
The Inhibition Effect of Cell DNA Oxidative Damage and LDL Oxidation by Bovine Colostrums


Abstract
:1. Introduction
2. Results
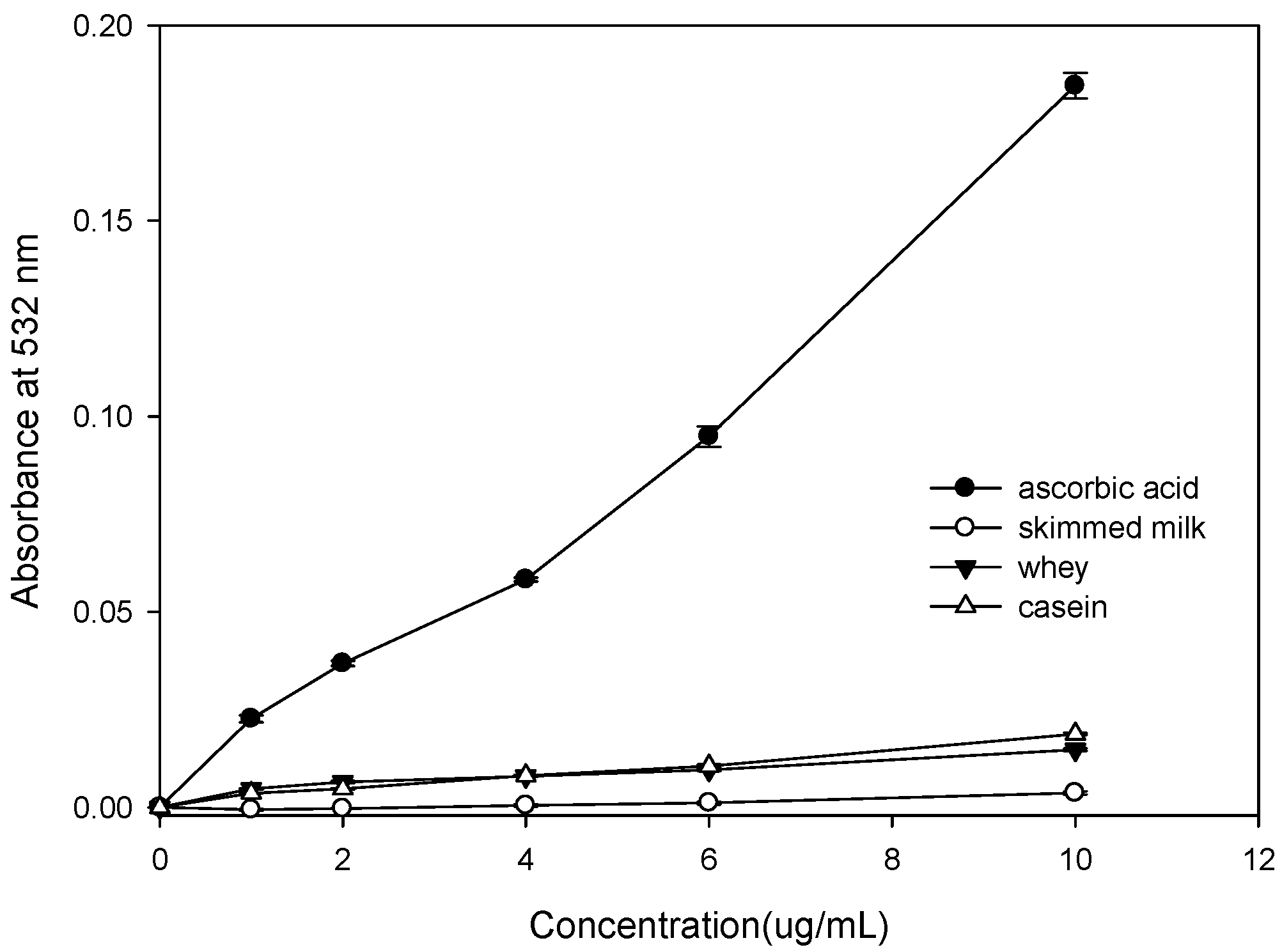
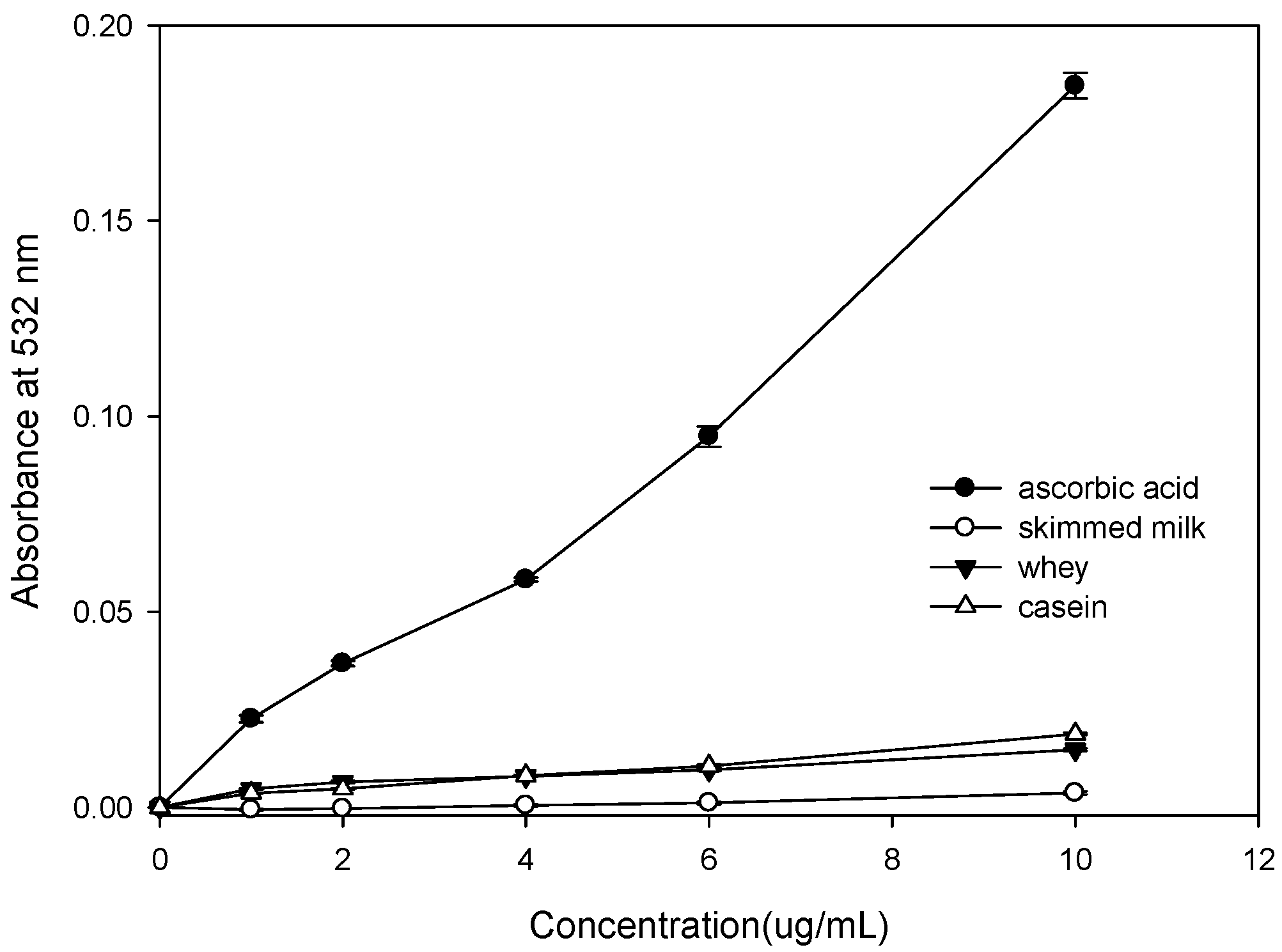
2.1. Effect of Bovine Colostrums Protein on the Fenton Reaction-Induced Oxidative Damage of Deoxyribose
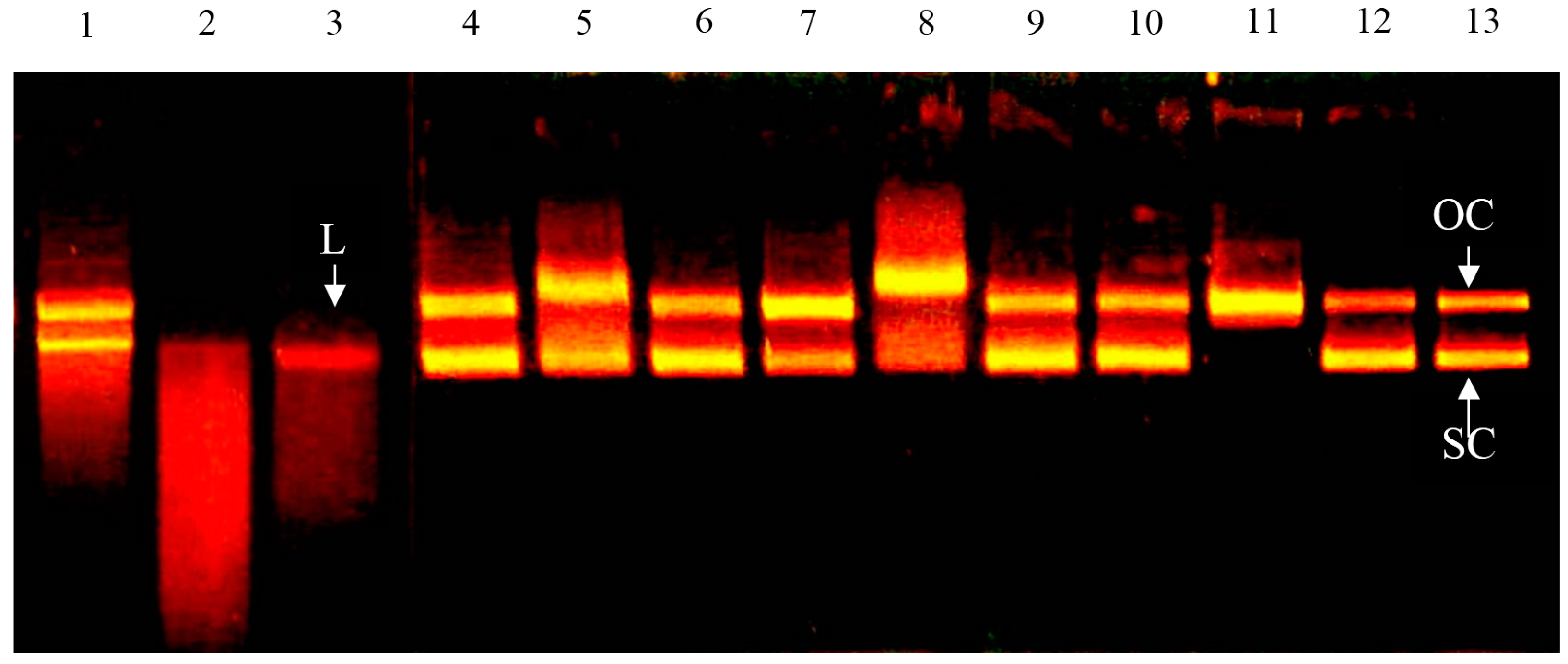
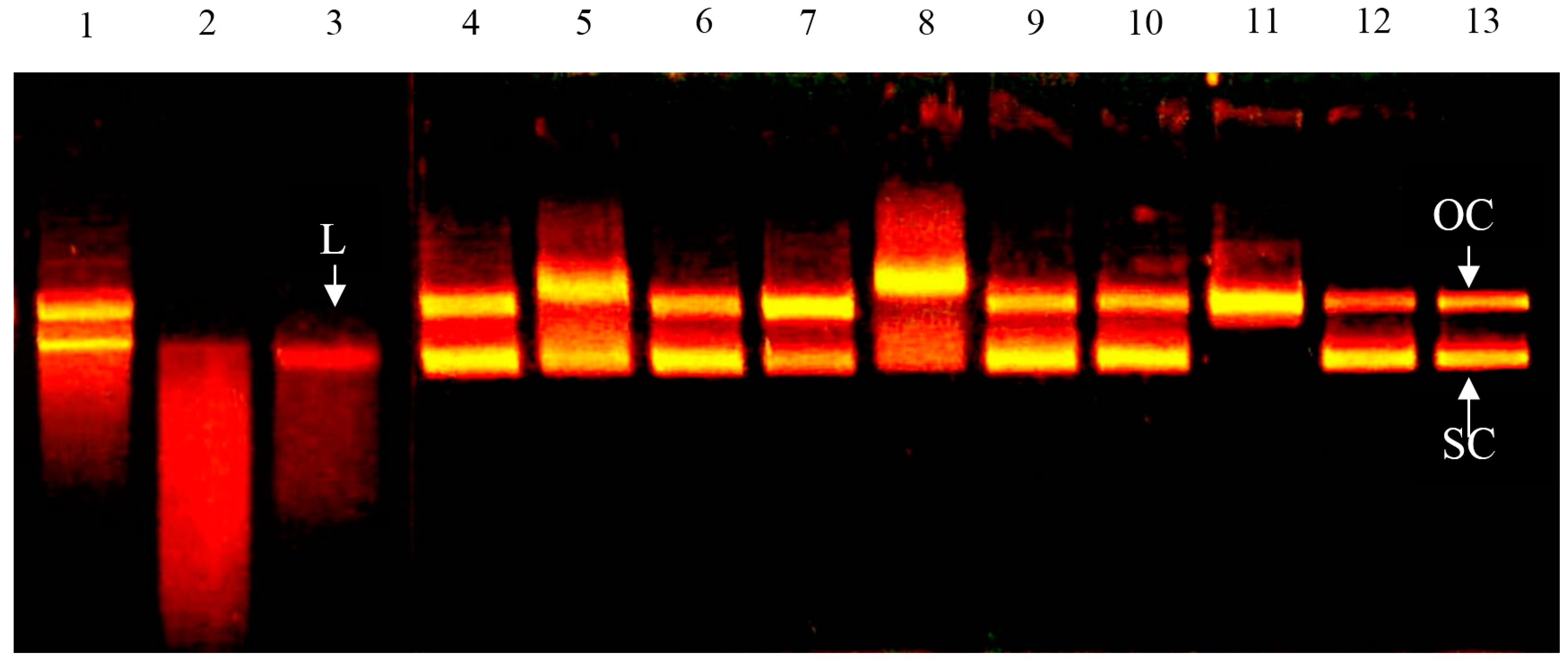
2.2. Effect of Bovine Colostrums Protein on DNA Single-Strand Cleavage Induced by Fenton Reaction
2.3. Effect of Bovine Colostruns Protein on the Oxidation of 2′-dG to 8-OH-2′-dG Induced by Fenton Reaction
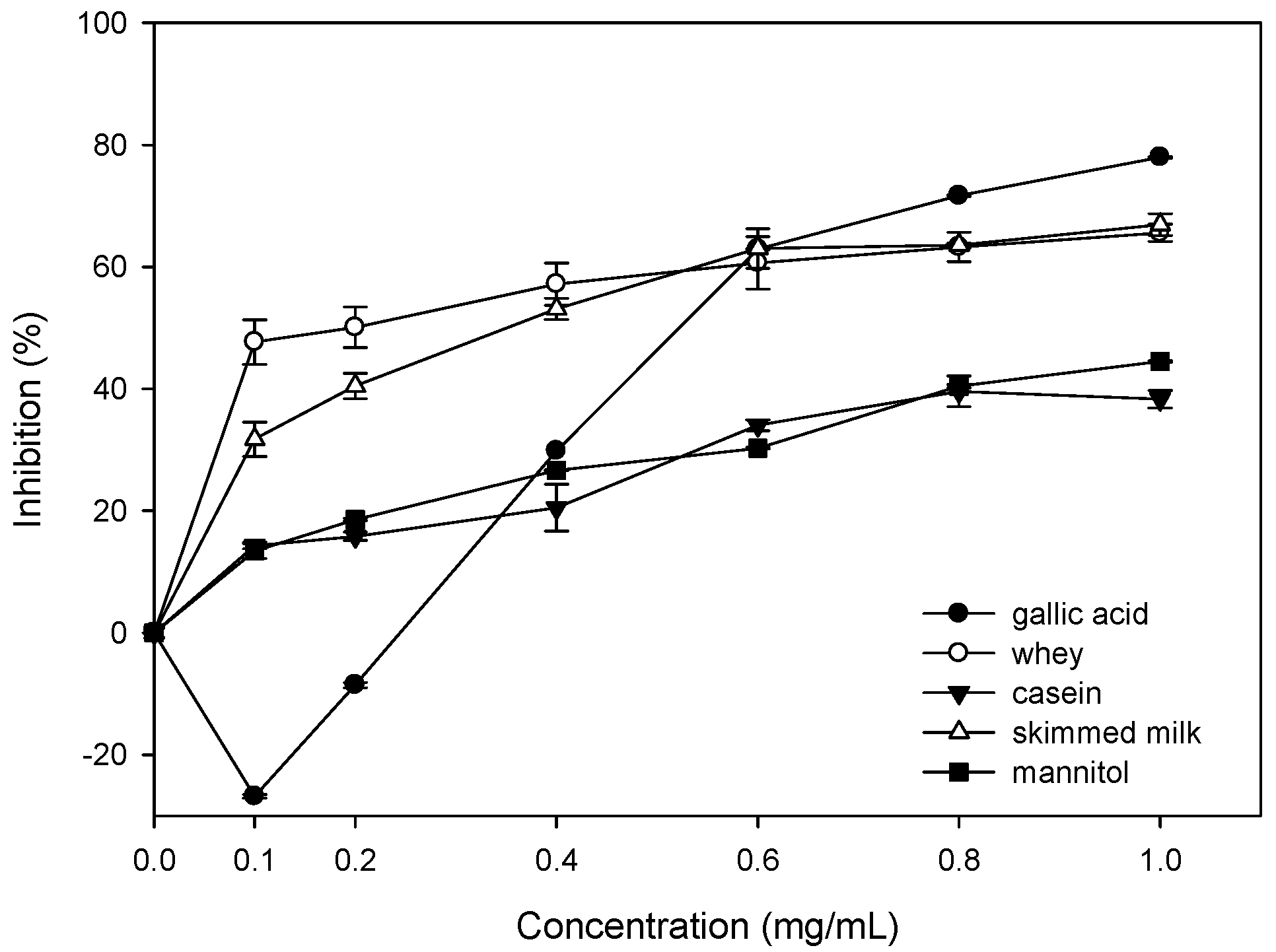
2.4. Effect of Bovine Colostrums Protein on Bleomycin-Dependent DNA Damage
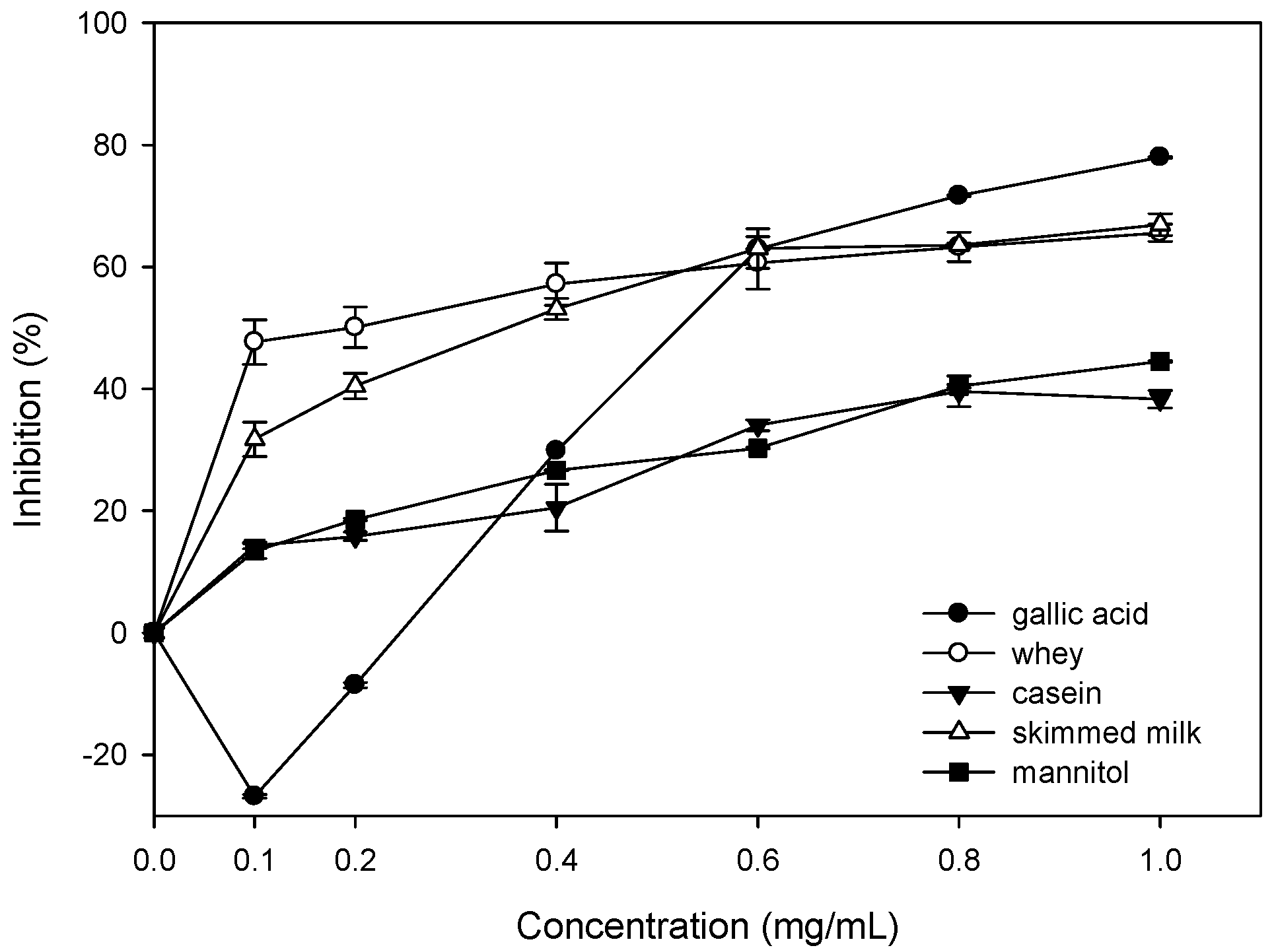
2.5. Inhibition of Oxidative Damages of Biomolecules by Bovine Colostrums Protein
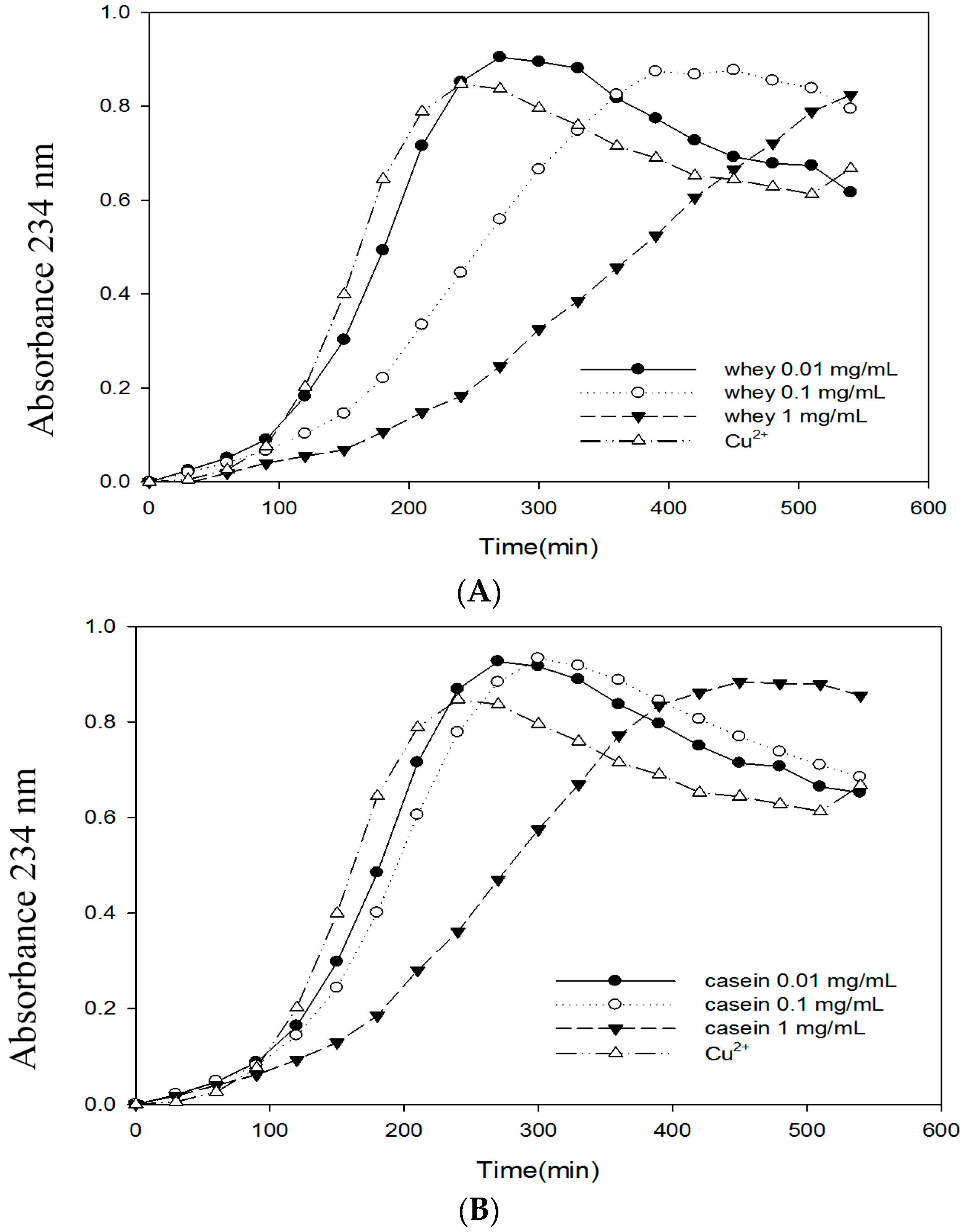
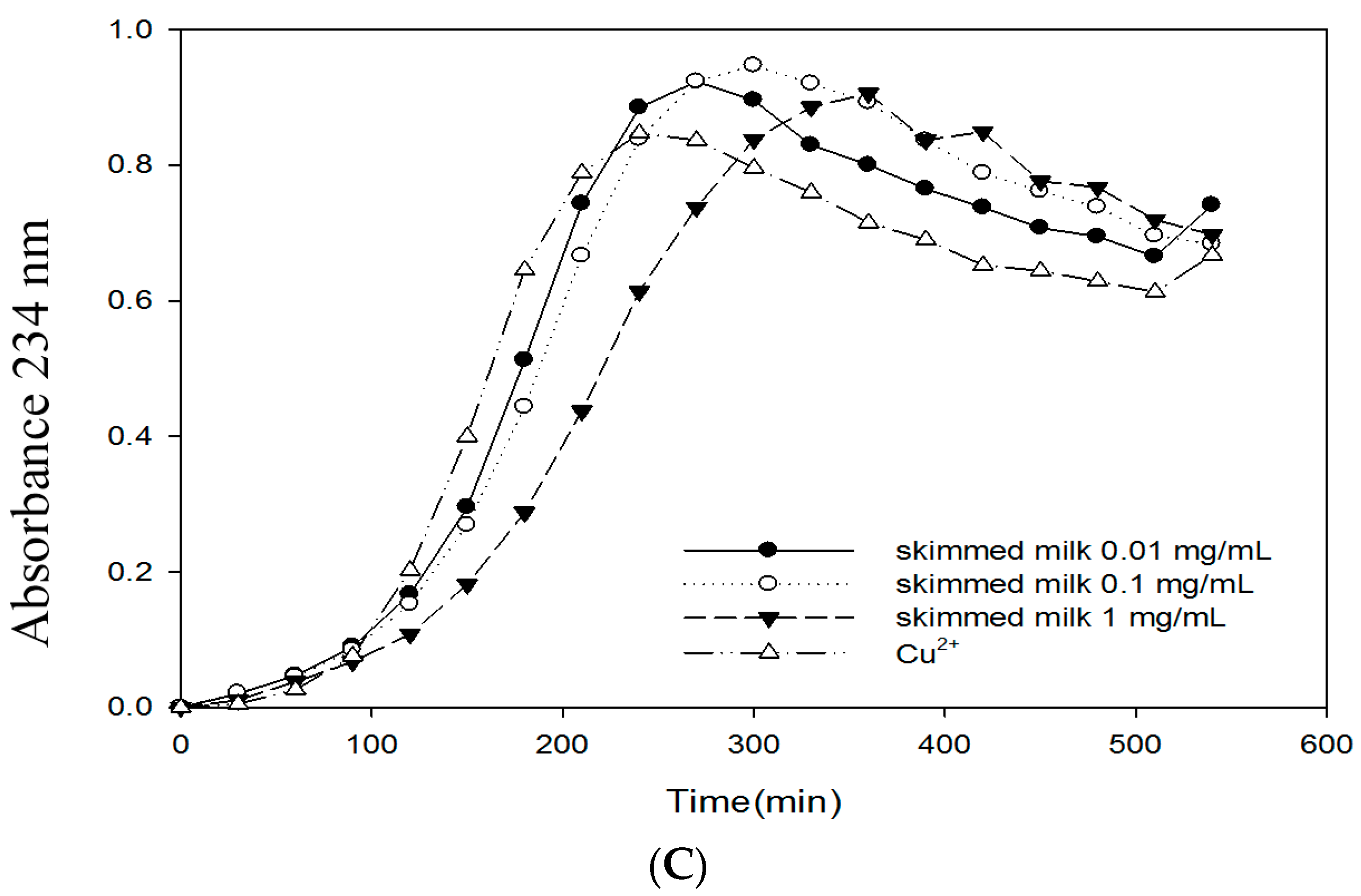
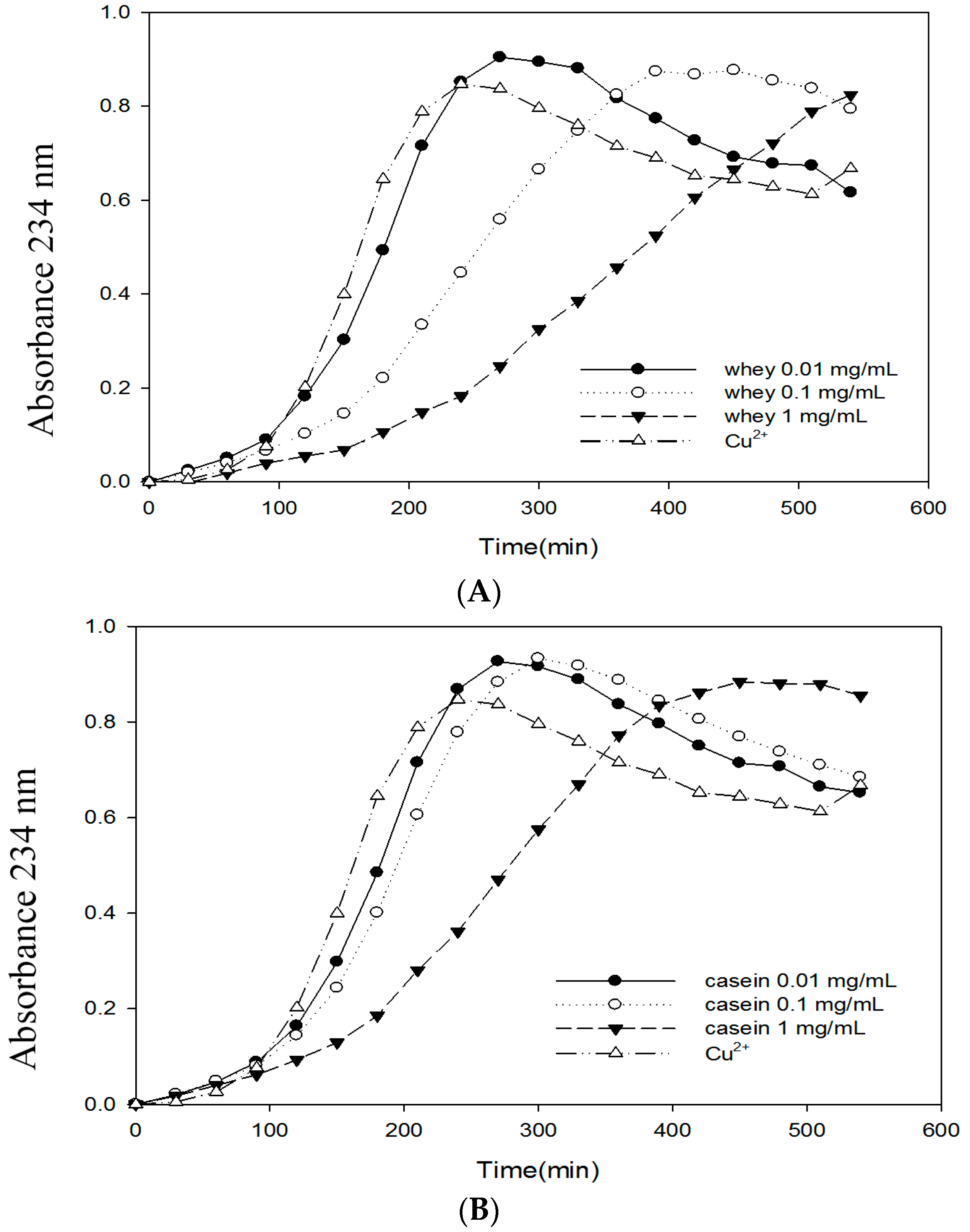
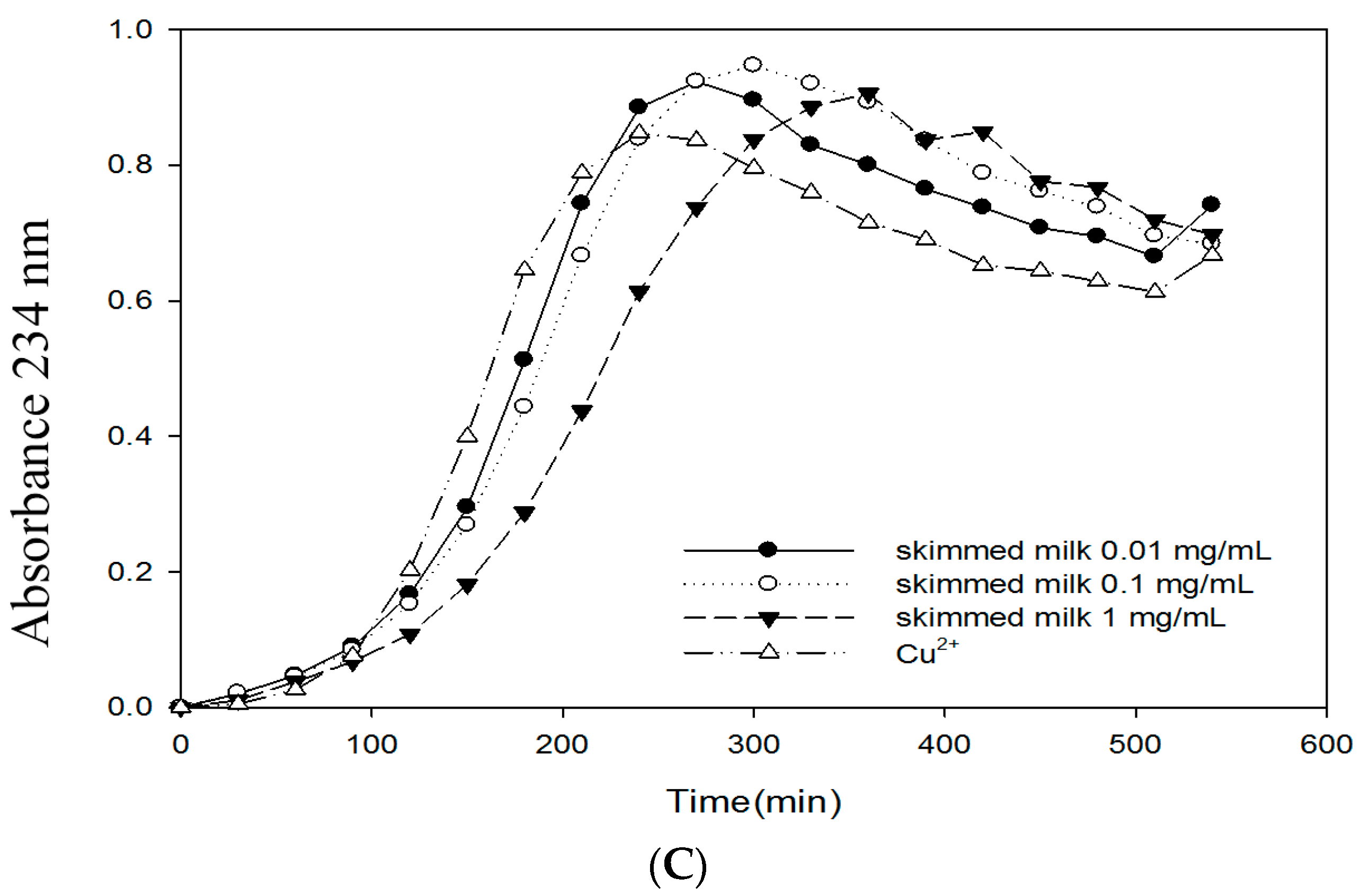
2.6. Effects of Bovine Colostrums Protein on the Formation of TBARS and Conjugated Dienes on LDL Oxidation Induced by Cu2+
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Skimmed Milk, Caseins and Whey Proteins
4.3. Effect of Bovine Colostrums Proteinon Deoxyribose Damage (Fenton Reaction)
4.4. Effect of Bovine Colostrums Protein on DNA Damage (Fenton Reaction)
4.5. Effect of Bovine Colostrums Protein on Oxidation of 2′-Deoxyguanosine (Fenton Reaction)
