
Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats

Abstract
:
1. Introduction
2. Results
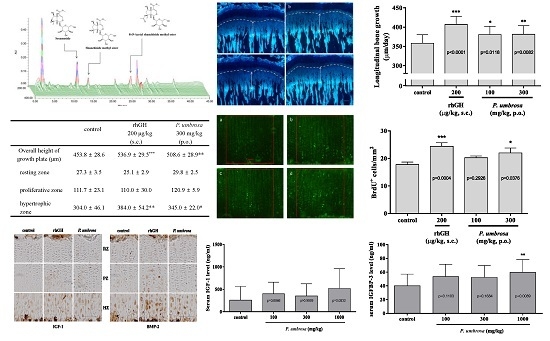
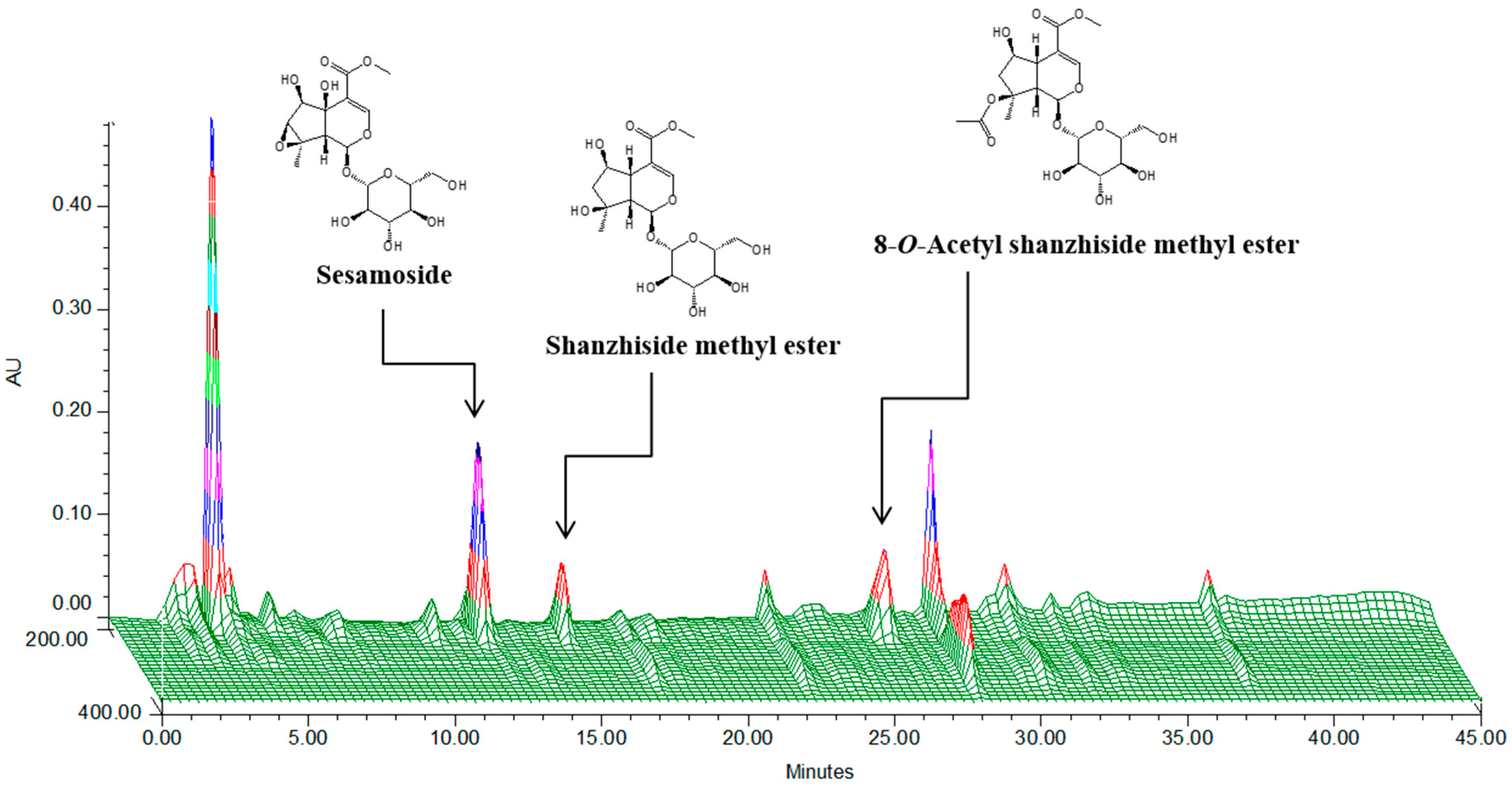
2.1. HPLC Analysis of P. umbrosa Extract
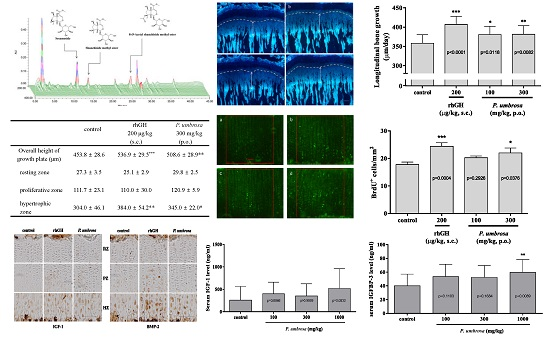
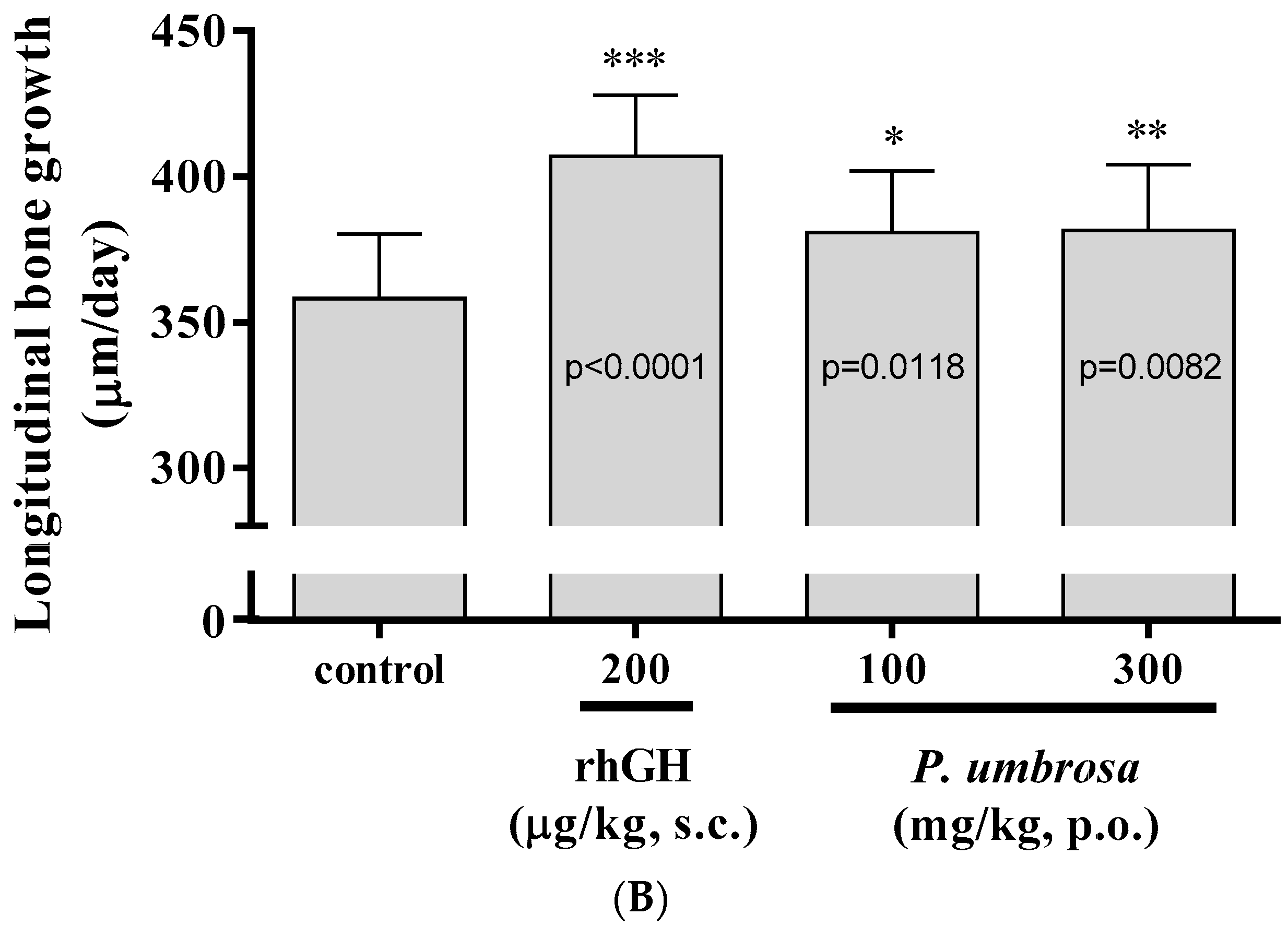
2.2. Effect on Longitudinal Bone Growth Rate
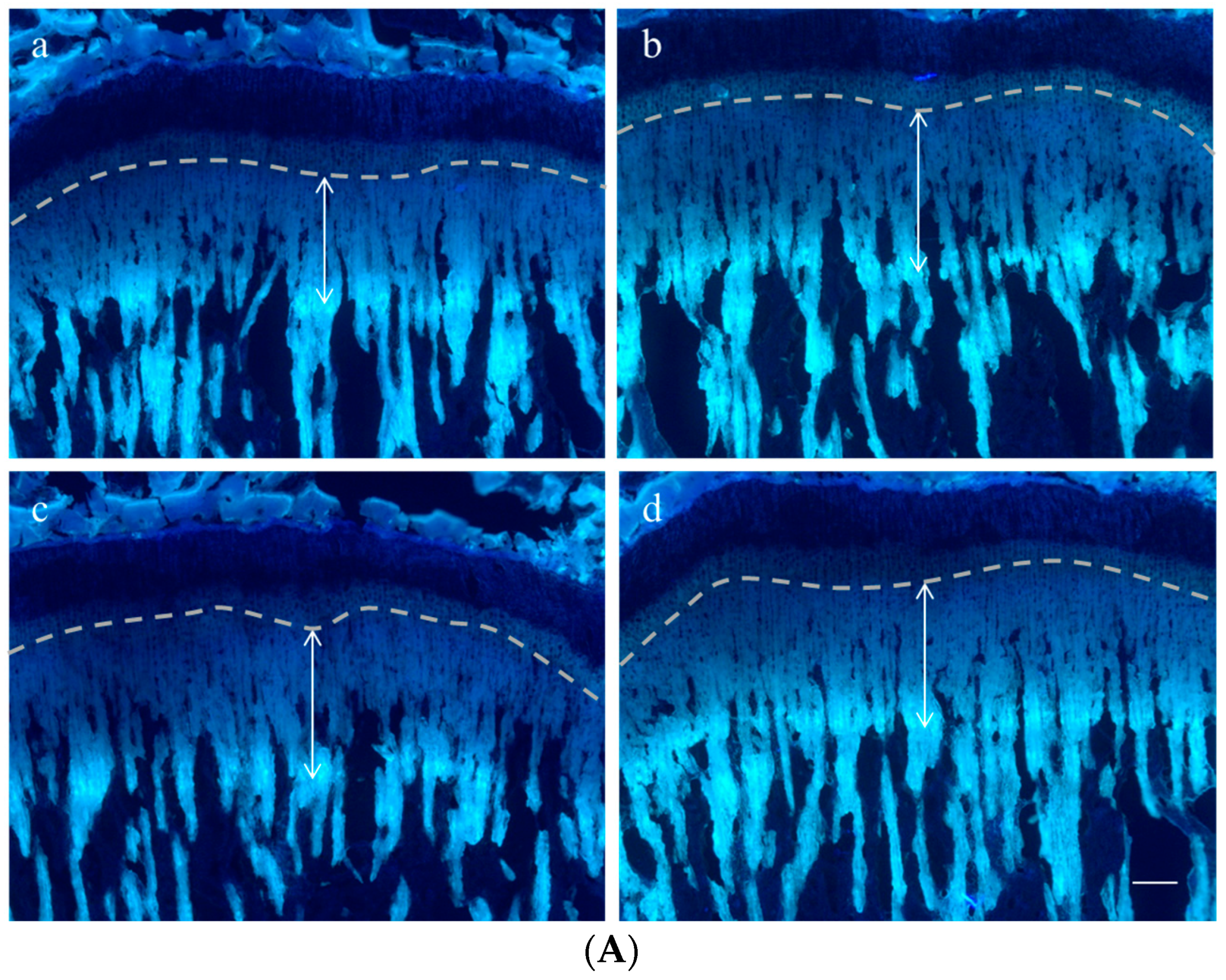
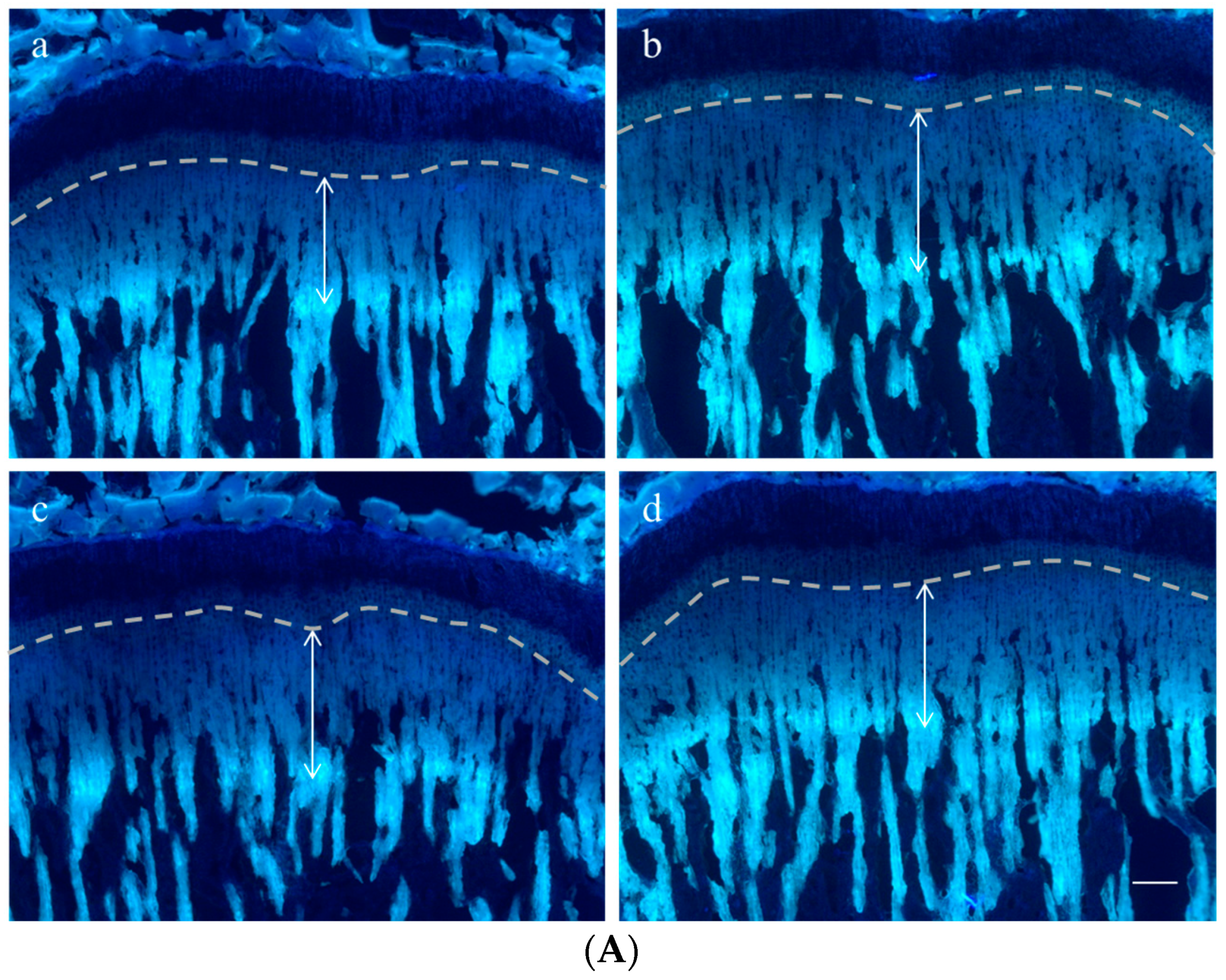
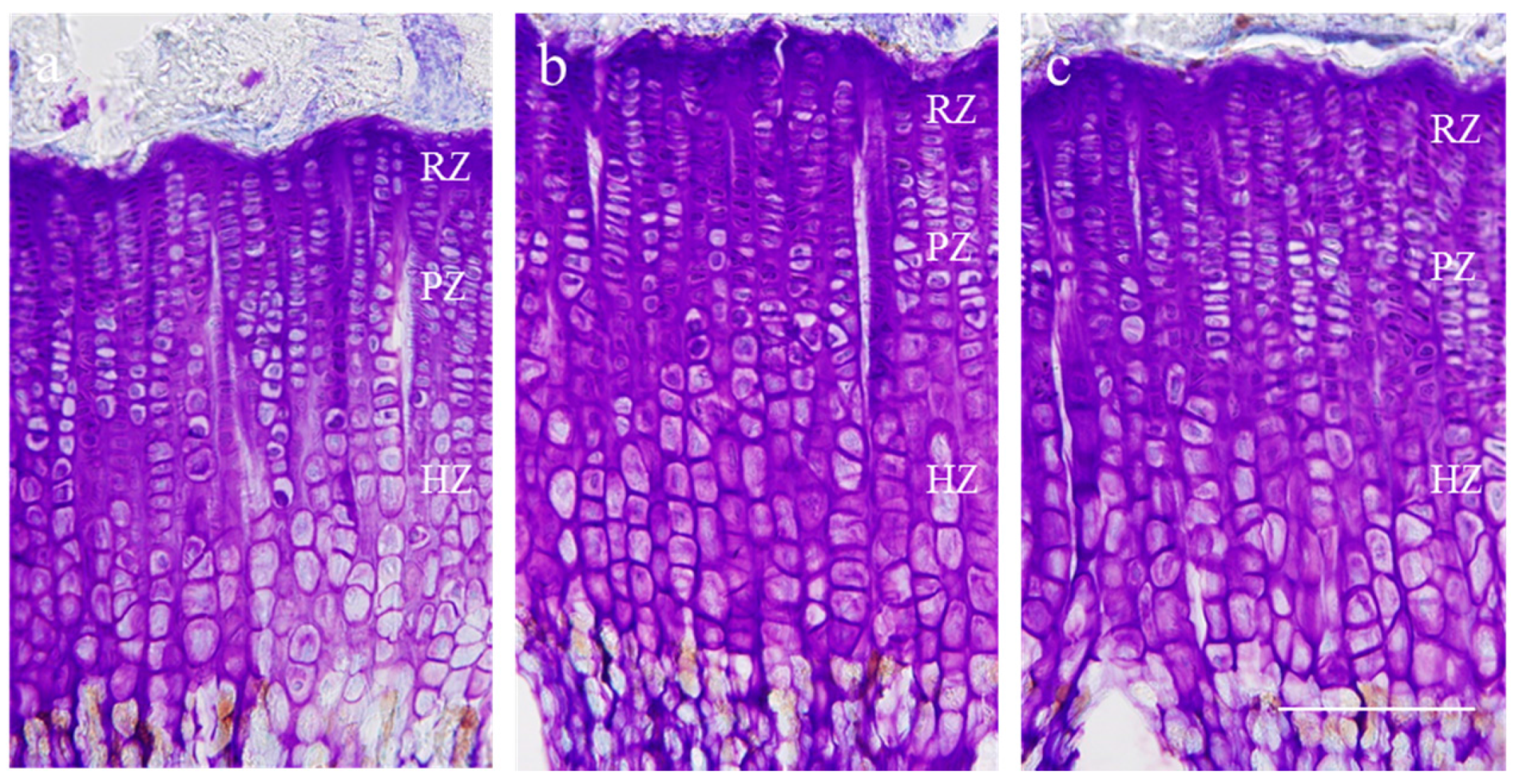
2.3. Effect on Growth Plate Height
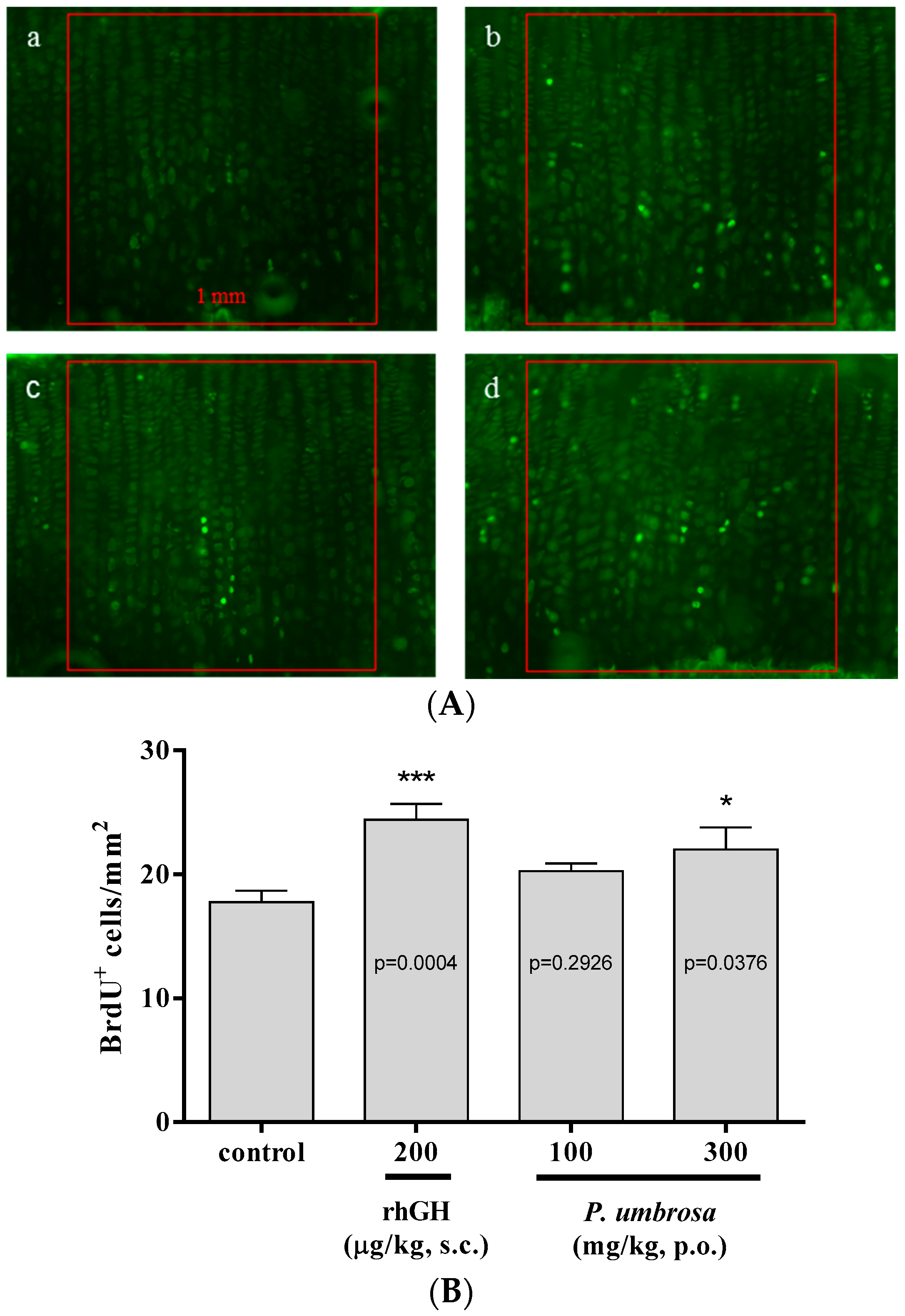
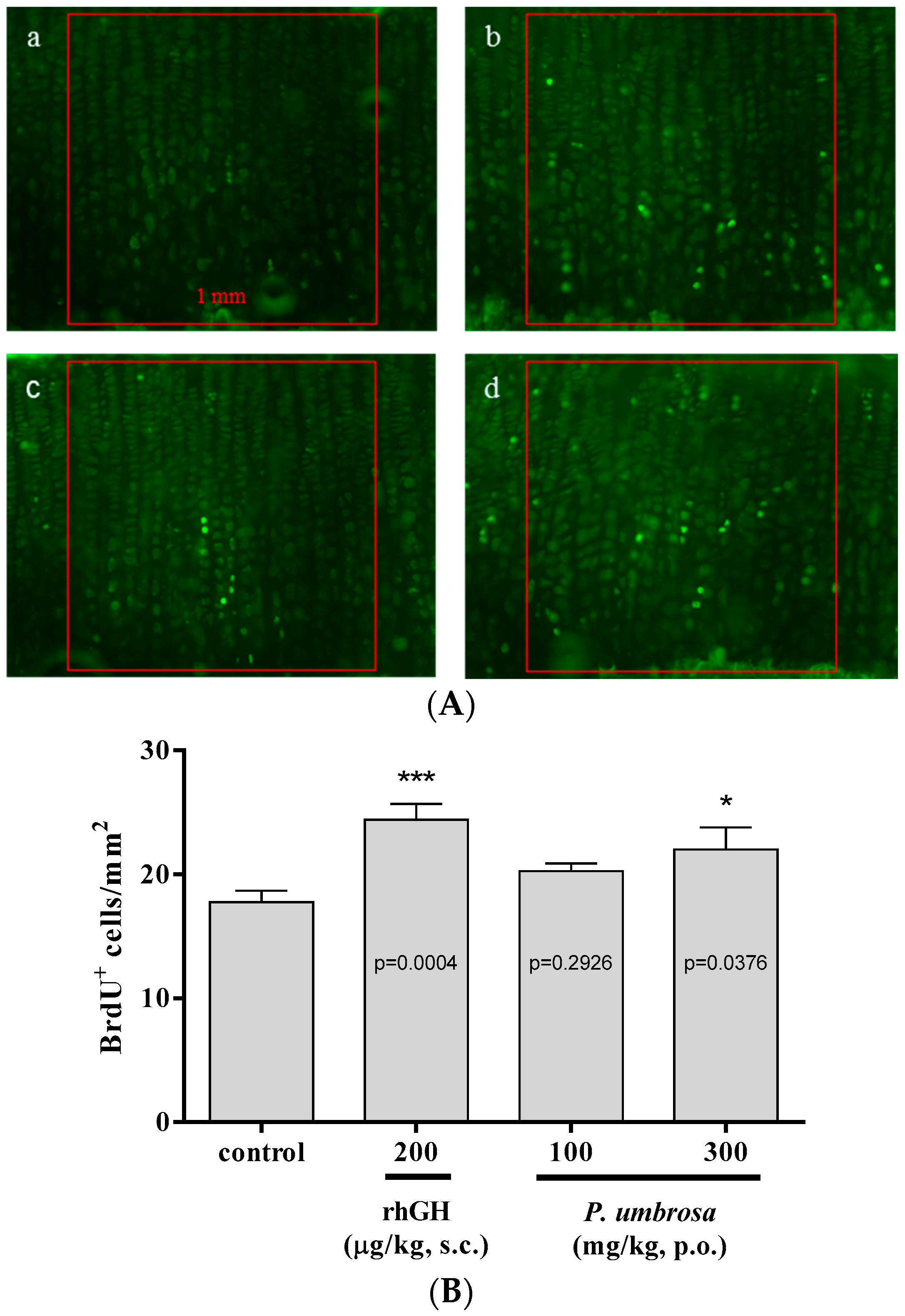
2.4. Effect on Chondrocyte Proliferation
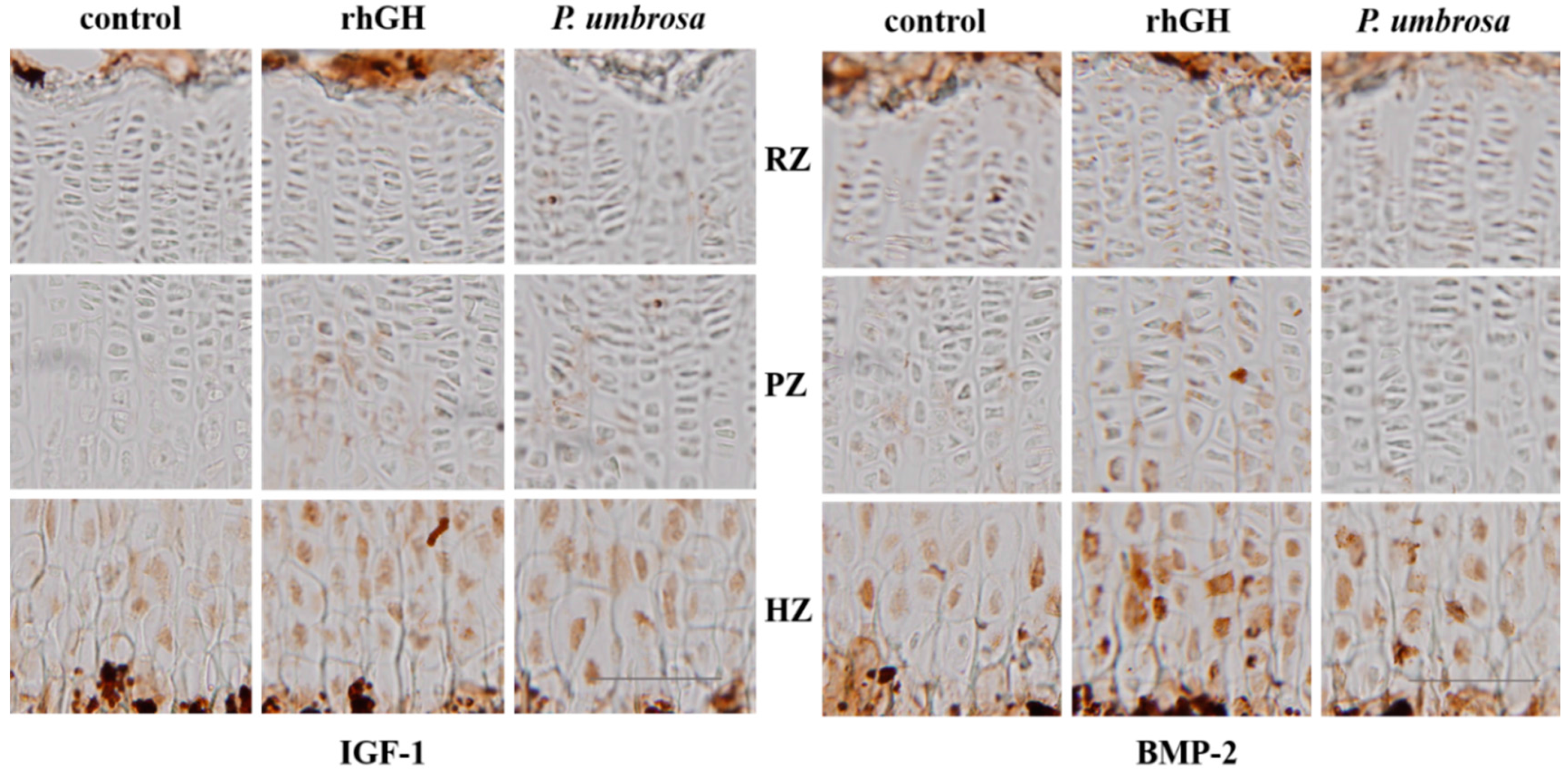
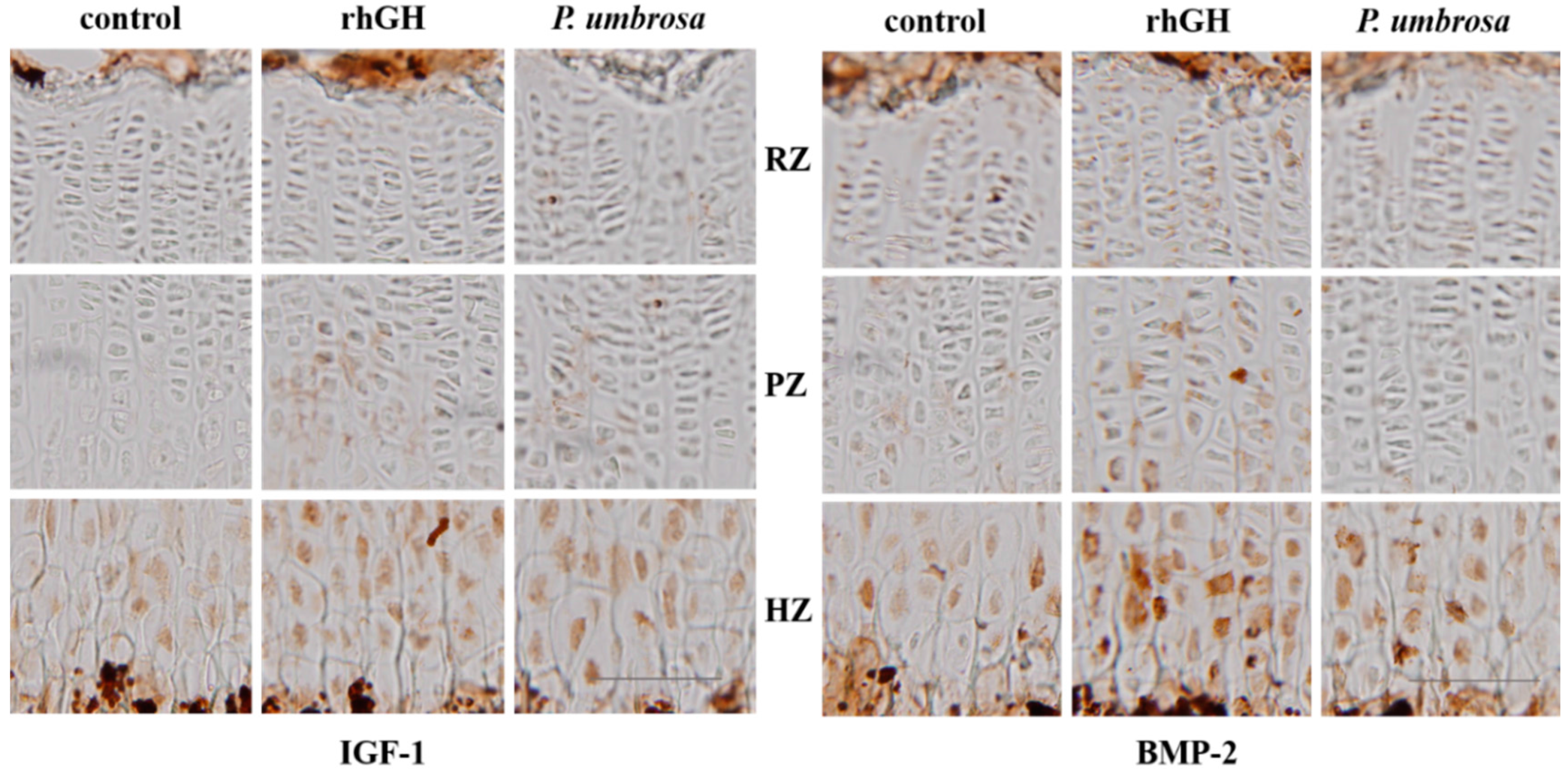
2.5. Effects on IGF-1 and BMP-2 Expression
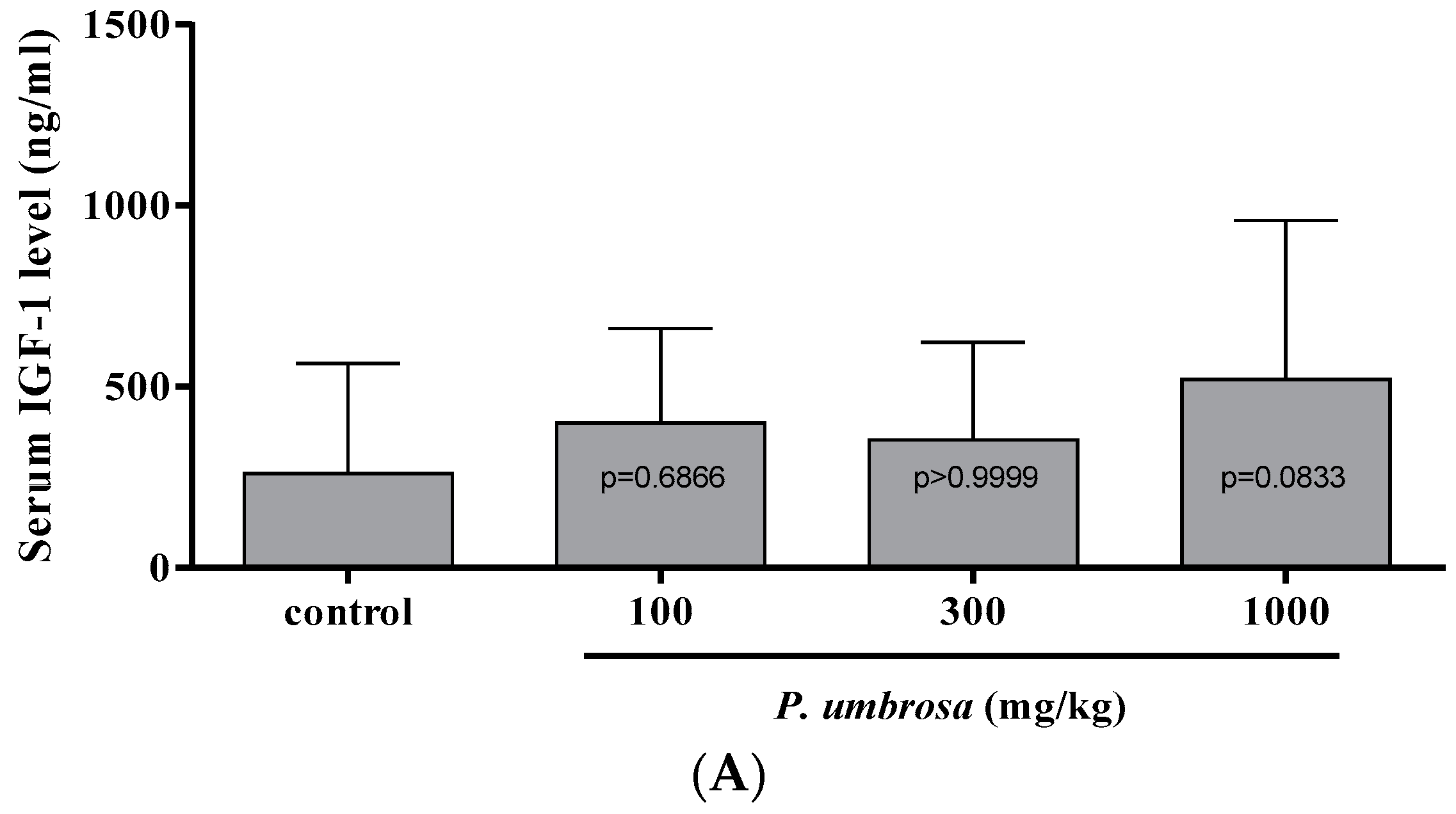
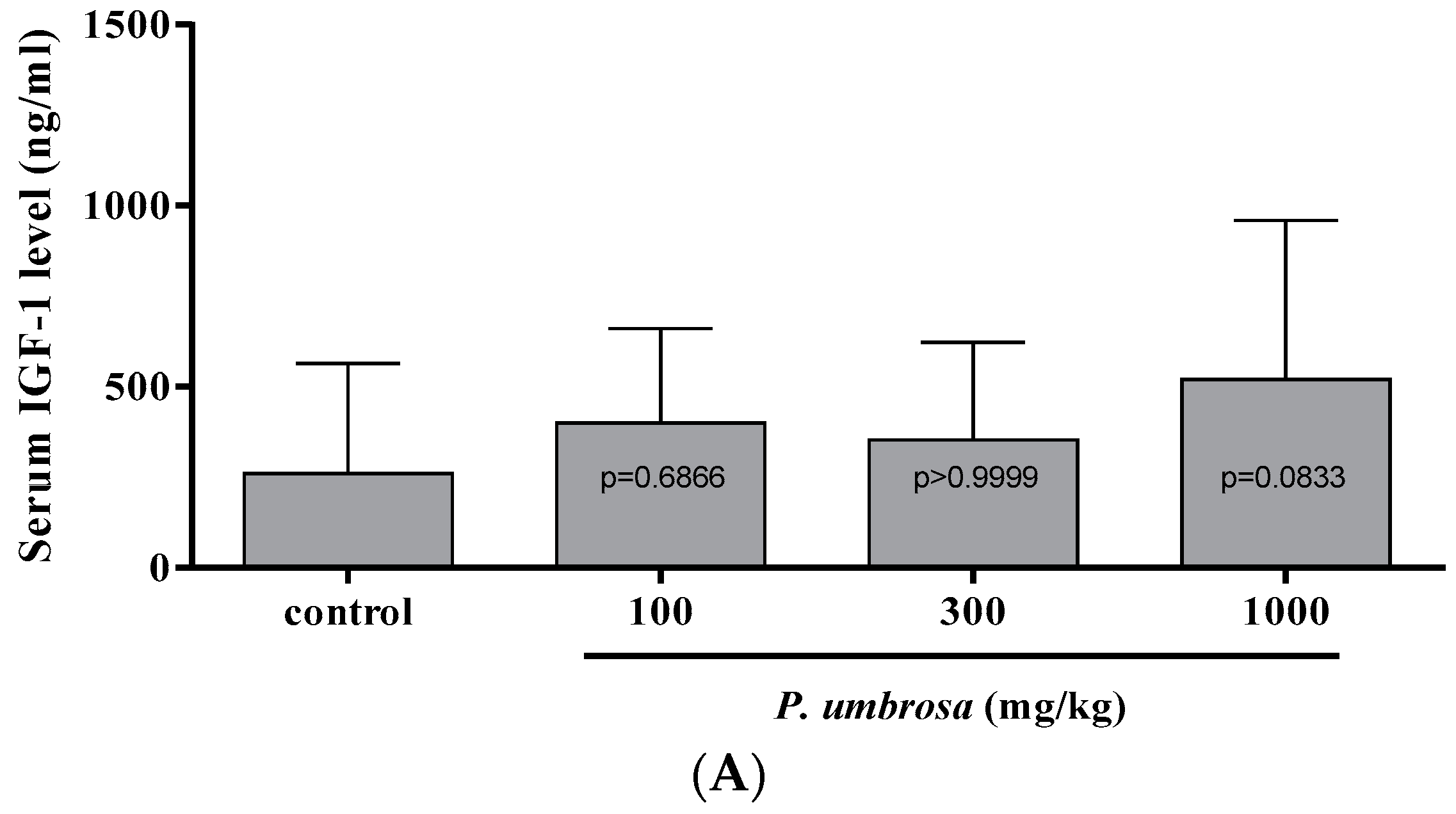
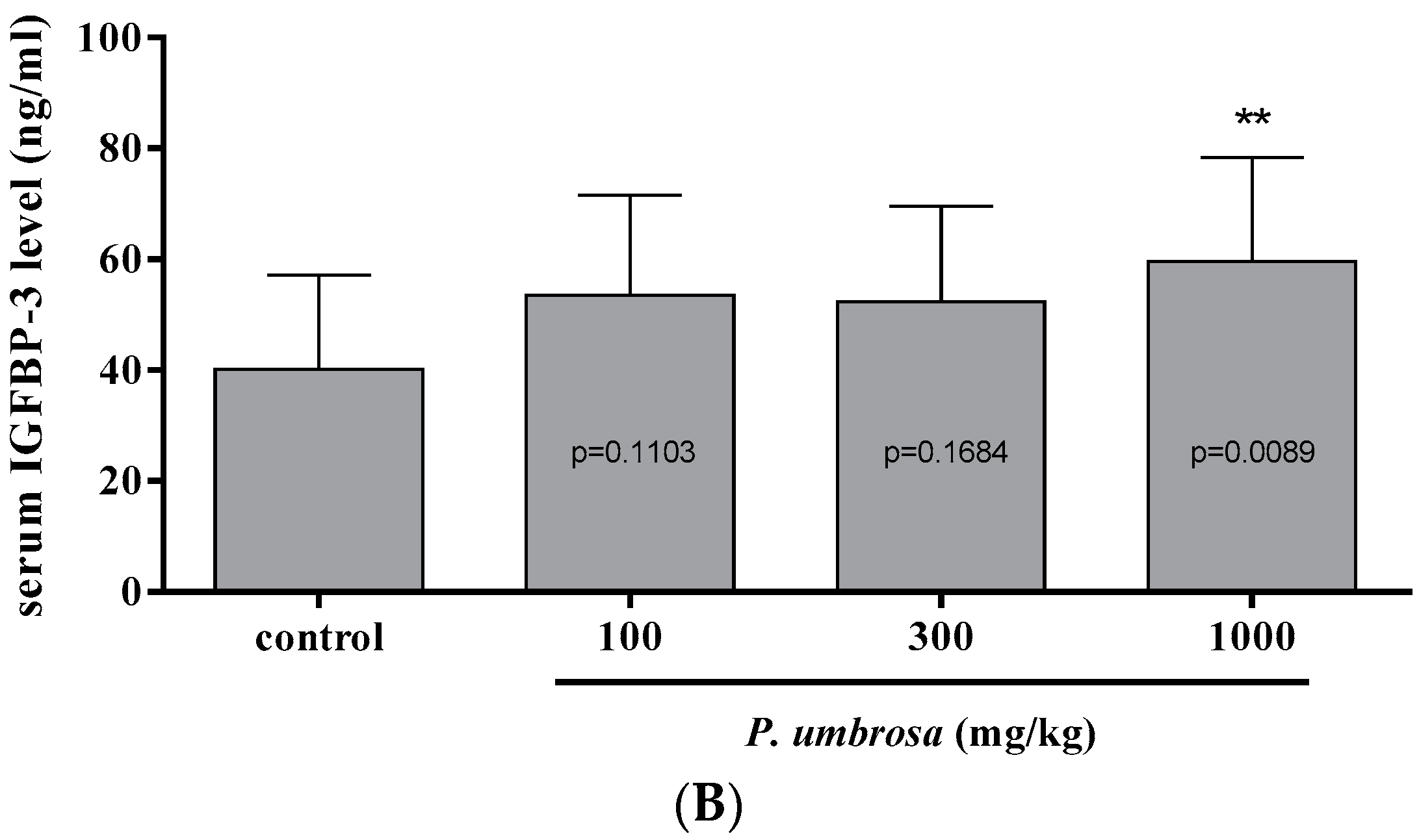
2.6. Effect on Serum IGF-1 and IGFBP-3 Concentrations
3. Discussion
4. Experimental Section
4.1. Plant Material
4.2. Sample Preparation and HPLC Analysis
4.3. Animals
4.4. Treatment
4.5. Longitudinal Bone Growth Rate
4.6. Growth Plate Height
4.7. Chondrocyte Proliferation
4.8. Immunohistochemistry
4.9. Serum IGF-1 and IGFBP-3
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cohen, P.; Rogol, A.D.; Deal, C.L.; Saenger, P.; Reiter, E.O.; Ross, J.L.; Chernausek, S.D.; Savage, M.O.; Wit, J.M. Consensus statement on the diagnosis and treatment of children with idiopathic short stature: A summary of the growth hormone research society, the lawson wilkins pediatric endocrine society, and the european society for paediatric endocrinology workshop. J. Clin. Endocrinol. Metab. 2008, 93, 4210–4217. [Google Scholar] [PubMed]
- Downie, A.B.; Mulligan, J.; Stratford, R.J.; Betts, P.R.; Voss, L.D. Are short normal children at a disadvantage? The wessex growth study. BMJ 1997, 314, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Stabler, B.; Clopper, R.R.; Siegel, P.T.; Stoppani, C.; Compton, P.G.; Underwood, L.E. Academic achievement and psychological adjustment in short children. The national cooperative growth study. J. Dev. Behav. Pediatr. 