
Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice

Abstract
:1. Introduction
2. Results and Discussion
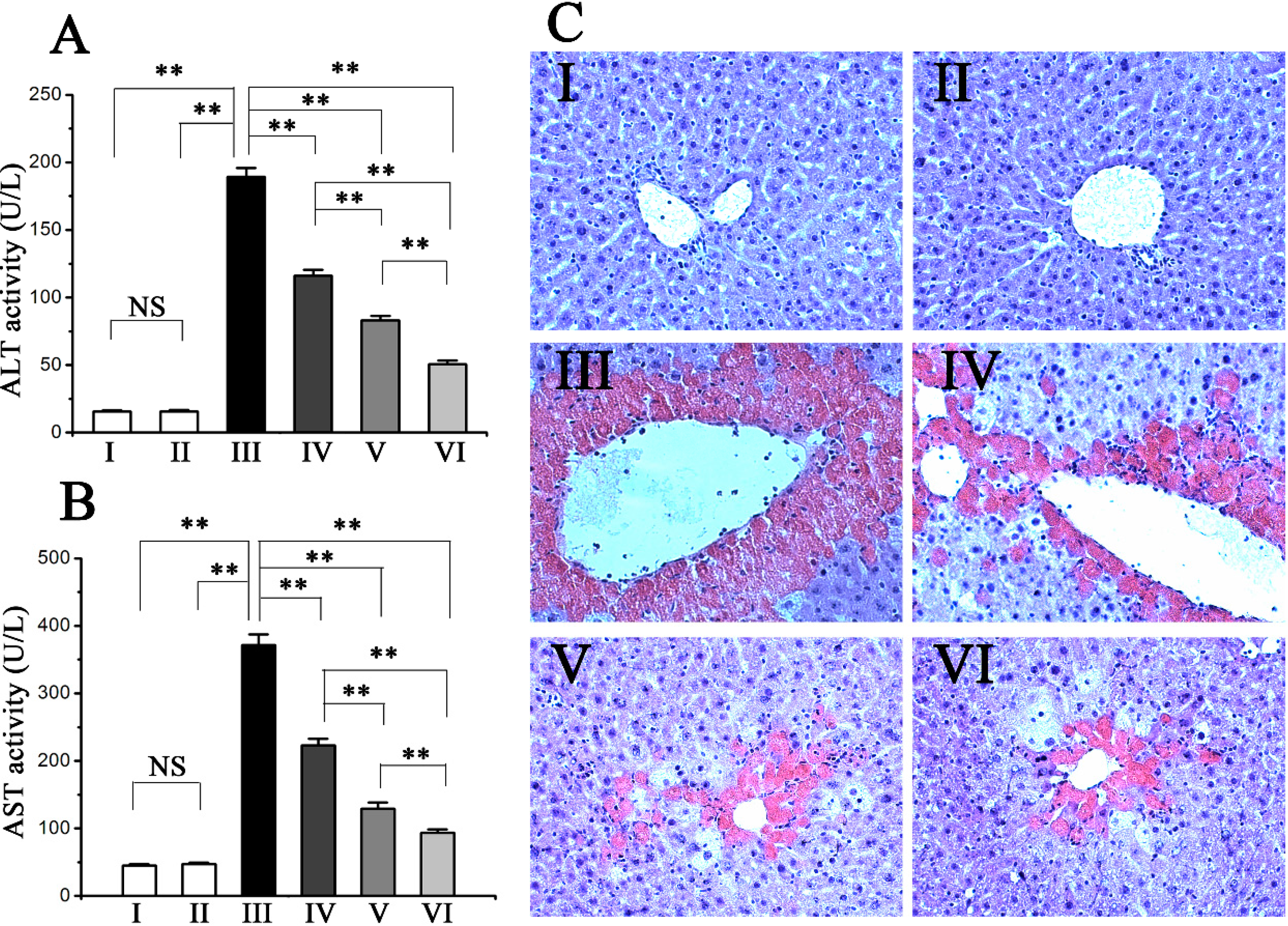
2.1. Procyanidin B2 Protects against CCl4-Induced Hepatic Dysfunction
2.2. Procyanidin B2 Alleviated CCl4-Induced Histopathological Changes in the Liver
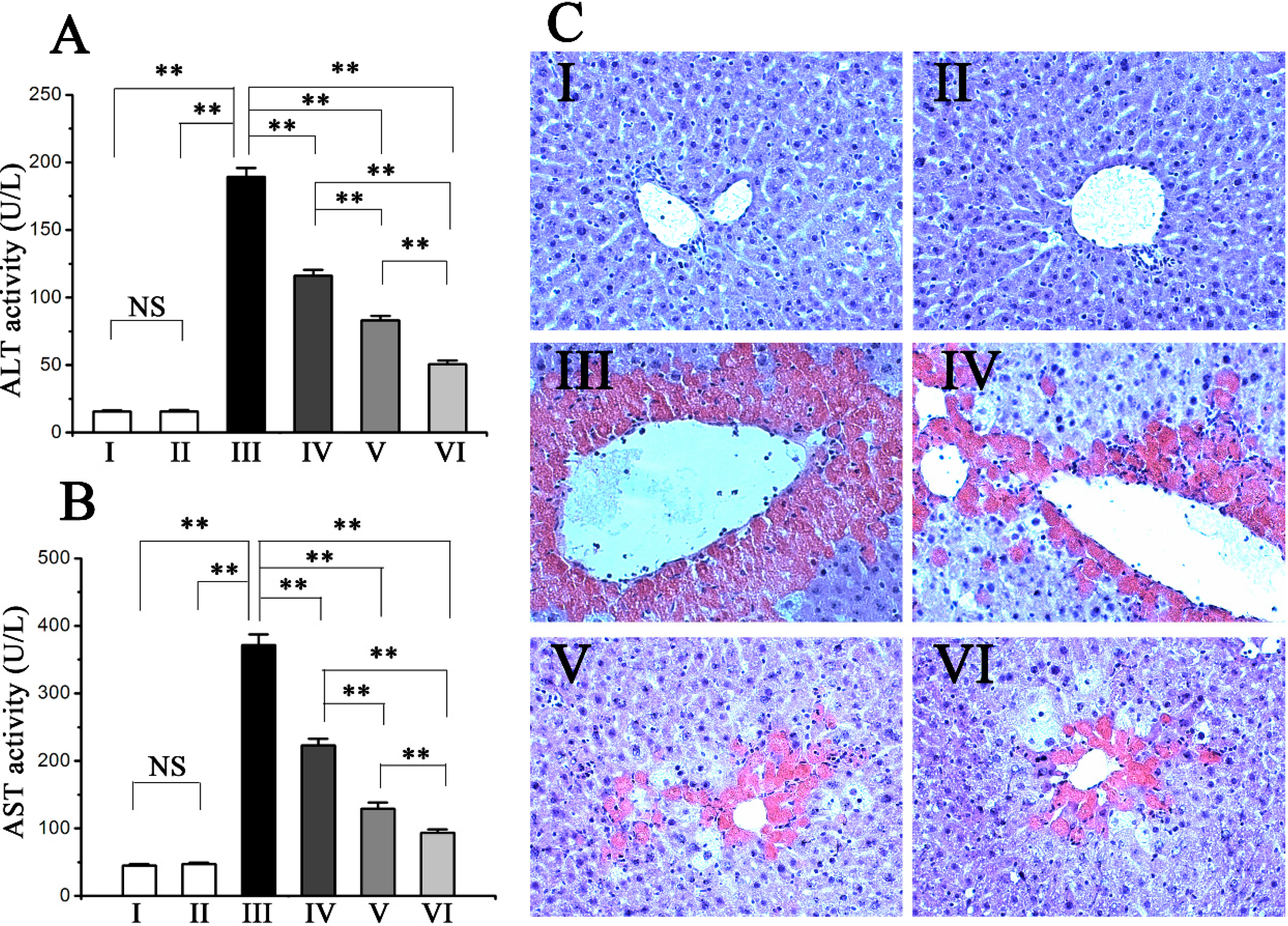
2.3. Procyanidin B2 Suppressed CCl4-Induced Oxidative Liver Injury

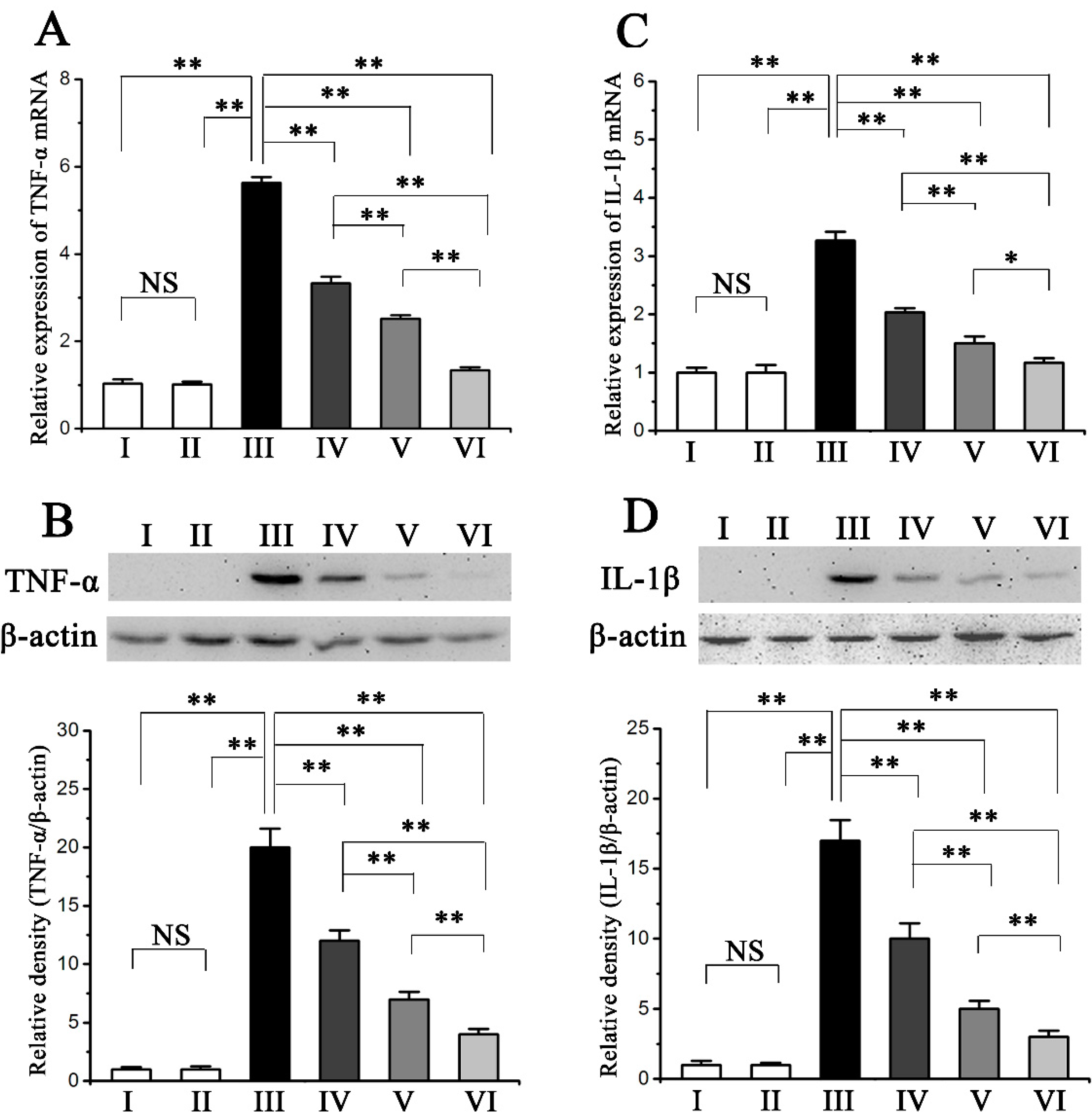
2.4. Procyanidin B2 Inhibited CCl4-Induced Pro-Inflammatory Response
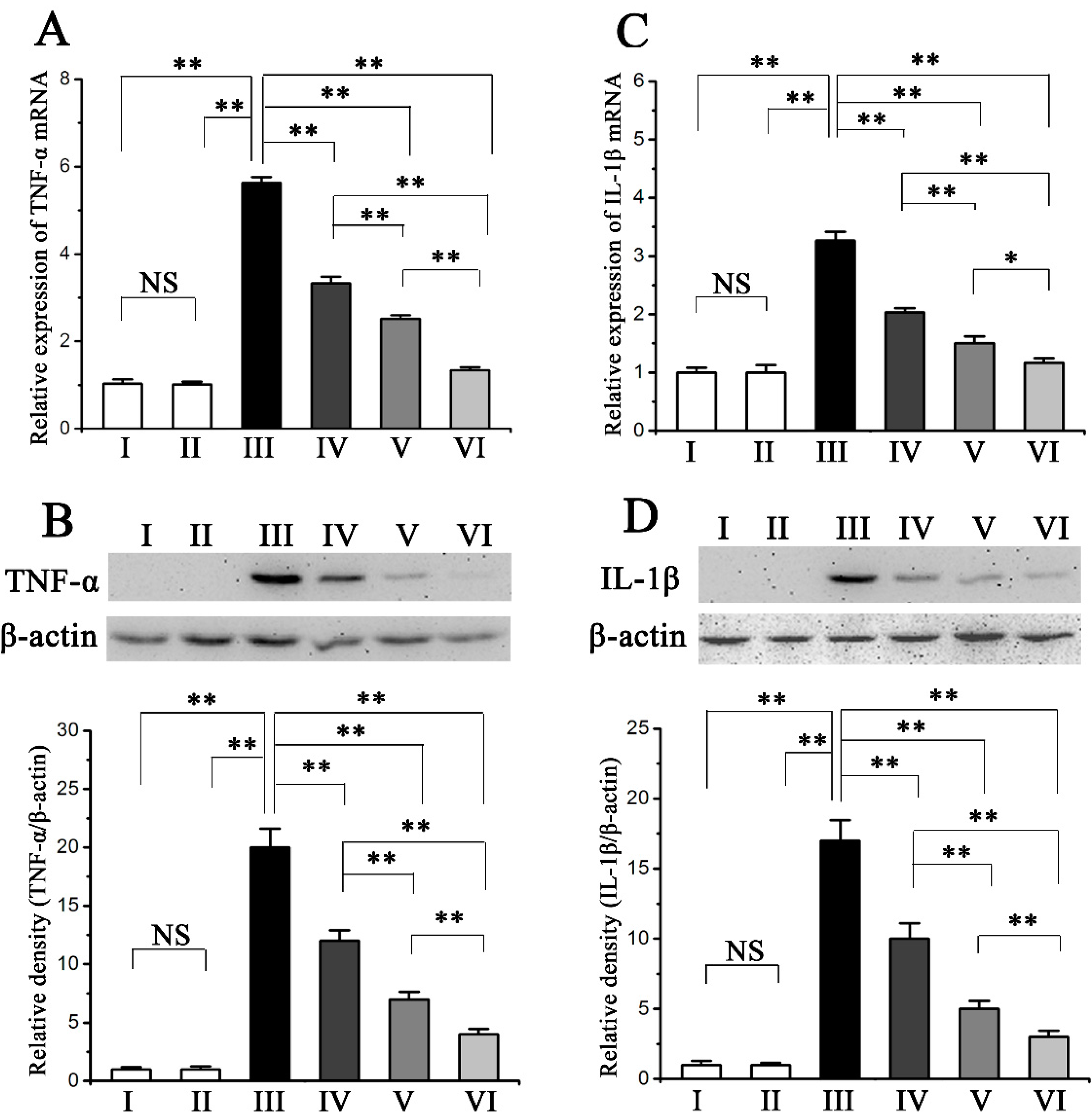
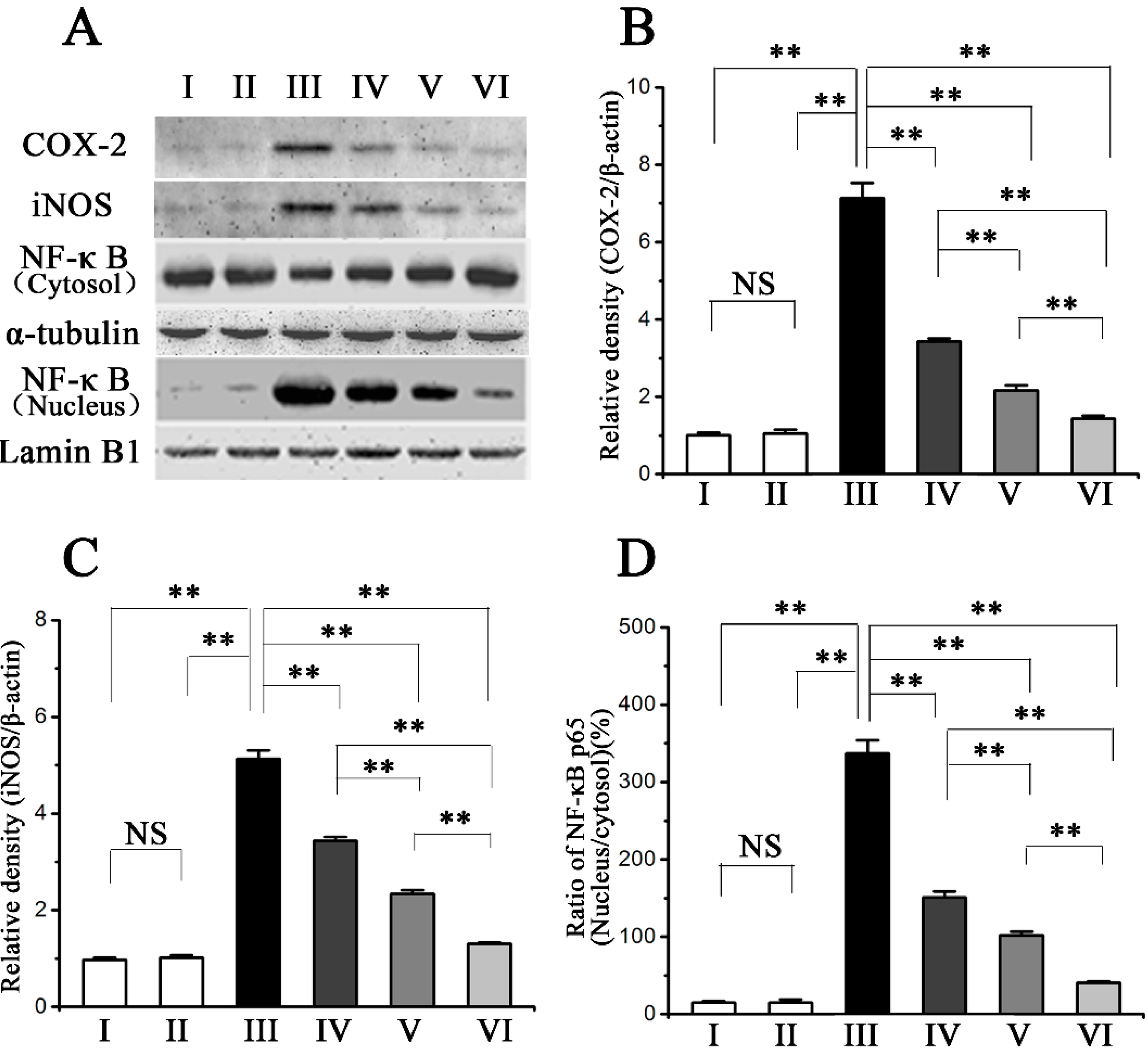
2.5. Procyanidin B2 Decreases CCl4-Induced Apoptosis of Hepatocytes
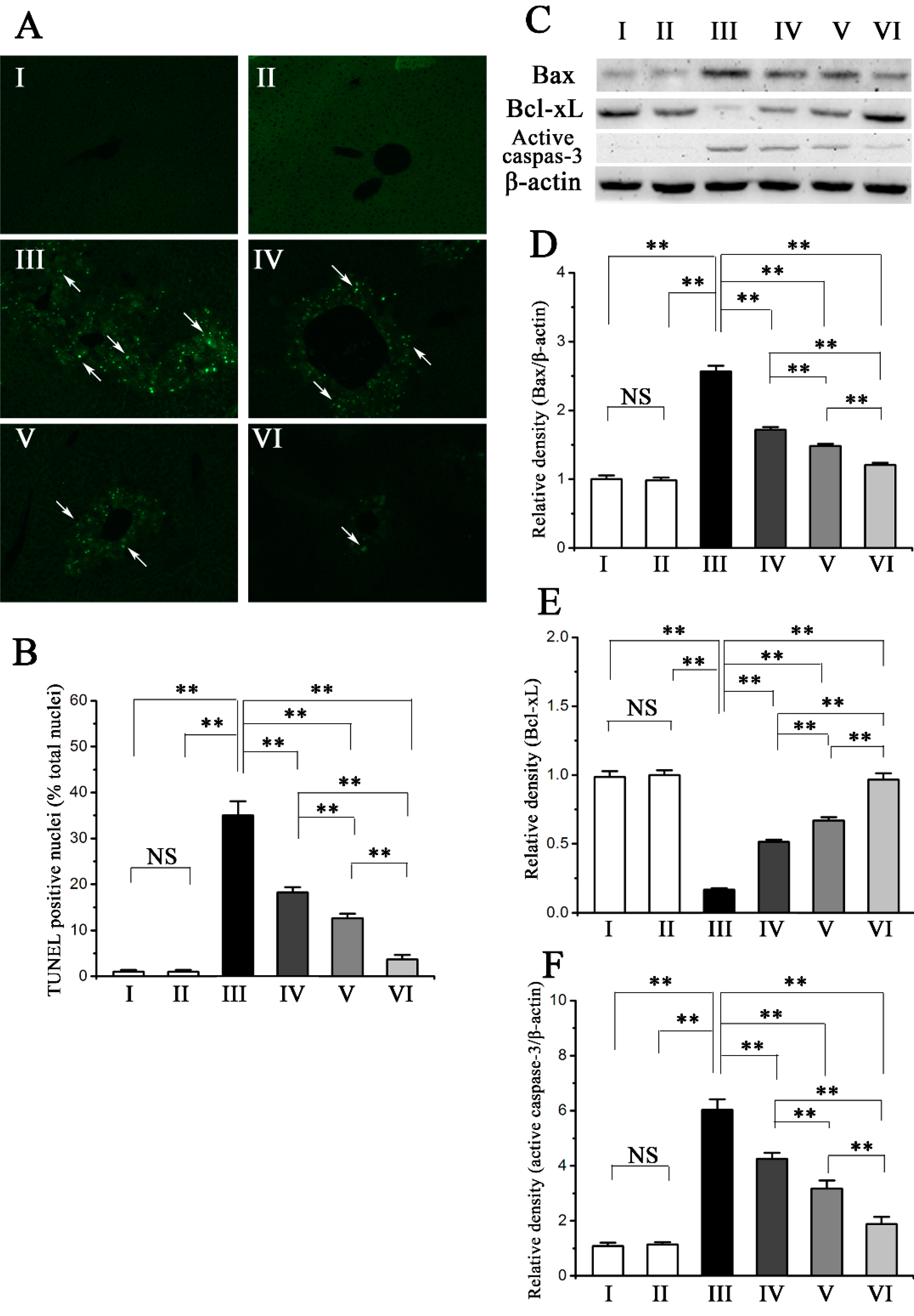
2.6. Discussion
3. Experimental Section
3.1. Chemical and Reagents
3.2. Animals
3.3. Experimental Design
3.4. Assessment of Liver Function
3.5. Assay of Hepatic MDA, GSH-Px, SOD and CAT Levels
3.6. RNA Extraction and Quantitative Real Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Primer Sequences (5′-3′) | Product Length (bp) |
|---|---|---|
| TNF-α (M11731) | Forward: GGCAGGTCTACTTTGGAGTC | 233 |
| Reverse: CACTGTCCCAGCATCTTGTG | ||
| IL-1β (NM_008361.3) | Forward: GCAGGCAGTATCACTCATTG | 165 |
| Reverse: CACACCAGCAGGTTATCATC | ||
| GAPDH (NM_008084.2) | Forward: CATCAACGGGAAGCCCATC | 211 |
| Reverse: CTCGTGGTTCACACCCATC |
3.7. Histological Examination of Liver Tissue