4.6. Effect of Bovine Colostrums Protein on Bleomycin-Dependent DNA Damage
4.7. Inhibition of Oxidative Damages of Biomolecules by Bovine Colostrums Protein
4.8. LDL Preparation
4.9. LDL Oxidation
4.10. Thiobarbituric Acid Reactive Substances (TBARS)
4.11. Conjugated Diene
4.12. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, X.; Jaio, C.; Wang, T.; Yu, Z. Study on DNA damage induced by the reactive oxygen species generated in situ based on the multi-walled carbon nanotubes and hemoglobin. J. Electroanal. Chem. 2016, 767, 182–187. [Google Scholar] [CrossRef]
- Oliveira, S.C.B.; Oliveira-Brett, A.M. In situ DNA oxidative damage by electrochemically generated hydroxyl free radicals on a boron-doped diamond electrode. Langmuir 2012, 28, 4896–4901. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Sugihara, N.; Kishimoto, Y.; Saita, E.; Taguchi, C.; Kobayashi, M.; Ichitani, M.; Ukawa, Y.; Sagesaka, Y.M.; Suzuki, E.; Kondo, K. Green tea catechins prevent low-density liporptein oxidation via their accumulation in low-density lipoprotein particles in humans. Nutr. Res. 2016, 36, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D. The LDL, modification hypothesis of atherogenesis: An update. J. Lipid Res. 2009, 50, S376–S381. [Google Scholar] [CrossRef] [PubMed]
- Huxley, P.R.; Neil, H.A. The relation between dietary flavonols and coronary heart disease mortality: A meta-analysis of prospective cohort studied. Eur. J. Clin. Nutr. 2009, 57, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Schildermann, P.A.E.L.; Ten