1994, 15, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Voss, L.D.; Sandberg, D.E. The psychological burden of short stature: Evidence against. Eur. J. Endocrinol. 2004, 151 (Suppl. S1), S29–S33. [Google Scholar] [CrossRef] [PubMed]
- Zimet, G.D.; Owens, R.; Dahms, W.; Cutler, M.; Litvene, M.; Cuttler, L. Psychosocial outcome of children evaluated for short stature. JAMA Pediatr. 1997, 151, 1017–1023. [Google Scholar] [CrossRef]
- Christensen, T.L.; Djurhuus, C.B.; Clayton, P.; Christiansen, J.S. An evaluation of the relationship between adult height and health-related quality of life in the general UK population. Clin. Endocrinol. 2007, 67, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Paajanen, T.A.; Oksala, N.K.; Kuukasjarvi, P.; Karhunen, P.J. Short stature is associated with coronary heart disease: A systematic review of the literature and a meta-analysis. Eur. Heart J. 2010, 31, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Wannamethee, S.G.; Shaper, A.G.; Whincup, P.H.; Walker, M. Adult height, stroke, and coronary heart disease. Am. J. Epidemiol. 1998, 148, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Hindmarsh, P.C.; Dattani, M.T. Use of growth hormone in children. Nat. Clin. Pract. Endoc. 2006, 2, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Wit, J.M.; Kamp, G.A.; Rikken, B. Spontaneous growth and response to growth hormone treatment in children with growth hormone deficiency and idiopathic short stature. Pediatr. Res. 1996, 39, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Hintz, R.L.; Attie, K.M.; Baptista, J.; Roche, A. Effect of growth hormone treatment on adult height of children with idiopathic short stature. Genentech collaborative group. N. Engl. J. Med. 1999, 340, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Leschek, E.W.; Rose, S.R.; Yanovski, J.A.; Troendle, J.F.; Quigley, C.A.; Chipman, J.J.; Crowe, B.J.; Ross, J.L.; Cassorla, F.G.; Blum, W.F.; et al. Effect of growth hormone treatment on adult height in peripubertal children with idiopathic short stature: A randomized, double-blind, placebo-controlled trial. J. Clin. Endocrinol. Metab. 2004, 89, 3140–3148. [Google Scholar] [CrossRef] [PubMed]
