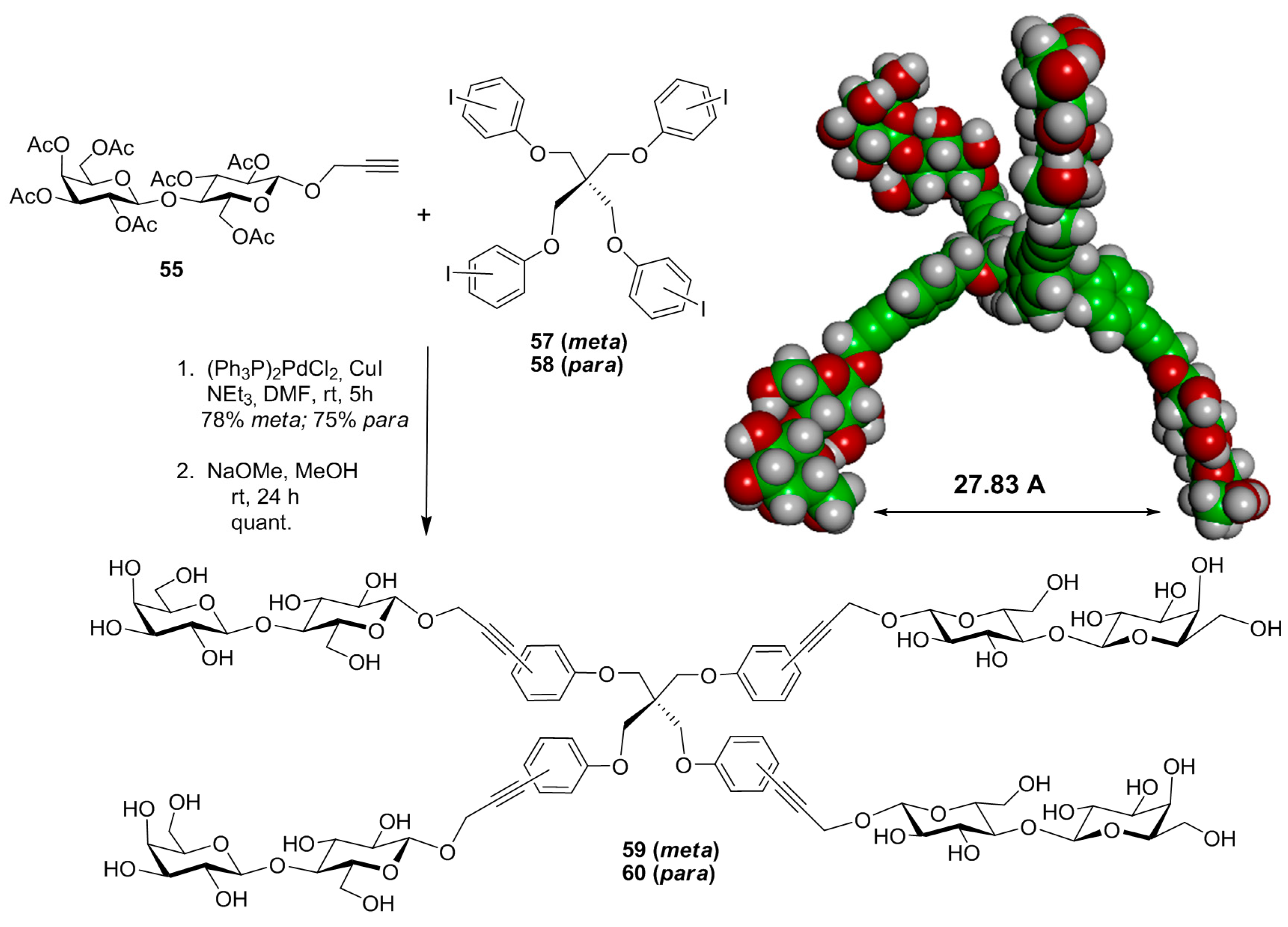
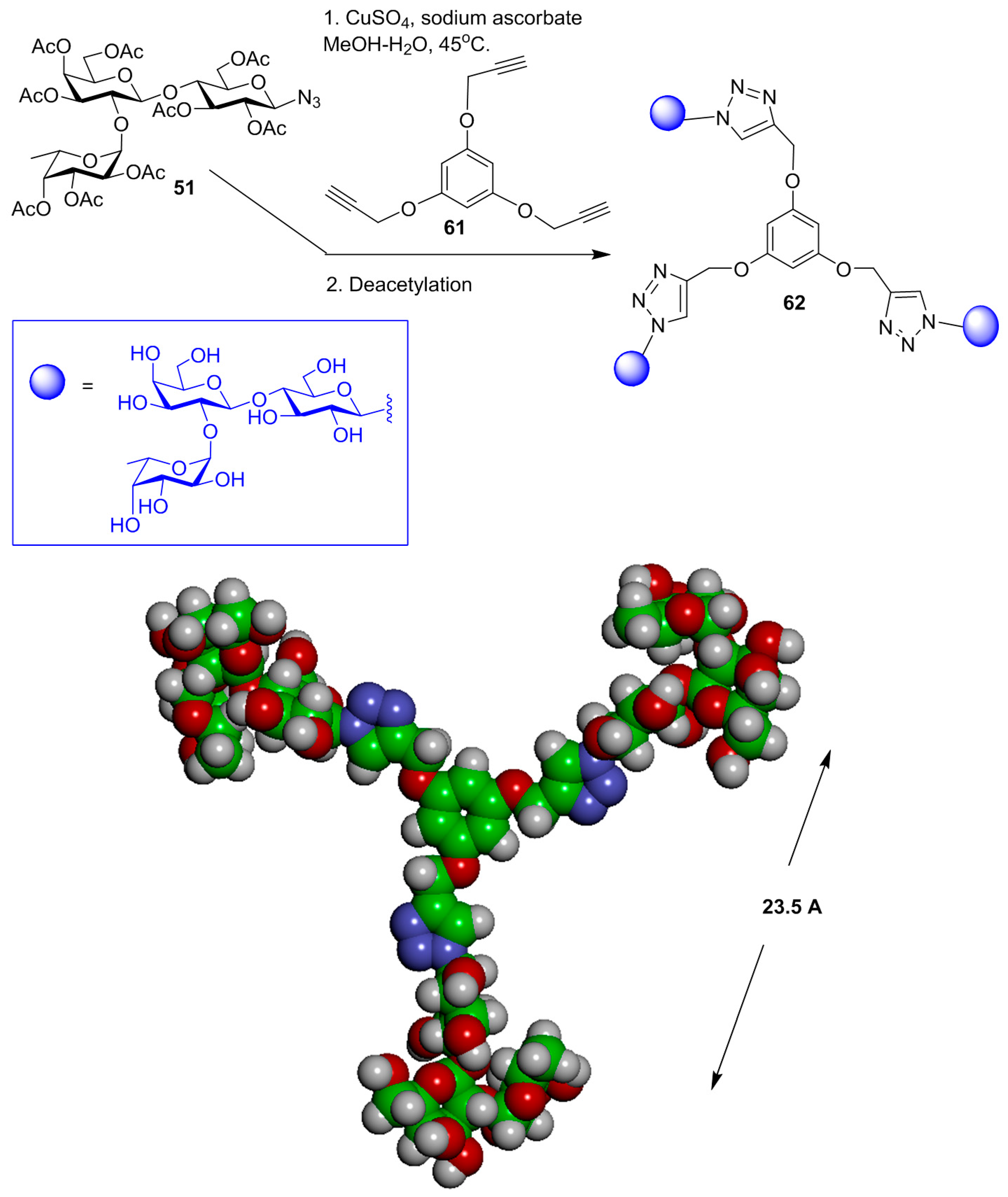
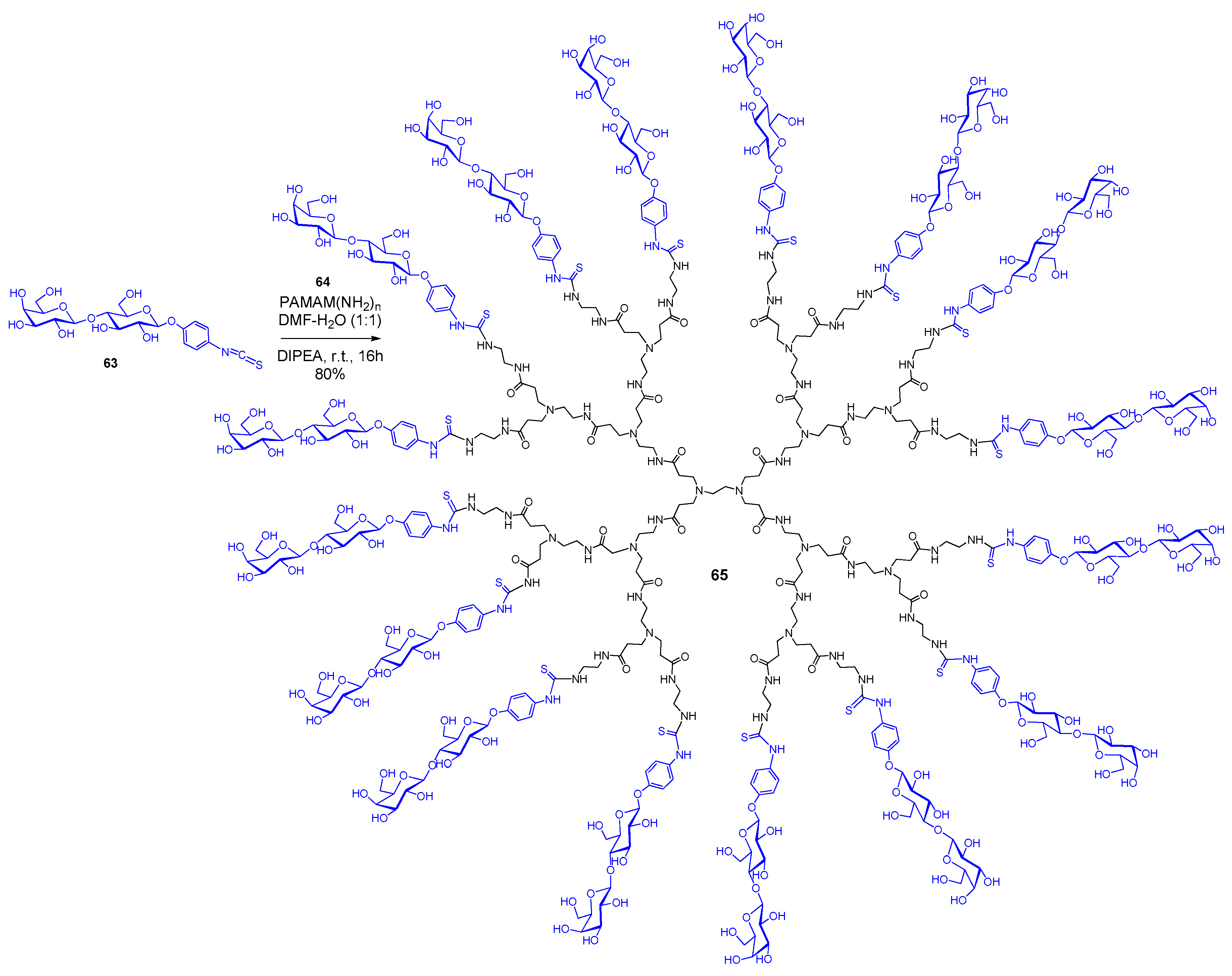
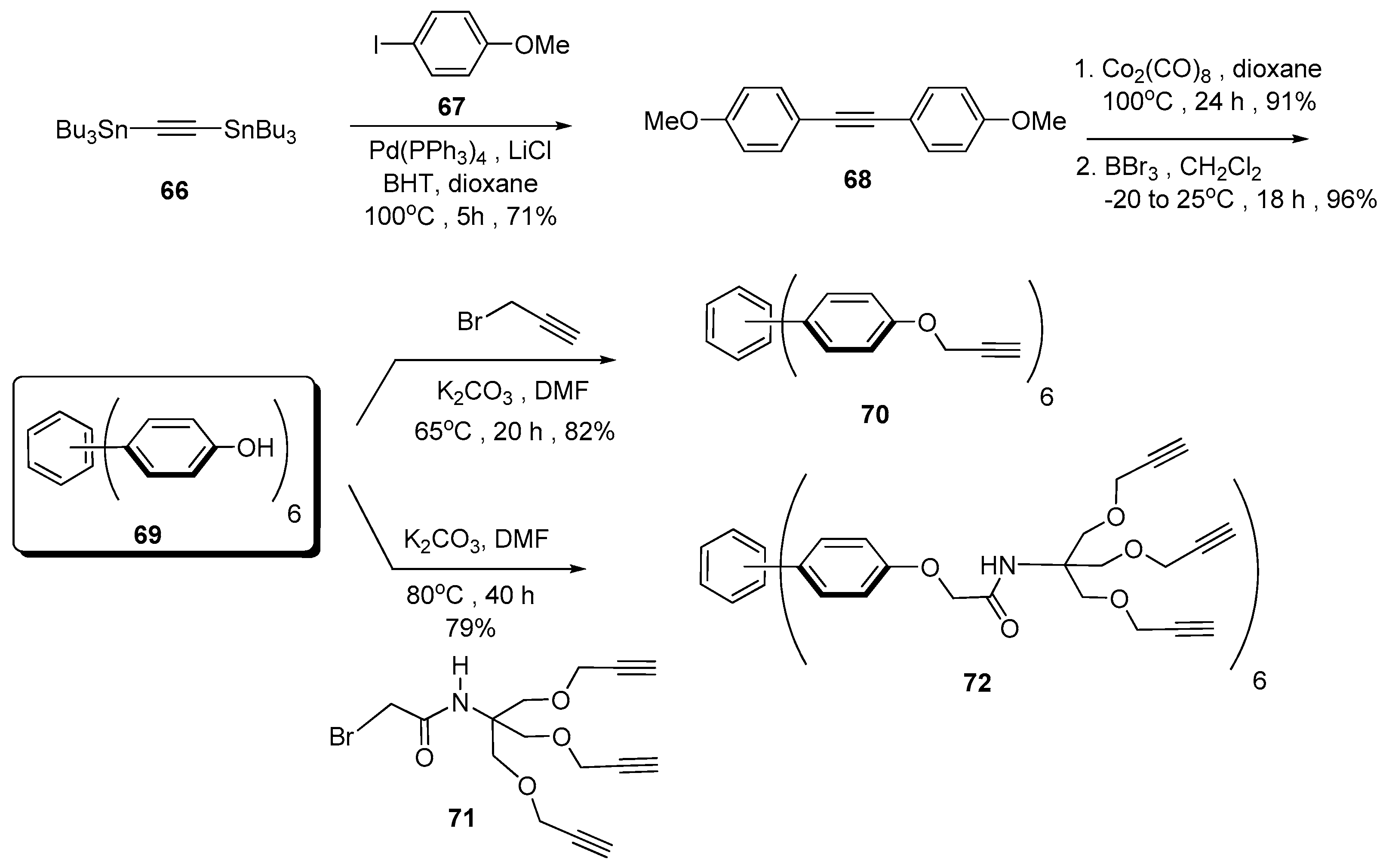
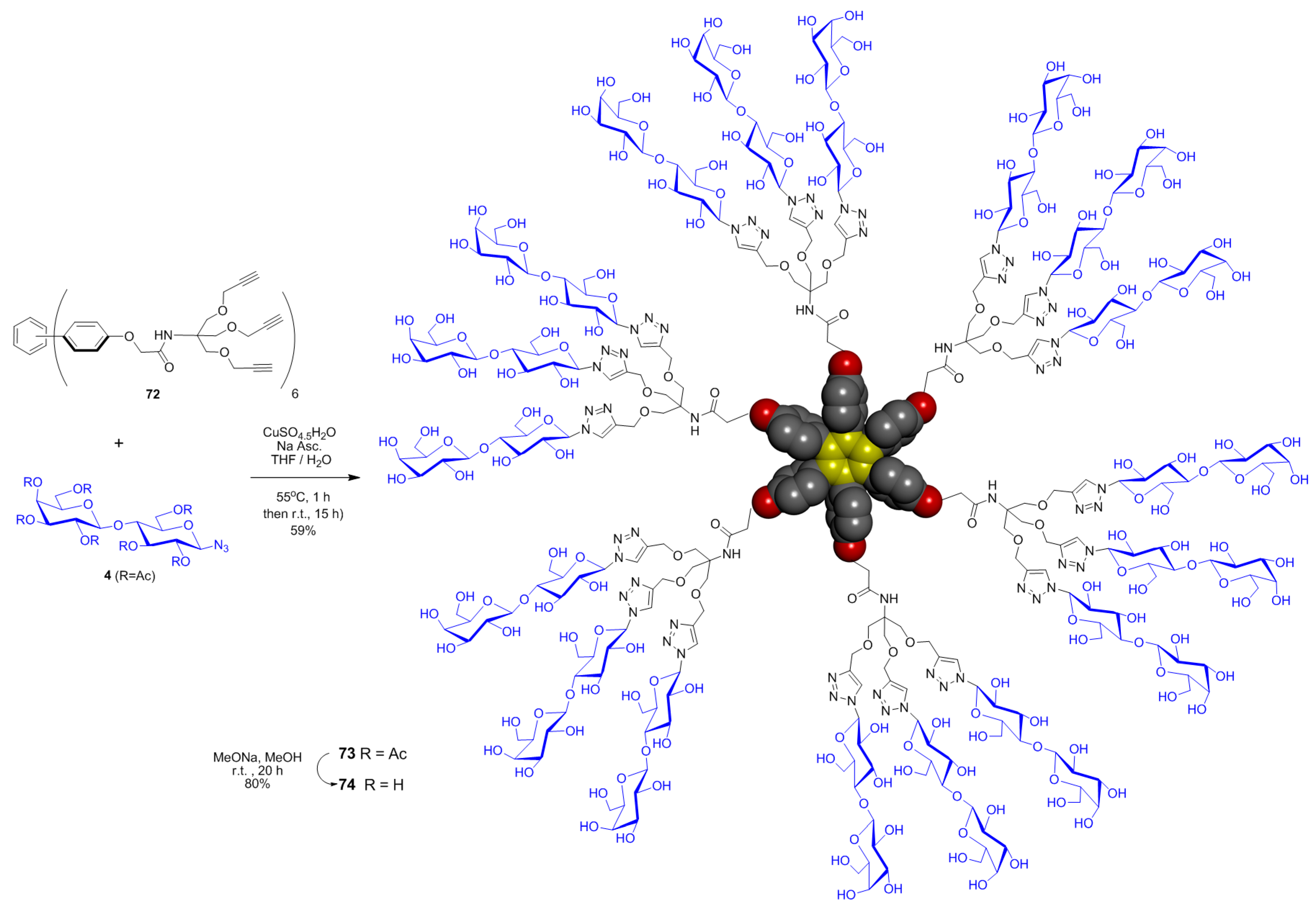



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
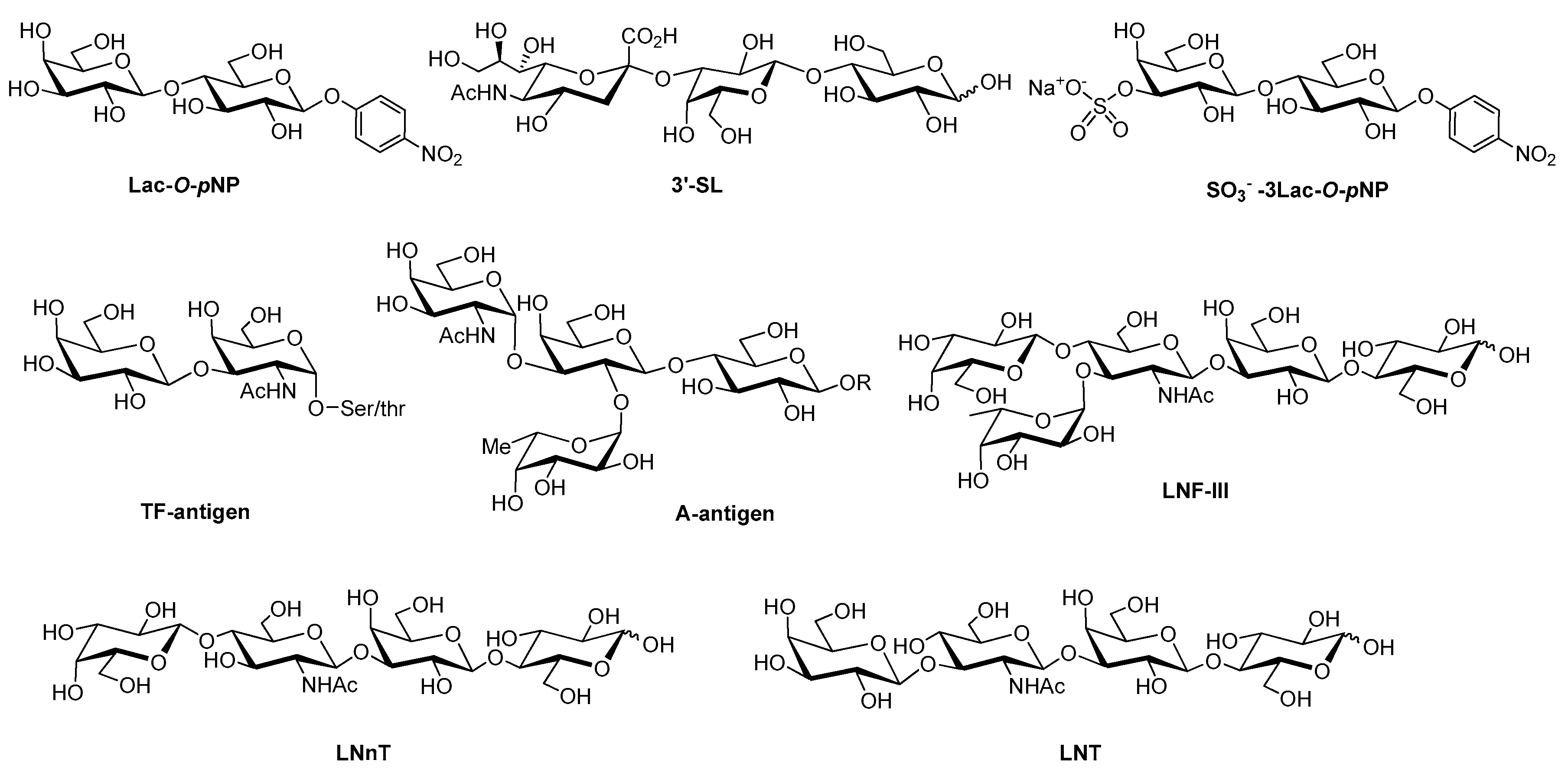{kind=link}
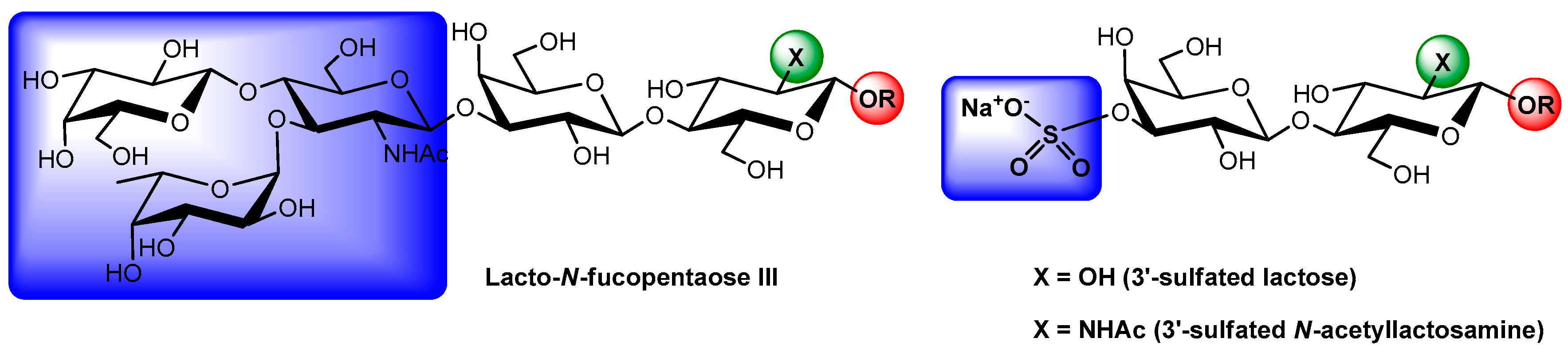{kind=link}