Hoor, F.; Kleinjas, J.C.S. Induction of oxidative DNA damage and early lesions in rat gastro-intestinal epithelium in relation to prostaglandin H synthase-mediated metabolism of butylatedhydroxyanisole. Food Chem. Toxicol. 1995, 33, 99–109. [Google Scholar] [CrossRef]
- Chen, C.W.; Chiang, S.H.; Wang, S.Y.; Lin, Y.T.; Lin, F.Y.; Chang, C.Y. Inhibitory effects of bovine colostrum protein hydrolysates on human leukemic U937 cell growth. J. Food Drug Anal. 2011, 20, 699–740. [Google Scholar]
- Chen, C.W.; Chiang, S.H.; Wang, S.Y.; Lin, Y.T.; Chang, C.Y. Growth Inhibition and differentiating effects of protein hydrolysates from bovine colostrums on human leukemic U937 cell. J. Food Biochem. 2013, 37, 8–17. [Google Scholar] [CrossRef]
- Palyford, R.D.; Mcdonald, C.E.; Johnson, W.S. Colostrums and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am. J. Clin. Nutr. 2000, 72, 5–14. [Google Scholar]
- Ogawa, J.; Sasahara, A.; Yoshida, T.; Sira, M.M.; Futatani, T.; Kanegane, H.; Miyawaki, T. Role of transforming growth factor in breast milk for initiation of IgA production in newborn infants. Early Hum. Dev. 2004, 77, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Zhiwei, Y.; Palkovicova, L.; Droban, B.; Petrik, J.; Kocan, A.; Trnovec, T.; Picciotto, I.H. Comparison of organochlorine compound concentrations in colostrums and mature milk. Chemosphere 2007, 66, 1012–1018. [Google Scholar]