- August, G.P.; Julius, J.R.; Blethen, S.L. Adult height in children with growth hormone deficiency who are treated with biosynthetic growth hormone: The national cooperative growth study experience. Pediatrics 1998, 102, 512–516. [Google Scholar] [PubMed]
- Bajpai, A.; Menon, P.S. Growth hormone therapy. Indian J. Pediatr. 2005, 72, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.; Baxter, L.; Cave, C.B.; Milne, R. Recombinant growth hormone for idiopathic short stature in children and adolescents. Cochrane DB Syst. Rev. 2007. [Google Scholar] [CrossRef]
- Finkelstein, B.S.; Imperiale, T.F.; Speroff, T.; Marrero, U.; Radcliffe, D.J.; Cuttler, L. Effect of growth hormone therapy on height in children with idiopathic short stature: A meta-analysis. JAMA Pediatr. 2002, 156, 230–240. [Google Scholar] [CrossRef]
- Hintz, R.L. Growth hormone: Uses and abuses. BMJ 2004, 328, 907–908. [Google Scholar] [CrossRef] [PubMed]
- Silverman, B.L.; Blethen, S.L.; Reiter, E.O.; Attie, K.M.; Neuwirth, R.B.; Ford, K.M. A long-acting human growth hormone (nutropin depot): Efficacy and safety following two years of treatment in children with growth hormone deficiency. J. Pediatr. Endocr. Metab. 2002, 15 (Suppl. S2), 715–722. [Google Scholar] [CrossRef]
- Hero, M.; Wickman, S.; Dunkel, L. Treatment with the aromatase inhibitor letrozole during adolescence increases near-final height in boys with constitutional delay of puberty. Clin. Endocrinol. 2006, 64, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Alba-Roth, J.; Muller, O.A.; Schopohl, J.; von Werder, K. Arginine stimulates growth hormone secretion by suppressing endogenous somatostatin secretion. J. Clin. Endocrinol. Metab. 