3.8. TUNEL Assay
3.9. Western Blot Assay
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Dwivedi1, S.; Khatri1, P.; Rajwar1, S.; Dwivedi1, A. Pharmacognostic and pharmacological aspects of potent herbal hepatoprotective drugs—A review. Int. J. Res. Pharm. Biomed. Sci. 2011, 2, 492–499. [Google Scholar]
- Sun, H.; Chen, L.; Zhou, W.; Hu, L.; Li, L.; Tu, Q.; Chang, Y.; Liu, Q.; Sun, X.; Wu, M.; et al. The protective role of hydrogen-rich saline in experimental liver injury in mice. J. Hepatol. 2011, 54, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Berasain, C.; Castillo, J.; Perugorria, M.J.; Latasa, M.U.; Prieto, J.; Avila, M.A. Inflammation and liver cancer: New molecular links. Ann. N. Y. Acad. Sci. 2009, 1155, 206–221. [Google Scholar] [CrossRef] [PubMed]
- Karakus, E.; Karadeniz, A.; Simsek, N.; Can, I.; Kara, A.; Yildirim, S.; Kalkan, Y.; Kisa, F. Protective effect of Panax ginseng against serum biochemical changes and apoptosis in liver of rats treated with carbon tetrachloride (CCl4). J. Hazard Mater. 2011, 195, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Stohs, S.J. Oxygen free radical scavenging abilities of vitamins C and E, and a grape seed proanthocyanidin extract in vitro. Res. Commun. Mol. Pathol. Pharmacol. 1997, 95, 179–189. [Google Scholar] [PubMed]
