
Turkish Scorzonera Species Extracts Attenuate Cytokine Secretion via Inhibition of NF-κB Activation, Showing Anti-Inflammatory Effect in Vitro

 ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
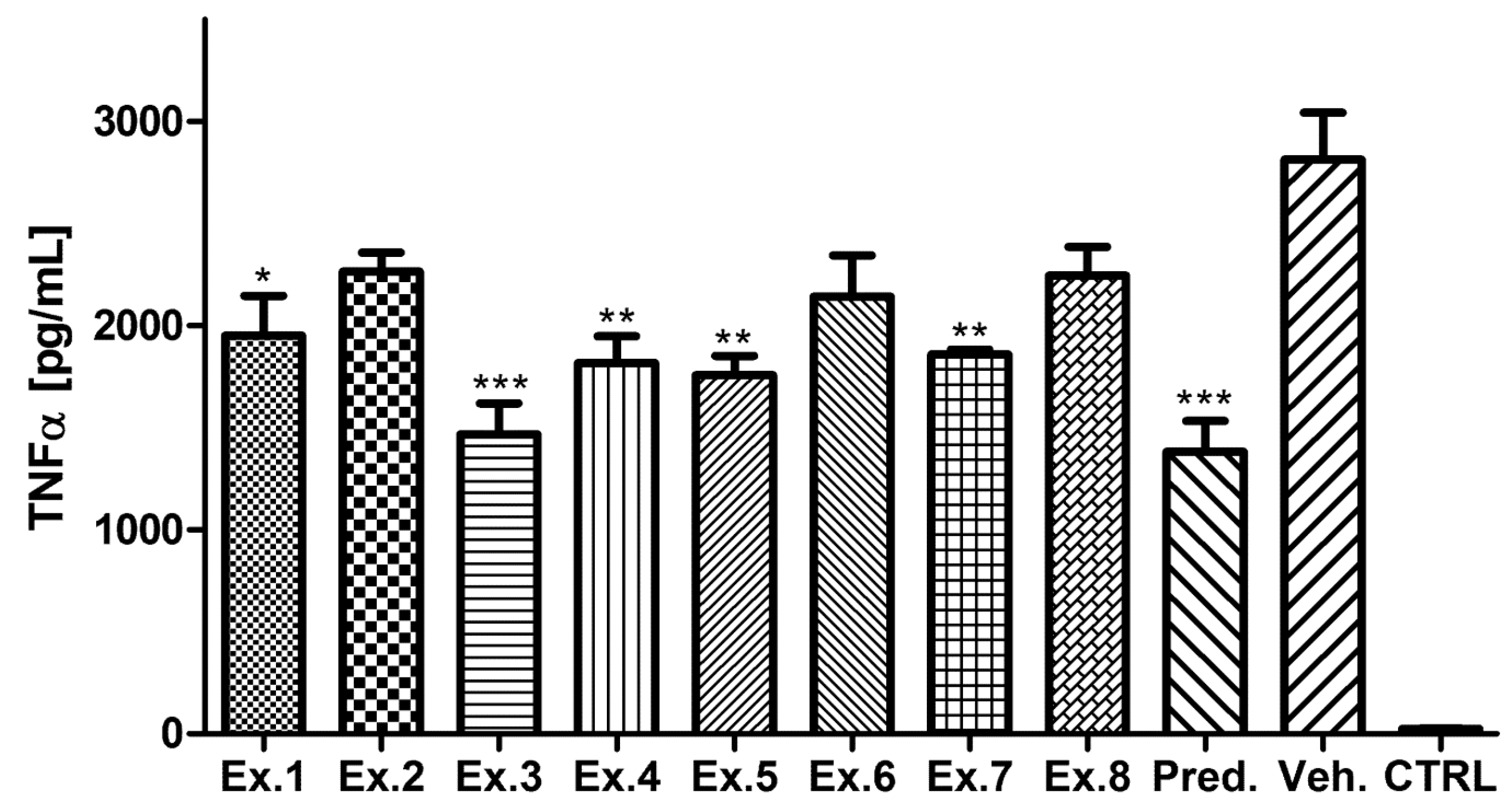
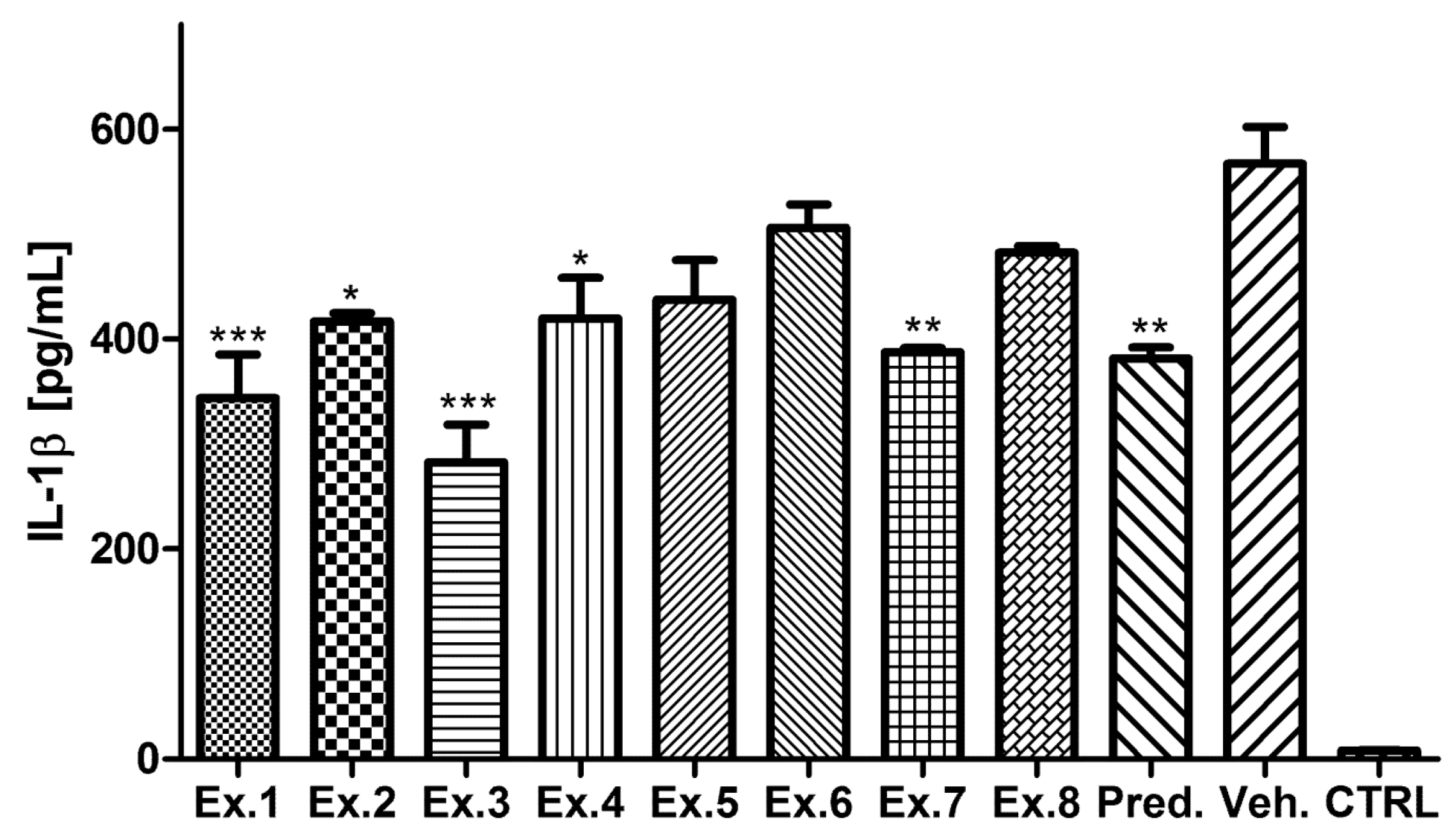
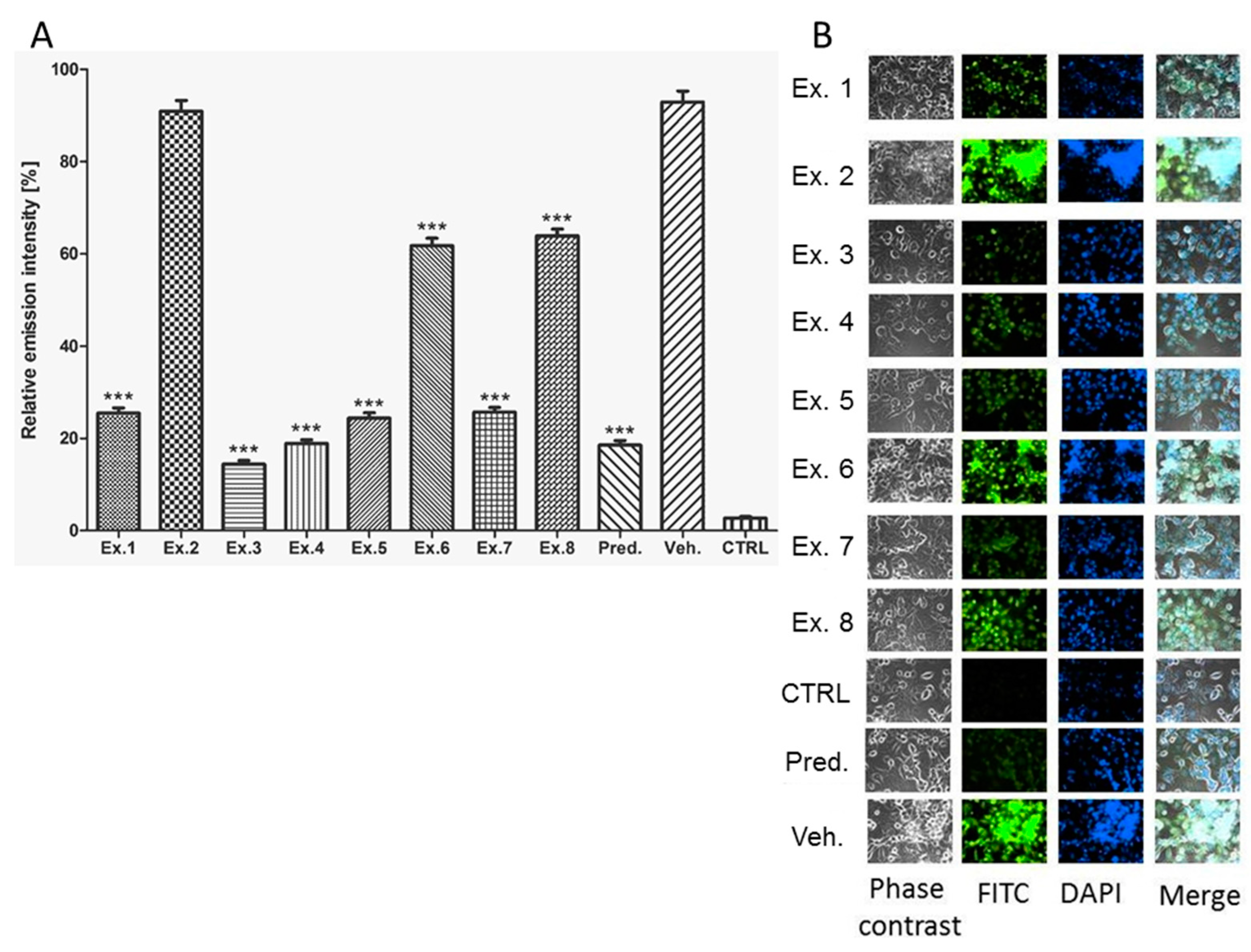
3. Experimental Section
3.1. Plant Material

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Locality | Date | Identification/Voucher Specimen No. | Code Name |
|---|---|---|---|---|
| S. latifolia (Fisch. & Mey.) DC. | Kars, Arpaçay | 2005 | H. Duman/23830 | Ex. 1 |
| S. cana (C.A. Meyer) Hoffm. var. jacquiniana (W. Koch) Chamb. | Ankara, Çamlıdere | 2008 | H. Duman/23834 | Ex. 2 |
| S. tomentosa L. | Yozgat, Akdağmadeni | 2005 | H. Duman/23841 | Ex. 3 |
| S. mollis Bieb. ssp. szowitzii (DC.) Chamb. | Ankara, Kızılcahamam | 2006 | M. Koyuncu/23844 | Ex. 4 |
| S. eriophora DC. | Ankara,Çubuk | 2007 | H. Duman/23832 | Ex. 5 |
| S. incisa DC. | Konya, Ermenek | 2005 | H. Duman/23833 | Ex. 6 |
| S. cinerea Boiss. | Sivas, Çetinkaya | 2005 | H. Duman/23829 | Ex. 7 |
| S. parviflora Jocq. | Ankara, Gölbaşı | 2008 | H. Duman/25894 | Ex. 8 |
3.2. Extraction of Plant Materials for Activity
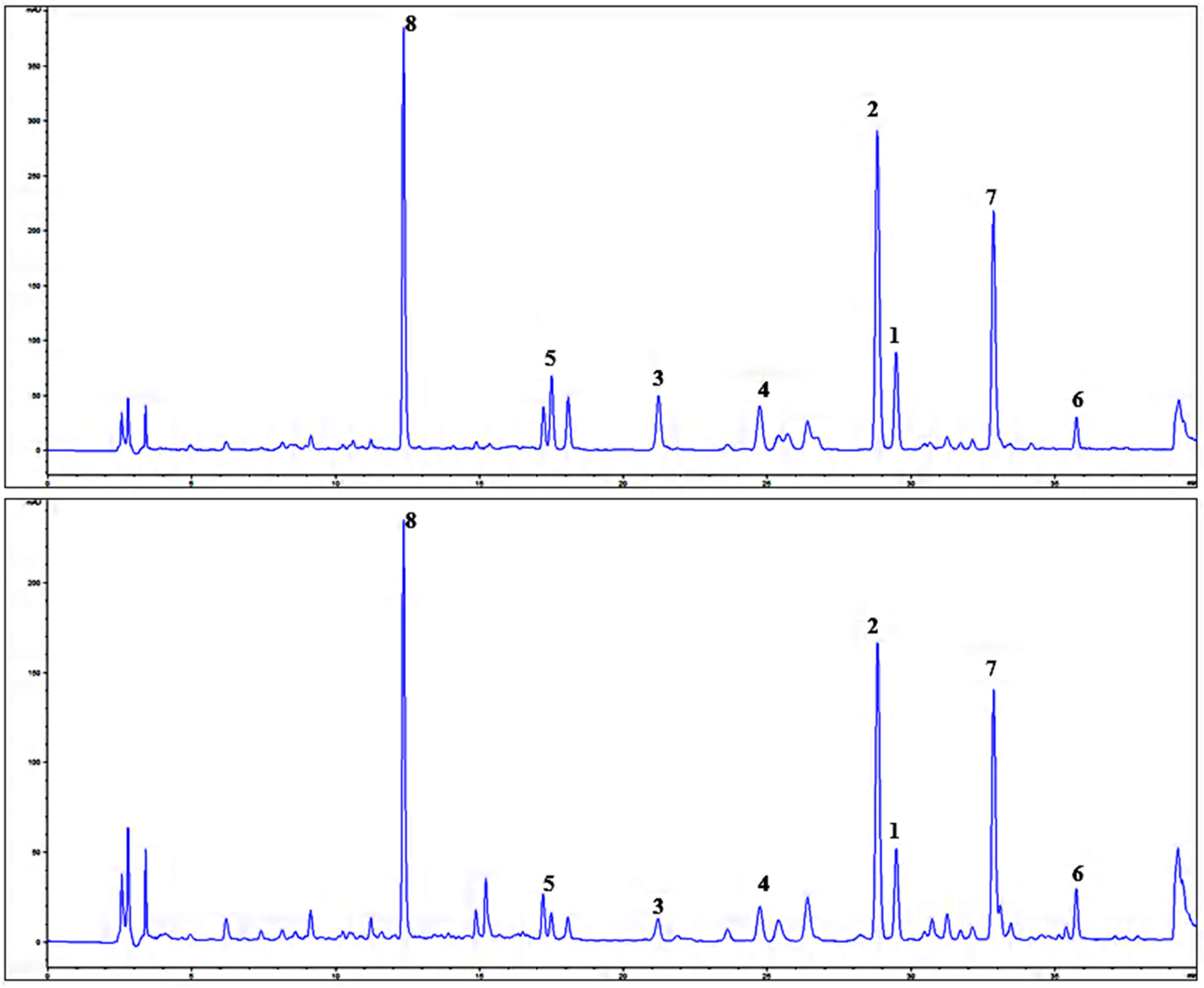
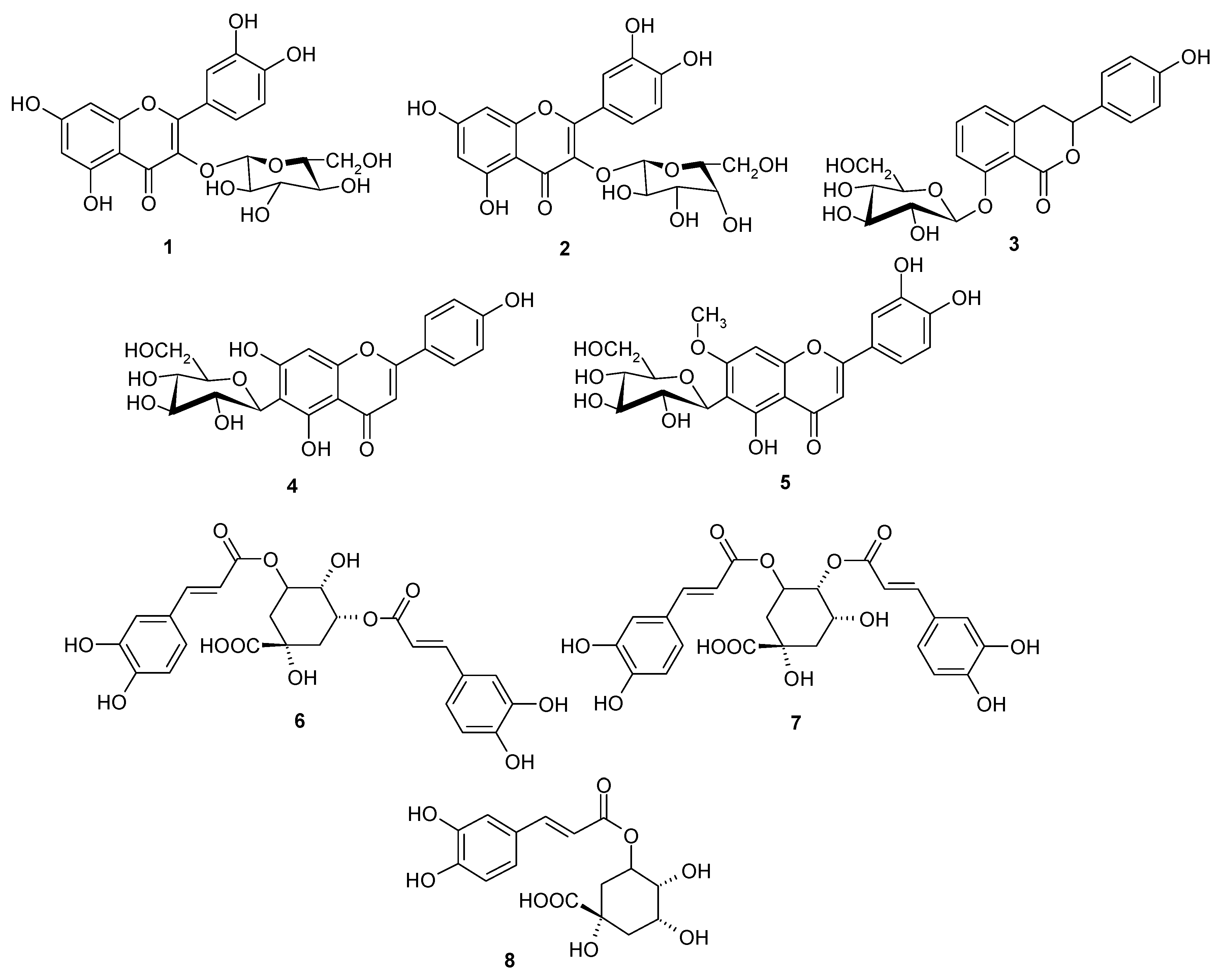
3.3. Isolation and Identification of Compounds