1988, 67, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Stammers, A.L.; Lowe, N.M.; Medina, M.W.; Patel, S.; Dykes, F.; Perez-Rodrigo, C.; Serra-Majam, L.; Nissensohn, M.; Moran, V.H. The relationship between zinc intake and growth in children aged 1–8 years: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2015, 69, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.T.; Hamed, A.I.; Sallam, M.T. Effect of zinc supplementation on growth hormone-insulin growth factor axis in short egyptian children with zinc deficiency. Ital. J. Pediatr. 2012, 38, 21. [Google Scholar] [CrossRef] [PubMed]
- Leem, K.; Park, S.Y.; Lee, D.H.; Kim, H. Lovastatin increases longitudinal bone growth and bone morphogenetic protein-2 levels in the growth plate of sprague-dawley rats. Eur. J. Pediatr. 2002, 161, 406–407. [Google Scholar] [CrossRef] [PubMed]
- Wilsman, N.J.; Farnum, C.E.; Leiferman, E.M.; Fry, M.; Barreto, C. Differential growth by growth plates as a function of multiple parameters of chondrocytic kinetics. J. Orthop. Res. 1996, 14, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, F. Bone development and its relation to fracture repair. The role of mesenchymal osteoblasts and surface osteoblasts. Eur. Cells Mater. 2008, 15, 53–76. [Google Scholar]
- Shin, T.Y.; Lee, J.K. Effect of phlomis umbrosa root on mast cell-dependent immediate-type allergic reactions by anal therapy. Immunopharmacol Immunotoxicol. 2003, 25, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Wang, J.; Li, M.; Miao, X.; Pan, H.; Yang, Y.; Wang, Y. Antinociceptive and anti-inflammatory activities of phlomis umbrosa turcz extract. Fitoterapia 2011, 82, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.Y.; Kim, S.H.; Kim, D.K.; Leem, K.H.; Park, J.S. Phlomis umbrosa root inhibits mast cell-dependent allergic reactions and inflammatory cytokine secretion. Phytother. Res. 2008, 22, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Limem-Ben Amor, I.; Boubaker, J.; Ben Sgaier, M.; Skandrani, I.; Bhouri, W.; Neffati, A.; Kilani, S.