
Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions

 ,
,  and
and 
Abstract
:1. Introduction
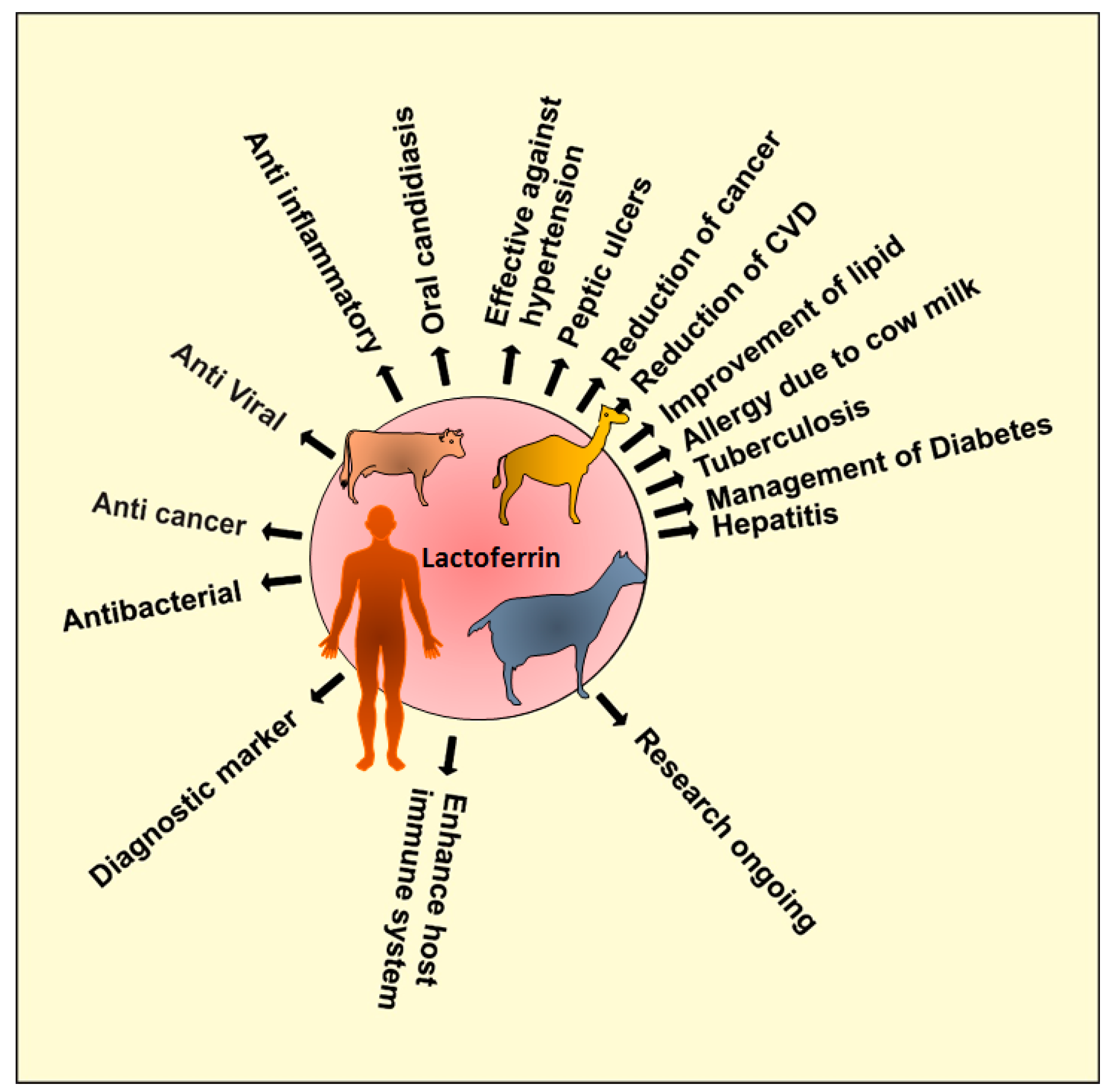
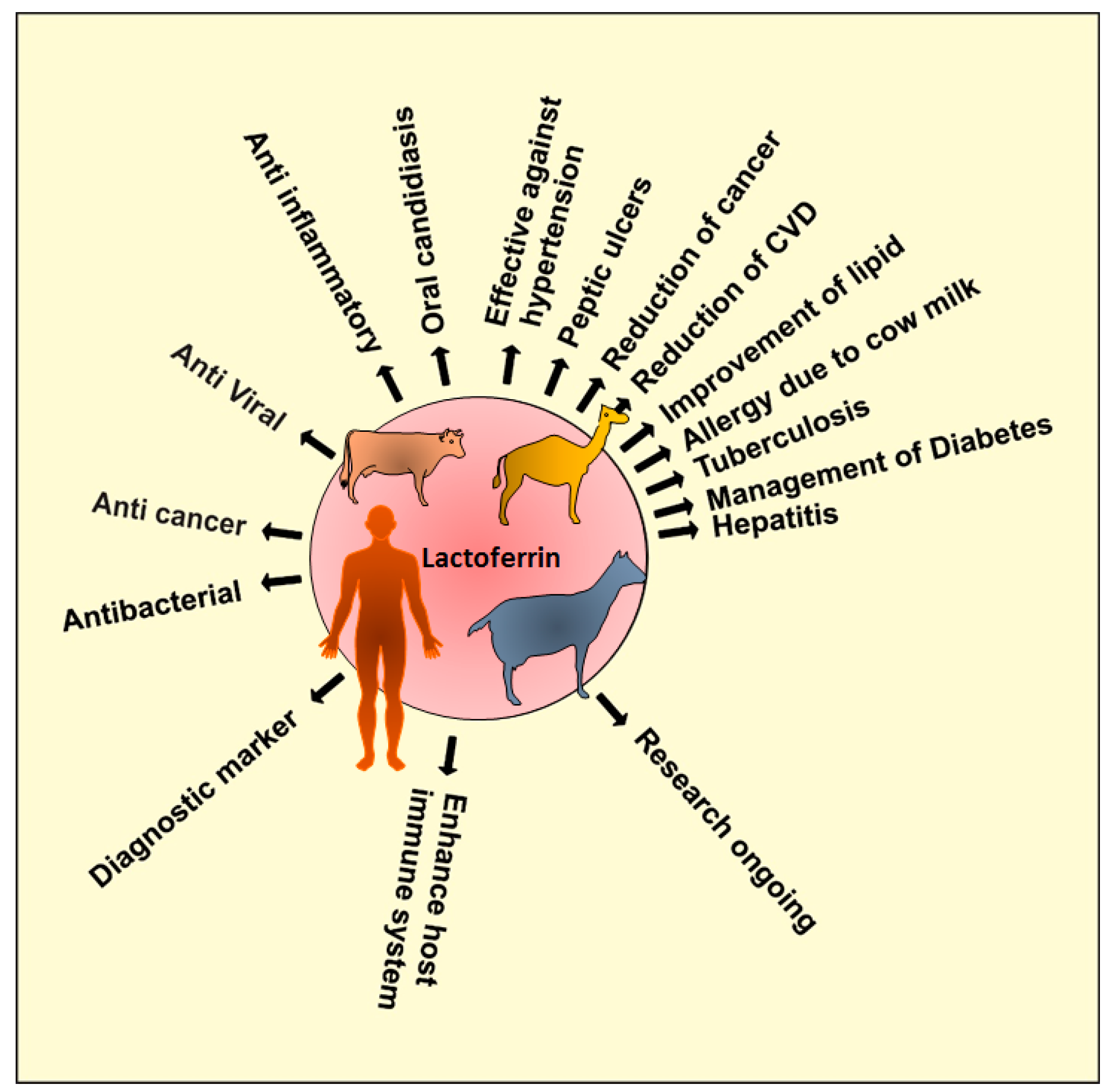
2. Sources of Lactoferrin

{kind=link}
{kind=link}
{kind=link}
| Lactoferrin Source | Action | Functional Role | Reference |
|---|---|---|---|
| Human Lactoferrin | Anti-microbial | Effective against Streptococcus, Salmonella, Shigella, Staphylococcus and Enterobacter. | [31–34] |
| Enhances the host immune system. | |||
| Anti-cancer | Diagnostic marker. | ||
| Goat Lactoferrin | Ongoing research | Still novel and further studies need to be conducted | [15] |
| Camel Lactoferrin | Anti-viral | Inhibits infection by Hepatitis C and B virus. It has hepatoprotective effect. | [16,17,35,36] |
| Anti-diabetic | Potential therapeutic molecule in targeting both type 1 and type 2 diabetes. More work needs to be done. | ||
| Bovine Lactoferrin | Anti-cancer | Anticancer activity against colorectal cancer and lung cancer. | [13,14,18,21,37,38] |
| Anti-microbial | Effective against oral candidiasis, influenza virus pneumonia and skin infections due to herpes virus. | ||
| Enhances host immune response | |||
| Anti-inflammatory. |
2.1. Human Lactoferrin
2.2. Goat Lactoferrin
2.3. Camel Lactoferrin
2.4. Bovine Lactoferrin
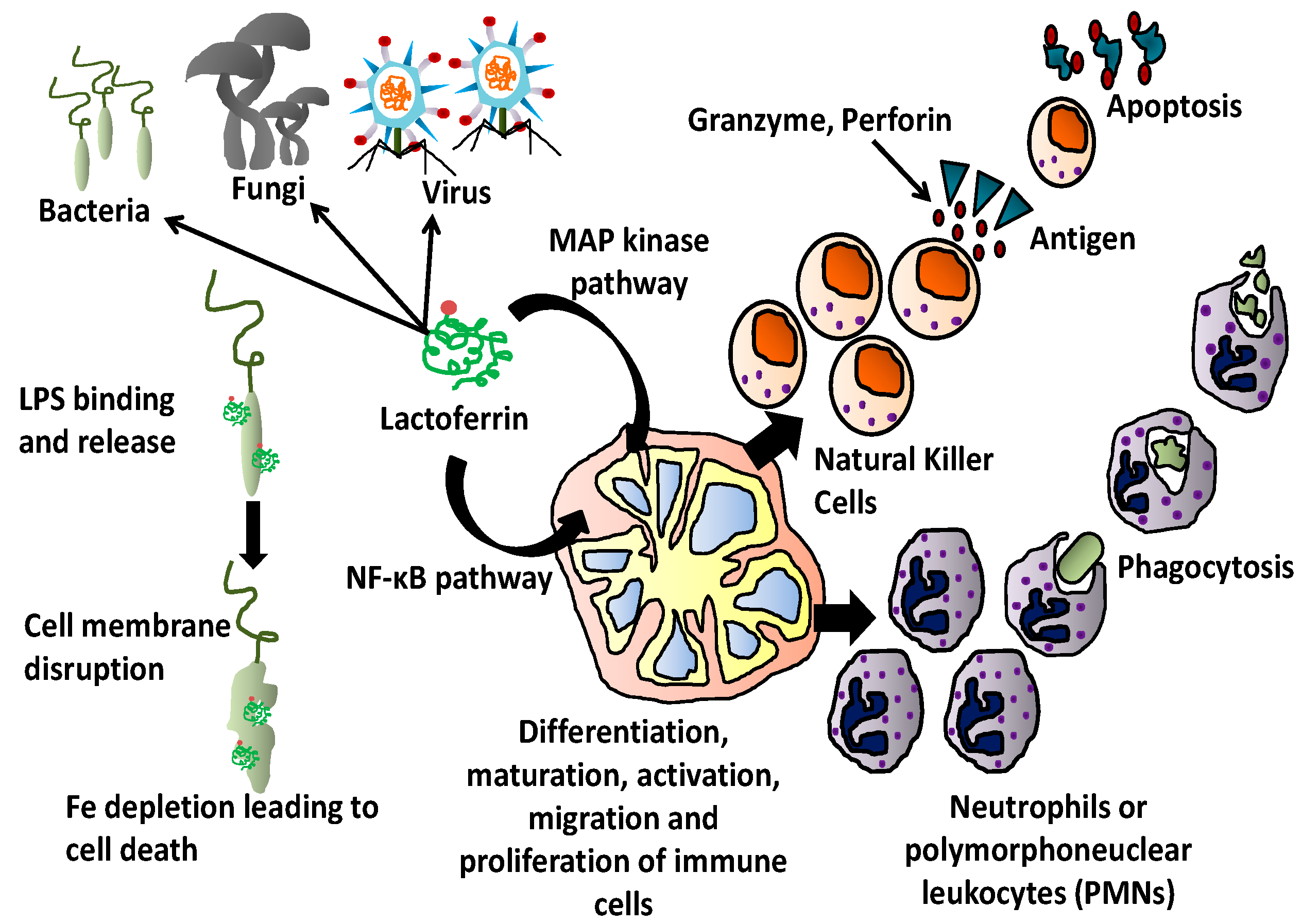
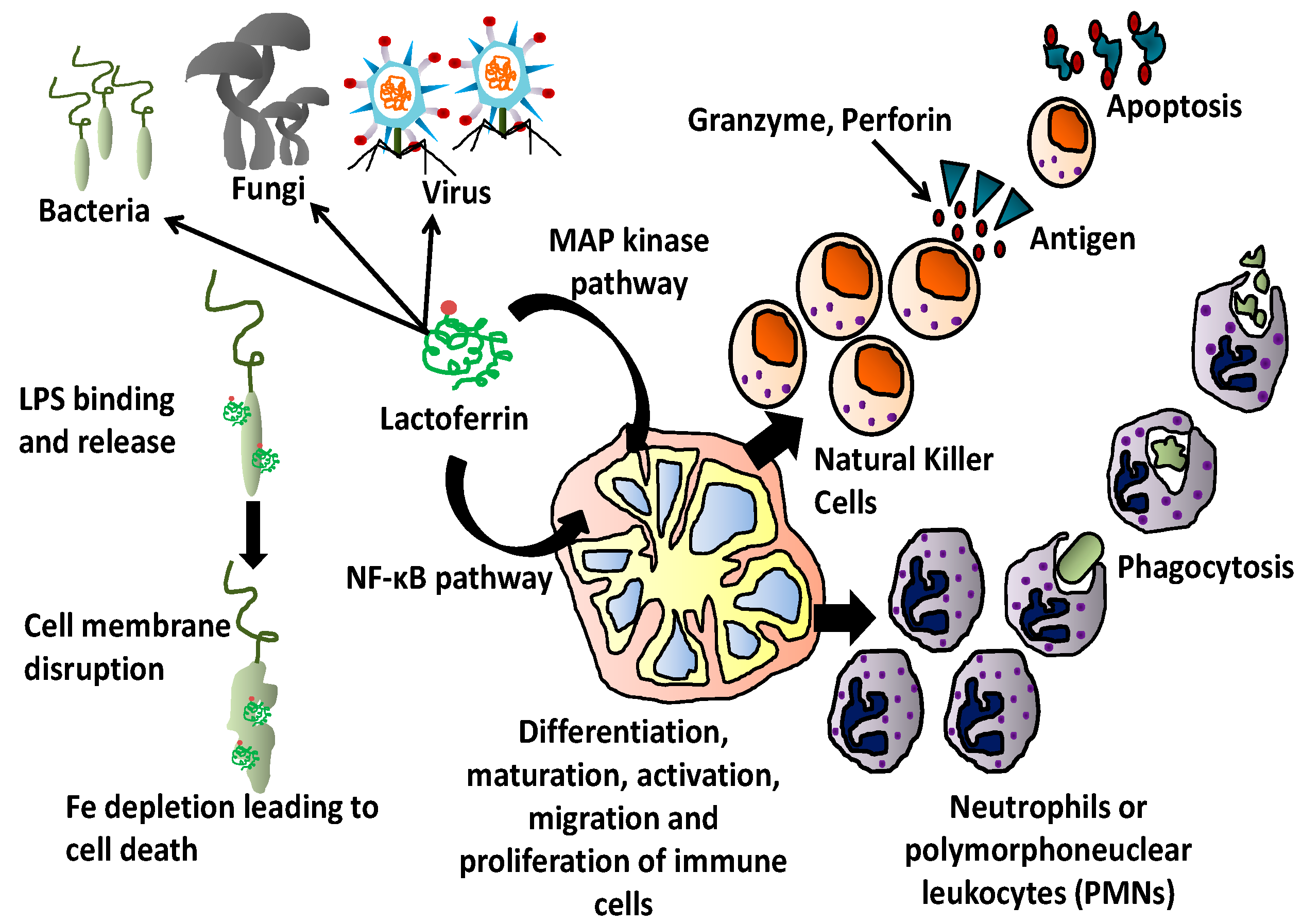
3. Antimicrobial Activity of Lactoferrin
4. Lactoferrin and Immunity
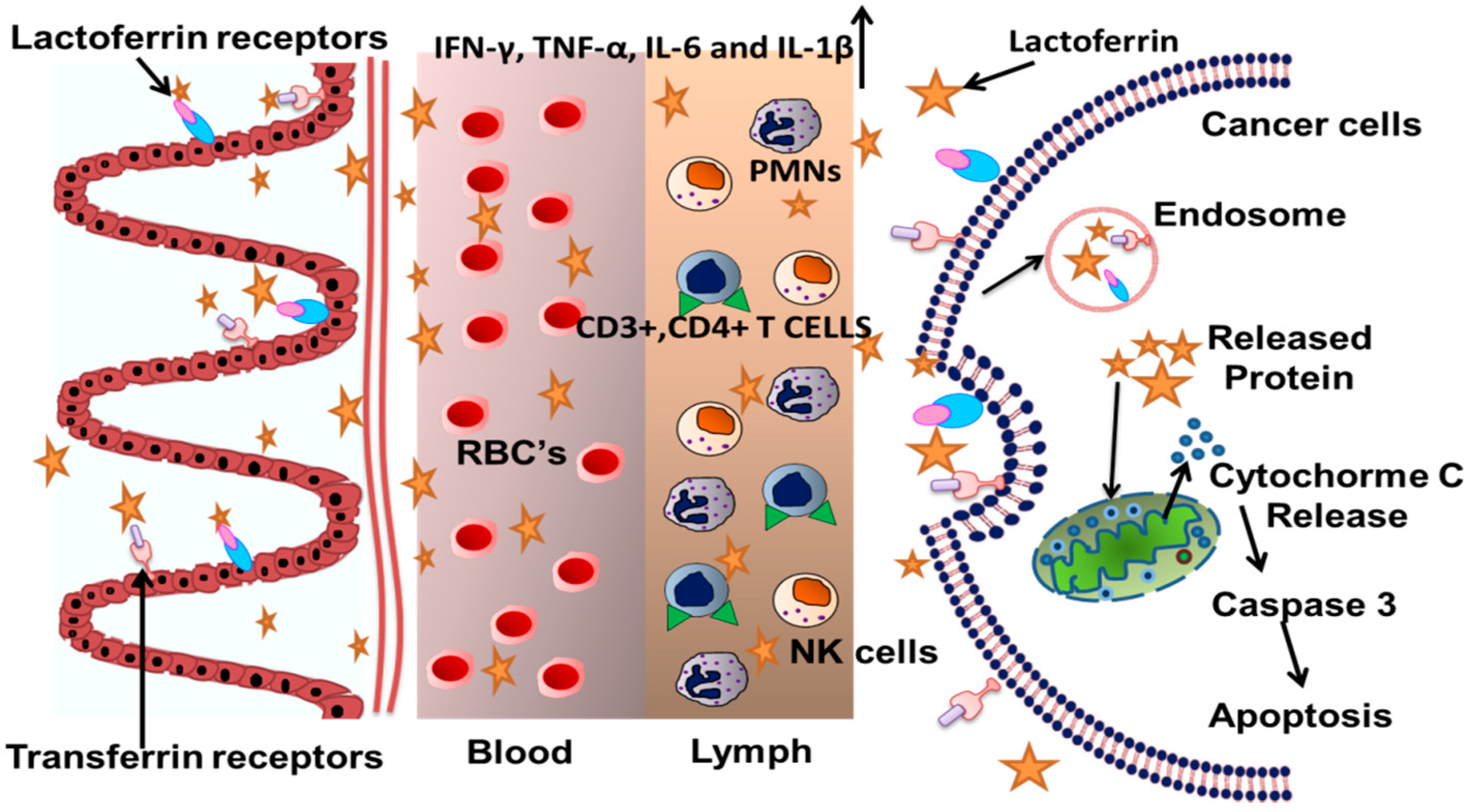
5. Nanoparticle-Based Cancer Therapy
5.1. Types of Nanoparticles Used
5.2. Nanoparticle Based Therapy Using Lactoferrin
| Type of Nanoparticle | Type of Cancer | Reference |
|---|---|---|
| Alginate enclosed chitosan conjugated, calcium phosphate-iron-saturated bovine lactoferrin nanocarriers | Colon cancer and colon cancer stem like cells | [14] |
| PEG-PLGA nanoparticles | 4T1 murine breast cancer | [166] |
| Silica nanoparticles targeted with anti-HER-2 Ab | MCF-7 breast cancer cells | [168] |
| PEG-PLA nanoparticles | Brain delivery | [172,173] |
| Super paramagnetic Iron oxide Nanoparticles | As a specific MRI contrast agent for detection of brain glioma | [174] |
| Biodegradable Polymersome | Chemotherapy of Glioma Rats | [175] |
| Procationic liposomes | Glioma | [176] |
| PEI/pDNA nanoparticles | Airway epithelial cells | [177] |
| Polyamidoamine (PAMAM) conjugated with lactoferrin | Rotenone-induced chronic Parkinson model | [178] |
| Chitosan/alginate/calcium complex micro particles loaded with lactoferrin | Carrageenan-induced edema in rats | [179] |
| Lf modified DNA loaded nanoparticles | Brain capillary endothelial cells | [180] |
| Alginate enclosed chitosan coated calcium phosphate nanoparticles | Colon cancer | [181] |
| Paclitaxel loaded Lf coupled solid lipid nanoparticles (SLPs) | Human bronchial epithelial cells (BEAS-2B) | [182] |
| Folic acid and Lf functionalized PLGA nanoparticles loaded with etoposide | Glioblastoma | [183] |
6. Lf in Clinical Trials
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nagpal, R.; Behare, P.; Rana, R.; Kumar, A.; Kumar, M.; Arora, S.; Morotta, F.; Jain, S.; Yadav, H. Bioactive peptides derived from milk proteins and their health beneficial potentials: An update. Food Funct. 2011, 2, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.; Baker, H. Lactoferrin. Cell. Mol. Life Sci. 2005, 62, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Iigo, M.; Alexander, D.B.; Long, N.; Xu, J.; Fukamachi, K.; Futakuchi, M.; Takase, M.; Tsuda, H. Anticarcinogenesis pathways activated by bovine lactoferrin in the murine small intestine. Biochimie 2009, 91, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Birgens, H.S. Lactoferrin in plasma measured by an elisa technique: Evidence that plasma lactoferrin is an indicator of neutrophil turnover and bone marrow activity in acute leukaemia. Scand. J. Haematol. 1985, 34, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between lactoferrin and beneficial microbiota in breast milk and infant’s feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Parhi, P.; Mohanty, C.; Sahoo, S.K. Nanotechnology-based combinational drug delivery: An emerging approach for cancer therapy. Drug Discov. Today 2012, 17–18, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, L.; Calvo, M.; Brock, J.H. Biological role of lactoferrin. Arch. Dis. Child. 1992, 67, 657–661. [Google Scholar] [CrossRef] [PubMed]
