1. Introduction
Astragalus Radix (RA) is a common Chinese medicinal material that has been used for more than 2000 years, with the first record appearing in “Shennong Bencao Jing” [
1]. RA has beneficial effects on the spleen and in treating diseases of deficiency of qi and blood [
2]. The Chinese Pharmacopoeia (2010 edition) specifies that a certified RA product must contain dried root of
Astragalus membranaceus (Fisch.) Bunge var.
mongholicus (Bunge) Hsiao (MG) or
A. membranaceus (Fisch.) Bunge (MJ) [
3].
At present, two kinds of RA are commercially available, namely, traditional RA with natural growth periods of more than five years, and fast-growing cultured RA with growth periods lasting one to three years. Traditional natural RA is mainly distributed in Shanxi, Inner Mongolia, Gansu, and Shaanxi; the production of MG (5–8 years old) from Shanxi and Inner Mongolia are considered genuine medicinal materials, which are characterized by “thick and long roots, fewer branches, mealiness, strong bean flavor, and sufficient sweetness” [
4]. Cultured RA of MG grown for two to three years is mainly distributed in Gansu and Ningxia, and cultured RA of MJ grown for one year is mainly distributed in Hebei and Shandong; these RAs are characterized by “short roots, more branches, hard texture, and tastelessness”, compared with the traditional natural RA [
5,
6,
7].
As the standard to evaluate the quality of the RA, the Chinese Pharmacopoeia (2010 edition) specifies that the content of Astragaloside ІV shall not be less than 0.04%. By determining the content of Astragaloside IV of 59 batches from different growth patterns, growth years, and commercial grades of RA samples, Tu
et al. found that the content of Astragaloside IV decreases with the increase of growth years and commercial grades; in addition, cultured RA contains more Astragaloside IV than traditional natural RA [
8,
9,
10].
The above finding indicates that under the current quality evaluation index of RA, the quality of cultured RA is similar to that of natural RA. However, the international market does not accept cultured RA, and some veteran TCM physicians still use natural RA. These conflicts show that the current quality evaluation method cannot be used for accurate assessment of the quality of RA. Therefore, a more rational evaluation method is urgently needed for the development of genuine medicinal materials.
In Traditional Chinese Medicine RA is believed to be a good medicine for “invigorating qi” and effective for treating deficiency of qi and blood [
1,
2,