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Phytochemistry and biological activities of phlomis species. J. Ethnopharmacol. 2009, 125, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Li, M.X.; Shang, X.F.; Jia, Z.P.; Zhang, R.X. Phytochemical and biological studies of plants from the genus phlomis. Chem. Biodivers. 2010, 7, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhang, S.; Fu, F.; Zhu, H.; Hou, J. Inhibition of nuclear factor-kappab by 6-O-acetyl shanzhiside methyl ester protects brain against injury in a rat model of ischemia and reperfusion. J. Neuroinflamm. 2010, 7. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.C.; Jiang, W.L.; Xu, Y.; Zhu, H.B.; Hou, J. Cardioprotection with 8-O-acetyl shanzhiside methylester on experimental myocardial ischemia injury. Eur. J. Pharm Sci. 2012, 47, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kan, Z.C.; Zhang, X.L.; Fang, H.; Jiang, W.L. 8-O-acetyl shanzhiside methylester attenuates cerebral ischaemia/reperfusion injury through an anti-inflammatory mechanism in diabetic rats. Basic Clin. Pharmacol. 2014, 115, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Clement-Lacroix, P.; Da Ponte, F.; Bouali, Y.; Binart, N.; Moriggl, R.; Goffin, V.; Coschigano, K.; Gaillard-Kelly, M.; Kopchick, J.; et al. Bone homeostasis in growth hormone receptor-null mice is restored by igf-i but independent of stat5. J. Clin. Investig. 2000, 106, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Rosen, C.J. From mouse to man: Redefining the role of insulin-like growth factor-I in the acquisition of bone mass. Exp. Biol. Med. 2003, 228, 245–252. [Google Scholar]
- Van der Eerden, B.C.; Karperien, M.; Wit, J.M. Systemic and local regulation of the growth plate. Endocr. Rev. 2003, 24, 782–801. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.I.; Stenstrom, A.; Thorngren, K.G. Skeletal deposition and toxicity of methacycline. Nature 1968, 219, 624–625. [Google Scholar] [CrossRef] [PubMed]