- González-Chávez, S.A.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. Lactoferrin: Structure, function and applications. Int. J. Antimicrob. Agents 2009, 33. [Google Scholar] [CrossRef] [PubMed]
- Iafisco, M.; Foggia, M.D.; Bonora, S.; Prat, M.; Roveri, N. Adsorption and spectroscopic characterization of lactoferrin on hydroxyapatite nanocrystals. Dalton Trans. 2011, 40, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Aisen, P.; Leibman, A. Lactoferrin and transferrin: A comparative study. BBA-Protein Struct. 1972, 257, 314–323. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Yamauchi, K.; Takase, M. Lactoferrin research, technology and applications. Int. Dairy J. 2006, 16, 1241–1251. [Google Scholar] [CrossRef]
- Burrow, H.; K Kanwar, R.; R Kanwar, J. Antioxidant enzyme activities of iron-saturated bovine lactoferrin (Fe-blf) in human gut epithelial cells under oxidative stress. Med. Chem. 2011, 7, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Palmano, K.P.; Sun, X.; Kanwar, R.K.; Gupta, R.; Haggarty, N.; Rowan, A.; Ram, S.; Krissansen, G.W. Iron-saturated lactoferrin is a potent natural adjuvant for augmenting cancer chemotherapy. Immunol. Cell Biol. 2008, 86, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Mahidhara, G.; Roy, K.; Sasidharan, S.; Krishnakumar, S.; Prasad, N.; Sehgal, R.; Kanwar, R.K. Fe-blf nanoformulation targets survivin to kill colon cancer stem cells and maintains absorption of iron, calcium and zinc. Nanomedicine 2014, 10, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Hiss, S.; Meyer, T.; Sauerwein, H. Lactoferrin concentrations in goat milk throughout lactation. Small Rumin. Res. 2008, 80, 87–90. [Google Scholar] [CrossRef]
- Hara, K.; Ikeda, M.; Saito, S.; Matsumoto, S.; Numata, K.; Kato, N.; Tanaka, K.; Sekihara, H. Lactoferrin inhibits hepatitis b virus infection in cultured human hepatocytes. Hepatol. Res. 2002, 24, 228–235. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Dogra, R.; Mohta, N.; Tiwari, R.; Singhal, S.; Sultania, S. Beneficial effect of camel milk in diabetic nephropathy. Acta Biomed. 2009, 80, 131–134. [Google Scholar] [PubMed]
- Gupta, I.; Sehgal, R.; Kanwar, R.K.; Punj, V.; Kanwar, J.R. Nanocapsules loaded with iron-saturated bovine lactoferrin have antimicrobial therapeutic potential and maintain calcium, zinc and iron metabolism. Nanomedicine 2014, 10, 1289–1314. [Google Scholar] [CrossRef] [PubMed]
- Roseanu, A.; Chelu, F.; Trif, M.; Motas, C.; Brock, J.H. Inhibition of binding of lactoferrin to the human promonocyte cell line thp-1 by heparin: The role of cell surface sulphated molecules. Biochim. Biophys. Acta 2000, 1475, 35–38. [Google Scholar] [CrossRef]
- Wilk, K.M.; Hwang, S.A.; Actor, J.K. Lactoferrin modulation of antigen-presenting-cell response to bcg infection. Postepy Hig. Med. Dosw. 2007, 61, 277–282. [Google Scholar]
- Mohamad, R.H.; Zekry, Z.K.; Al-Mehdar, H.A.; Salama, O.; El-Shaieb, S.E.; El-Basmy, A.A.; Al-said, M.G.; Sharawy, S.M. Camel milk as an adjuvant therapy for the treatment of type 1 diabetes: Verification of a traditional ethnomedical practice. J. Med. Food 2009, 12, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Ohshima, Y.; Nomoto, H.; Fujita, K.I.; Matsuda, E.; Iigo, M.; Takasuka, N.; Moore, M.A. Cancer prevention by natural compounds. Drug Metabol. Pharmacokinet. 2004, 19, 245–263. [Google Scholar] [CrossRef]
