
Aberrant Expression of Bacterial Pattern Recognition Receptor NOD2 of Basophils and Microbicidal Peptides in Atopic Dermatitis

Abstract
:1. Introduction
2. Results and Discussion
2.1. AD Patients and Control Subjects
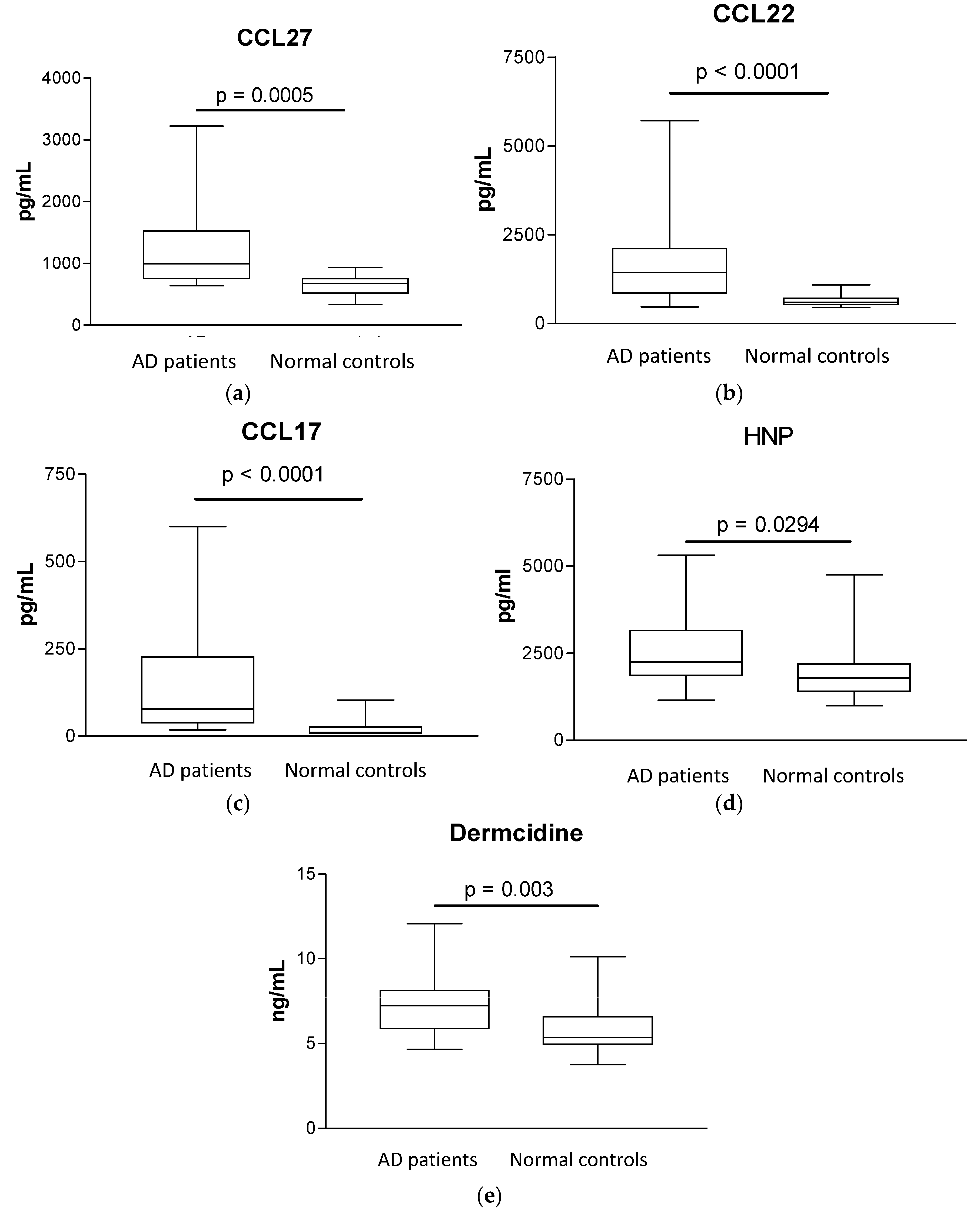
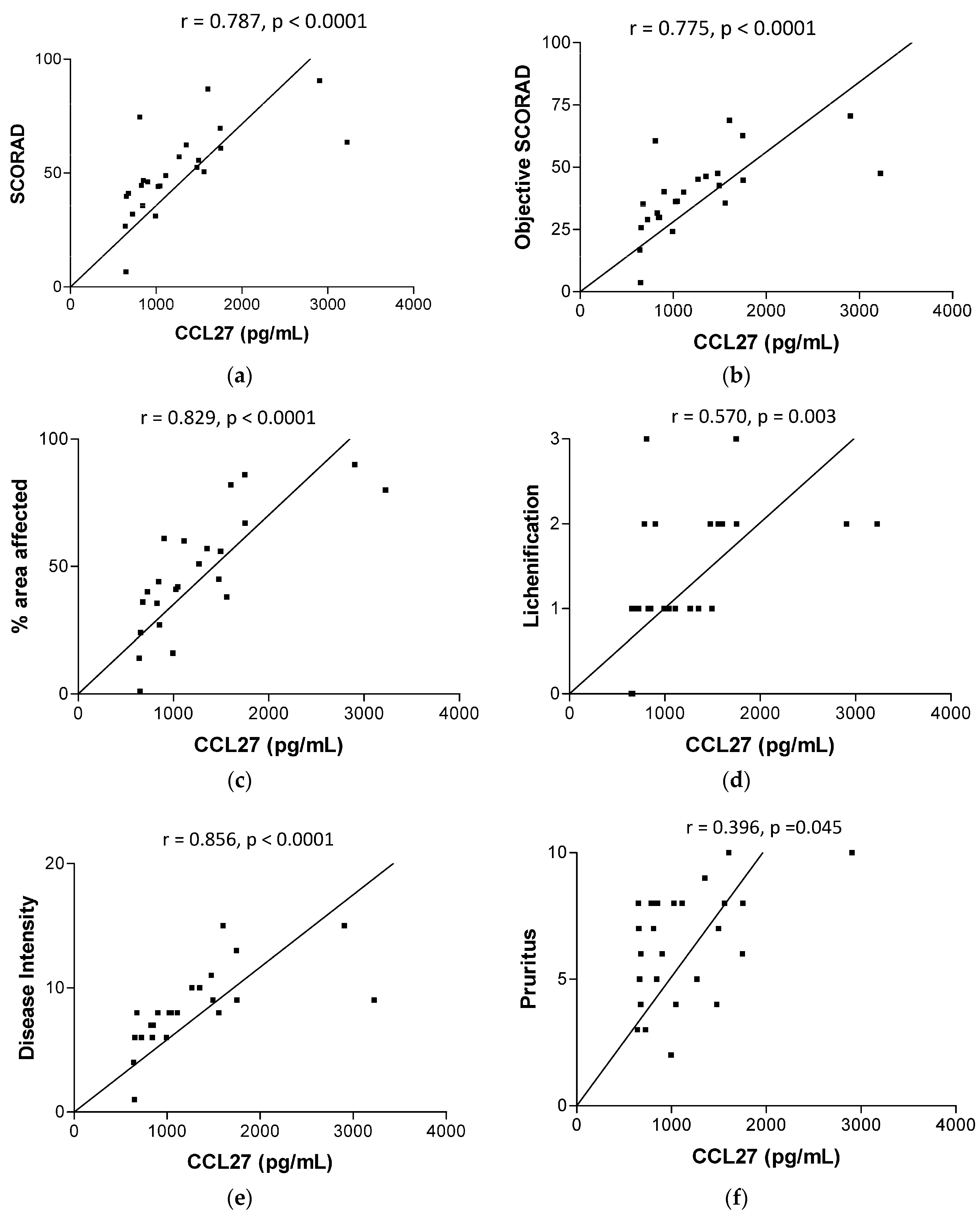
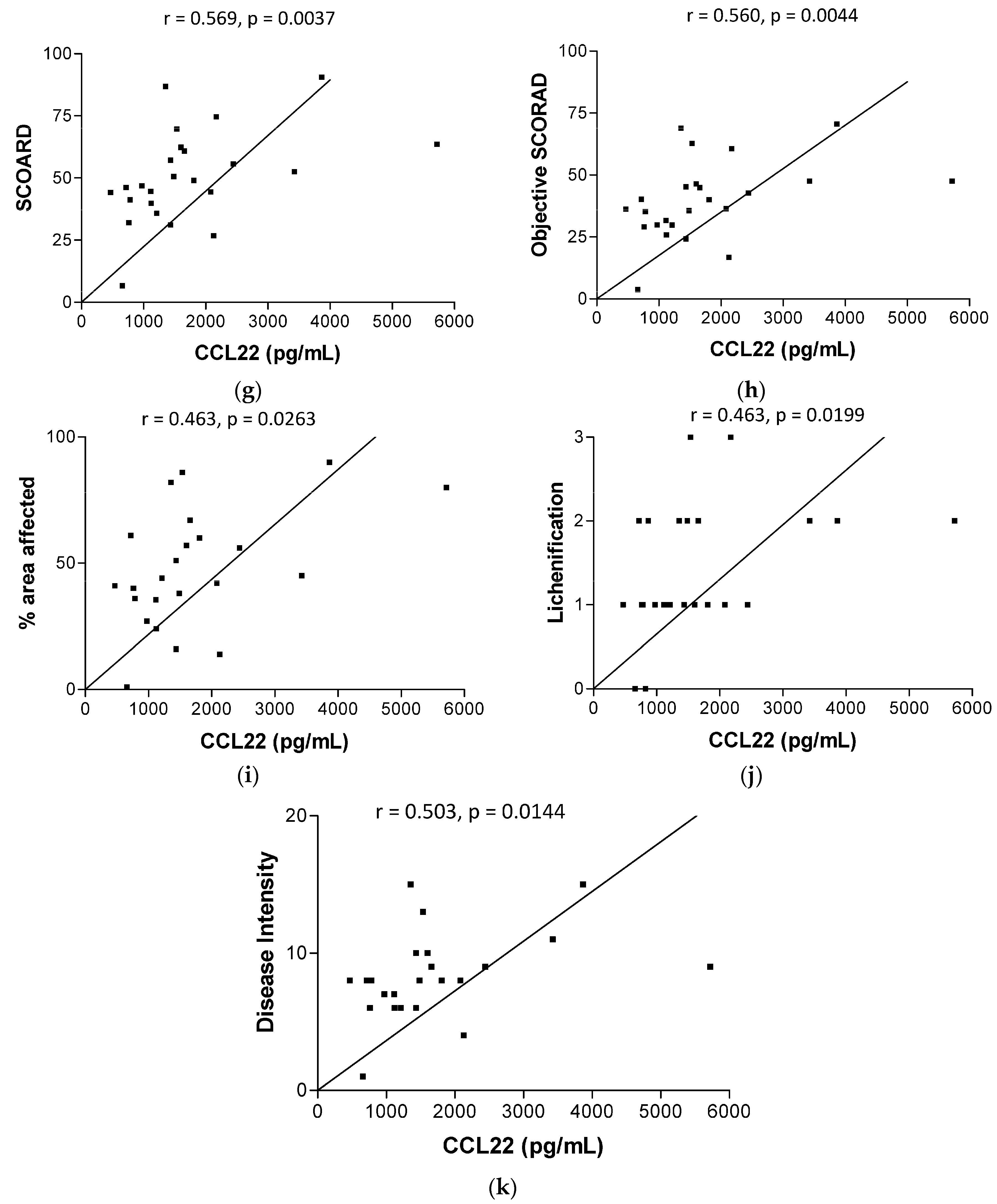
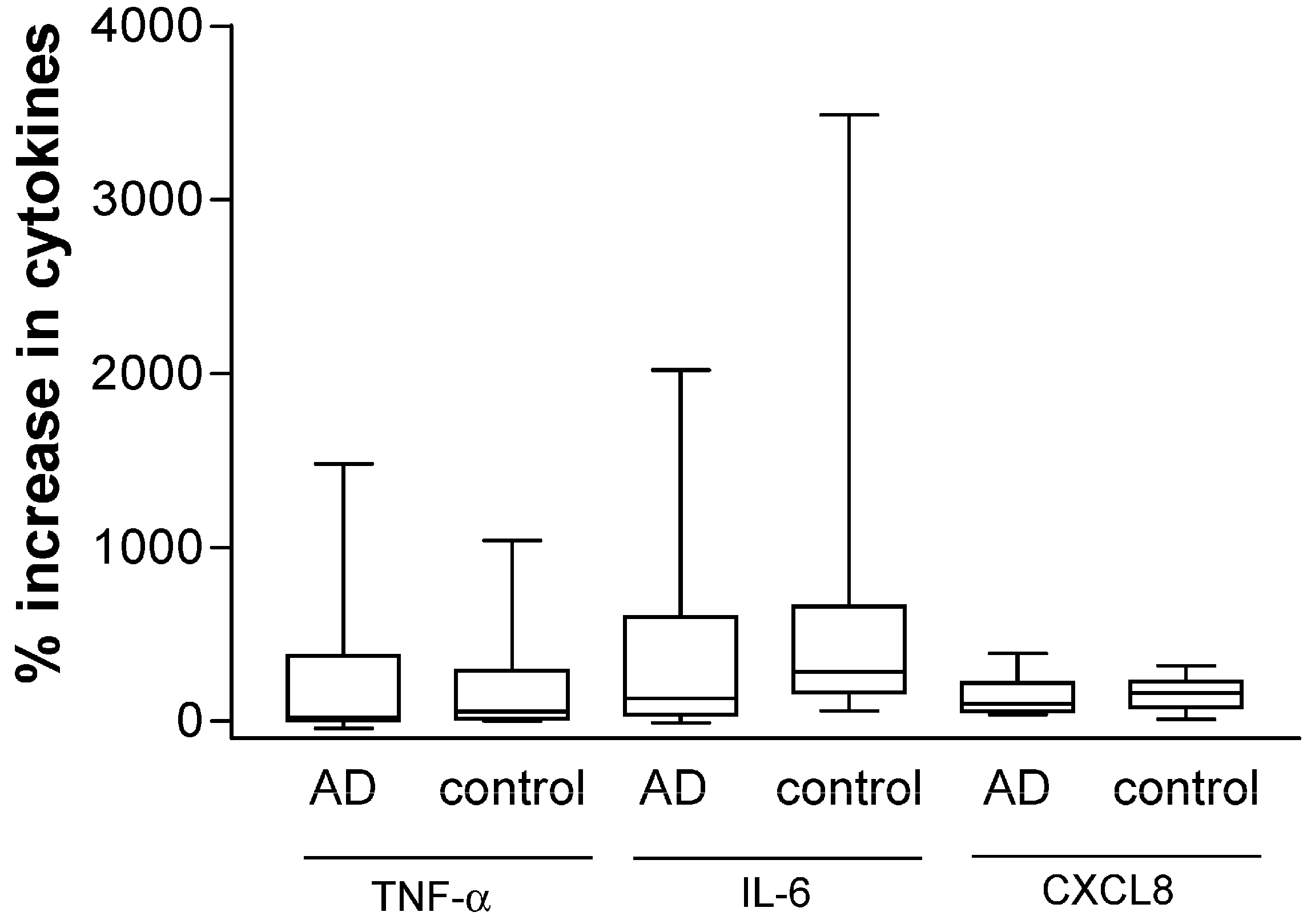
2.2. Plasma Concentrations of AD-Related Chemokines and Bacterial Infection-Related Dermcidin and HNP
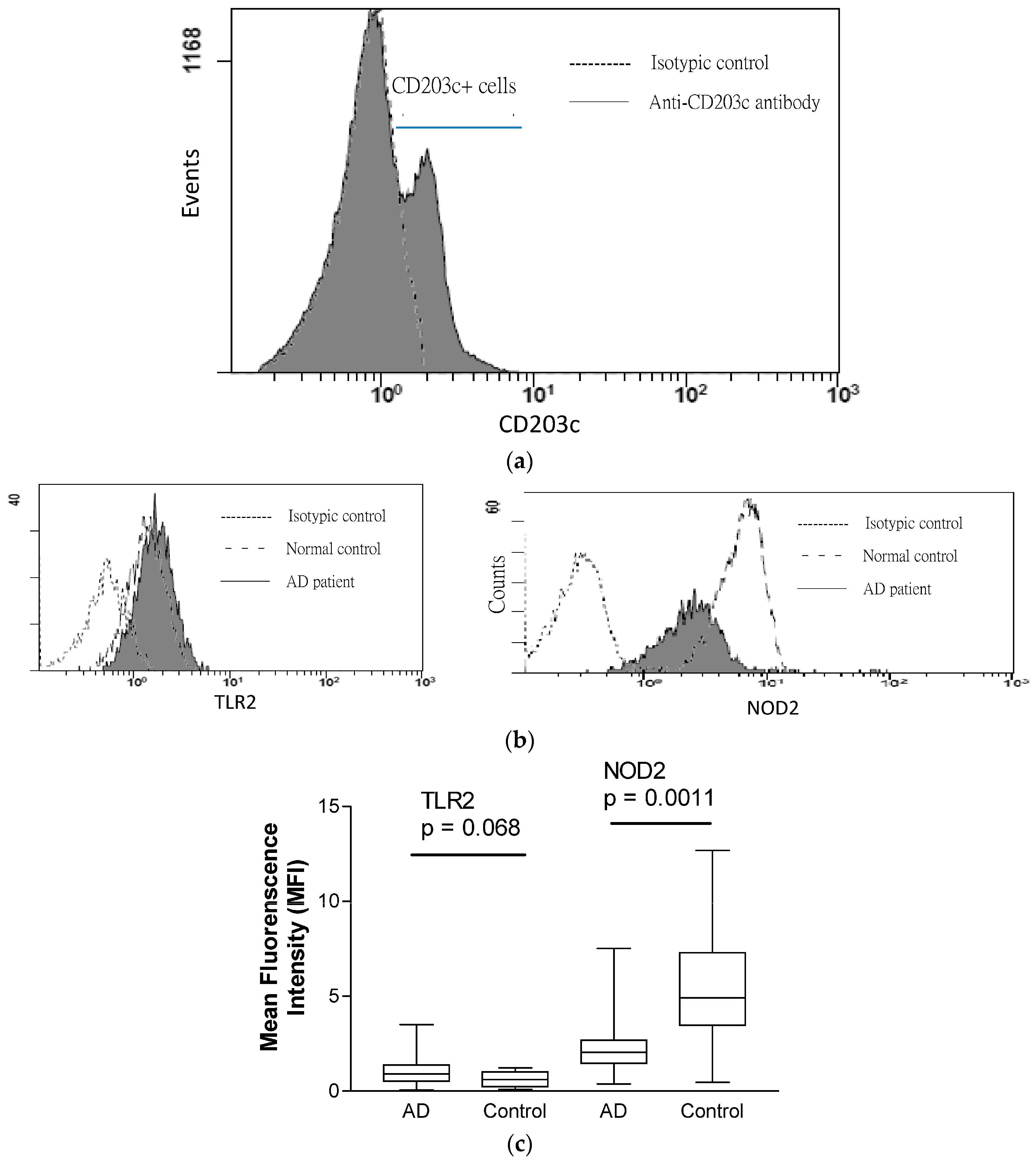
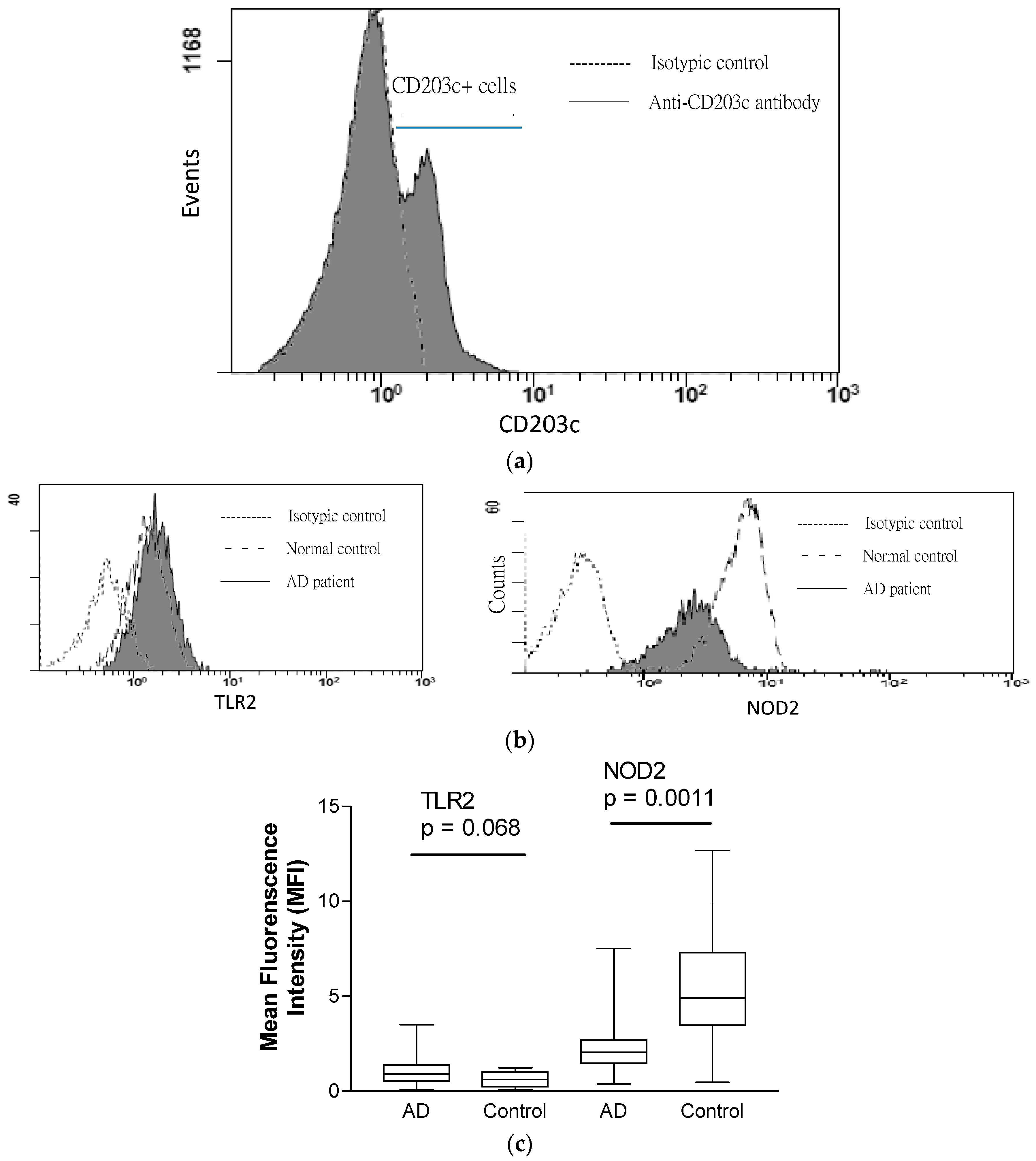
2.3. Expression of TLR2 and NOD2 in CD203+ Granulocytes
3. Materials and Methods
3.1. AD Patients, Non-Allergic Control Subjects and Blood Samples
3.2. Flow Cytometric Analysis for TLR2 and NOD2 Expression in CD203+ Basophils from AD Patients and Controls
3.3. Quantitative Analysis of Plasma Concentrations of Chemokines, Dermcidin and HNP
3.4. Assay of Serum IgE
3.5. Basophil Activation Test (BAT)
3.6. Ex Vivo Induction of Inflammatory Cytokines from PBMC
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Sokol, C.L.; Barton, G.M.; Farr, A.G.; Medzhitov, R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat. Immunol. 2008, 9, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Matsuoka, K.; Taya, C.; Suzuki, H.; Yokozeki, H.; Nishioka, K.; Hirokawa, K.; Etori, M.; Yamashita, M.; Kubota, T.; et al. Basophils play a critical role in the development of IgE-mediated chronic allergic inflammation independently of T cells and mast cells. Immunity 2005, 23, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Mukai, K.; Tsujimura, Y.; Ishiwata, K.; Kawano, Y.; Minegishi, Y.; Watanabe, N.; Karasuyama, H. Basophils are essential initiators of a novel type of chronic allergic inflammation. Blood 2007, 110, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Satoh, T.; Takayama, K.; Miyagishi, C.; Walls, A.F.; Yokozeki, H. Basophil recruitment and activation in inflammatory skin diseases. Allergy 2011, 66, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Ohnmacht, C.; Schwartz, C.; Panzer, M.; Schiedewitz, I.; Naumann, R.; Voehringer, D. Basophils orchestrate chronic allergic dermatitis and protective immunity against helminths. Immunity 2010, 33, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.W.; Kim, M.R.; Lee, E.Y.; Kim, J.H.; Kim, Y.S.; Jeon, S.G.; Yang, J.M.; Lee, B.J.; Pyun, B.Y.; Gho, Y.S.; et al. Extracellular vesicles derived from Staphylococcus aureus induce atopic dermatitis-like skin inflammation. Allergy 2011, 66, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Strandberg, K.; Lin, Y.C.; Peterson, M.L.; Leung, D.Y. Secreted virulence factor comparison between methicillin-resistant and methicillin-sensitive Staphylococcus aureus, and its relevance to atopic dermatitis. J. Allergy Clin. Immunol. 2010, 125, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Cheung, P.F.; Ip, W.K.; Lam, C.W. Intracellular signaling mechanisms regulating toll-like receptor-mediated activation of eosinophils. Am. J. Respir. Cell Mol. Biol. 2007, 37, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.F.; Wong, C.K.; Ip, W.K.; Lam, C.W. FAK-mediated activation of ERK for eosinophil migration: A novel mechanism for infection-induced allergic inflammation. Int. Immunol. 2008, 20, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Jiao, D.; Wong, C.K.; Qiu, H.N.; Dong, J.; Cai, Z.; Chu, M.; Hon, K.L.; Tsang, M.S.; Lam, C.W. NOD2 and TLR2 ligands trigger the activation of basophils and eosinophils by interacting with dermal fibroblasts in atopic dermatitis-like skin inflammation. Cell. Mol. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Mempel, M.; Voelcker, V.; Köllisch, G.; Plank, C.; Rad, R.; Gerhard, M.; Schnopp, C.; Fraunberger, P.; Walli, A.K.; Ring, J.; et al. Toll-like receptor expression in human keratinocytes: Nuclear factor kappaB controlled gene activation by Staphylococcus aureus is toll-like receptor 2 but not toll-like receptor 4 or platelet activating factor receptor dependent. J. Investig. Dermatol. 2003, 121, 1389–1396. [Google Scholar] [CrossRef] [PubMed]