- Beauloye, V.; Muaku, S.M.; Lause, P.; Portetelle, D.; Renaville, R.; Robert, A.R.; Ketelslegers, J.M.; Maiter, D. Monoclonal antibodies to growth hormone (GH) prolong liver GH binding and GH-induced IGF-I/Igfbp-3 synthesis. Am. J. Physiol. 1999, 277, E308–E315. [Google Scholar] [PubMed]
- Han, S.H. Effect of the Active Compounds in Phlomidis Radix on Bone Regeneration in Primary Cultured Osteoblasts Derived from Alveolar Bone. Ph.D. Thesis, Wonkwang University, Jeollabuk-do, Korea, 2004. [Google Scholar]
- Kim, S.Y. Effect of Physiologically Active Compounds in Phlomidis Radix on Bone Regeneration. Ph.D. Thesis, Wonkwang University, Jeollabuk-do, Korea, 2004. [Google Scholar]
- Kim, J.Y.; Lee, J.I.; Song, M.; Lee, D.; Song, J.; Kim, S.Y.; Park, J.; Choi, H.Y.; Kim, H. Effects of eucommia ulmoides extract on longitudinal bone growth rate in adolescent female rats. Phytother. Res. 2015, 29, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Hunziker, E.B. Mechanism of longitudinal bone growth and its regulation by growth plate chondrocytes. Microsc. Res. Tech. 1994, 28, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Farnum, C.E.; Lee, R.; O’Hara, K.; Urban, J.P. Volume increase in growth plate chondrocytes during hypertrophy: The contribution of organic osmolytes. Bone 2002, 30, 574–581. [Google Scholar] [CrossRef]
- Breur, G.J.; VanEnkevort, B.A.; Farnum, C.E.; Wilsman, N.J. Linear relationship between the volume of hypertrophic chondrocytes and the rate of longitudinal bone growth in growth plates. J. Orthop. Res. 1991, 9, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Hansson, L.I. Daily growth in length of diaphysis measured by oxytetracycline in rabbit normally and after medullary plugging. Acta Orthop. Scand. 1967. [Google Scholar] [CrossRef]
- Kuhn, J.L.; DeLacey, J.H.; Leenellett, E.E. Relationship between bone growth rate and hypertrophic chondrocyte volume in new zealand white rabbits of varying ages. J. Orthop. Res. 1996, 14, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Gratzner, H.G. Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: A new reagent for detection of DNA replication. Science 1982, 218, 474–475. [Google Scholar] [CrossRef] [PubMed]