- Terra, X.; Montagut, G.; Bustos, M.; Llopiz, N.; Ardevol, A.; Blade, C.; Fernandez-Larrea, J.; Pujadas, G.; Salvado, J.; Arola, L.; et al. Grape-seed procyanidins prevent low-grade inflammation by modulating cytokine expression in rats fed a high-fat diet. J. Nutr. Biochem. 2009, 20, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.Á. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2011, 51, 881–892. [Google Scholar]
- Tyagi, A.; Raina, K.; Shrestha, S.P.; Miller, B.; Thompson, J.A.; Wempe, M.F.; Agarwal, R.; Agarwal, C. Procyanidin B2 3,3″-di-O-gallate, a biologically active constituent of grape seed extract, induces apoptosis in human prostate cancer cells via targeting NF-κB, Stat3, and AP1 transcription factors. Nutr. Cancer 2013, 66, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xiao, L.; Yuan, Y.; Luo, X.; Jiang, M.; Ni, J.; Wang, N. Procyanidin B2 inhibits NLRP3 inflammasome activation in human vascular endothelial cells. Biochem. Pharmacol. 2014, 92, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Xu, Y.; Xu, L.; Cong, X.; Yin, L.; Li, H.; Peng, J. Mechanism investigation of dioscin against CCl4-induced acute liver damage in mice. Environ. Toxicol. Pharmacol. 2012, 34, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Boll, M.; Weber, L.W.; Becker, E.; Stampfl, A. Mechanism of carbon tetrachloride-induced hepatotoxicity. Hepatocellular damage by reactive carbon tetrachloride metabolites. Z. Naturforsch. C 2001, 56, 649–659. [Google Scholar]