- Hagiwara, K.; Domi, M.; Ando, J. Bovine colostral CD-8 positive cells are potent IFN-γ producing cells. Vet. Immunol. Immunopathol. 2008, 124, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Murcia, A.; Butler, J.; Halliwell, B. Evaluation of antioxidant and prooxidant actions of gallic acid and its derivatives. J. Agric. Food Chem. 1993, 41, 1880–1885. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Yen, G.C. Antioxidant actions of Du-Zhong (Eucommiaulmoides Oliv.) toward oxidative damage in biomolecules. Life Sci. 2000, 66, 1387–1400. [Google Scholar] [CrossRef]
- Kobayashi, S.; Uead, K.; Komano, T. The effects of metal ions on the DNA damage induced by hydrogen peroxide. Agric. Biol. Chem. 1990, 54, 69–76. [Google Scholar] [PubMed]
- Smith, C.; Halliwell, B.; Aruoma, O.I. Protection by albumin against the pro-oxidant actions of phenolic dietary components. Food Chem. Toxicol. 1992, 30, 483–489. [Google Scholar] [CrossRef]
- Treml, J.; Šmejkal, K. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Chiang, S.H.; Chang, C.Y. Antioxidant properties of bovine colostrum casein and whey protein. J. Food Drug Anal. 2005, 13, 57–63. [Google Scholar]
- Aruoma, O.I.; Halliwell, B.; Williamson, G. In vitro methods for characterizing potential prooxidant and antioxidant actions of nonnutritive substances in plant foods. In Antioxidant Methodology In Vivo and In Vitro Concepts; Aruoma, O.I., Cuppett, S.L., Eds.; AOCS press: Champaign, IL, USA, 1998; Volume 10, pp. 173–204. [Google Scholar]