- Yakar, S.; Liu, J.L.; Stannard, B.; Butler, A.; Accili, D.; Sauer, B.; LeRoith, D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1999, 96, 7324–7329. [Google Scholar] [CrossRef] [PubMed]
- Sjogren, K.; Liu, J.L.; Blad, K.; Skrtic, S.; Vidal, O.; Wallenius, V.; LeRoith, D.; Tornell, J.; Isaksson, O.G.; Jansson, J.O.; et al. Liver-derived insulin-like growth factor I (IGF-I) is the principal source of IGF-I in blood but is not required for postnatal body growth in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 7088–7092. [Google Scholar] [CrossRef] [PubMed]
- De Luca, F.; Barnes, K.M.; Uyeda, J.A.; de-Levi, S.; Abad, V.; Palese, T.; Mericq, V.; Baron, J. Regulation of growth plate chondrogenesis by bone morphogenetic protein-2. Endocrinology 2001, 142, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yang, W.; de Luca, F. Insulin-like growth factor-independent effects of growth hormone on growth plate chondrogenesis and longitudinal bone growth. Endocrinology 2015, 156, 2541–2551. [Google Scholar] [CrossRef] [PubMed]
- Barrios, V.; Argente, J.; Munoz, M.T.; Pozo, J.; Chowen, J.A.; Hernandez, M. Diagnostic interest of acid-labile subunit measurement in relationship to other components of the igf system in pediatric patients with growth or eating disorders. Eur. J. Endocrinol. 2001, 144, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, R.G. Is growth hormone deficiency a viable diagnosis? J. Clin. Endocrinol. MeTab. 1997, 82, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Van der Kaay, D.C.; Hendriks, A.E.; Ester, W.A.; Leunissen, R.W.; Willemsen, R.H.; de Kort, S.W.; Paquette, J.R.; Hokken-Koelega, A.C.; Deal, C.L. Genetic and epigenetic variability in the gene for IGFBP-3 (IGFBP3): Correlation with serum IGFBP-3 levels and growth in short children born small for gestational age. Growth Horm. IGF