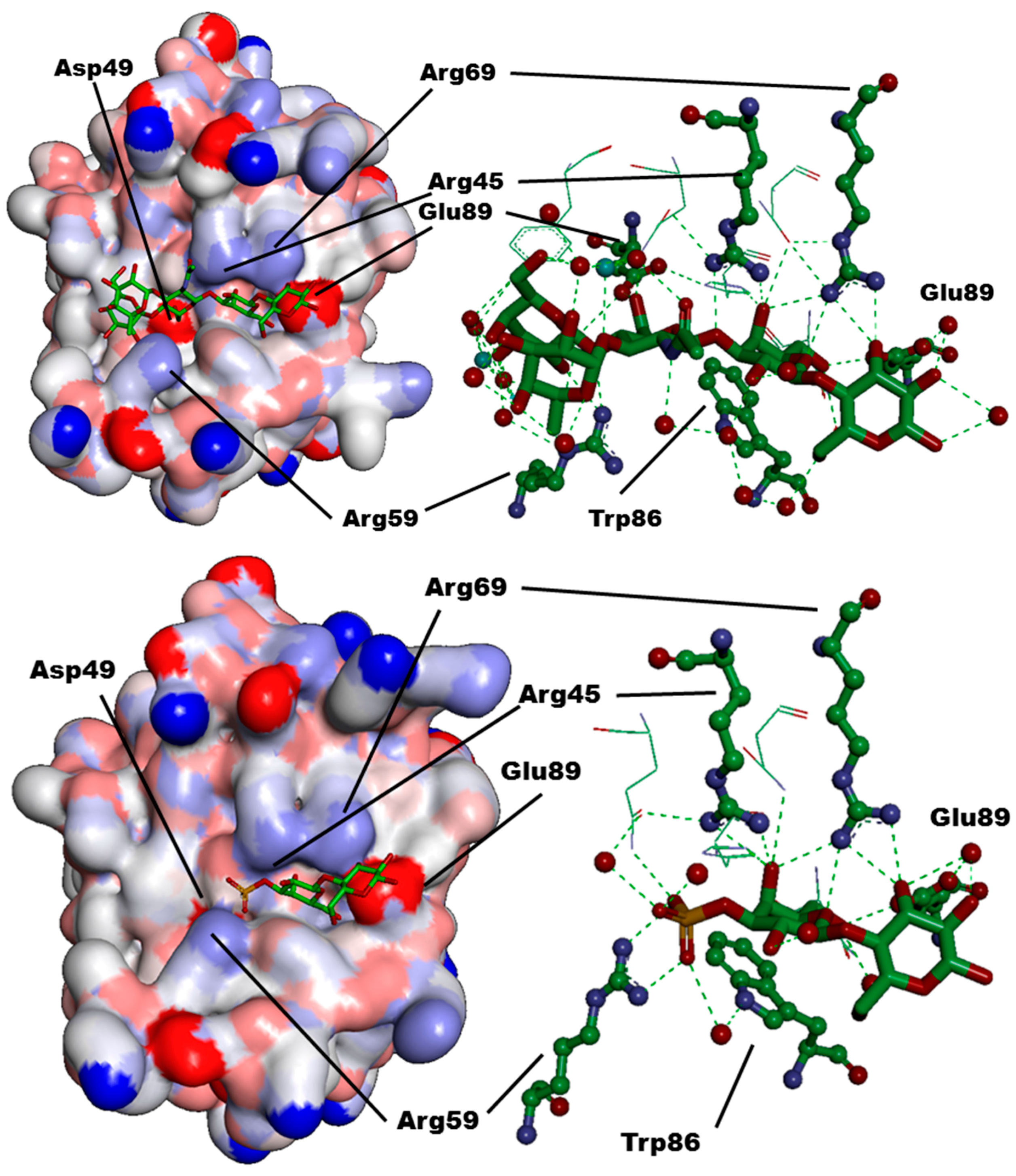{kind=link}
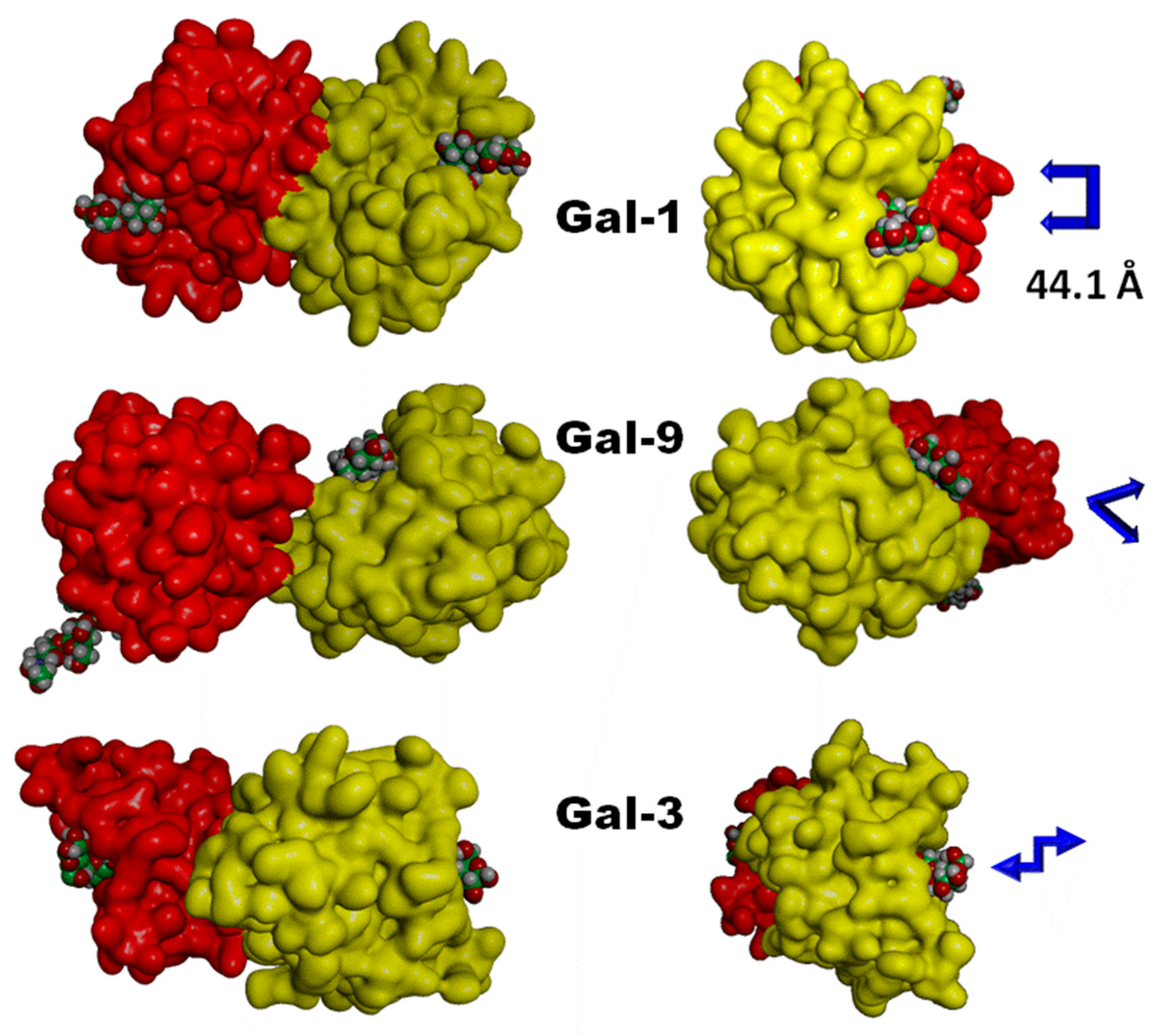{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
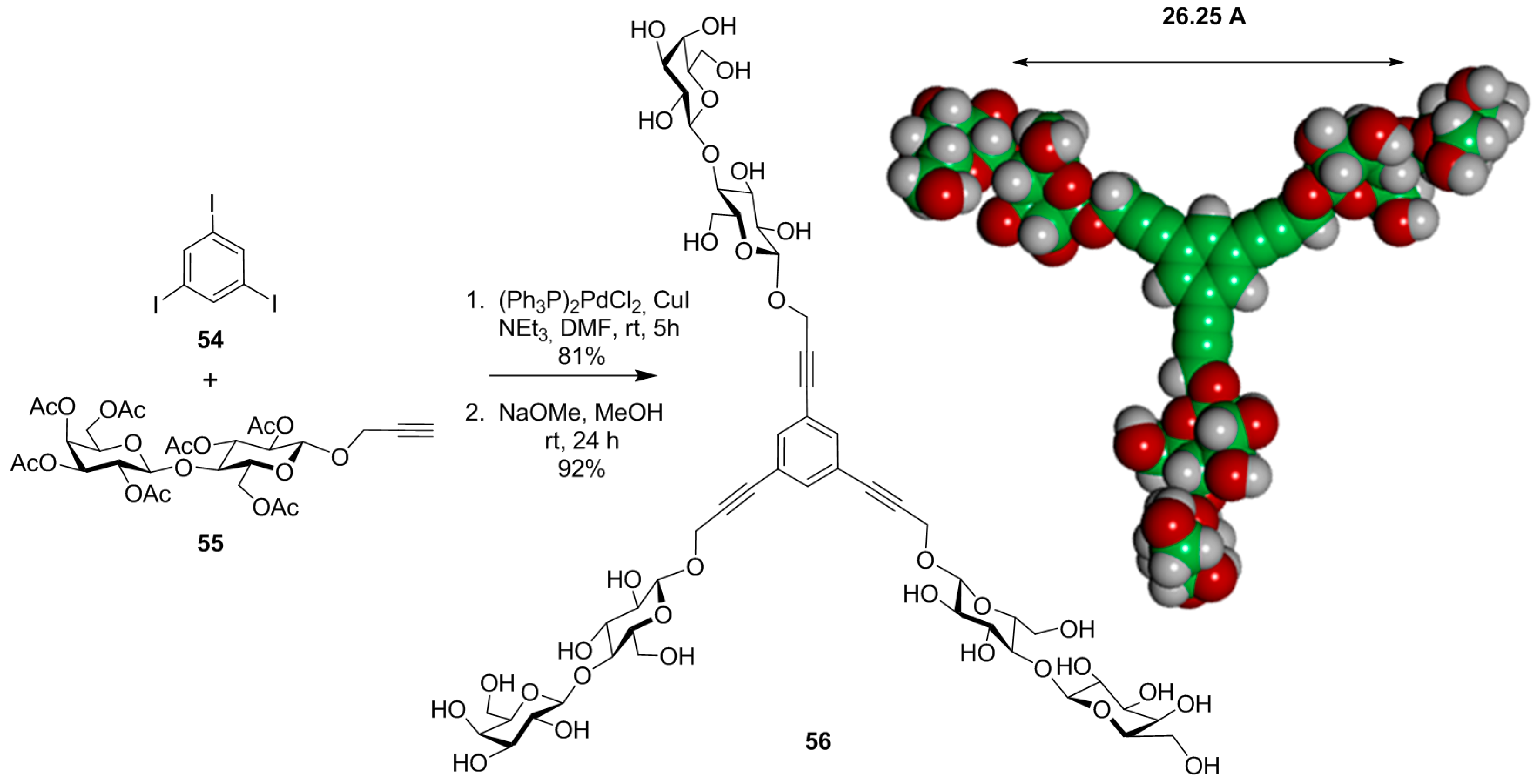{kind=link}
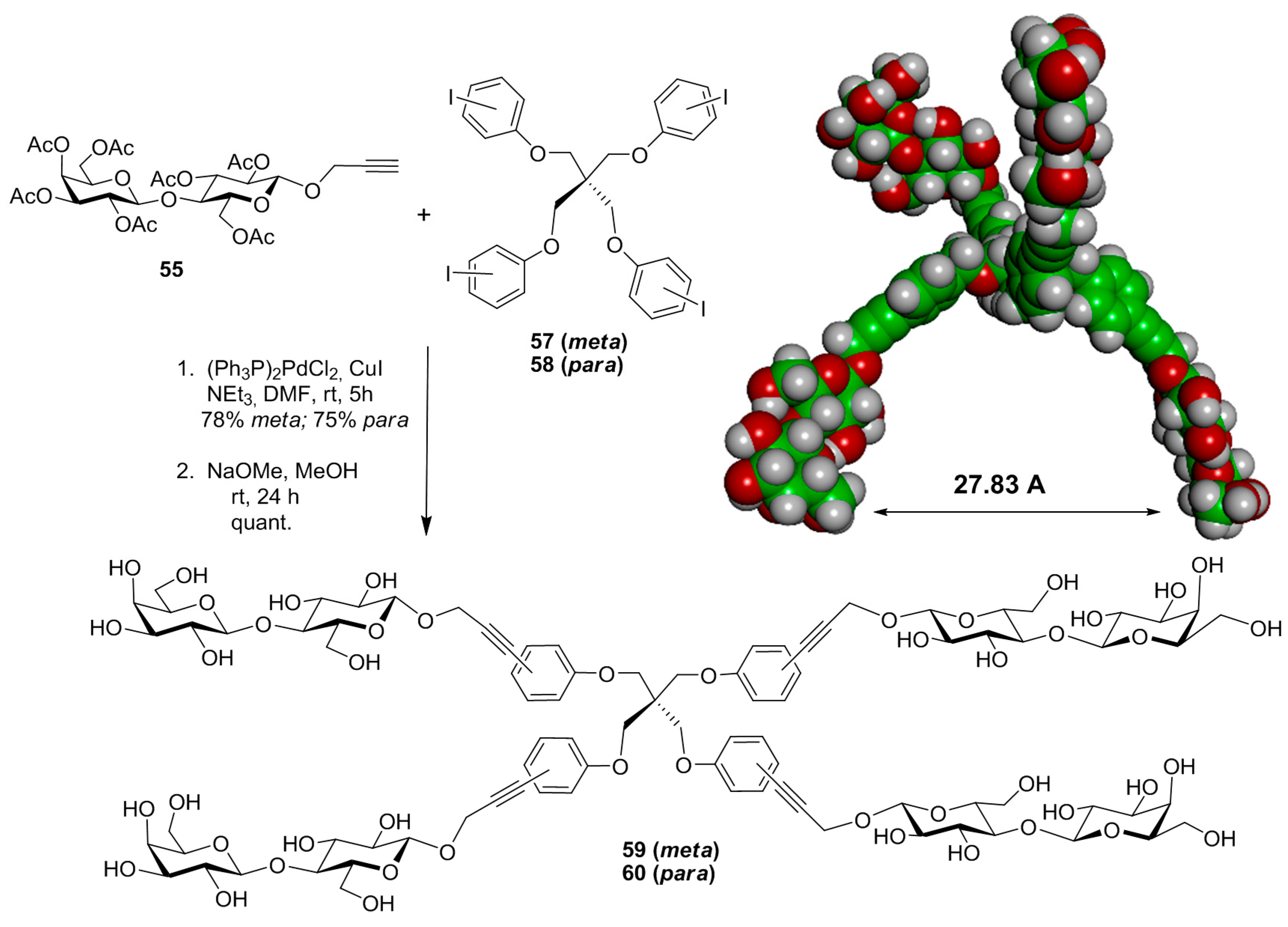{kind=link}
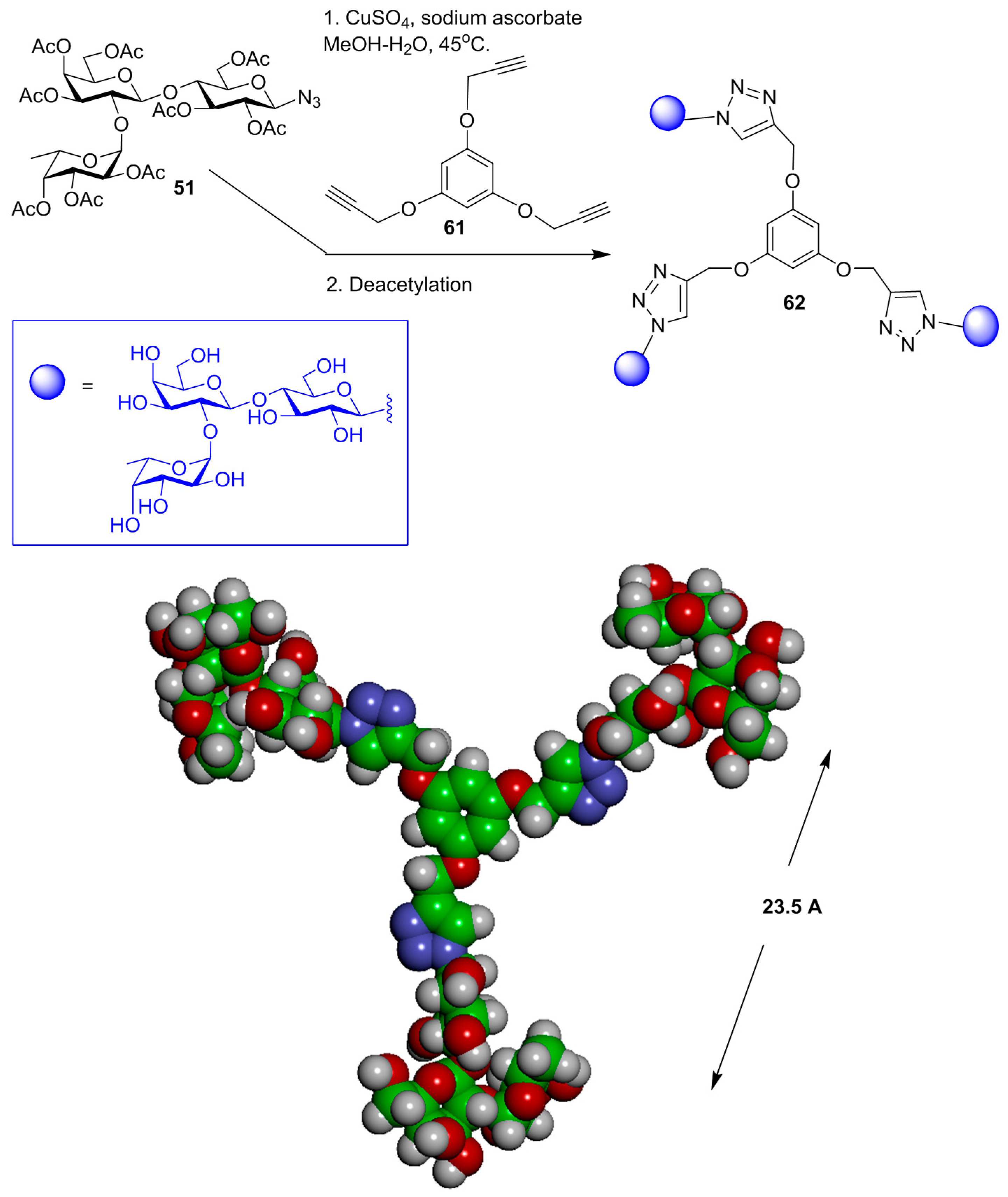{kind=link}
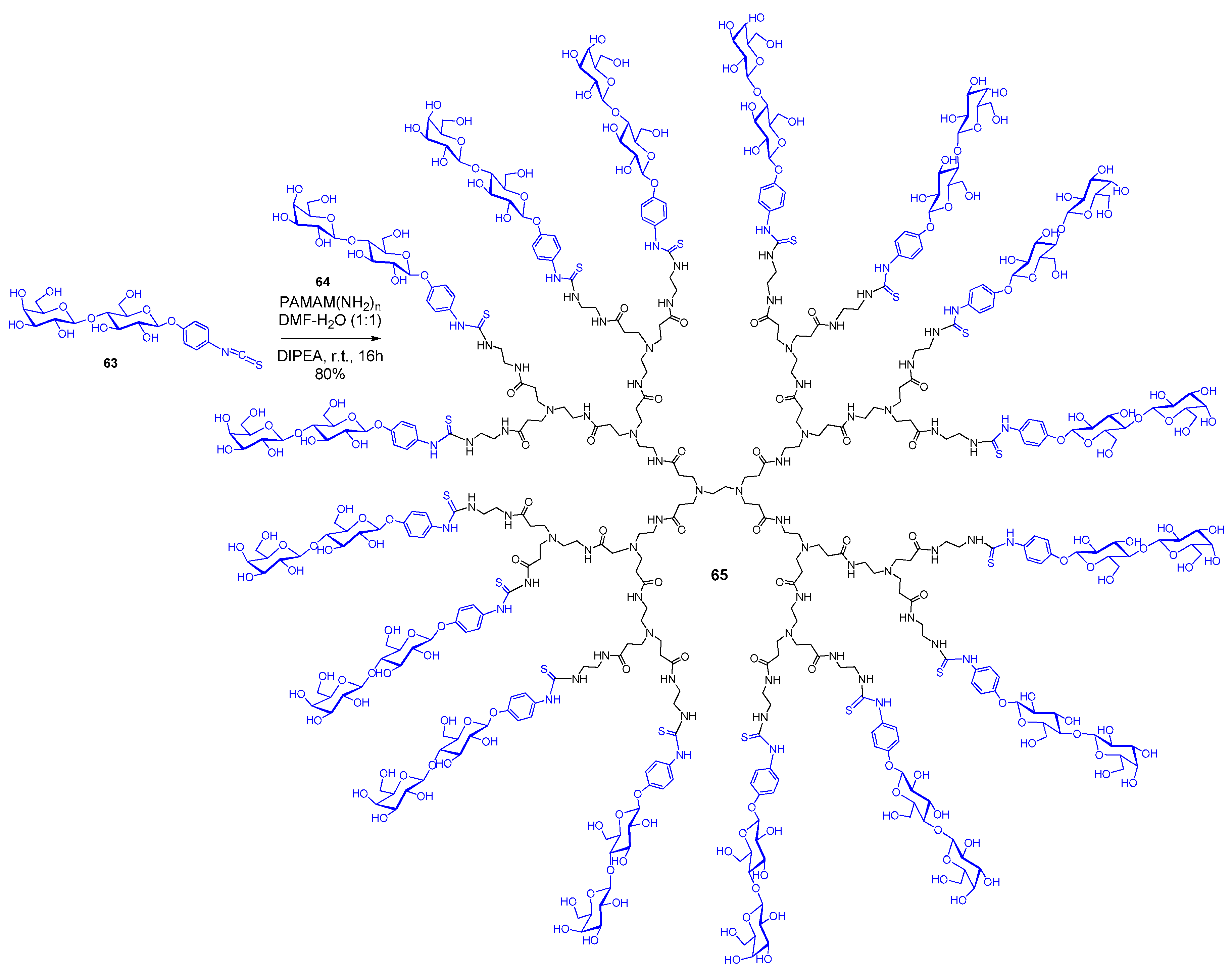{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