- Yoshinoa, M.; Hanedaa, M.; Naruseb, M.; Htaya, H.H.; Iwataa, S.; Tsubouchia, R.; Murakamia, K. Prooxidant action of gallic acid compounds: Copper-dependent strand breaks and the formation of 8-hydroxy-2′-deoxyguanosine in DNA. Toxicol. In Vitro 2002, 16, 705–709. [Google Scholar] [CrossRef]
- Erdem, G.; Öner, C.; Önal, A.M.; Kisakürek, D.; Öhüs, A. Free radical mediated interaction of ascorbic acid and ascorbate/Cu(II) with viral and plasmid DNAs. J. Biosci. 1994, 19, 9–17. [Google Scholar] [CrossRef]
- Stadtman, E.R. Ascorbic acid and oxidative inactivation of protein. Am. J. Clin. Nutr. 1991, 54, 1125–1128. [Google Scholar]
- Hu, M.L.; Shih, M.K. Ascorbic acid inbibits lipid peroxidation but enhances DNA damage in rat liver nuclei incubated with iron ions. Free Radic. Res. 1997, 26, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Sai, K.; Umemura, T.; Hasegawa, R.; Kurokawa, Y. Inhibitory effects of vitamin E and ellagic acid on 8-hydroxydeoxyguanosine formation in liver nuclear DNA of rats treated with 2-nitropropane. Cancer Lett. 1995, 91, 139–144. [Google Scholar] [CrossRef]
- Kasai, H.; Nishimura, S. Hydroxylation of deoxyguanosine at the C-8 position by ascorbic acid and other reducing agents. Nucleic Acids Res. 1984, 12, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Shibutani, S.; Takeshita, M.; Grollman, A.P. Insertion of specific bases during DNA synthesis past the oxidation-damage base 8-oxo-dG. Nature 1991, 349, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, M.K.; Ames, B. Assay for 8-hydroxy-2′-deoxyguanosine: A biomarker of in vivo oxidative DNA damage. Free Radic. Biol. Med. 1991, 10, 211–216. [Google Scholar] [CrossRef]