Res. 2009, 19, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Costalonga, E.F.; Antonini, S.R.; Guerra-Junior, G.; Mendonca, B.B.; Arnhold, I.J.; Jorge, A.A. The -202 a allele of insulin-like growth factor binding protein-3 (IGFBP3) promoter polymorphism is associated with higher IGFBP-3 serum levels and better growth response to growth hormone treatment in patients with severe growth hormone deficiency. J. Clin. Endocrinol. Metab. 2009, 94, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Firth, S.M.; Baxter, R.C. Cellular actions of the insulin-like growth factor binding proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Sample of the P. umbrosa extract is available from the authors.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height (μm) | Control | rhGH 200 μg/kg (s.c.) | P. umbrosa 300 mg/kg (p.o.) |
|---|---|---|---|
| Overall growth plate | 453.8 ± 28.6 | 536.9 ± 29.5 *** | 508.6 ± 28.9 ** |
| Resting zone | 27.3 ± 3.5 | 25.1 ± 2.9 | 29.8 ± 2.5 |
| Proliferative zone | 111.7 ± 23.1 | 110.0 ± 30.0 | 120.9 ± 5.9 |
| Hypertrophic zone | 304.0 ± 46.1 | 384.0 ± 54.2 ** | 345.0 ± 22.0 * |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Kim, Y.-S.; Song, J.; Kim, H.S.; Lee, H.J.; Guo, H.; Kim, H. Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats. Molecules 2016, 21, 461. https://doi.org/10.3390/molecules21040461
Lee D, Kim Y-S, Song J, Kim HS, Lee HJ, Guo H, Kim H. Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats. Molecules. 2016; 21(4):461. https://doi.org/10.3390/molecules21040461
Chicago/Turabian StyleLee, Donghun, Young-Sik Kim, Jungbin Song, Hyun Soo Kim, Hyun Jung Lee, Hailing Guo, and Hocheol Kim. 2016. "Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats" Molecules 21, no. 4: 461. https://doi.org/10.3390/molecules21040461
APA StyleLee, D., Kim, Y.-S., Song, J., Kim, H. S., Lee, H. J., Guo, H., & Kim, H. (2016). Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats. Molecules, 21(4), 461. https://doi.org/10.3390/molecules21040461