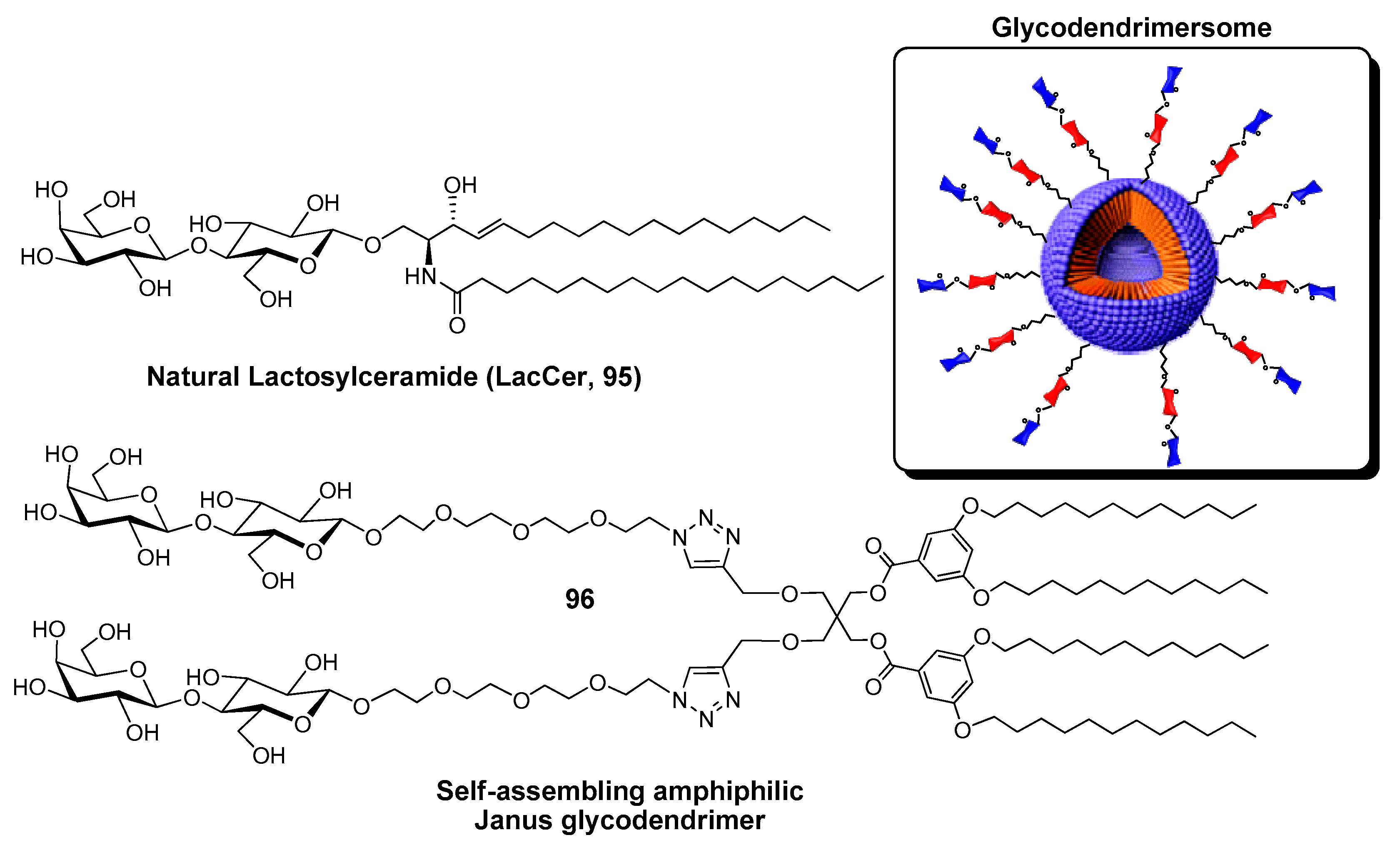{kind=link}
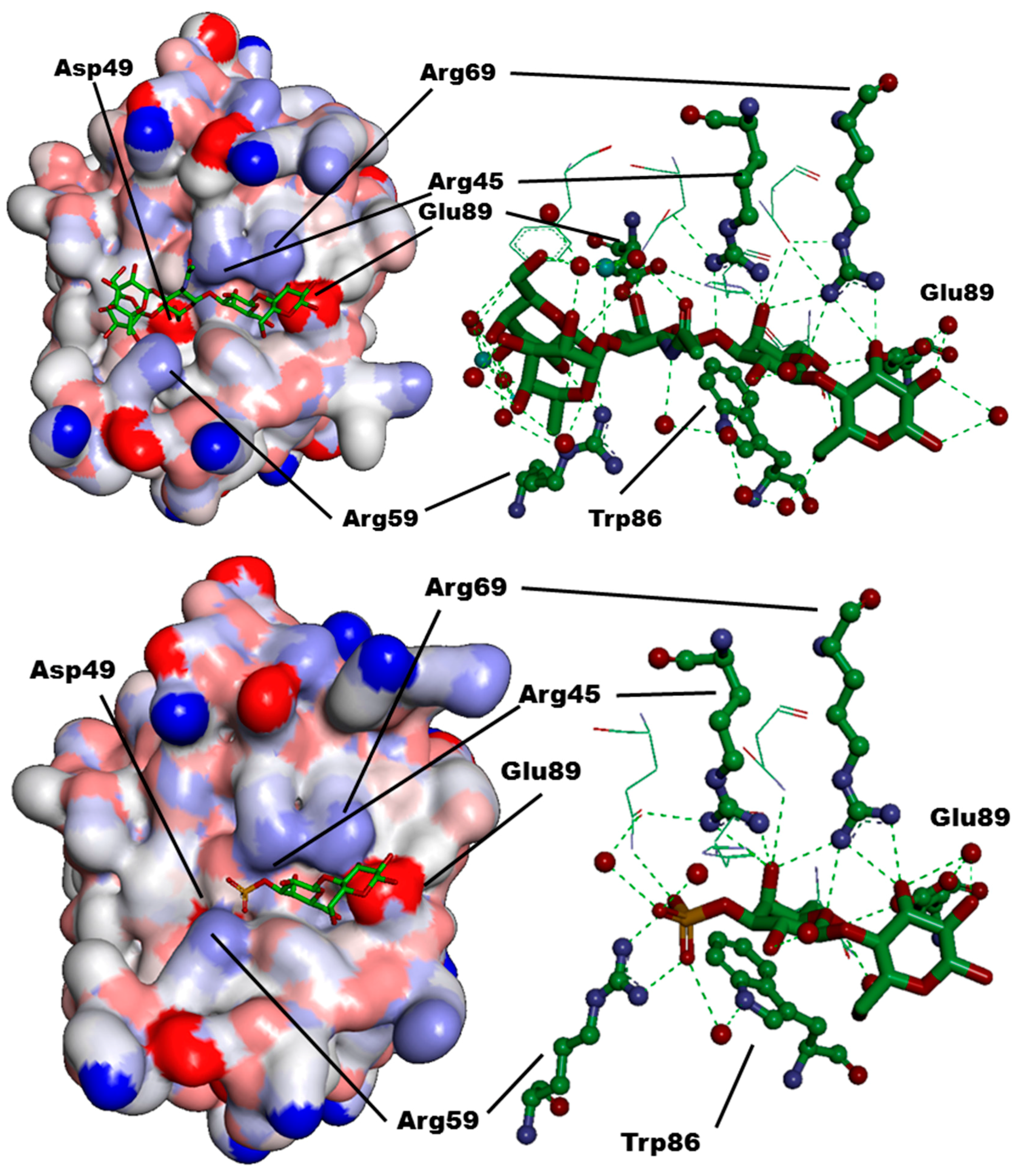
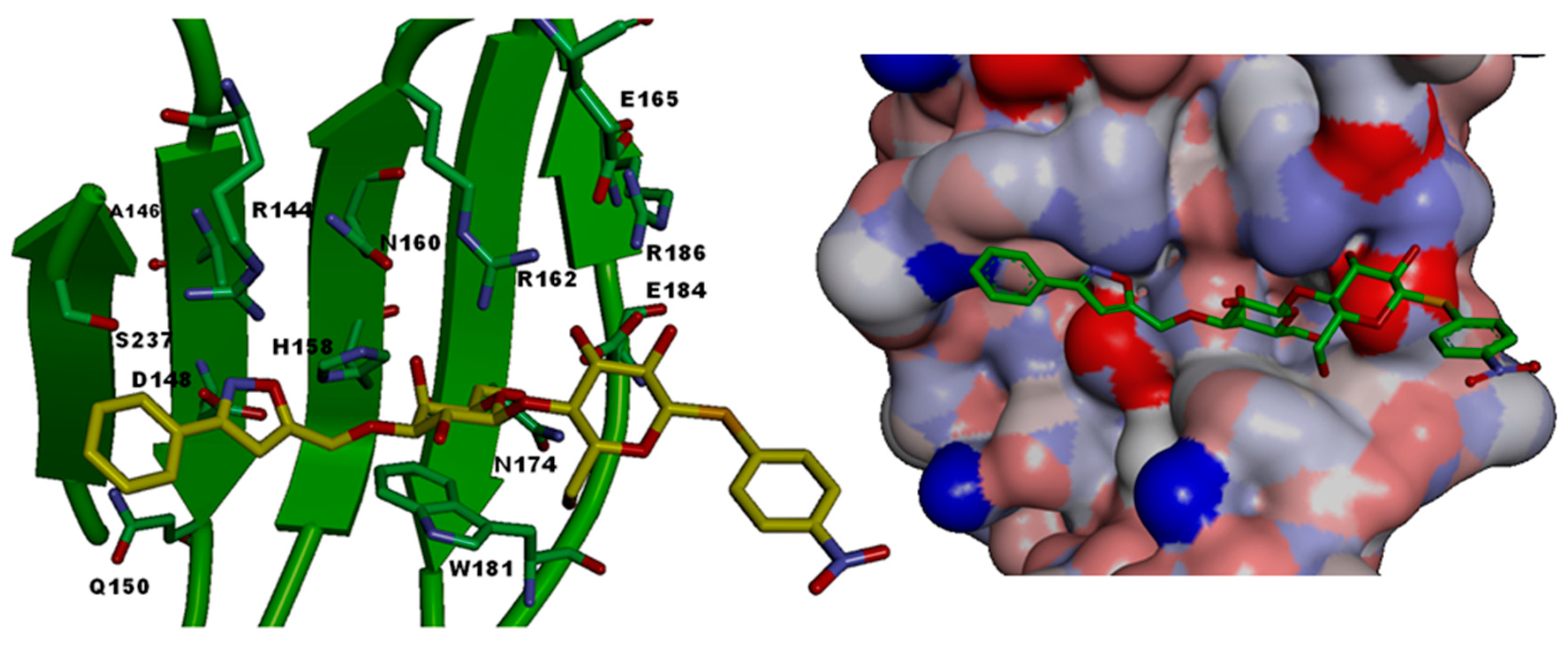
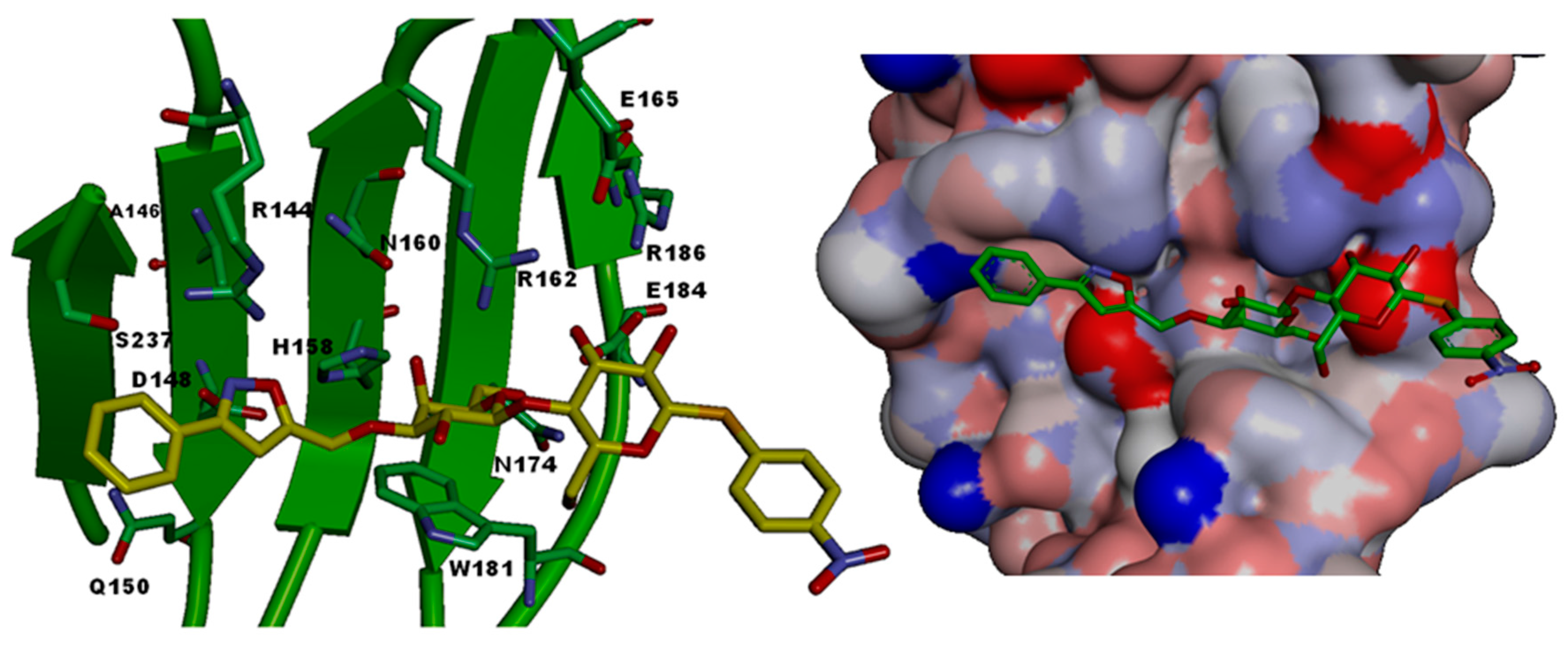
| CRD-Subsite | Galectin-1 | Galectin-2 | Galectin-3 | Galectin-4C | Galectin-7 | Galectin-8N |
|---|---|---|---|---|---|---|