7]. An experimental and clinical study showed that RA can significantly increase immunity with a variety of active anti-inflammatory, anti-cancer, and anti-viral properties [
11]. Numerous studies have shown that saccharides are the main substances responsible for immunomodulation, and different saccharides have different immunomodulatory activities [
12,
13]. Therefore, the content and variety of saccharides can be used as an index of quality evaluation for RA. At present, UV spectrophotometry is used to determine the total saccharide content for quality evaluation of RA. However, this method lacks specificity for RA, which limits its application in the Chinese Pharmacopoeia.
The successful establishment of peptide mapping technology using polypeptide hydrolase with different specificities has revolutionized the analysis of a biological system at protein level [
14,
15]. Similar to peptide mapping, saccharide mapping is developed based on saccharide hydrolysis [
16,
17,
18,
19]. Saccharide components in the cell are sequentially extracted according to the differences in the solubility of saccharide compounds in different solvents. Coupled with multivariate analysis, saccharide mapping of different medicinal materials from enzymatic hydrolysis or acid hydrolysis of sugar compounds extracts are utilized to reflect the specificity of herbs. The carbohydrate hydrolysis technology is the key factor of this technique. The specificity of the enzymatic hydrolysis method is prominent and can be used for identifying the varieties and linkages of saccharide [
16,
17]. However, more than 10 kinds of hydrolases are used in this method because of the complexity of sugar structure, thereby increasing its cost and complexity by employing different hydrolysis conditions. The acid hydrolysis method is mainly based on trifluoroacetic acid (TFA) hydrolysis with gas chromatography–mass spectrometry (GC-MS) [
18]. This method not only can quantitatively analyze the variety and content of monosaccharides, but also has the advantages of simple operation, high specificity, and low cost. Guan
et al. found differences in natural and cultivated
Cordyceps sinensis by monosaccharide mapping based on acid hydrolysis [
18]. Wu
et al. distinguished the 12 kinds of different Ganoderma medicinal materials by the above method [
19]. The technology laid foundations for the use of sugar compounds as a quality index for evaluation of TCMs.
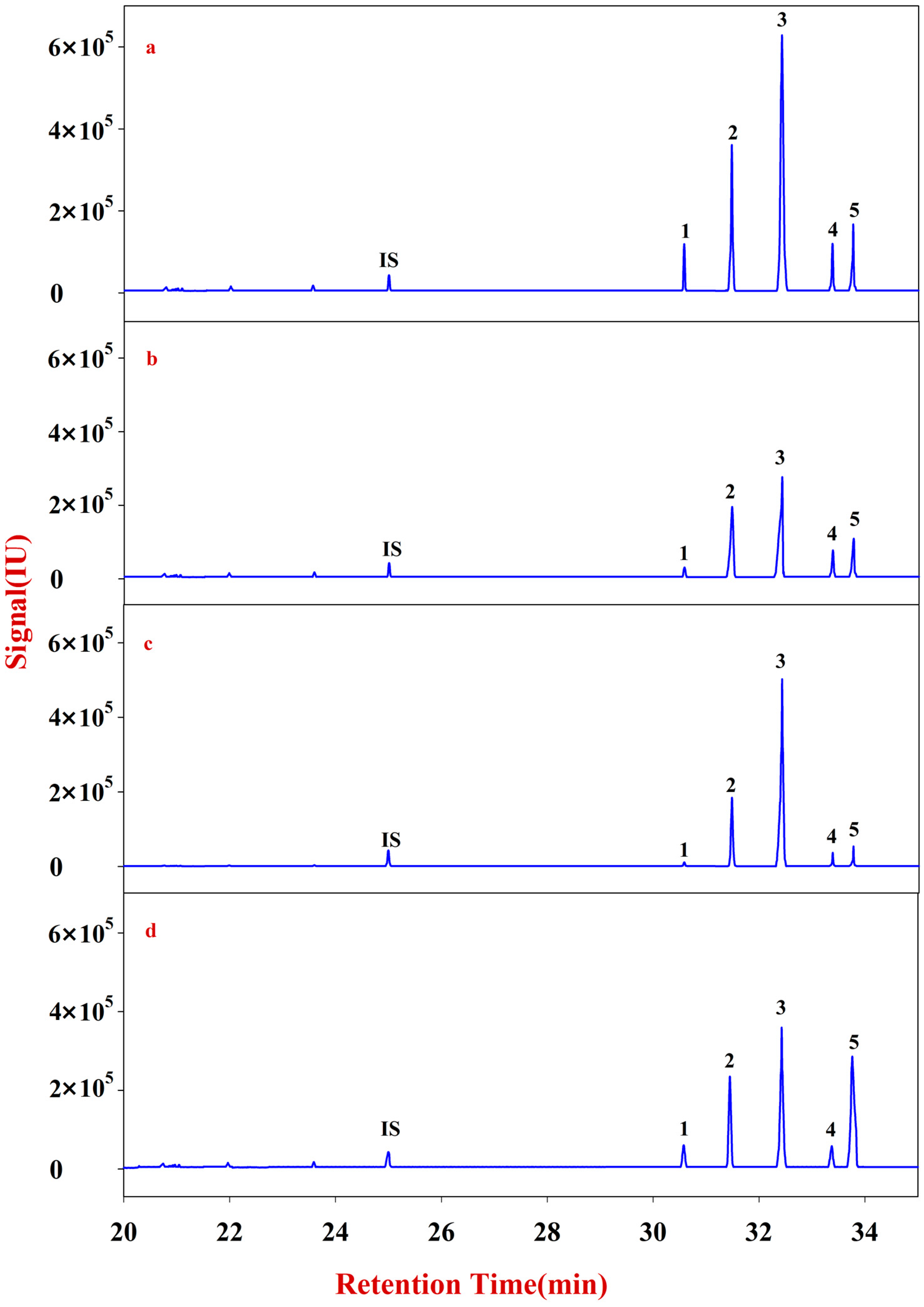
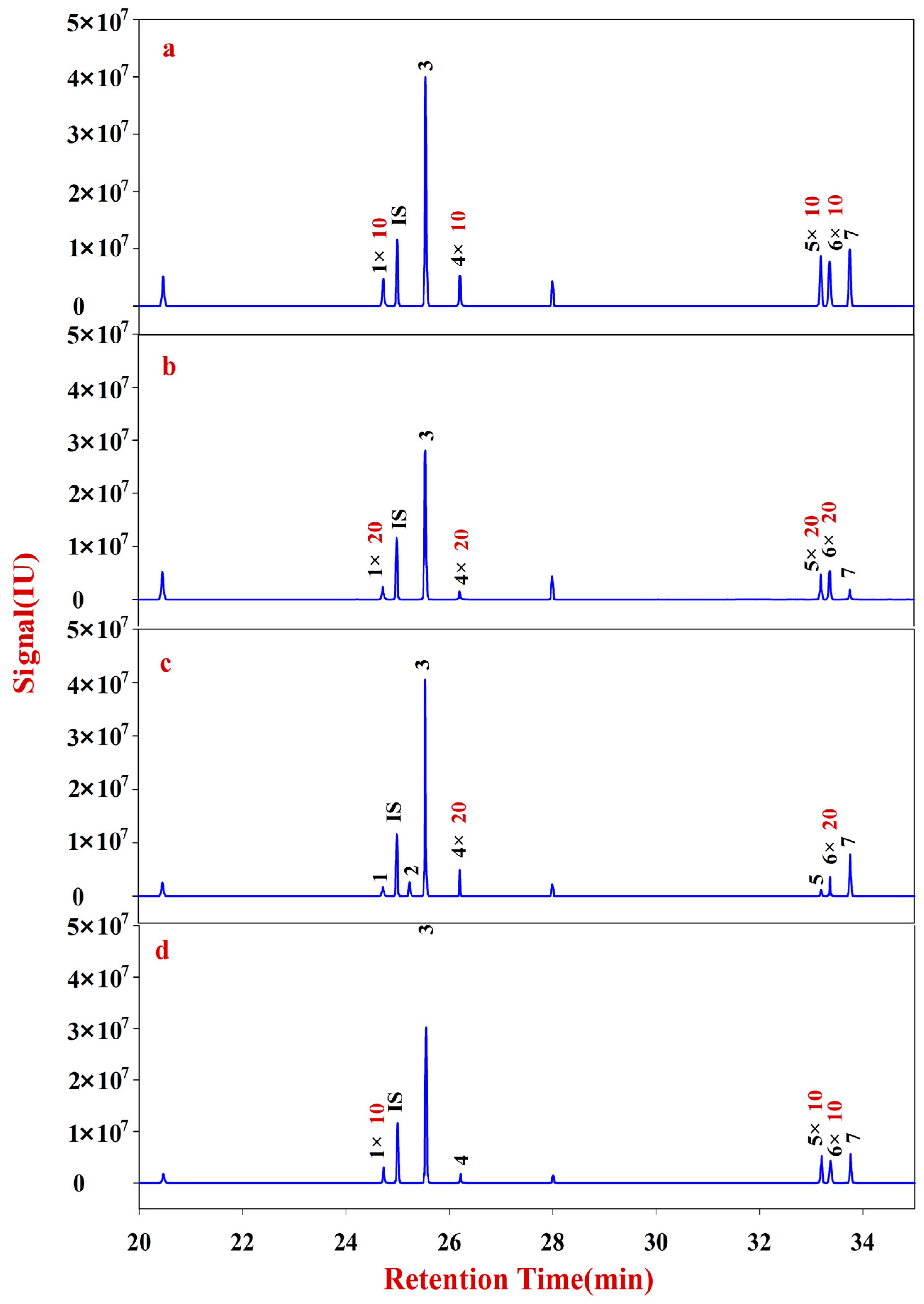
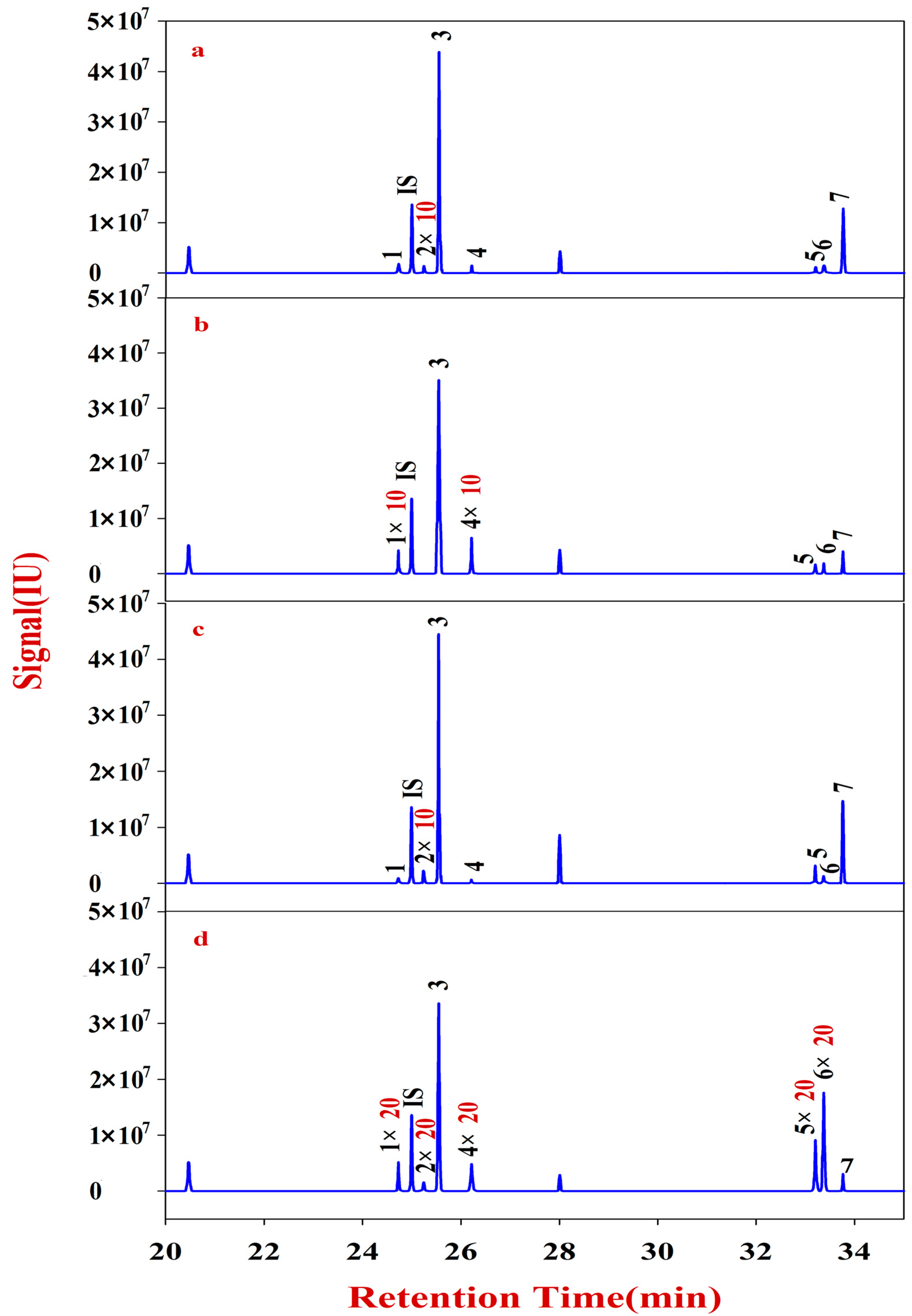
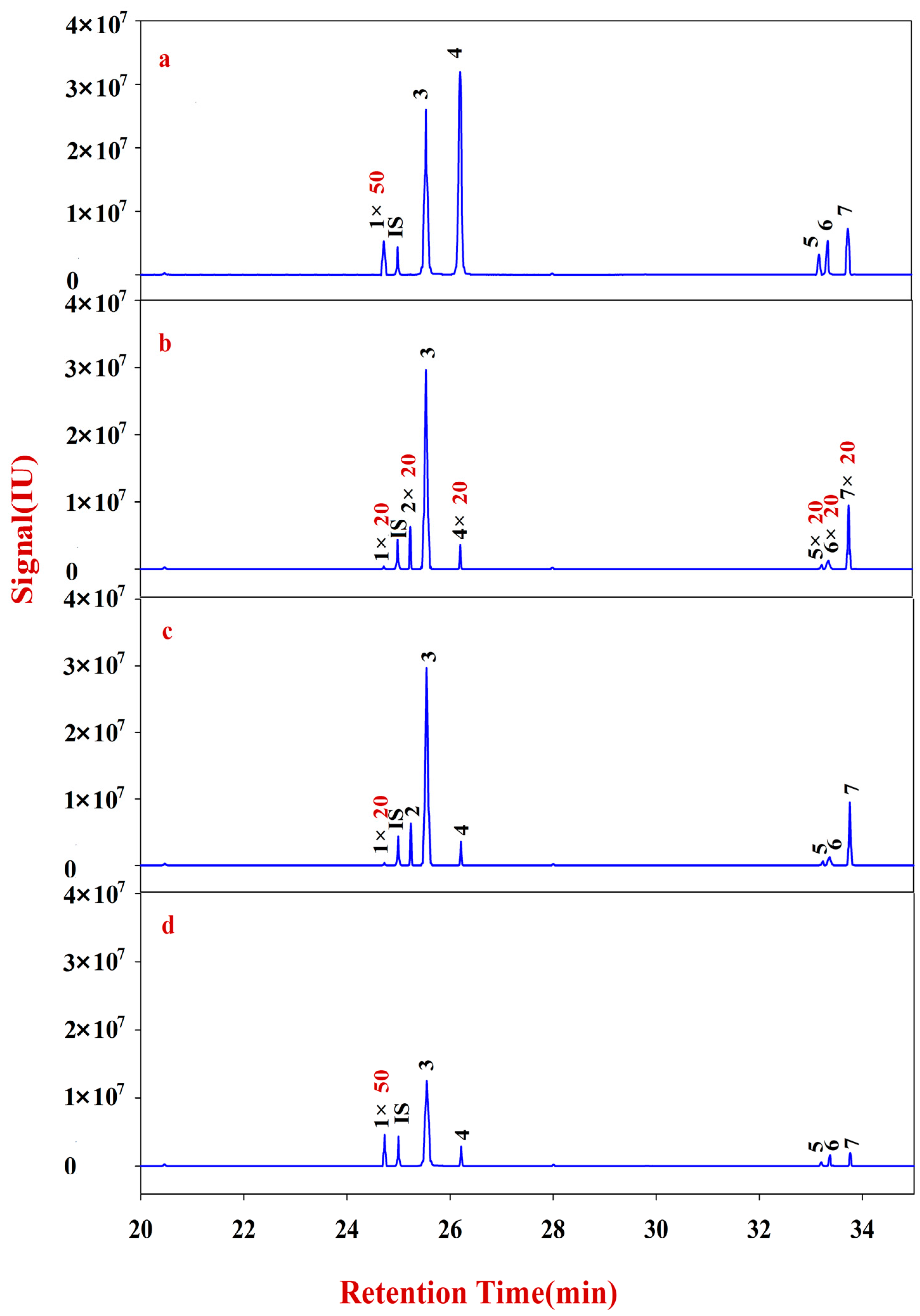
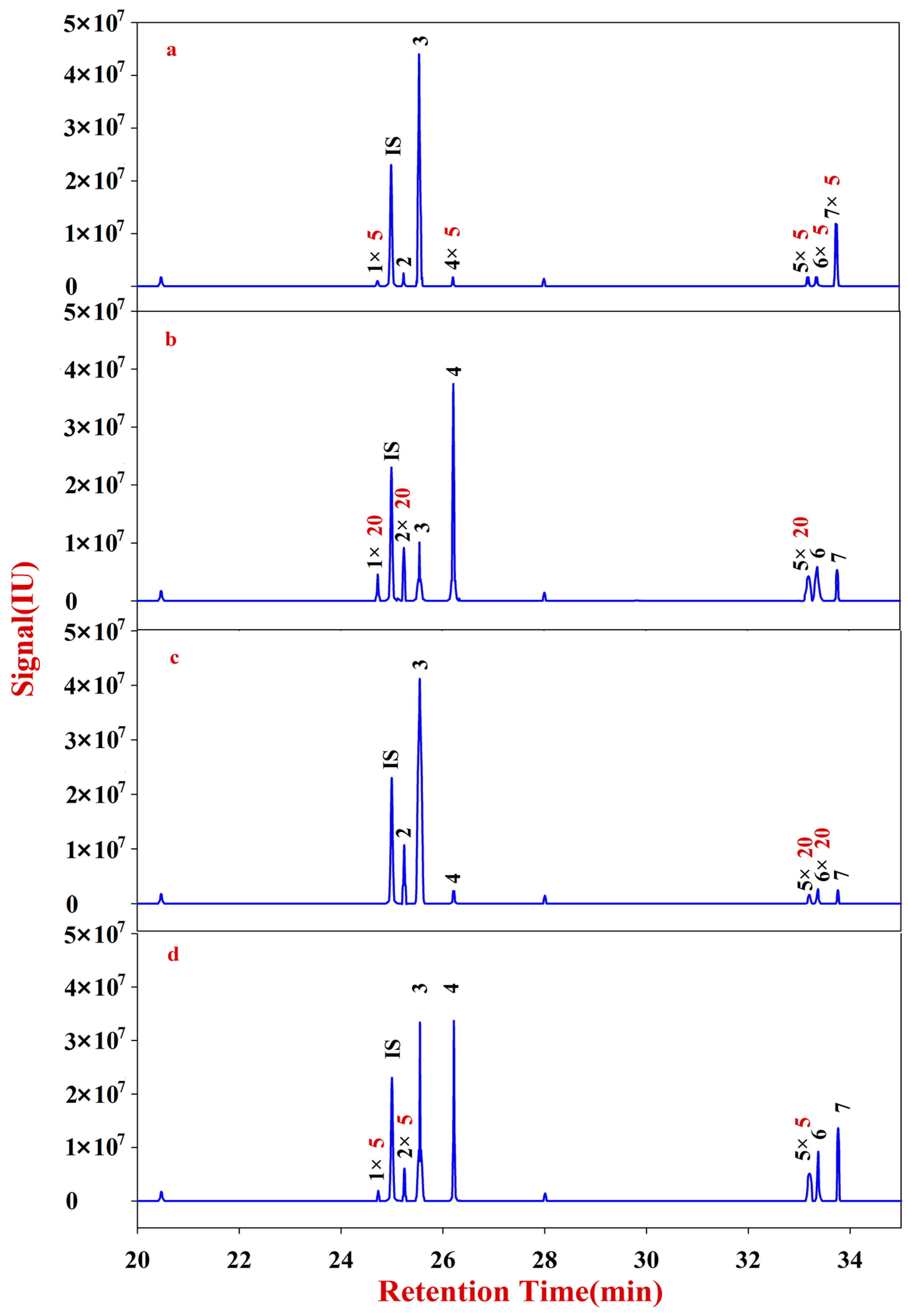