- Zhang, Y.; Lima, C.F.; Rodrigues, L.R. Anticancer effects of lactoferrin: Underlying mechanisms and future trends in cancer therapy. Nutr. Rev. 2014, 72, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Takakura, N.; Yamauchi, K.; Tamura, Y. Modulation of immunity-related gene expression in small intestines of mice by oral administration of lactoferrin. Clin. Vaccine Immunol. 2006, 13, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Reitamo, S.; Konttinen, Y.; Segerberg-Konttinen, M. Distribution of lactoferrin in human salivary glands. Histochemistry 1980, 66, 285–291. [Google Scholar] [CrossRef] [PubMed]
- McClellan, K. Mucosal defense of the outer eye. Surv. Ophthalmol. 1997, 42, 233–246. [Google Scholar] [CrossRef]
- Valore, E.V.; Park, C.H.; Igreti, S.L.; Ganz, T. Antimicrobial components of vaginal fluid. Am. J. Obstet. Gynecol. 2002, 187, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Breton-Gorius, J.; Mason, D.; Buriot, D.; Vilde, J.; Griscelli, C. Lactoferrin deficiency as a consequence of a lack of specific granules in neutrophils from a patient with recurrent infections. Detection by immunoperoxidase staining for lactoferrin and cytochemical electron microscopy. Am. J. Pathol. 1980, 99, 413–428. [Google Scholar] [PubMed]
- Masson, P.L.; Heremans, J.F.; Schonne, E. Lactoferrin, an iron-binding protein in neutrophilic leukocytes. J. Exp. Med. 1969, 130, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Caccavo, D.; Garzia, P.; Sebastiani, G.D.; Ferri, G.M.; Galluzzo, S.; Vadacca, M.; Rigon, A.; Afeltra, A.; Amoroso, A. Expression of lactoferrin on neutrophil granulocytes from synovial fluid and peripheral blood of patients with rheumatoid arthritis. J. Rheumatol. 2003, 30, 220–224. [Google Scholar] [PubMed]
- Arnold, R.R.; Brewer, M.; Gauthier, M.M. Bactericidal activity of human lactoferrin: Sensitivity of a variety of microorganisms. Infect. Immun. 2001, 28, 893–898. [Google Scholar]
- Bezault, J.; Bhimani, R.; Wiprovnick, J.; Firmanski, P. Human lactoferrin inhibits growth of solid tumors and development of metastasis in mice. Cancer Res. 1994, 54, 2310–2313. [Google Scholar] [PubMed]
- Nichols, B.L.; Mckee, K.S.; Henry, J.F.; Putman, M. Human lactoferrin stimulates thymidine incorporation into DNA of rat crypt cells. Pediat. Res. 1987, 21, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Matsuse, R.; Tomiota, S.; Sugi, K.; Saitoh, O.; Ohshiba, S. Immunochemical detection of human lactoferrin in feces as a new marker for inflammatory gastrointestinal disorders and colon cancer. Clin. Biochem. 1994, 22, 259–264. [Google Scholar] [CrossRef]
- Redwan, E.R.M.; Tabll, A. Camel lactoferrin markedly inhibits hepatitis c virus genotype 4 infection of human peripheral blood leukocytes. J. Immun. Immunochem. 2007, 28, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, A.; Tanaka, K.; Naganuma, A.; Kato, N. Recent advances of basic research and clinical application of lactoferrin as an antiviral reagent against chronic hepatitis c. Nippon Rinsho 2002, 60, 819–829. [Google Scholar] [PubMed]
- Iigo, M.; Alexander, D.B.; Xu, J.; Futakuchi, M.; Suzui, M.; Kozu, T.; Akasu, T.; Saito, D.; Kakizoe, T.; Yamauchi, K. Inhibition of intestinal polyp growth by oral ingestion of bovine lactoferrin and immune cells in the large intestine. Biometals 2014, 27, 1017–1029. [Google Scholar] [CrossRef] [PubMed]
- Samarasinghe, R.M.; Kanwar, R.K.; Kanwar, J.R. The effect of oral administration of iron saturated-bovine lactoferrin encapsulated chitosan-nanocarriers on osteoarthritis. Biomaterials 2014, 35, 7522–7534. [Google Scholar] [CrossRef] [PubMed]
- Crouch, S.P.; Slater, K.J.; Fletcher, J. Regulation of cytokine release from mononuclear cells by the iron-binding protein lactoferrin. Blood 1992, 80, 235–240. [Google Scholar] [PubMed]
- Yamaguchi, M.; Matsuura, M.; Kobayashi, K.; Sasaki, H.; Yajima, T.; Kuwata, T. Lactoferrin protects against development of hepatitis caused by sensitization of kupffer cells by lipopolysaccharide. Clin. Diagn. Lab. Immunol. 2001, 8, 1234–1239. [Google Scholar] [CrossRef] [PubMed]