- Niebuhr, M.; Baumert, K.; Werfel, T. TLR-2-mediated cytokine and chemokine secretion in human keratinocytes. Exp. Dermatol. 2010, 19, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.K.; Wen, H.; Ting, J.P. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu. Rev. Immunol. 2011, 29, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Hruz, P.; Zinkernagel, A.S.; Jenikova, G.; Botwin, G.J.; Hugot, J.P.; Karin, M.; Nizet, V.; Eckmann, L. NOD2 contributes to cutaneous defense against Staphylococcus aureus through alpha-toxin-dependent innate immune activation. Proc. Natl. Acad. Sci. USA 2009, 106, 12873–12878. [Google Scholar] [CrossRef] [PubMed]
- Doss, M.; White, M.R.; Tecle, T.; Hartshorn, K.L. Human defensins and LL-37 in mucosal immunity. J. Leukoc. Biol. 2010, 87, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Morel, E.; Alvarez, L.; Cabañas, R.; Fiandor, A.; Díaz, R.; Escamochero, S.; Prior, N.; Blanca, M.; Bellón, T. Expression of α-defensin 1–3 in T cells from severe cutaneous drug-induced hypersensitivity reactions. Allergy 2011, 66, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.R.; Gallo, R.L. Antimicrobial peptides, skin infections, and atopic dermatitis. Semin. Cutan. Med. Surg. 2008, 27, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Rieg, S.; Garbe, C.; Sauer, B.; Kalbacher, H.; Schittek, B. Dermcidin is constitutively produced by eccrine sweat glands and is not induced in epidermal cells under inflammatory skin conditions. Br. J. Dermatol. 2004, 151, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.P.; Peng, Y.F.; Zuo, Y.; Li, J.; Huang, J.; Wang, L.F.; Wu, Z.R. Functional and structural characterization of recombinant dermcidin-1L, a human antimicrobial peptide. Biochem. Biophys. Res. Commun. 2005, 328, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Nomura, I.; Goleva, E.; Howell, M.D.; Hamid, Q.A.; Ong, P.Y.; Hall, C.F.; Darst, M.A.; Gao, B.; Boguniewicz, M.; Travers, J.B.; et al. Cytokine milieu of atopic dermatitis, as compared to psoriasis, skin prevents induction of innate immune response genes. J. Immunol. 2003, 171, 3262–3269. [Google Scholar] [CrossRef] [PubMed]