3.4. HPLC Analysis
3.5. Anti-Inflammatory Assay
3.5.1. Compounds for In Vitro Anti-Inflammatory Assays

3.5.2. TNF-α and IL-1β Inhibitory Activity Measurement
3.5.3. Measurement of Inhibition of Activation of NF-κB
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015. [Google Scholar] [CrossRef] [PubMed]
- Yesilada, E. Evaluation of the anti-inflammatory activity of the Turkish medicinal plant Sambucus ebulus. Chem. Nat. Prod. 1997, 33, 539–540. [Google Scholar]
- Bandgar, B.P.; Patil, S.A.; Totre, J.V.; Korbad, B.L.; Gacche, R.N.; Hote, B.S.; Jalde, S.S.; Chavan, H.V. Synthesis and biological evaluation of nitrogen-containing benzophenone analogues as TNF-a and IL-6 inhibitors with antioxidant activity. Bioorg. Med. Chem. Lett. 2010, 20, 2292–2296. [Google Scholar] [CrossRef] [PubMed]
- Leiro, J.M.; Varela, M.; Piazzon, M.C.; Arranz, J.A.; Noya, M.; Lamas, J. The anti-inflammatory activity of the polyphenol resveratrol may be partially related to inhibition of tumour necrosis factor-α (TNF-α) pre-mRNA splicing. Mol. Immunol. 2010, 47, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Satti, N.K.; Sharma, V.K.; Dutt, P.; Suri, K.A.; Bani, S. Amelioration of inflammatory responses by chlorogenic acid via suppression of pro-inflammatory mediators. J. Appl. Pharm. Sci. 2011, 1, 67–75. [Google Scholar]