- Haversen, L.; Ohlsson, B.G.; Hahn-Zoric, M.; Hanson, L.A.; Mattsby-Baltzer, I. Lactoferrin down-regulates the LPS-induced cytokine production in monocytic cells via NF-kappa b. Cell. Immunol. 2002, 220, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Sato, A.; Shimizu, H.; Ando, K.; Takeuchi, T. Pegylated lactoferrin enhances its hepatoprotective effects on acute liver injury induced by d-galactosamine and lipopolysaccharide in rats. J. Vet. Med. Sci. 2010, 72, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.F.; Arnold, R.R.; Mestecky, J.; Prince, S.; Kulhavy, R.; McGhee, J.R. Studies with human lactoferrin and s. Mutans. In Microbial Aspects of Dental Caries; Stiles, M., Loesche, W.J., O’Brien, T.C., Eds.; Information Retrieval, Inc.: Washington, DC, USA, 1977; Volume 2, pp. 359–373. [Google Scholar]
- Le Parc, A.; Dallas, D.C.; Duaut, S.; Leonil, J.; Martin, P.; Barile, D. Characterization of goat milk lactoferrin n-glycans and comparison with the N-glycomes of human and bovine milk. Electrophoresis 2014, 35, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- El-Fakharany, E.M.; Tabil, A.; El-Wahab, A.A.; Haroun, B.M.; Redwan, E. Potential activity of camel milk-amylase and lactoferrin against hepatitis c virus infectivity in hepg2 and lymphocytes. Hepat. Mon. 2008, 8, 101–109. [Google Scholar]
- Bonkovsky, H.L.; Banner, B.F.; Rothman, A.L. Iron and chronic viral hepatitis. Hepatology 1997, 25, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Farinati, F.; Cardin, R.; Maria, N.D.; Libera, G.D.; Marafin, C.; Lecis, E. Iron storage, lipid peroxidation and glutathione turnover in chronic anti-HCV positive hepatitis. J. Hepatol. 1995, 22, 449–456. [Google Scholar] [CrossRef]
- Khan, J.A.; Kumar, P.; Paramasivam, M.; Yadav, R.S.; Sahani, M.S.; Sharma, S.; Srinivasan, A.; Singh, T.P. Camel lactoferrin, a transferrin-cum-lactoferrin crystal structures of camel apolactoferrin at a 2.6 Å resolution and structural basis of its dual role. J. Mol. Biol. 2001, 309, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.I.; Yamauchi, K.; Sugimoto, R.; Fujita, N.; Kobayashi, Y.; Watanabe, S. Lactoferrin inhibits lipid peroxidation in patients with chronic hepatitis c. Hepatol. Res. 2006, 36, 27–32. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, E.I. Bioactive components in camel milk. In Bioactive Components in Milk and Dairy Products, 1st ed.; Park, Y.W., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; Volume 1, pp. 159–194. [Google Scholar]
- Saltanat, H.; Li, H.; Xu, Y.; Wang, J.; Liu, F.; Geng, H.H. The influences of camel milk on the immune response of chronic hepatitis b patients. Milk Res. 2009, 25, 431–433. [Google Scholar]
- Al-Hashem, F.; Dallak, M.; Bashir, N.; Abbas, M. Camel’s milk protects against cadmium chloride induced toxicity in white albino rats. Am. J. Pharmacol. Toxicol. 2009, 4, 107–117. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. BBA-Protein Struct. Mol. Enzymol. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Chierici, R.; Sawatzki, G.; Tamisari, L.; Volpato, S.; Vigi, V. Supplementation of an adapted formula with bovine lactoferrin. 2. Effects on serum iron, ferritin and zinc levels. Acta Paediatr. 1992, 81, 475–479. [Google Scholar] [CrossRef] [PubMed]
- King, J.C., Jr.; Cummings, G.E.; Guo, N.; Trivedi, L.; Readmond, B.X.; Keane, V.; Feigelman, S.; de Waard, R. A double-blind, placebo-controlled, pilot study of bovine lactoferrin supplementation in bottle-fed infants. J. Pediat. Gastroenterol. Nutr. 2007, 44, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Takakura, N.; Wakabayashi, H.; Ishibashi, H.; Yamauchi, K.; Teraguchi, S.; Tamura, Y.; Yamaguchi, H.; Abe, S. Effect of orally administered bovine lactoferrin on the immune response in the oral candidiasis murine model. J. Med. Microbiol. 