- Rieg, S.; Steffen, H.; Seeber, S.; Humeny, A.; Kalbacher, H.; Dietz, K.; Garbe, C.; Schittek, B. Deficiency of dermcidin-derived antimicrobial peptides in sweat of patients with atopic dermatitis correlates with an impaired innate defense of human skin in vivo. J. Immunol. 2005, 174, 8003–8010. [Google Scholar] [CrossRef] [PubMed]
- Thijs, J.; Krastev, T.; Weidinger, S.; Buckens, C.F.; de Bruin-Weller, M.; Bruijnzeel-Koomen, C.; Flohr, C.; Hijnen, D. Biomarkers for atopic dermatitis: A systematic review and meta-analysis. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Machura, E.; Rusek-Zychma, M.; Jachimowicz, M.; Wrzask, M.; Mazur, B.; Kasperska-Zajac, A. Serum TARC and CTACK concentrations in children with atopic dermatitis, allergic asthma, and urticaria. Pediatr. Allergy Immunol. 2012, 23, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Homey, B.; Alenius, H.; Müller, A.; Soto, H.; Bowman, E.P.; Yuan, W.; McEvoy, L.; Lauerma, A.I.; Assmann, T.; Bünemann, E.; et al. CCL27-CCR10 interactions regulate T cell-mediated skin inflammation. Nat. Med. 2002, 8, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Vulcano, M.; Albanesi, C.; Stoppacciaro, A.; Bagnati, R.; D’Amico, G.; Struyf, S.; Transidico, P.; Bonecchi, R.; del Prete, A.; Allavena, P.; et al. Dendritic cells as a major source of macrophage-derived chemokine/CCL22 in vitro and in vivo. Eur. J. Immunol. 2001, 31, 812–822. [Google Scholar] [CrossRef]
- Imai, T.; Baba, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Yoshie, O. The T cell-directed CC chemokine TARC is a highly specific biological ligand for CC chemokine receptor 4. J. Biol. Chem. 1997, 272, 15036–15042. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Nagira, M.; Takagi, S.; Kakizaki, M.; Nishimura, M.; Wang, J.; Gray, P.W.; Matsushima, K.; Yoshie, O. Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage derived chemokine. Int. Immunol. 1999, 11, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Bianchi, G.; Bordignon, P.P.; D’Ambrosio, D.; Lang, R.; Borsatti, A.; Sozzani, S.; Allavena, P.; Gray, P.A.; Mantovani, A.; et al. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J. Exp. Med. 1998, 187, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Chantry, D.; Raport, C.J.; Wood, C.L.; Nishimura, M.; Godiska, R.; Yoshie, O.; Gray, P.W. Macrophage derived chemokine is a functional ligand for the CC chemokine receptor 4. J. Biol. Chem. 1998, 273, 1764–1768. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, T.; Nakamura, K.; Wakugawa, M.; Mitsui, H.; Tada, Y.; Saeki, H.; Torii, H.; Asahina, A.; Onai, N.; Matsushima, K.; et al. Thymus and activation regulated chemokine in atopic dermatitis: Serum thymus and activation regulated chemokine is closely related with disease activity. J. Allergy Clin. Immunol. 2001, 107, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, T.; Nakamura, K.; Wakugawa, M.; Mitsui, H.; Tada, Y.; Saeki, H.; Torii, H.; Komine, M.; Asahina, A.; Tamaki, K. Serum macrophage-derived chemokine (MDC) levels is closely related with the disease activity of atopic dermatitis. Clin. Exp. Immunol. 2002, 127, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Nakamura, K.; Oyama, N.; Kaneko, F.; Tsunemi, Y.; Saeki, H.; Tamaki, K. Macrophage-derived chemokine (MDC)/CCL22 produced by monocyte derived dendritic cells reflects the disease activity in patients with atopic dermatitis. J. Dermatol. Sci. 2006, 44, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Homey, B.; Steinhoff, M.; Ruzicka, T.; Leung, D.Y. Cytokines and chemokines orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 118, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Shiohara, T.; Doi, T.; Hayakawa, J. Defective sweating responses in atopic dermatitis. Curr. Probl. Dermatol. 2011, 41, 68–79. [Google Scholar] [PubMed]