- Benbarek, H.; Deby-Dupont, G.; Deby, C.; Serteyn, D. Direct stimulation of the oxidative activity of isolated equine neutrophils by TNF-α and IL-1β. Vet. Immunol. Immunopathol. 2008, 121, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.P.N.; Kabir, S.N.; Mohamed, T. Nonsteroidal Anti-Inflammatory Drugs (NSAIDs): Progress in Small Molecule Drug Development. Pharmaceuticals 2010, 3, 1530–1549. [Google Scholar] [CrossRef]
- Zelova, H.; Hosek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, M.; Wallace, D.; Tith, S.; Arrhenius, T.; Kashiwagi, H.; Ono, Y.; Ishikawa, A.; Sato, H.; Kozono, T.; et al. Discovery and structure-activity relationship of coumarin derivatives as TNF-α inhibitors. Bioorg. Med. Chem. Lett. 2004, 14, 2411–2415. [Google Scholar] [PubMed]
- Folmer, F.; Jaspars, M.; Solano, G.; Cristofanon, S.; Henry, E.; Tabudravu, J.; Black, K.; Green, D.H.; Küpper, F.C.; Aalbersberg, W.; et al. The inhibition of TNF-α-induced NF-κB activation by marine natural products. Biochem. Pharmacol. 2009, 78, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Nam, N.H.; Jae, Y.Y. NF-κB Inhibitory Activities of the Methanol Extracts and some Constituents therein of some Vietnamese Medicinal Plants. Sci. Pharm. 2009, 77, 389–399. [Google Scholar] [CrossRef]