2004, 53, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.; Wakabayashi, H.; Yamauchi, K.; Teraguchi, S.; Tamura, Y.; Kurokawa, M.; Shiraki, K. Effects of orally administered bovine lactoferrin and lactoperoxidase on influenza virus infection in mice. J. Med. Microbiol. 2005, 54, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Pisani, S.; Antonini, G.; Valenti, P.; Seganti, L.; Orsi, N. Metal complexes of bovine lactoferrin inhibit in vitro replication of herpes simplex virus type 1 and 2. Biometals 1998, 11, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Debbabi, H.; Dubarry, M.; Boyaka, P.; Tome, D. Regulation of physiological and pathological th1 and th2 responses by lactoferrin this paper is one of a selection of papers published in this special issue, entitled 7th international conference on lactoferrin: Structure, function, and applications, and has undergone the journal's usual peer review process. Biochem. Cell Biol. 2006, 84, 303–311. [Google Scholar] [PubMed]
- Zhang, Y.; Nicolau, A.; Lima, C.F.; Rodrigues, L.R. Bovine lactoferrin induces cell cycle arrest and inhibits mtor signaling in breast cancer cells. Nutr. Cancer 2014, 66, 1371–1385. [Google Scholar] [CrossRef] [PubMed]
- Sekine, K.; Watanabe, E.; Nakamura, J.; Takasuka, N.; Kim, D.J.; Asamoto, M.; Krutovskikh, V.; Baba-Toriyama, H.; Ota, T.; Moore, M.A. Inhibition of azoxymethane-initiated colon tumor by bovine lactoferrin administration in f344 rats. Cancer Sci. 1997, 88, 523–526. [Google Scholar] [CrossRef]
- Ushida, Y.; Sekine, K.; Kuhara, T.; Takasuka, N.; Iigo, M.; Maeda, M.; Tsuda, H. Possible chemopreventive effects of bovine lactoferrin on esophagus and lung carcinogenesis in the rat. Cancer Sci. 1999, 90, 262–267. [Google Scholar] [CrossRef]
- Takakura, N.; Wakabayashi, H.; Ishibashi, H.; Teraguchi, S.; Tamura, Y.; Yamaguchi, H.; Abe, S. Oral lactoferrin treatment of experimental oral candidiasis in mice. Antimicrob. Agents Chemother. 2003, 47, 2619–2623. [Google Scholar] [CrossRef] [PubMed]
- Mulder, A.M.; Connellan, P.A.; Oliver, C.J.; Morris, C.A.; Stevenson, L.M. Bovine lactoferrin supplementation supports immune and antioxidant status in healthy human males. Nutr. Res. 2008, 28, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, I.; Kohno, K.; Tanimoto, T.; Ikegami, H.; Kurimoto, M. Development of CD8+ effector T cells is differentially regulated by IL-18 and IL-12. J. Immunol. 1999, 162, 3202–3211. [Google Scholar] [PubMed]
- Iigo, M.M.; Kuhara, T.T.; Ushida, Y.Y.; Sekine, K.K.; Moore, M.A.M.; Tsuda, H.H. Inhibitory effects of bovine lactoferrin on colon carcinoma 26 lung metastasis in mice. Clin. Exp. Metastas. 1999, 17, 35–40. [Google Scholar] [CrossRef]
- Yoo, Y.C.; Watanabe, S.; Watanabe, R.; Hata, K.; Shimazaki, K.I.; Azuma, I. Bovine lactoferrin and lactoferricin, a peptide derived from bovine lactoferrin, inhibit tumor metastasis in mice. Cancer Sci. 1997, 88, 184–190. [Google Scholar] [CrossRef]
- Macciò, A. Efficacy and safety of oral lactoferrin supplementation in combination with rhuepo-β for the treatment of anemia in advanced cancer patients undergoing chemotherapy: Open-label, randomized controlled study. Oncologist 2010, 15, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Kuhara, T.; Yamauchi, K.; Iwatsuki, K. Bovine lactoferrin induces interleukin-11 production in a hepatitis mouse model and human intestinal myofibroblasts. Eur. J. Nutr. 2012, 51, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Paesano, R.; Pacifici, E.; Benedetti, S.; Berlutti, F.; Frioni, A.; Polimeni, A.; Valenti, P. Safety and efficacy of lactoferrin versus ferrous sulphate in curing iron deficiency and iron deficiency anaemia in hereditary thrombophilia pregnant women: An interventional study. Biometals 2014, 27, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H. Lactoferrin delivery systems: Approaches for its more effective use. Expert Opin. Drug Deliv. 2011, 8, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Tan, A.X.; Vlassara, H. Antibacterial activity of lysozyme and lactoferrin is inhibited by binding of advanced glycation-modified proteins to a conserved motif. Nature Med. 1995, 1, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Bullen, J.J.; Rogers, H.J.; Leigh, L. Iron binding proteins in milk and resistance to escherichia coli infection in infants. Br. Med. J. 1972, 1, 68–75. [Google Scholar] [CrossRef]