In the present study, RA samples from 24 batches with different growth patterns were selected to elucidate the corresponding monosaccharide mapping, including soluble sugar in cytoplasm, polysaccharides in pectin, and hemicelluloses of cell wall, through sequential chemical extraction, TFA hydrolysis, acetylation, and GC-MS analysis. The components and classification indexes of the natural and cultured RA were compared and analyzed by quantitative monosaccharide mapping combined with multivariate and correlation analyses.
3. Materials and Methods
3.1. Plant Materials
RA samples were obtained from two species of
A. membranaceus. All plant materials were identified by Prof. Xue-Mei Qin, and the voucher specimens were deposited in the herbarium of the Modern Research Center for Traditional Chinese Medicine of Shanxi University. Detailed information on the samples is presented in
Table 8. The 24 RA samples were dried, pulverized, and then stored at 20 °C in a vacuum desiccator until further analysis.
Table 8.
List of RA plant materials.
Table 8.
List of RA plant materials.
| Species | Cultivation Pattern | Regions | No. | Harvesting Time | Growth Year |
|---|
| MG | Cultured RA | Gs, Longxi County | 1–2 | 2010.12 | 2 |
| Gs, Dangchang County | 3 | 2010.12 | 2 |
| Sx, Daixian County | 4 | 2011.1 | 2 |
| Sx, Yingxian County | 5 | 2011.1 | 2 |
| Sx, Yanggao County | 6 | 2011.1 | 2 |
| Natural RA | Ssx, Yulin | 7–9 | 2011 | 5 |
| Sx, Hunyuan County | 10 | 2011.11 | ≥5 |
| 11–12 | 2011.1 | 5 |
| MJ | Cultured RA | Hlj, Hulan County | 13–15 | 2011.1 | 1 |
| Sd, Wendeng County | 16–18 | 2011 | 1 |
| Natural RA | Hlj, Jiagedaqi | 19–20 | 2011 | 5 |
| Hlj, Hulan County | 21 | 2011.11 | 5 |
| Hlj | 22–24 | 2011 | 5 |
3.2. Solvents and Chemicals
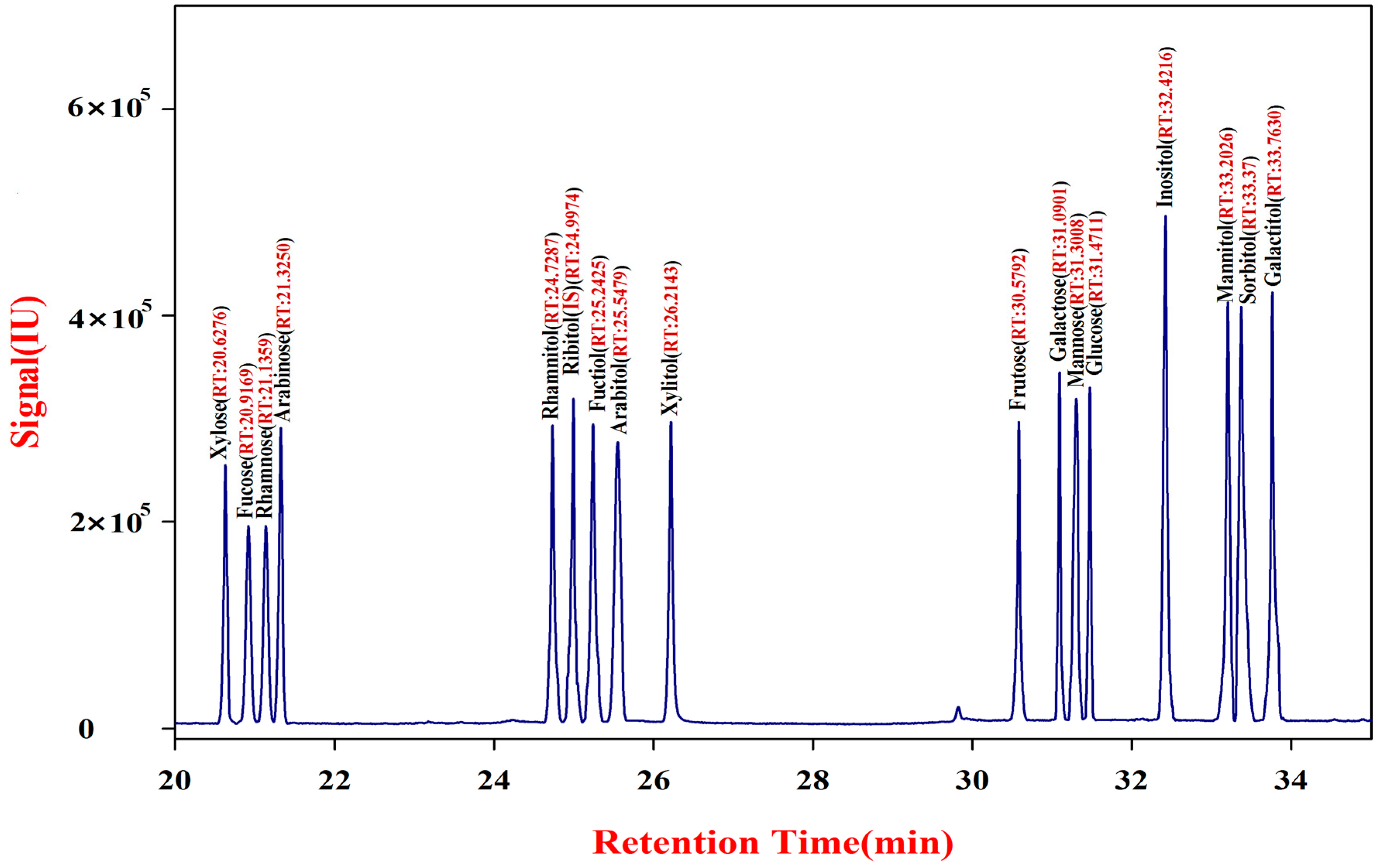
Analytical-grade Na2CO3, KOH, acetic anhydride, NaBH4, and tris-(hydroxymethyl) methyl aminomethane were purchased from Fengchuan Chemical Co. Ltd. (Tianjin, China). Analytical-grade ethanol acetic anhydride, CH2Cl2, and acetone were purchased from Beijing Chemical Works (Beijing, China). TFA, dimethyl sulfoxide (DMSO), and 1-methylimidazole were obtained from Sangon Biotech Co., Ltd. (Shanghai, China). The 17 standard compounds of monosaccharides and alditols (d-glucose, d-galactose, d-mannose, d-fructose, d-xylose, d-arabinose, l-fucose, l-rhamnose, l-inositol, d-ribitol, sorbitol, galactitol, mannitol, xylitol, arabinitol, fucitol, and rhamnitol), were purchased from Sigma-Aldrich China (Shanghai, China). Pectinase (>500 µ/mg) from Aspergillus was purchased from Sangon (Shanghai, China). Type-I porcine a-amylase (250 µ/mg) was obtained from Sigma-Aldrich China.