- Yen, G.C.; Chen, H.Y.; Peng, H.H. Antioxidant and pro-oxidant effects of various tea extracts. J. Agric. Food Chem. 1997, 45, 30–34. [Google Scholar] [CrossRef]
- Povirk, L.F.; Austin, M.J.F. Genotoxicity of bleomycin. Mutat. Res. 1991, 257, 127–143. [Google Scholar] [CrossRef]
- Huet, J.; Laval, F. Potentiation of cell killing by inhibitors of poly (adenosinediphosphate-ribose) synthesis in bleomycin-treated Chineses hamster ovary cells. Cancer Res. 1985, 45, 987–991. [Google Scholar] [PubMed]
- Moran, J.F.; Klucas, R.V.; Grayer, R.J.; Abian, J.; Becana, M. Complexes of iron with phenolic compounds from soybean nodules and other legume tissues: Prooxidant and antioxidant properties. Free Radic. Biol. Med. 1997, 22, 861–870. [Google Scholar] [CrossRef]
- Yamanaka, N.; Oda, O.; Nagao, S. Prooxidant activity of caffeic acid, dietary non-flavonoid phenolic acid, on Cu2+ induced low density lipoprotein oxidation. FEBS Lett. 1997, 405, 186–190. [Google Scholar] [CrossRef]
- Yagi, K. A simple fluorometric assay for lipid peroxides in blood serum or plasma. In CRC Hand Book of Free Radicals and Antioxidants in Biomedicine; Miquel, J., Quintanilha, A.T., Weber, H., Eds.; CRC Press: Boca Raton, FL, USA, 1989; Volume 3, pp. 215–218. [Google Scholar]