| A | Ser29 | Gly30 | Arg144 (R) | Ser220 | Arg31 | Arg45 |
| B | Val31 | Val32 | Ala146 (A) | Ala222 | His33 | Gln47 |
| C | Asn33 | Asn34 | Asp148 (D) | Asn224 | Asn35 | Asp49 |
| D | Gly35 | Gly36 | Gln150 (Q) | Lys226 | Leu37 | Gln51 |
| E | - | - | - | - | - | Arg59 |
| F | His44 | His45 | His158 (H) | His236 | His49 | His65 |
| G | Asn46 | Asn47 | Asn160 (N) | Asn238 | Asn51 | Asn67 |
| H | Arg48 | Arg49 | Arg162 (R) | Arg240 | Arg53 | Arg69 |
| I | His52 | - | - | - | - | - |
| J | Asp54 | Gln52 | Glu165 (E) | - | - | - |
| K | Asn61 | Asn58 | Asn174 (N) | Asn249 | Asn62 | Asn79 |
| L | Trp68 | Trp65 | Trp181 (W) | Trp256 | Trp69 | Trp86 |
| M | Glu71 | Glu68 | Glu184 (E) | Glu259 | Glu72 | Glu89 |
| N | Arg73 | Arg70 | Arg186 (R) | Lys261 | Arg74 | Ile91 |
| O | - | Arg120 | Ser237 (S) | Gln313 | Gly124 | Tyr141 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, R.; Murphy, P.V.; Gabius, H.-J. Multivalent Carbohydrate-Lectin Interactions: How Synthetic Chemistry Enables Insights into Nanometric Recognition. Molecules 2016, 21, 629. https://doi.org/10.3390/molecules21050629
Roy R, Murphy PV, Gabius H-J. Multivalent Carbohydrate-Lectin Interactions: How Synthetic Chemistry Enables Insights into Nanometric Recognition. Molecules. 2016; 21(5):629. https://doi.org/10.3390/molecules21050629
Chicago/Turabian StyleRoy, René, Paul V. Murphy, and Hans-Joachim Gabius. 2016. "Multivalent Carbohydrate-Lectin Interactions: How Synthetic Chemistry Enables Insights into Nanometric Recognition" Molecules 21, no. 5: 629. https://doi.org/10.3390/molecules21050629
APA StyleRoy, R., Murphy, P. V., & Gabius, H.-J. (2016). Multivalent Carbohydrate-Lectin Interactions: How Synthetic Chemistry Enables Insights into Nanometric Recognition. Molecules, 21(5), 629. https://doi.org/10.3390/molecules21050629