3.3. Preparation of 17 Standard Monosaccharide and Alditol Acetylation Products and Methodology Validation
Derivatization of authentic standard compounds of soluble saccharides was done following the previously described method [
32]. This method was concurrently used to generate GC-MS chromatograms of the 17 standard compounds of monosaccharides and alditols.
Authentic standard saccharide solution of ribitol (IS)-DMSO (1:10 w/v; 200 µL) was mixed with 150 μL of acetic anhydride and 30 μL of 1-methylimidazole into 10 mL glass centrifuge tubes and then left to stand for 10 min. Double-distilled H2O (ddH2O, 600 µL) was added to the tubes to remove excess acetic anhydride. Then, 200 μL of CH2Cl2 was added to the tubes. The tubes were centrifuged for 1 min to partition the organic phase. Finally, 1 μL of the lower methylene chloride fraction was injected for GC-MS analysis.
A series of standard solutions of ribitol in DMSO at seven levels including 0.5, 2.5, 6.25, 25, 50, 100 and 200 ng/μL was prepared for determining the standard equation. These solutions were derivatized as described in the manuscript and three replications of each level were performed. According to the following formula, the standard equation of internal standard ribitol and the relative correction factor for each standard compound was calculated using ribitol as an internal standard:
where
mi and
ms respectively stand for the detection masses of monosaccharide and inner standard of ribitol;
Ai and
As respectively stand for the peak areas of monosaccharide and inner standard of ribitol; f stands for the relative correction factor of monosaccharide to ribitol. We calculated the standard equation of ribitol, detection limit, and the recovery rate of each standard compound.
3.4. GC-MS Detection Conditions
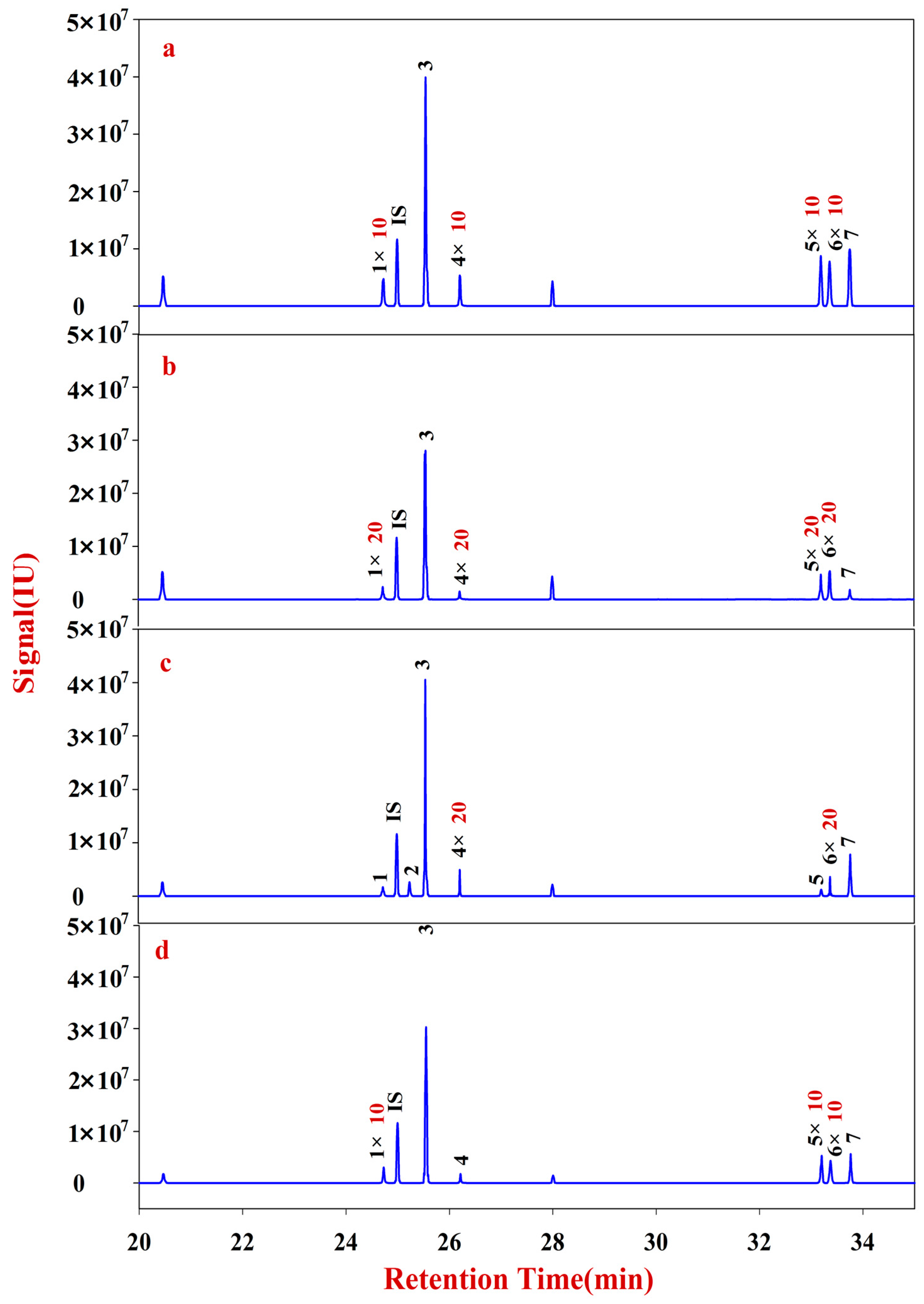
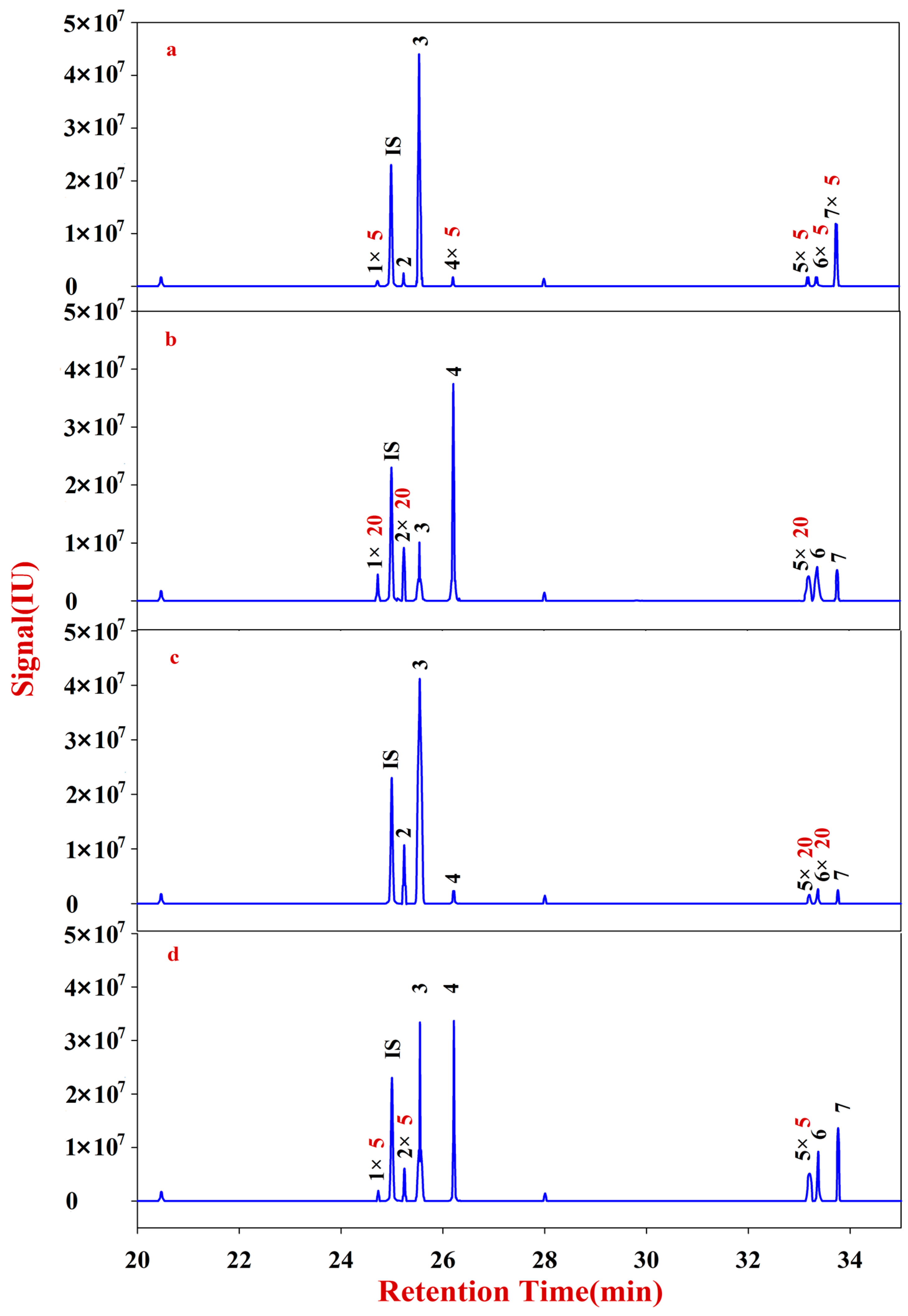
An ion trap MS system (Trace-GC Ultra) connected to a Trace-Plase Q Mass Selective Detector (Thermo Finnigan, San Jose, CA, USA) was used in the analysis of acetyl derivatives on a DB-5 MS fused silica capillary column (30 m × 0.25 mm, 0.25 μm film thickness, Agilent, Santa Clara, California, CA, USA) and with ionization of full scan and selected-ion monitoring (selected ion ± 1.0 mass unit). The oven temperatures were programmed as follows: initiation at 100 °C, gradual ramped to 180 °C (5 °C/min), and held for 1 min; 190 °C (1 °C/min) and held for 2 min; then increased to 220 °C (30 °C/min), holding for 2 min; then increased to 230 °C (1 °C/min), holding for 2 min; ramped to 280 °C (20 °C/min), and held for 10 min. The temperature at the injector port was 250 °C, and that at the detector port was 220 °C. Helium was used as the carrier gas at a constant flow rate of 1.0 mL/min. An Xcalibur 2.0 workstation was used for data acquisition and quantitative data processing.