- Wang, E.; Qiang, X.; Li, J.; Zhu, S.; Wang, P. The in Vitro Immune-Modulating Properties of a Sweat Gland-Derived Antimicrobial Peptide Dermcidin. Shock 2016, 45, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Song, P.I.; Seo, C.H.; Cheong, H.; Park, Y. Colonization and infection of the skin by S. aureus: Immune system evasion and the response to cationic antimicrobial peptides. Int. J. Mol. Sci. 2014, 15, 8753–8772. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Leung, T.F.; Chu, I.M.; Dong, J.; Lam, Y.Y.; Lam, C.W. Aberrant Expression of Regulatory Cytokine IL-35 and Pattern Recognition Receptor NOD2 in Patients with Allergic Asthma. Inflammation 2015, 38, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Hanifin, J.M.; Rajka, G. Diagnostic features of atopic dermatitis. Acta Derm. Venereol. 1980, 2, 44–47. [Google Scholar]
- Severity scoring of atopic dermatitis: The SCORAD index. Consensus Report of the European Task Force on Atopic Dermatitis. Dermatology 1993, 186, 23–31. [Google Scholar]
- Hon, K.L.; Wong, K.Y.; Leung, T.F.; Chow, C.M.; Ng, P.C. Comparison of skin hydration evaluation sites and correlations among skin hydration, transepidermal water loss, SCORAD Index, Nottingham Eczema Severity Score, and quality of life in patients with atopic dermatitis. Am. J. Clin. Dermatol. 2008, 9, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wehner, J.; Neuber, K. Staphylococcus aureus enterotoxins induce histamine and leukotriene release in patients with atopic eczema. Br. J. Dermatol. 2001, 145, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Ho, C.Y.; Ko, F.W.; Chan, C.H.; Ho, A.S.; Hui, D.S.; Lam, C.W. Proinflammatory cytokines (IL-17, IL-6, IL-18 and IL-12) and Th cytokines (IFN-gamma, IL-4, IL-10 and IL-13) in patients with allergic asthma. Clin. Exp. Immunol. 