- Ellison, R.T.; Giehl, T.J.; la Force, F.M. Damage of the outer membrane of enteric gram-negative bacteria by lactoferrin and transferrin. Infect. Immun. 1988, 56, 2774–2781. [Google Scholar] [PubMed]
- Leitch, E.C.; Willcox, M.D. Lactoferrin increases the susceptibility of s. Epidermidis biofilms to lysozyme and vancomycin. Curr. Eye Res. 1999, 19, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Leitch, E.C.; Willcox, M.D. Elucidation of the antistaphylococcal mechanism of lactoferrin and lysozyme. J. Med. Microbiol. 1999, 48, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Leitch, E.C.; Willcox, M.D. Lactoferrin induced reduction of vanb vancomycin resistance in enterococci. Int. J. Antimicrob. Agents 2000, 18, 399–402. [Google Scholar] [CrossRef]
- Roseanu, A.; Florian, P.; Condei, M.; Cristea, D.; Damian, M. Antibacterial activity of lactoferrin and lactoferricin against oral streptococci. Rom. Biotechnol. Lett. 2010, 15, 5788–5792. [Google Scholar]
- Turchany, J.M.; Aley, S.B.; Gillin, F.D. Giardicidal activity of lactoferrin and N-terminal peptides. Infect. Immun. 1995, 63, 4550–4552. [Google Scholar] [PubMed]
- Ochoa, T.J.; Woo, E.W.; Campos, M.; Pecho, M.; Prada, A.; McMahon, R.J.; Cleary, T.G. Impact of lactoferrin supplementation on growth and prevalence of giardia colonization in children. Clin. Infect. Dis. 2008, 46, 1881–1883. [Google Scholar] [PubMed]
- Elbarbary, H.A.; Abdou, A.M.; Park, E.Y.; Nakamura, Y.; Mohamed, H.A.; Sato, K. Novel antibacterial lactoferrin peptides generated by rennet digestion and autofocusing technique. Int. Dairy J. 2010, 20, 646–651. [Google Scholar] [CrossRef]
- Oda, H.; Wakabayashi, H.; Yamauchi, K.; Sato, T.; Xiao, J.Z.; Abe, F.; Iwatsuki, K. Isolation of a bifidogenic peptide from the pepsin hydrolysate of bovine lactoferrin. Appl. Environ. Microbiol. 2013, 79, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K. Potent antimicrobial effects of the glycosylated lactoferrin. Food Preserv. Sci. 2002, 28, 243–246. [Google Scholar] [CrossRef]
- Wakabayashi, H.; Uchida, K.; Yamauchi, K.; Teraguchi, S.; Hayasawa, H.; Yamaguchi, H. Lactoferrin given in food facilitates dermatophytosis cure in guinea pig models. J. Antimicrob. Chemother. 2000, 46, 595–660. [Google Scholar] [CrossRef] [PubMed]
- Cintra, W.M.; Silva-Filho, F.C.; de Souza, W. The surface charge of toxoplasma gondii: A cytochemical and electrophoretic study. J. Submicrosc. Cytol. 1986, 18, 773–781. [Google Scholar] [PubMed]
- Tachezy, J.; Suchan, P.; Schrevel, J.; Kulda, J. The host-protein independent iron uptake by tritrichomonas foetus. Exp. Parasitol. 1998, 90, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Sugiyama, K.; Tanaka, T.; Tanaka, K.; Sekihara, H.; Shimotohno, K.; Kato, N. Lactoferrin markedly inhibits hepatitis c virus infection in cultured human hepatocytes. Biochem. Biophys. Res. Commun. 1998, 245, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.C.; Swart, P.J.; de Bethune, M.P.; Pauwels, R.; de Clercq, E.; The, T.H.; Meijer, D.K. Antiviral effects of plasma and milk proteins: Lactoferrin shows potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J. Infect. Dis. 1995, 172, 380–388. [Google Scholar] [PubMed]
- Roy, K.; Kanwar, R.K.; Kanwar, J.R. Targeting viral hepatitis using natural milk protein and traditional medicinal herbs. J. Clin. Cell. Immunol. 2012, 3. [Google Scholar] [CrossRef]
- Farnaud, S.; Evans, R.W. Lactoferrin—A multifunctional protein with antimicrobial properties. Mol. Immunol. 2003, 40, 395–405. [Google Scholar] [CrossRef]