3.5. Sequential Extractive Process of Fractions A and B of RA
The saccharide components in the cytoplasm of RA preparation were determined as previously described [
18]. Briefly, 20 g of RA fine powder was placed into a 500 mL round-bottom flask, to which 200 mL of 70% ethanol aqueous solution was added for extraction. The solution was extracted at 100 °C for 1 h, cooled to room temperature, and then centrifuged. The supernatant was freeze-dried to obtain a powder of Fraction A. The residual powder was placed into a 500 mL round-bottom flask, to which ddH
2O (1:80,
w/
v) was added. The solution was extracted at 100 °C for 3 h and then centrifuged. The extraction process was repeated, in which two times supernatant were combined, concentrated, and freeze-dried to obtain a powder Fraction B.
3.6. Alcohol Insoluble Residue (AIR) Preparation
AIR of RA was prepared as previously described [
33]. The precipitate was placed into a 500 mL conical flask, to which 80% ethanol aqueous solution (1:100,
w/
v) was added. The reaction was placed in a water bath at 60 °C for 1 h, and then centrifuged at 3500 rpm for 10 min, the supernatant was discarded, and the above experiment was repeated several times until the supernatant was colorless. Acetone (1:10,
w/
v) was added to the conical flask. The solution was allowed to settle for some time, then the supernatant was discarded, and the residue dried in an oven at 45 °C. The residual powder was placed into a 500 mL conical flask, to which a buffer (1:100,
w/
v; 50 mM Tris-HCl, pH 7.0) containing type-I porcine α-amylase (20 units/mL) was added. The reaction was placed in a shaking table at 37 °C for 12 h, and then centrifuged at 3500 rpm for 10 min; the supernatant was discarded. The residue was washed for several times by ddH
2O, and then centrifuged. The supernatant was discarded and acetone (1:10,
w/
v) was added to the residue. The solution was left to stand for some time and the supernatant was discarded. The residue was dried in an oven at 45 °C.
3.7. Sequential Extraction Process of Fractions C and D of RA