- Agin, D.; Gallagher, D.; Wang, J. Effects of whey protein and resistance exercise on body cell mass, muscle strength, and quality of life in women with HIV. AIDS 2001, 7, 431–440. [Google Scholar] [CrossRef]
- Bounous, G. Whey protein concentrate (WPC) and glutathione modulation in cancer treatment. Anticancer Res. 2000, 20, 4785–4792. [Google Scholar] [PubMed]
- Watanabe, A.; Okada, K.; Shimizu, Y. Nutritional therapy of chronic hepatitis by whey protein (non-heated). J. Med. 2000, 31, 283–302. [Google Scholar] [PubMed]
- Lands, L.C.; Grey, V.L.; Smountas, A.A. Effect of supplementation with a cysteine donor on muscular performance. J. Appl. Physiol. 1999, 87, 1381–1385. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compounds bovine colostrum protein are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Addition to RM * | 8-OH-2′-dG (μg/mL) | ||
|---|---|---|---|
| Whey | Casein | Skimmed Milk | |
| PBS buffer | 0.36 e,** | 0.36 b,** | 0.36 d,** |
| 15 mM Ascorbic acid | 10.03 a | 10.03 a | 10.03 a |
| 1 mg/mL | 0.46 b,c | 0.33 b | 1.45 b,c |
| 2 mg/mL | 0.33 d | 0.24 b | 1.46 b,c |
| 4 mg/mL | 0.35 c,d | 0.32 b | 1.38 b,c |
| 6 mg/mL | 0.24 d | 0.38 b | 1.24 c |
| 8 mg/mL | 0.55 b | 0.38 b | 1.36 b,c |
| 10 mg/mL | 0.33 d | 0.41 b | 1.49 b |
| Addition to RM * | Absorbance at 532 nm | Inhibition (%) | 8-OH-2′-dG (μg/mL) | Inhibition (%) |
|---|---|---|---|---|
| Ascorbic acid | 0.186 a,** | 10.13 a,** | ||
| Whey | 0.171 c | 8.41 | 8.12 c | 20.01 |
| Casein | 0.174 b | 6.62 | 8.29 c | 18.28 |
| Skimmed milk | 0.182 a | 2.33 | 9.02 b | 11.07 |