- Zidorn, C.; Ellmerer, E.P.; Sturm, S.; Stuppner, H. Tyrolobibenzyls E and F from Scorzonera humilis and distribution of caffeic acid derivatives, lignans and tyrolobibenzyls in European taxa of the subtribe Scorzonerinae (Lactuceae, Asteraceae). Phytochemistry 2003, 63, 61–67. [Google Scholar] [CrossRef]
- Tsevegsuren, N.; Edrada, R.A.; Lin, W.; Ebel, R.; Torre, C.; Ortlepp, S.; Wray, V.; Proksch, P. Biologically Active Natural Products from Mongolian Medicinal Plants Scorzonera divaricata and Scorzonera pseudodivaricata. J. Nat. Prod. 2007, 70, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Edrada-Ebel, R.A.; Tsevegsuren, N.; Sendker, J.; Braun, M.; Wray, V.; Lin, W.; Proksch, P. Dihydrostilbene derivatives from the Mongolian medicinal plant Scorzonera radiata. J. Nat. Prod. 2009, 72, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Zidorn, C.; Ellmerer-Müller, E.P.; Stuppner, H. Sesquiterpenoids from Scorzonera hispanica L. Pharmazie 2000, 55, 550–551. [Google Scholar] [PubMed]
- Auzi, A.R.; Hawisa, N.T.; Sherif, F.M.; Sarker, S.D. Neuropharmacological properties of Launaea resedifolia. Rev. Bras. Farmacogn. 2007, 17, 160–165. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, Q.; Hu, P.; Wu, W. Biguaiascorzolides A and B: Two novel dimeric guaianolides with a rare skeleton, from Scorzonera austriaca. Food Chem. 2009, 114, 1316–1320. [Google Scholar] [CrossRef]