Portions of the Fraction C were solubilized from the cell walls of RA by each of the following sequential treatments: EPG and Na
2CO
3 as previously described [
34]. Briefly, AIR fine power material (2 g) was weighed and placed into a 100 mL centrifuge tube, to which 70 mL of buffer (sodium acetate 50 mM, pH 5.0) containing pectinase (20 μ/mL) were added. The reaction was placed in a shaking table at 30 °C for 24 h, and then centrifuged at 3500 rpm for 10 min; the supernatant was removed and stored at 4 °C. The above steps were repeated for two or three times, and then the residue was washed with 20 mL ddH
2O for three times. The supernatant and washes were combined in a 500 mL conical flask, which was placed in a water bath at 70 °C for 30 min. Then, the solution was run through a dialysis tubing (3500 molecular-weight cutoff), concentrated, and then lyophilized.
The residual walls were added with 70 mL of 50 mM Na2CO3 and 5 mM EGTA. The reaction was placed in a shaking table at 30 °C for 24 h, and then centrifuged at 3500 rpm for 10 min. The supernatant was removed and stored at 4 °C. The above steps were repeated two or three times, and then the residue was washed with 20 mL of ddH2O three times. The supernatant and washes were combined, neutralized with acetic acid, dialyzed, concentrated, and lyophilized.
After Na2CO3 extraction, the walls were treated sequentially with 1 M and 4 M KOH to solubilize Fraction D. The remaining residual walls after Na2CO3, extraction were incubated in 70 mL of 1 M KOH solution containing 1% NaBH4 at 30 °C for 24 h. Then, the wall suspension was centrifuged at 3500 rpm for 10 min, and the supernatant was removed and stored at 4 °C. The above steps were repeated two or three times, and then the residue was washed with 20 mL ddH2O three times. The supernatant and washes were combined, neutralized with acetic acid, dialyzed, concentrated and lyophilized.