- Szymonik-Lesiuk, S.; Czechowska, G.; Stryjecka-Zimmer, M.; Słomka, M.; Mądro, A.; Celiński, K.; Wielosz, M. Catalase, superoxide dismutase, and glutathione peroxidase activities in various rat tissues after carbon tetrachloride intoxication. J. Hepatobiliary Pancreat. Surg. 2003, 10, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.; Mikkelsen, B.B.; Nielsen, J.B.; Andersen, H.R.; Grandjean, P. Plasma malondialdehyde as biomarker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem. 1997, 43, 1209–1214. [Google Scholar] [PubMed]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of malondialdehyde (MDA) by high-performance liquid chromatography in serum and liver as a biomarker for oxidative stress. Application to a rat model for hypercholesterolemia and evaluation of the effect of diets rich in phenolic antioxidants from fruits. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 827, 76–82. [Google Scholar]
- Bowling, A.C.; Schulz, J.B.; Brown, R.H., Jr.; Beal, M.F. Superoxide dismutase activity, oxidative damage, and mitochondrial energy metabolism in familial and sporadic amyotrophic lateral sclerosis. J. Neurochem. 1993, 61, 2322–2325. [Google Scholar] [CrossRef] [PubMed]
- Heck, D.E.; Shakarjian, M.; Kim, H.D.; Laskin, J.D.; Vetrano, A.M. Mechanisms of oxidant generation by catalase. Ann. N. Y. Acad. Sci. 2010, 1203, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Zamocky, M.; Furtmuller, P.G.; Obinger, C. Evolution of catalases from bacteria to humans. Antioxid. Redox Signal. 2008, 10, 1527–1548. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Ren, N.; Gao, H.; Lei, X.; Zheng, J.; Cao, W. Antioxidant and hepatoprotective effects of Schisandra chinensis pollen extract on CCl4-induced acute liver damage in mice. Food Chem. Toxicol. 2013, 55, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, G.D.; D’Souza, N.B. Hepatic antioxidant enzymes and lipid peroxidation in carbon tetrachloride-induced liver cirrhosis in rats. Biochem. Med. Metab. Biol. 1988, 40, 42–45. [Google Scholar] [CrossRef]
- Ebaid, H.; Bashandy, S.A.; Alhazza, I.M.; Rady, A.; El-Shehry, S. Folic acid and melatonin ameliorate carbon tetrachloride-induced hepatic injury, oxidative stress and inflammation in rats. Nutr. Metab. (Lond.) 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Kiso, K.; Ueno, S.; Fukuda, M.; Ichi, I.; Kobayashi, K.; Sakai, T.; Fukui, K.; Kojo, S. The role of Kupffer cells in carbon tetrachloride intoxication in mice. Biol. Pharm. Bull. 2012, 35, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.J.; Keller, B.J.; Kauffman, F.C.; Thurman, R.G. The involvement of Kupffer cells in carbon tetrachloride toxicity. Toxicol. Appl. Pharmacol. 