- Sezik, E.; Yeşilada, E.; Tabata, M.; Honda, G.; Takaishi, Y.; Fujita, T.; Tanaka, T.; Takeda, Y. Traditional medicine in Turkey VIII. Folk medicine in East Anatolia; Erzurum, Erzincan, Ağrı, Kars, Iğdır Provinces. Econ. Bot. 1997, 51, 195–211. [Google Scholar] [CrossRef]
- Baytop, T. Türkiye’de Bitkiler ile Tedavi (Theraphy with Medicinal Plants in Turkey); Nobel publishers: Istanbul, Turkey, 1999. [Google Scholar]
- Donia, A.M. Phytochemical and pharmacological studies on Scorzonera alexandrina Bioss. J. Saudi Chem. Soc. 2013, in press. [Google Scholar] [CrossRef]
- Bahadir, Ö.; Saltan Çitoğlu, G.; Šmejkal, K.; Dall'Acqua, S.; Özbek, H.; Cvačka, J.; Žemlička, M. Analgesic Compounds from Scorzonera latifolia (Fisch. and Mey.) DC. J. Ethnopharmacol. 2010, 131, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Küpeli Akkol, E.; Acıkara Bahadır, Ö.; Süntar, İ.; Çitoğlu Saltan, G.; Keleş, H.; Ergene, B. Enhancement of wound healing by topical application of Scorzonera species: Determination of the constituents by HPLC with new validated reverse phase method. J. Ethnopharmacol. 2011, 137, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Küpeli Akkol, E.; Acıkara Bahadır, Ö.; Süntar, İ.; Ergene, B.; Çitoğlu Saltan, G. Ethnopharmacological evaluation of some Scorzonera species: In vivo anti-inflammatory and antinociceptive effects. J. Ethnopharmacol. 