The remaining residual walls after extraction with 1 M KOH were incubated in 70 mL of 4 M KOH solution containing 1% NaBH4 at 30 °C for 24 h. Then, the wall suspension was centrifuged at 3500 rpm for 10 min, and the supernatant was removed and stored at 4 °C. The above steps were repeated two or three times, and then the residue was washed with 20 mL of ddH2O three times. The supernatant and washes were combined, neutralized with acetic acid, dialyzed, concentrated, and lyophilized.
3.8. Hydrolization by TFA of Carbohydrates of RA
Acid hydrolysis of RA was determined as described previously [
34]. Fraction A powder (20 mg) was placed into a 10 mL centrifuge tube, to which 2 mL of 2 M TFA was added. The tube was sealed and placed in an oil bath at 110 °C for 1h. Then, after drying under a gentle steam of air, the sample was redissolved with 1 mL of methanol and then dried again. The experiment was repeated for three times, and 1 mL of internal standard solution ribitol (1:10,
w/
v) was added. Fractions B, C, and D powders (20 mg each) were separately placed into 10 mL centrifuge tubes. Hydrolysis products obtained by the above method were dissolved in 1 mL of ribitol (1:10,
w/
v).
3.9. Preparation of Carbohydrates of RA Acetylation Products
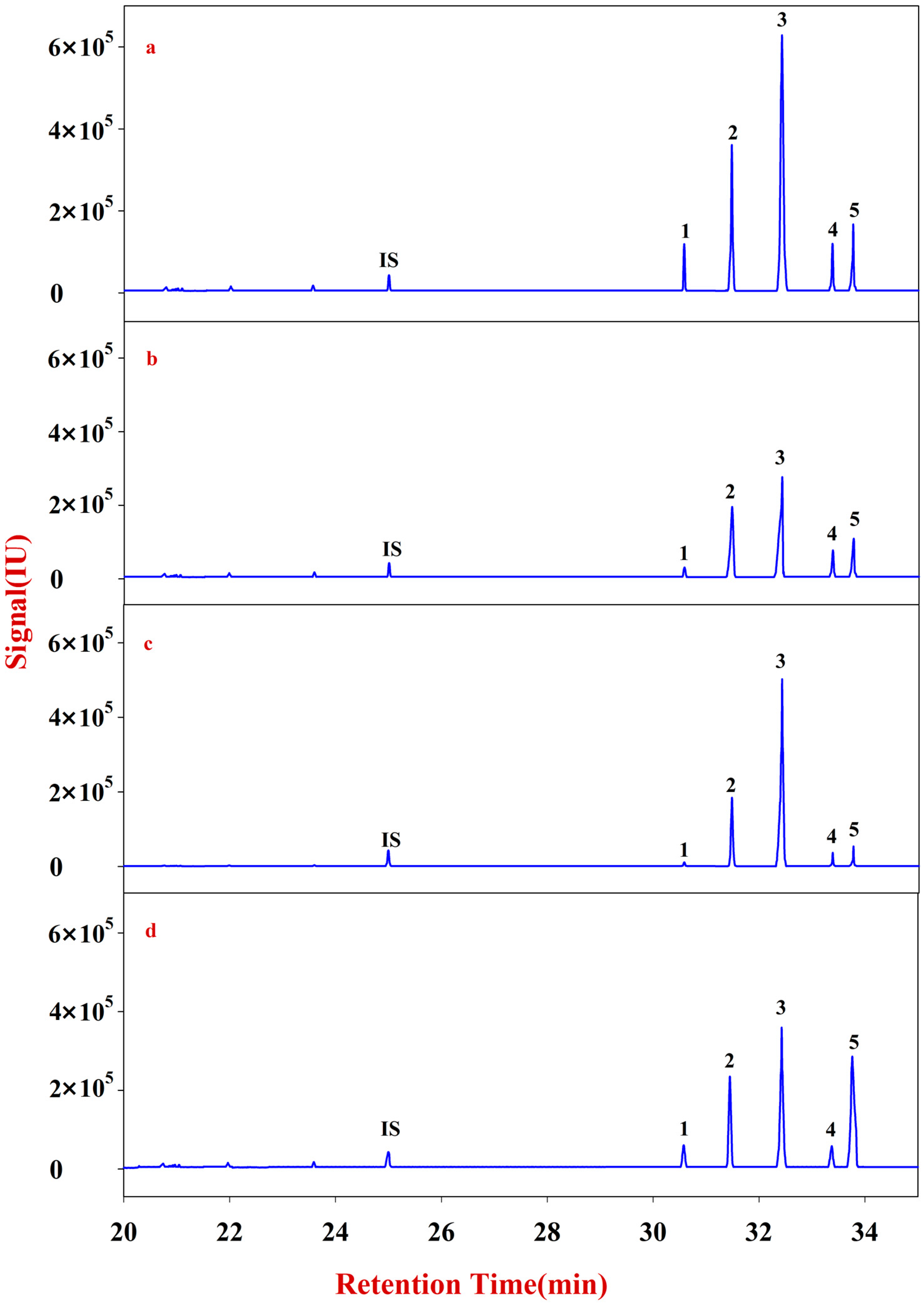
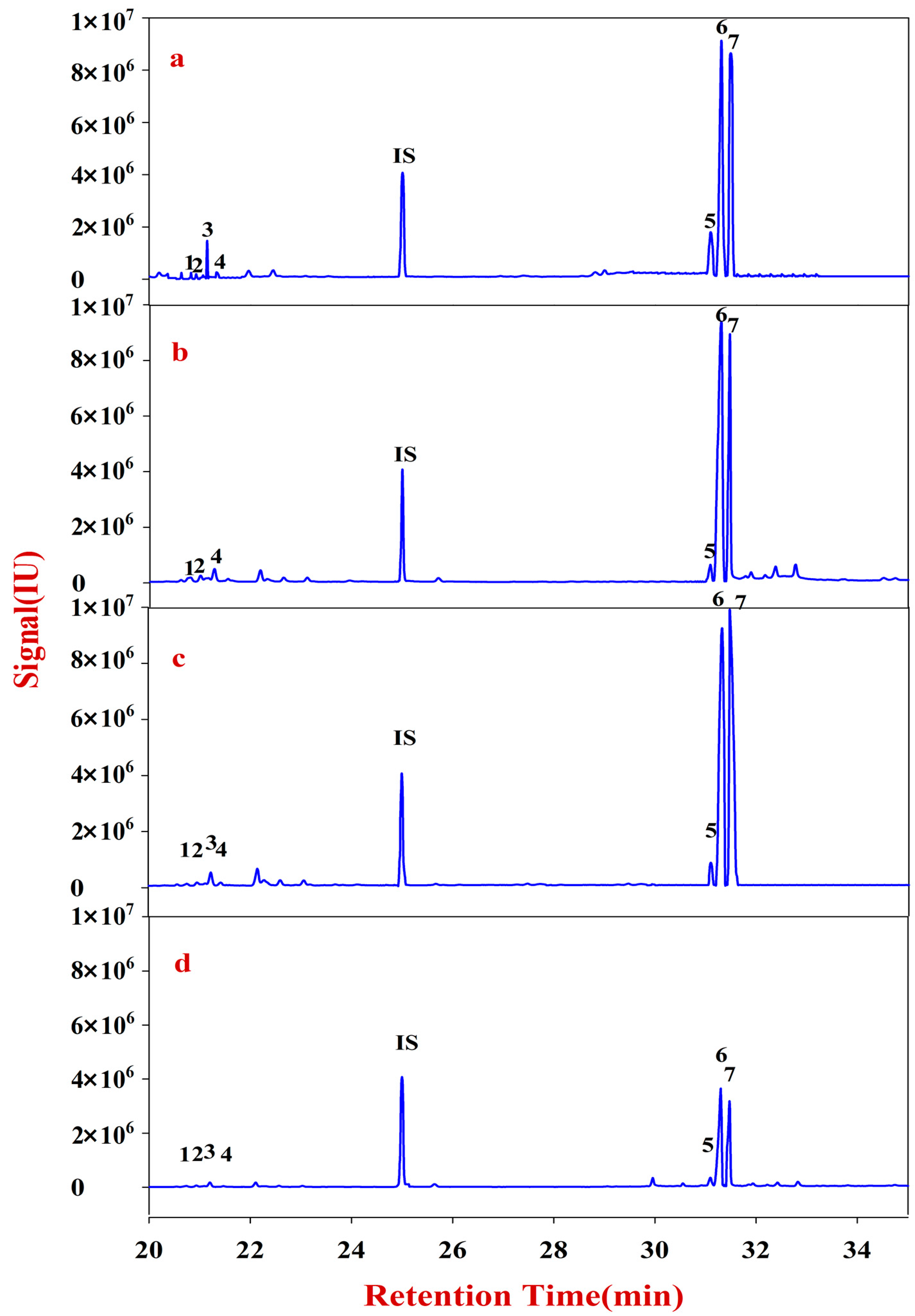
The solutions of Fractions A and B were transferred into 10 mL glass centrifuge tubes for derivatization. After drying, the samples were redissolved into 200 μL of CH2Cl2 for filtering. The reaction mixture (1 μL) was injected into the GC-MS for analysis. Fractions A and B of RA were quantitatively analyzed according to an internal standard.
The solutions of Fractions C and D were transferred into 10 mL glass centrifuge tubes, to which 500 μL of DMSO containing 2% NaHB4 was added. The reaction was placed in a shaking table at 40 °C for 90 min, neutralized with acetic acid, and then derivatized. After drying, the sample was redissolved into 200 μL of CH2Cl2 for filtering. The reaction mixture (1 μL) was injected into the GC-MS for analysis. Fractions C and D of RA were quantitatively analyzed according to an internal standard.
3.10. Statistical Analysis
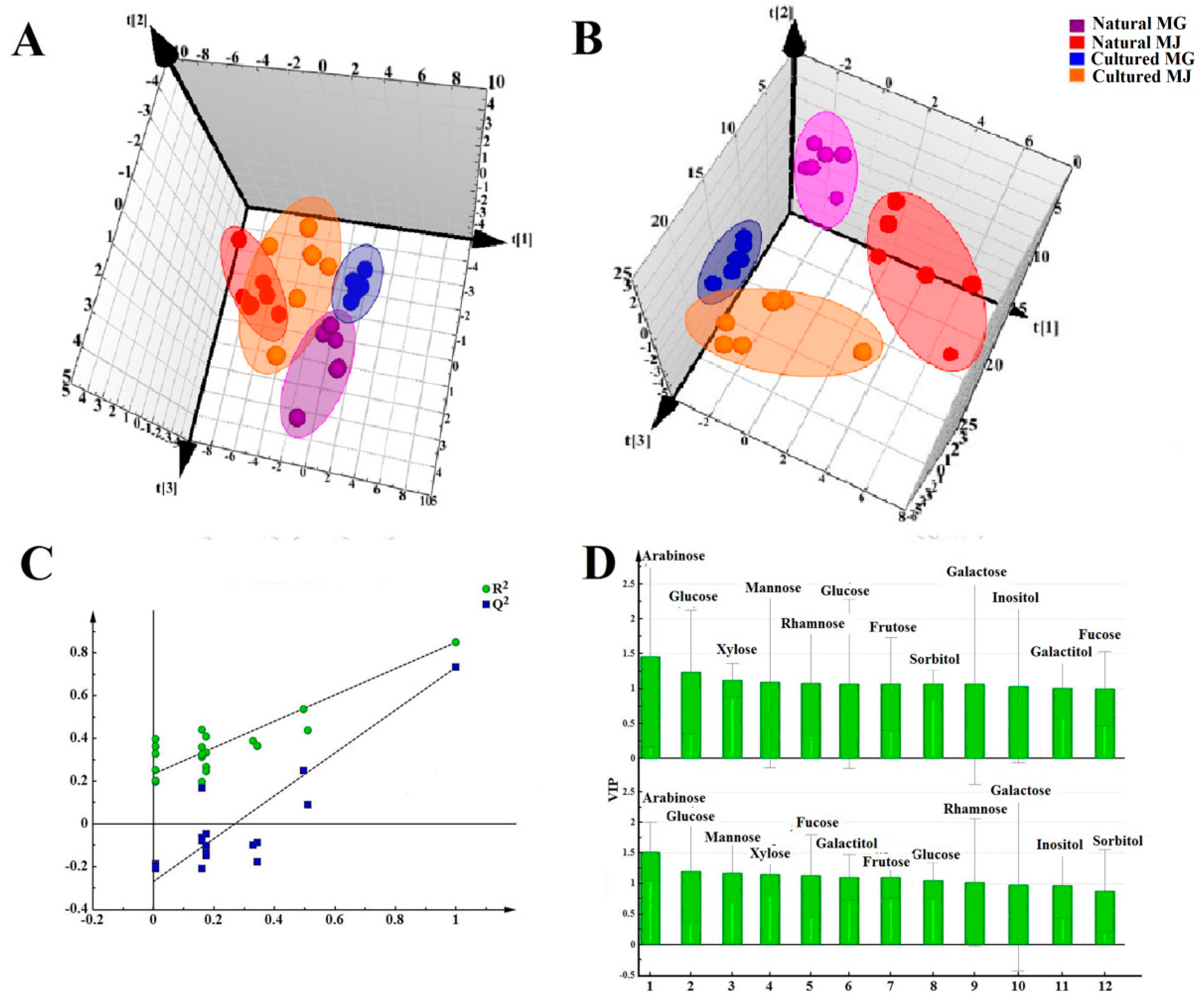