2001, 125, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.F.; Sy, H.Y.; Wong, G.W.; Ng, M.C.; Chan, I.H.; Li, C.Y.; Tang, N.L.; Waye, M.W.; Lam, C.W. Asthma and atopy are associated with chromosome 17q21 markers in Chinese children. Allergy 2009, 64, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Hu, S.; Leung, K.M.; Dong, J.; He, L.; Chu, Y.J.; Chu, I.M.; Qiu, H.N.; Liu, K.Y.; Lam, C.W. NOD-like receptors mediated activation of eosinophils interacting with bronchial epithelial cells: A link between innate immunity and allergic asthma. Cell. Mol. Immunol. 2013, 10, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.F.; Wong, C.K.; Chan, I.H.; Ip, W.K.; Lam, C.W.; Wong, G.W. Plasma concentration of thymus and activation-regulated chemokine is elevated in childhood asthma. J. Allergy Clin. Immunol. 2002, 110, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S. Basophil activation test in allergy: Time for an update? Int. Arch. Allergy Immunol. 2012, 158, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Lit, L.C.; Tam, L.S.; Li, E.K.; Wong, P.T.; Lam, C.W. Hyperproduction of IL-23 and IL-17 in patients with systemic lupus erythematosus: Implications for Th17-mediated inflammation in auto-immunity. Clin. Immunol. 2008, 127, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compound muramyl dipeptide are available from the authors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | AD Patients | Control Subjects |
|---|---|---|
| (n = 27) | (n = 15) | |
| Sex (male/female) | 15/12 | 5/10 |
| Age at evaluation, year (mean ± SD, range) | 14.3 ± 2.4 (9–18) | 21.2 ± 6.1 (10–29) |
| SCORAD index | 48.0 (40.5–61.7) | N.A. |
| Objective SCORAD index | 38.2 (29.9–47.0) | N.A. |
| Intensity (Erythema, Edema/Papulation, Oozing/Crusting, Excoriation, Lichenification Dryness) | 8.0 (6.5–10.0) | N.A. |
| Pruritus | 7.0 (4.5–8.0) | N.A. |
| Sleep loss | 5.5 (0.0–7.5) | N.A. |
| SH | 24.6 (15.4–34.7) | N.A. |
| TWEL | 8.5 (7.8–10.8) | N.A. |
| Area affected (%) | 44 (35.8–64.0) | N.A. |
| Lichenification | 1.0 (1.0–2.0) | N.A. |
| CDLQI | 10.0 (4.5–15.5) | N.A. |