2012, 140, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Süntar, İ.; Acıkara Bahadır, Ö.; Çitoğlu Saltan, G.; Keleş, H.; Ergene, B.; Küpeli Akkol, E. In vivo and In vitro Evaluation of the Therapeutic Potential of Some Scorzonera Species as Wound healing Agent. Curr. Pharm. Des. 2011, 137, 1018–1027. [Google Scholar]
- Amdekar, S.; Roy, P.; Singh, V.; Kumar, A.; Singh, R.; Sharma, P. Anti-Inflammatory Activity of Lactobacillus on Carrageenan-Induced Paw Edema in Male Wistar Rats. Int. J. Inflamm. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.R.; Jeong, C.K.; Park, K.K.; Choi, J.H.; Park, J.H.Y.; Lim, S.S.; Chung, W.Y. Anti-Inflammatory Effects of Licorice and Roasted Licorice Extracts on TPA-Induced Acute Inflammation and Collagen-Induced Arthritis in Mice. J. Biomed. Biotechnol. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Yeşilada, E.; Üstün, O.; Sezik, E.; Takaishi, Y.; Ono, Y.; Honda, G. Inhibitory effects of Turkish folk remedies on inflammatory cytokines: Interleukin-1α, interleukin-1β and tumor necrosis factor α. J. Ethnopharmacol. 1997, 58, 59–73. [Google Scholar] [CrossRef]
- Schwaiger, S.; Zeller, I.; Pölzelbauer, P.; Frotschnig, S.; Laufer, G.; Messner, B.; Pieri, V.; Stuppner, H.; Bernhard, D. Identification and pharmacological characterization of the anti-inflammatory principal of the leaves of dwarf elder (Sambucus ebulus L.). J. Ethnopharmacol. 