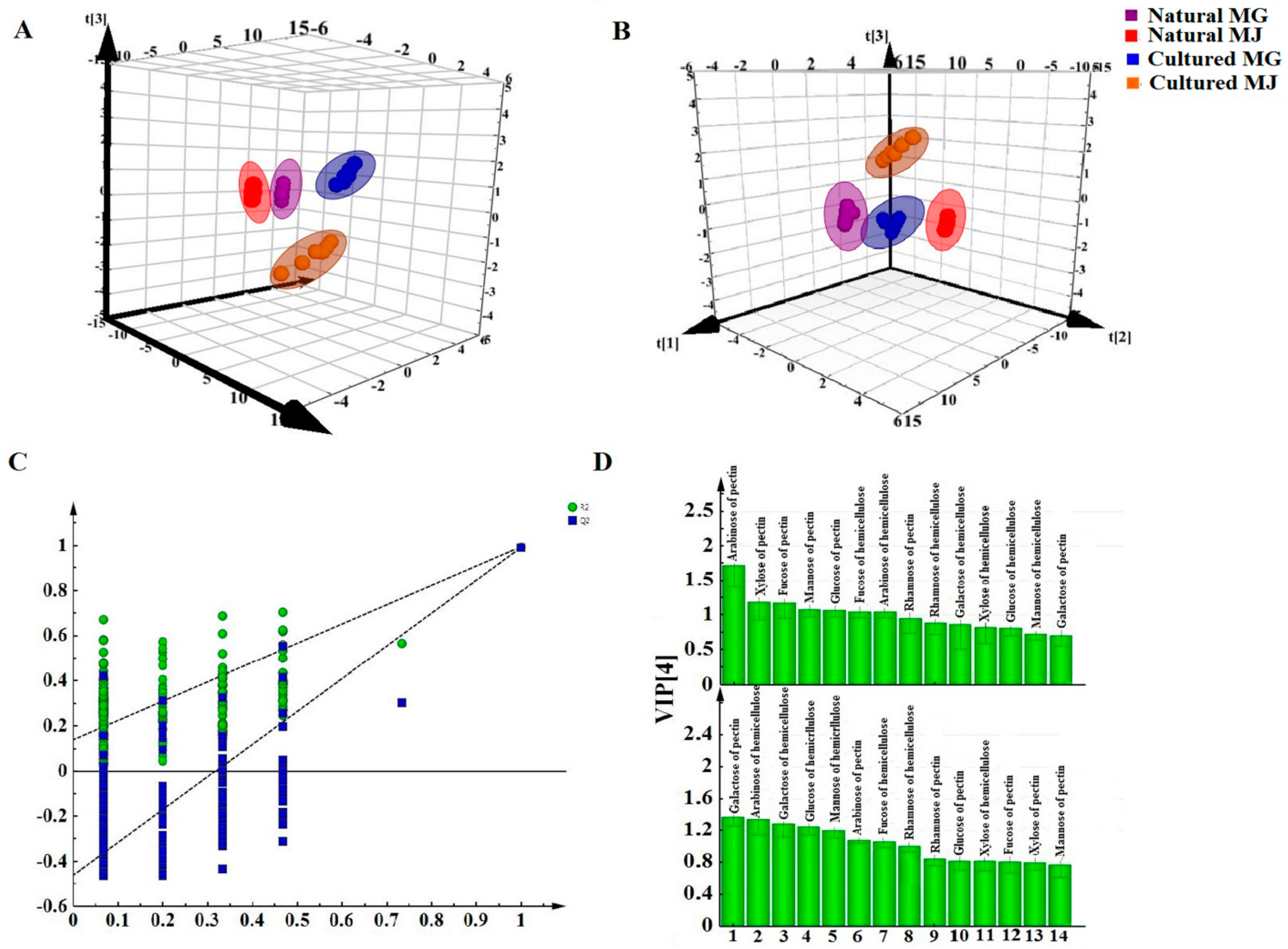
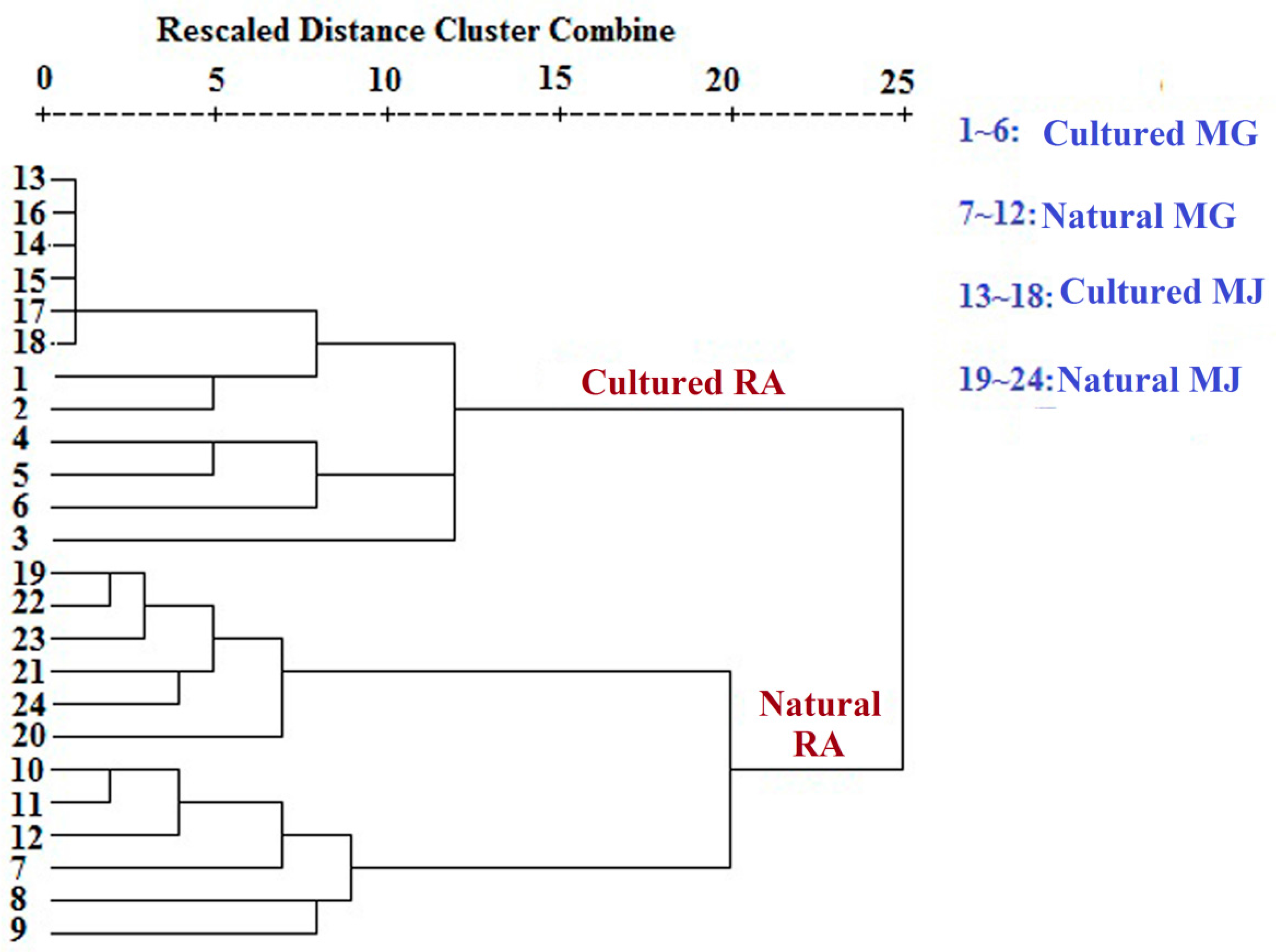
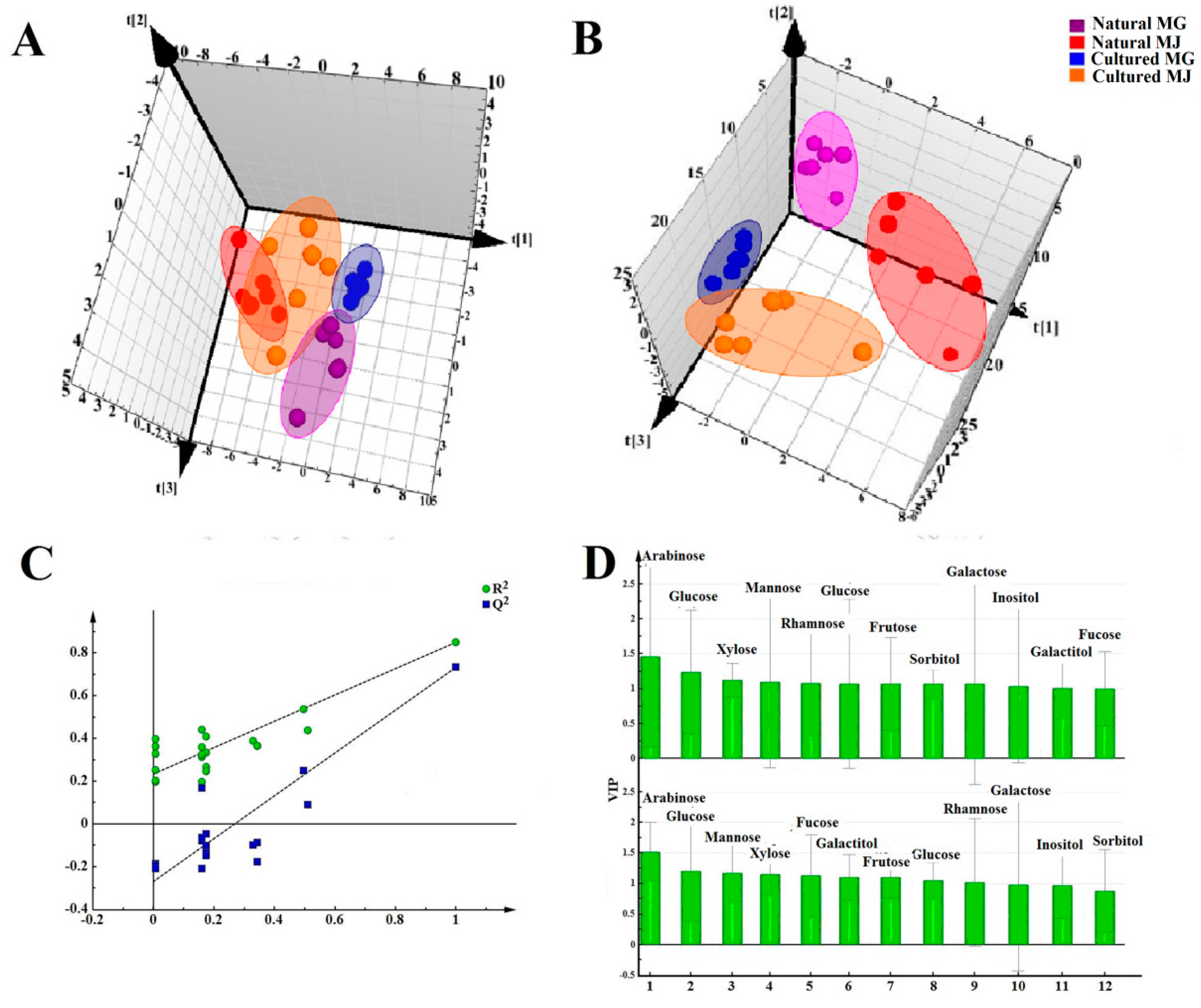
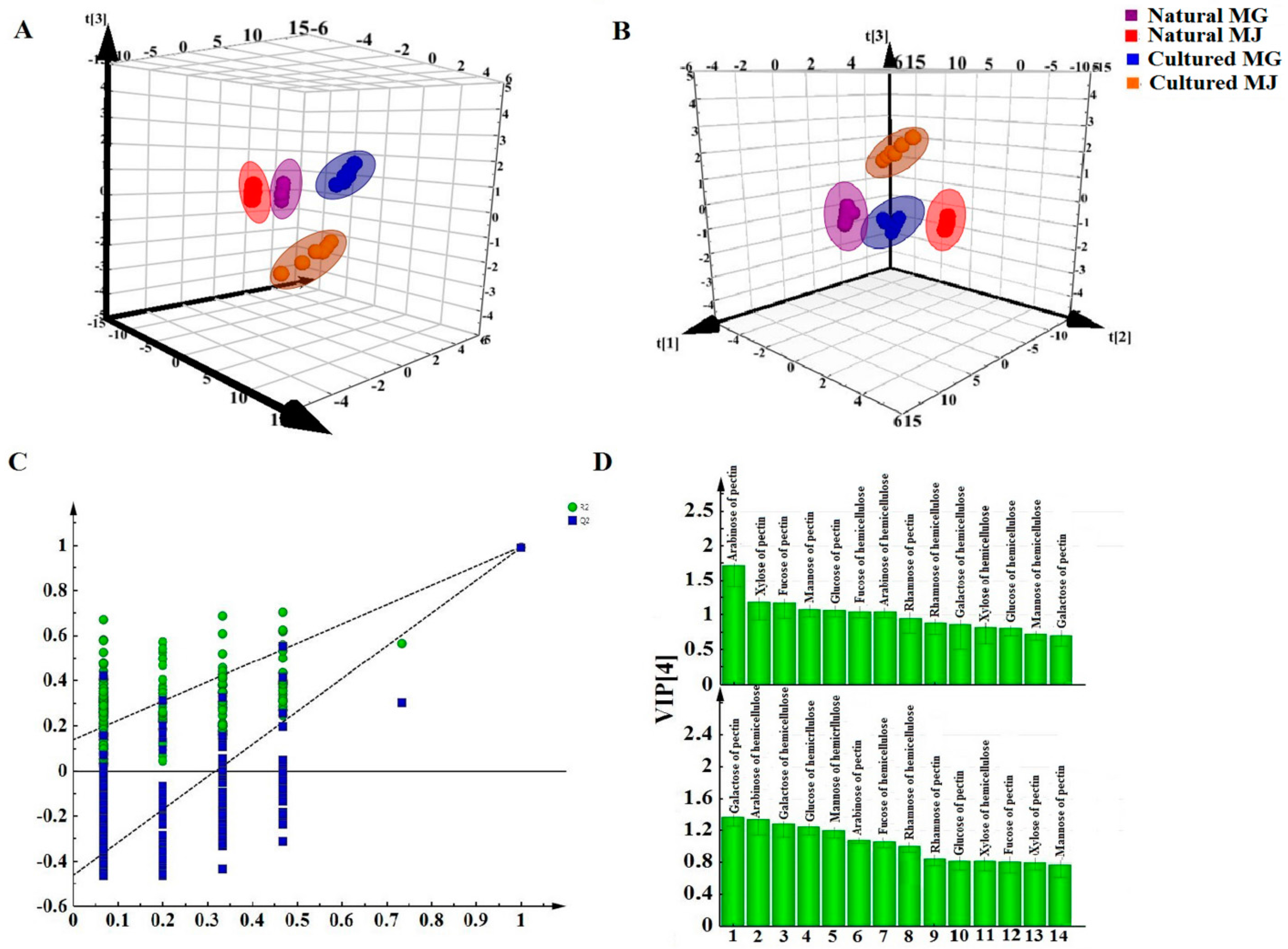
Multivariate analyses for all data were performed using SIMCA-P 13.0. Correlation analyses for all data (mean ± SD) were performed using SPSS 16.0. Monosaccharides mapping obtained by SigmaPlot 12.0 were used to establish all figures.
4. Conclusions
In this study, monosaccharide mapping of Fractions A, B, C, and D of 24 batches of RA samples with different growth patterns were obtained based on TFA hydrolysis followed by GC/MS. Fractions A, B, C, and D were mainly composed of glucose, fructose, inositol, sorbitol, galactitol, xylose, fucose, rhamnose, arabinose, galactose, and mannose. Statistical analyses showed that the contents of monosaccharides in natural RA were higher than those of cultured RA.
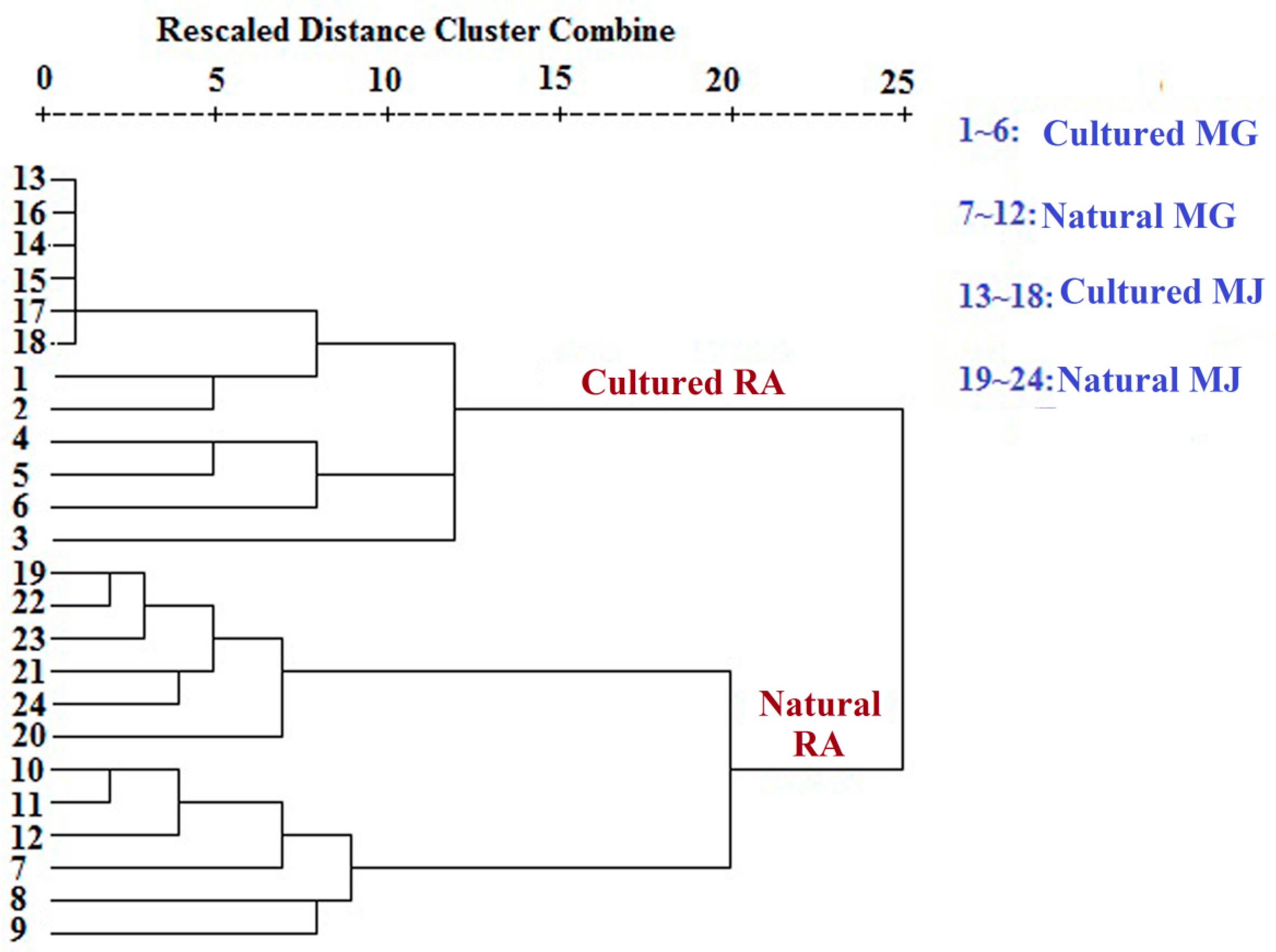
Monosaccharide mapping of RA by multivariate analysis showed that natural RA and cultured RA can be separated. Results indicated that the mapping and molar ratios of saccharide compositions of the cultured and natural RA samples were different both in cytoplasm and cell wall. The molar ratio of mannose to arabinose of Fraction A was higher than 3.5:1 in cytoplasm in cultured RA, where the ratio was less than 3.5:1 in natural RA. The ratio of the content of arabinose of Fraction D to that of Fraction C was more than 0.5:1 in polysaccharides of cell wall in cultured RA, whereas the ratio was less than 0.5:1 in natural RA. Thus, these ratios may be used to distinguish the growth patterns of RA.
The difference in monosaccharide composition and carbohydrate content between natural RA and cultured RA were compared by GC-MS. The molar ratios of different monosaccharides between natural and cultured RA were also determined. This research not only provides a basis for the identification of RA in terms of growth patterns, but also for evaluation of the quality of Chinese herbal medicines.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}