1993, 119, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Liong, E.C.; Ling, M.T.; Ching, Y.P.; Fung, M.L.; Tipoe, G.L. S-allylmercaptocysteine reduces carbon tetrachloride-induced hepatic oxidative stress and necroinflammation via nuclear factor kappa B-dependent pathways in mice. Eur. J. Nutr. 2012, 51, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J.; Karin, M. Nuclear factor-kappaB: A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar] [PubMed]
- Chang, F.; Steelman, L.S.; Shelton, J.G.; Lee, J.T.; Navolanic, P.M.; Blalock, W.L.; Franklin, R.; McCubrey, J.A. Regulation of cell cycle progression and apoptosis by the Ras/Raf/MEK/ERK pathway (Review). Int. J. Oncol. 2003, 22, 469–480. [Google Scholar] [PubMed]
- Ma, S.; Shan, L.Q.; Xiao, Y.H.; Li, F.; Huang, L.; Shen, L.; Chen, J.H. The cytotoxicity of methacryloxylethyl cetyl ammonium chloride, a cationic antibacterial monomer, is related to oxidative stress and the intrinsic mitochondrial apoptotic pathway. Braz. J. Med. Biol. Res. 2011, 44, 1125–1133. [Google Scholar] [PubMed]
- Chou, A.H.; Yeh, T.H.; Kuo, Y.L.; Kao, Y.C.; Jou, M.J.; Hsu, C.Y.; Tsai, S.R.; Kakizuka, A.; Wang, H.L. Polyglutamine-expanded ataxin-3 activates mitochondrial apoptotic pathway by upregulating Bax and downregulating Bcl-xL. Neurobiol. Dis. 2006, 21, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Rosse, T.; Olivier, R.; Monney, L.; Rager, M.; Conus, S.; Fellay, I.; Jansen, B.; Borner, C. Bcl-2 prolongs cell survival after Bax-induced release of cytochrome c. Nature 1998, 391, 496–499. [Google Scholar] [PubMed]