2011, 133, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Kaileh, M.; Berghe, W.V.; Boone, E.; Essawi, T.; Haegeman, G. Screening of indigenous Palestinian medicinal plants for potential anti-inflammatory and cytotoxic activity. J. Ethnopharmacol. 2007, 113, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Siriwatanametanon, N.; Fiebich, B.L.; Efferth, T.; Prieto, J.M.; Heinrich, M. Traditionally used Thai medicinal plants: In vitro anti-inflammatory, anticancer and antioxidant activities. J. Ethnopharmacol. 2010, 130, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Borchers, A.T.; Keen, C.L.; Stern, J.S.; Gershwin, M.E. Inflammation and Native American medicine: The role of botanicals. Am J Clin Nutr 2000, 72, 339–347. [Google Scholar] [PubMed]
- Dos Santos, M.D.; Almeida, M.C.; Lopes, N.P.; Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef] [PubMed]
- Yonathan, M.; Asres, K.; Assefa, A.; Bucar, F. In vivo anti-inflammatory and antinociceptive activities of Cheilanthes farinose. J. Ethnopharmacol. 2006, 108, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Hošek, J.; Šmejkal, K. Flavonoids as Anti-inflammatory Agents. In Encyclopedia of Inflammatory Diseases; Parnham, M., Ed.; Springer: New York, NY, USA; ISBN 978-3-0348-0620-6. (Online).