| Concentration (mg/mL) | Whey | Casein | Skimmed Milk | Whey | Casein | Skimmed Milk |
|---|---|---|---|---|---|---|
| TBARS (n mol/mL) | Lag Time * (min) | |||||
| Blank | 5.13 ± 0.01 a,* | 5.13 ± 0.01 a | 5.13 ± 0.01 a | 90–120 | 90–120 | 90–120 |
| 1 mg/mL | 4.19 ± 0.01 b | 4.07 ± 0.06 b | 4.72 ± 0.05 a | 180–210 | 150–180 | 120–150 |
| 0.1 mg/mL | 4.13 ± 0.05 b | 4.21 ± 0.09 b | 4.80 ± 0.14 a | 120–150 | 90–120 | 90–120 |
| 0.01 mg/mL | 4.42 ± 0.06 b | 4.53 ± 0.01 b | 4.72 ± 0.05 a | 90–120 | 90–120 | 90–120 |
| 0.001 mg/mL | 4.98 ± 0.02 a | 4.85 ± 0.05 a | 5.02 ± 0.06 a | - | - | - |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-W.; Chang, C.-Y.; Chiang, S.-H. The Inhibition Effect of Cell DNA Oxidative Damage and LDL Oxidation by Bovine Colostrums. Molecules 2016, 21, 1378. https://doi.org/10.3390/molecules21101378
Chen C-W, Chang C-Y, Chiang S-H. The Inhibition Effect of Cell DNA Oxidative Damage and LDL Oxidation by Bovine Colostrums. Molecules. 2016; 21(10):1378. https://doi.org/10.3390/molecules21101378
Chicago/Turabian StyleChen, Chih-Wei, Chi-Yue Chang, and Shu-Hua Chiang. 2016. "The Inhibition Effect of Cell DNA Oxidative Damage and LDL Oxidation by Bovine Colostrums" Molecules 21, no. 10: 1378. https://doi.org/10.3390/molecules21101378
APA StyleChen, C.-W., Chang, C.-Y., & Chiang, S.-H. (2016). The Inhibition Effect of Cell DNA Oxidative Damage and LDL Oxidation by Bovine Colostrums. Molecules, 21(10), 1378. https://doi.org/10.3390/molecules21101378