- Abrink, M.; Larsson, E.; Gobl, A.; Hellman, L. Expression of lactoferrin in the kidney: Implications for innate immunity and iron metabolism. Kidney Int. 2000, 57, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.J.; Han, S.B.; Kang, J.S.; Yoon, Y.D.; Park, S.K.; Kim, H.M.; Yang, K.H.; Joe, C.O. Lactoferrin works as a new LPS-binding protein in inflammatory activation of macrophages. Int. Immunopharmacol. 2004, 4, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Elass-Rochard, E.; Roseanu, A.; Legrand, D.; Trif, M.; Salmon, V.; Motas, C.; Montreuil, J.; Spik, G. Lactoferrin-lipopolysaccharide interaction: Involvement of the 28–34 loop region of human lactoferrin in the high-affinity binding to Escherichia coli 055b5 lipopolysaccharide. Biochem. J. 1995, 312, 839–845. [Google Scholar] [PubMed]
- Legrand, D.; Pierce, A.; Elass, E.; Carpentier, M.; Mariller, C.; Mazurier, J. Lactoferrin Structure and Functions Bioactive Components of Milk; Bösze, Z., Ed.; Springer: New York, NY, USA, 2008; Volume 606, pp. 163–194. [Google Scholar]
- Gahr, M.; Speer, C.; Damerau, B.; Sawatzki, G. Influence of lactoferrin on the function of human polymorphonuclear leukocytes and monocytes. J. Leukoc. Biol. 1991, 49, 427–433. [Google Scholar] [PubMed]
- Damiens, E.; Mazurier, J.; El Yazidi, I.; Masson, M.; Duthille, I.; Spik, G.; Boilly-Marer, Y. Effects of human lactoferrin on nk cell cytotoxicity against haematopoietic and epithelial tumour cells. BBA-Mol. Cell Res. 1998, 1402, 277–287. [Google Scholar] [CrossRef]
- Dashper, S.G.; Pan, Y.; Veith, P.D.; Chen, Y.Y.; Toh, E.C.; Liu, S.W.; Cross, K.J.; Reynolds, E.C. Lactoferrin inhibits porphyromonas gingivalis proteinases and has sustained biofilm inhibitory activity. Antimicrob. Agents Chemother. 2012, 56, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Shimazaki, K.; Higuchi, H.; Nagahata, H. Antibacterial activity of bovine lactoferrin hydrolysate against mastitis pathogens and its effect on superoxide production of bovine neutrophils. Zoonoses Public Health 2007, 54, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, P.; Stolfi, I.; Messner, H.; Cattani, S.; Laforgia, N.; Romeo, M.G.; Bollani, L.; Rinaldi, M.; Gallo, E.; Quercia, M.; et al. Bovine lactoferrin prevents invasive fungal infections in very low birth weight infants: A randomized controlled trial. Pediatrics 2012, 129, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Malet, A.; Bournaud, E.; Lan, A.; Mikogami, T.; Tome, D.; Blais, A. Bovine lactoferrin improves bone status of ovariectomized mice via immune function modulation. Bone 2011, 48, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M. Oxidative stress and vascular damage in hypertension. Curr. Hypertens. Rep. 2000, 2, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zheng, Y.; Fan, S.; Qin, Z.; Li, N.; Tang, A.; Ai, F.; Zhang, X.; Bian, Y.; Dang, W. Lactoferrin deficiency promotes colitis-associated colorectal dysplasia in mice. PLoS ONE 2014, 9, e103298. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.; Richmond, R.; Halliwell, B. Inhibition of the iron-catalysed formation of hydroxyl radicals from superoxide and of lipid peroxidation by desferrioxamine. Biochem. J. 1979, 184, 469–472. [Google Scholar] [PubMed]
- Raetz, C.R.; Ulevitch, R.J.; Wright, S.D.; Sibley, C.H.; Ding, A.; Nathan, C.F. Gram-negative endotoxin: An extraordinary lipid with profound effects on eukaryotic signal transduction. FASEB J. 1991, 5, 2652–2660. [Google Scholar] [PubMed]
- Pajkrt, D.; Doran, J.E.; Koster, F.; Lerch, P.G.; Arnet, B.; van der Poll, T.; ten Cate, J.W.; van Deventer, S.J. Antiinflammatory effects of reconstituted high-density lipoprotein during human endotoxemia. J. Exp. Med. 1996, 184, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Zagulski, T.; Lipinski, P.; Zagulska, A.; Broniek, S.; Jarzabek, Z. Lactoferrin can protect mice against a lethal dose of Escherichia coli in experimental infection in vivo. Br. J. Exp. Pathol. 1989, 70, 697–704. [Google Scholar] [PubMed]