- Comalada, M.; Ballester, I.; Bailon, E.; Sierra, S.; Xaus, J.; Galvez, J.; Sanchez de Medina, F.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mouse macrophages by naturally occurring flavonoids: Analysis of the structure-activity relationship. Biochem. Pharmacol. 2006, 72, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.T.; Gohil, V.M.; Bhutani, K.K. Modulating TNF-α signaling with natural products. Drug Discov. Today 2006, 11, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-κB pathway in the treatment of inflammation and cancer. J. Clin. Investig. 2001, 107, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.-S.; Bae, J.-S. Anti-Inflammatory Effects of Triterpenoids; Naturally Occurring and Synthetic Agents. Mini-Rev. Org. Chem. 2014, 11, 316–329. [Google Scholar] [CrossRef]
- Sultana, N.; Saify, Z.S. Naturally occurring and synthetic agents as potential anti-inflammatory and immunomodulants. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2012, 11, 3–19. [Google Scholar] [CrossRef]
- Shen, W.; Qi, R.; Zhang, J.; Wang, Z.; Wang, H.; Hu, C.; Zhao, Y.; Bie, M.; Wang, Y.; Fu, Y.; Chen, M.; Lu, D. Chlorogenic acid inhibits LPS-induced microglial activation and improves survival of dopaminergic neurons. Brain Res. Bull. 2012, 88, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.K.; Zhou, W.; Lee, W.; Han, M.S.; Na, M.; Bae, J.S. Anti-inflammatory effects of hyperoside in human endothelial cells and in mice. Inflammation 2015, 38, 784–799. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Mohanti, S.; Bawankule, D.U.; Khan, F.; Shanker, K. Effect of Pluchea lanceolata bioactives in LPS-induced neuroinflammation in C6 rat glial cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-J.; Tsai, Y.-C.; Hwang, T.-L.; Wang, T.-C. Thymol, Benzofuranoid, and Phenylpropanoid Derivatives: Anti-inflammatory Constituents from Eupatorium cannabinum. J. Nat. Prod. 2011, 74, 1021–1027. [Google Scholar]
- Hashimoto, T.; Tori, M.; Asakawa, Y. Three dihydroisocoumarine glucosides from Hydrangea macrophylla subsp. Serrate. Phytochemistry 1987, 26, 3323–3330. [Google Scholar] [CrossRef]
- Frank, J.H.; Powder-George, Y.M.; Ramsewak, R.S.; Reynolds, W.F. Variable-Temperature 1H-NMR Studies on Two C-Glycosylflavones. Molecules 2012, 17, 7914–7926. [Google Scholar] [CrossRef] [PubMed]
- Pukalskas, A.; Venskutonis, P.R.; Dijkgraaf, I.; van Beek, T.A. Isolation, identification and activity of natural antioxidants from costmary (Chrysanthemum balsamita) cultivated in Lithuania. Food Chem. 2010, 122, 804–811. [Google Scholar] [CrossRef]
- Acikara Bahadir, Ö.; Saltan Citoglu, G.; Dall'Acqua, S.; Özbek, H.; Cvačka, J.; Žemlička, M.; Šmejkal, K. Bioassay-guided isolation of the antinociceptive compounds motiol and β-sitosterol from Scorzonera latifolia root extract. Pharmazie 2014, 69, 711–714. [Google Scholar]
- Paraschos, S.; Magiatis, P.; Kalpoutzakis, E.; Harvala, C.; Skaltsounis, A.-L. Three New Dihydroisocoumarins from the Greek Endemic Species Scorzonera cretica. J. Nat. Prod. 2001, 64, 1585–1587. [Google Scholar] [CrossRef] [PubMed]
- Acikara Bahadir, Ö.; Saltan Citoglu, G.; Dall’Acqua, S.; Šmejkal, K.; Cvačka, J.; Žemlička, M. A new triterpene from Scorzonera latifolia (Fisch. and Mey.) DC. Nat. Prod. Res. 2012, 26, 1892–1897. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds 1–16 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahadır Acikara, Ö.; Hošek, J.; Babula, P.; Cvačka, J.; Budešínský, M.; Dračinský, M.; Saltan İşcan, G.; Kadlecová, D.; Ballová, L.; Šmejkal, K. Turkish Scorzonera Species Extracts Attenuate Cytokine Secretion via Inhibition of NF-κB Activation, Showing Anti-Inflammatory Effect in Vitro. Molecules 2016, 21, 43. https://doi.org/10.3390/molecules21010043
Bahadır Acikara Ö, Hošek J, Babula P, Cvačka J, Budešínský M, Dračinský M, Saltan İşcan G, Kadlecová D, Ballová L, Šmejkal K. Turkish Scorzonera Species Extracts Attenuate Cytokine Secretion via Inhibition of NF-κB Activation, Showing Anti-Inflammatory Effect in Vitro. Molecules. 2016; 21(1):43. https://doi.org/10.3390/molecules21010043
Chicago/Turabian StyleBahadır Acikara, Özlem, Jan Hošek, Petr Babula, Josef Cvačka, Miloš Budešínský, Martin Dračinský, Gülçin Saltan İşcan, Daniela Kadlecová, Ludmila Ballová, and Karel Šmejkal. 2016. "Turkish Scorzonera Species Extracts Attenuate Cytokine Secretion via Inhibition of NF-κB Activation, Showing Anti-Inflammatory Effect in Vitro" Molecules 21, no. 1: 43. https://doi.org/10.3390/molecules21010043
APA StyleBahadır Acikara, Ö., Hošek, J., Babula, P., Cvačka, J., Budešínský, M., Dračinský, M., Saltan İşcan, G., Kadlecová, D., Ballová, L., & Šmejkal, K. (2016). Turkish Scorzonera Species Extracts Attenuate Cytokine Secretion via Inhibition of NF-κB Activation, Showing Anti-Inflammatory Effect in Vitro. Molecules, 21(1), 43. https://doi.org/10.3390/molecules21010043