| Presence of at least one positive allergen-specific IgE, n (%) | 27 (100) | 0 (0) |
| D. pteronyssinus, n (%) | 27 (100) | 0 (0) |
| Cockroaches, n (%) | 16 (59) | 0 (0) |
| Cat dander, n (%) | 13 (48) | 0 (0) |
| Dog dander, n (%) | 21 (78) | 0 (0) |
| Mixed molds, n (%) | 18 (67) | 0 (0) |
| Staphylococcal enterotoxin A, n (%) | 16 (59) | 0 (0) |
| Staphylococcal enterotoxin B, n (%) | 14 (52) | 0 (0) |
| Staphylococcal enterotoxin C, n (%) | 12 (44) | 0 (0) |
| Positive basophil activation test | ||
| House dust mite, D. pteronyssinus, n (%) | 26 (96) | 0 (0) |
| Cockroaches, Bla g 2 protein, n (%) | 21 (78) | 0 (0) |
| Cat dander, Fel d 1 protein, n (%) | 19 (70) | 0 (0) |
| Positive S. aureus growth (wound swabs), n (%) | 18 (70) | N.A. |
| Treatment | ||
| Topical corticosteroid | 24 (89) | N.A. |
| Emollient | 27 (100) | N.A. |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, C.-K.; Chu, I.M.-T.; Hon, K.-L.; Tsang, M.S.-M.; Lam, C.W.-K. Aberrant Expression of Bacterial Pattern Recognition Receptor NOD2 of Basophils and Microbicidal Peptides in Atopic Dermatitis. Molecules 2016, 21, 471. https://doi.org/10.3390/molecules21040471
Wong C-K, Chu IM-T, Hon K-L, Tsang MS-M, Lam CW-K. Aberrant Expression of Bacterial Pattern Recognition Receptor NOD2 of Basophils and Microbicidal Peptides in Atopic Dermatitis. Molecules. 2016; 21(4):471. https://doi.org/10.3390/molecules21040471
Chicago/Turabian StyleWong, Chun-Kwok, Ida Miu-Ting Chu, Kam-Lun Hon, Miranda Sin-Man Tsang, and Christopher Wai-Kei Lam. 2016. "Aberrant Expression of Bacterial Pattern Recognition Receptor NOD2 of Basophils and Microbicidal Peptides in Atopic Dermatitis" Molecules 21, no. 4: 471. https://doi.org/10.3390/molecules21040471
APA StyleWong, C.-K., Chu, I. M.-T., Hon, K.-L., Tsang, M. S.-M., & Lam, C. W.-K. (2016). Aberrant Expression of Bacterial Pattern Recognition Receptor NOD2 of Basophils and Microbicidal Peptides in Atopic Dermatitis. Molecules, 21(4), 471. https://doi.org/10.3390/molecules21040471