- El-Beshbishy, H.A.; Mohamadin, A.M.; Nagy, A.A.; Abdel-Naim, A.B. Amelioration of tamoxifen-induced liver injury in rats by grape seed extract, black seed extract and curcumin. Indian J. Exp. Biol. 2010, 48, 280–288. [Google Scholar] [PubMed]
- Pinheiro, F.V.; Pimentel, V.C.; de Bona, K.S.; Scola, G.; Salvador, M.; Funchal, C.; Moretto, M.B. Decrease of adenosine deaminase activity and increase of the lipid peroxidation after acute methotrexate treatment in young rats: Protective effects of grape seed extract. Cell Biochem. Funct. 2010, 28, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Sehirli, O.; Ozel, Y.; Dulundu, E.; Topaloglu, U.; Ercan, F.; Sener, G. Grape seed extract treatment reduces hepatic ischemia-reperfusion injury in rats. Phytother. Res. 2008, 22, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Dai, Y.; Li, X.; Niu, N.; Li, W.; Liu, F.; Zhao, Y.; Yu, Z. Inhibition of arsenic-induced rat liver injury by grape seed exact through suppression of NADPH oxidase and TGF-beta/Smad activation. Toxicol. Appl. Pharmacol. 2011, 254, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, S.; He, B.; Mi, Y.; Cao, H.; Zhang, C.; Li, L. Ameliorative effect of grape seed proanthocyanidin extract on thioacetamide-induced mouse hepatic fibrosis. Toxicol. Lett. 2012, 213, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.O.; Yoon, S.; Moon, J.O. The proanthocyanidins inhibit dimethylnitrosamine-induced liver damage in rats. Arch. Pharm. Res. 2010, 33, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.; Osakabe, N.; Natsume, M.; Terao, J. Absorption and urinary excretion of procyanidin B2 [epicatechin-(4beta-8)-epicatechin] in rats. Free Radic. Biol. Med. 2002, 33, 142–148. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds procyanidin B2 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.-Y.; Zhang, X.-Y.; Guan, S.-W.; Hua, Z.-C. Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice. Molecules 2015, 20, 12250-12265. https://doi.org/10.3390/molecules200712250
Yang B-Y, Zhang X-Y, Guan S-W, Hua Z-C. Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice. Molecules. 2015; 20(7):12250-12265. https://doi.org/10.3390/molecules200712250
Chicago/Turabian StyleYang, Bing-Ya, Xiang-Yu Zhang, Sheng-Wen Guan, and Zi-Chun Hua. 2015. "Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice" Molecules 20, no. 7: 12250-12265. https://doi.org/10.3390/molecules200712250
APA StyleYang, B.-Y., Zhang, X.-Y., Guan, S.-W., & Hua, Z.-C. (2015). Protective Effect of Procyanidin B2 against CCl4-Induced Acute Liver Injury in Mice. Molecules, 20(7), 12250-12265. https://doi.org/10.3390/molecules200712250