- Zagulski, T.; Lipinski, P.; Zagulska, A.; Jarzabek, Z. Antibacterial system generated by lactoferrin in mice in vivo is primarily a killing system. Int. J. Exp. Pathol. 1998, 79, 117–123. [Google Scholar] [PubMed]
- Foster, T.J. Immune evasion by staphylococci. Nat. Rev. Microbiol. 2005, 3, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Holtfreter, S.; Broker, B.M. Staphylococcal superantigens: Do they play a role in sepsis? Arch. Immunol. Ther. Exp. 2005, 53, 13–27. [Google Scholar]
- Ellison, R. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Elass, E.; Carpentier, M.; Mazurier, J. Lactoferrin: A modulator of immune and inflammatory responses. Cell. Mol. Life Sci. 2005, 62, 2549–2559. [Google Scholar] [CrossRef] [PubMed]
- Kruzel, M.L.; Harari, Y.; Mailman, D.; Actor, J.K.; Zimecki, M. Differential effects of prophylactic, concurrent and therapeutic lactoferrin treatment on LPS-induced inflammatory responses in mice. Clin. Exp. Immunol. 2002, 130, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Baynes, R.D.; Bezwoda, W.R. Lactoferrin and the inflammatory response. Adv. Exp. Med. Biol. 1994, 357, 133–141. [Google Scholar] [PubMed]
- Shau, H.; Kim, A.; Golub, S.H. Modulation of natural killer and lymphokine-activated killer cell cytotoxicity by lactoferrin. J. Leukoc. Biol. 1992, 51, 343–349. [Google Scholar] [PubMed]
- Miyauchi, H.; Hashimoto, S.; Nakajima, M.; Shinoda, I.; Fukuwatari, Y.; Hayasawa, H. Bovine lactoferrin stimulates the phagocytic activity of human neutrophils: Identification of its active domain. Cell. Immunol. 1998, 187, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Takakura, N.; Teraguchi, S.; Tamura, Y. Lactoferrin feeding augments peritoneal macrophage activities in mice intraperitoneally injected with inactivated candida albicans. Microbiol. Immunol. 2003, 47, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.A.; Lopez, V.; Lonnerdal, B. Mammalian lactoferrin receptors: Structure and function. Cell. Mol. Life Sci. 2005, 62, 2560–2575. [Google Scholar] [CrossRef] [PubMed]
- Hammarstrom, M.L.; Mincheva-Nilsson, L.; Hammarstrom, S. Functional lactoferrin receptors on activated human lymphocytes. Adv. Exp. Med. Biol. 1995, 371A, 47–53. [Google Scholar]
- Sorimachi, K.; Akimoto, K.; Hattori, Y.; Ieiri, T.; Niwa, A. Activation of macrophages by lactoferrin: Secretion of tnf-alpha, il-8 and no. Biochem. Mol. Biol. Int. 1997, 43, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Wlaszczyk, A.; Wojciechowski, R.; Dawiskiba, J.; Kruzel, M. Lactoferrin regulates the immune responses in post-surgical patients. Arch. Immunol. Ther. Exp. 2001, 49, 325–333. [Google Scholar]
- Machnicki, M.; Zimecki, M.; Zagulski, T. Lactoferrin regulates the release of tumour necrosis factor alpha and interleukin 6 in vivo. Int. J. Exp. Pathol. 1993, 74, 433–439. [Google Scholar] [PubMed]
- Puddu, P.; Valenti, P.; Gessani, S. Immunomodulatory effects of lactoferrin on antigen presenting cells. Biochimie 2009, 91, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Tanida, T.; Rao, F.; Hamada, T.; Ueta, E.; Osaki, T. Lactoferrin peptide increases the survival of candida albicans-inoculated mice by upregulating neutrophil and macrophage functions, especially in combination with amphotericin b and granulocyte-macrophage colony-stimulating factor. Infect. Immun. 2001, 69, 3883–3890. [Google Scholar] [CrossRef] [PubMed]
- Cumberbatch, M.; Dearman, R.J.; Uribe-Luna, S.; Headon, D.R.; Ward, P.P.; Conneely, O.M.; Kimber, I. Regulation of epidermal langerhans cell migration by lactoferrin. Immunology 2000, 100, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Cumberbatch, M.; Bhushan, M.; Dearman, R.J.; Kimber, I.; Griffiths, C.E. IL-1beta-induced langerhans’ cell migration and tnf-alpha production in human skin: Regulation by lactoferrin. Clin. Exp. Immunol. 2003, 132, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, T.L.; Bakman, S.; Sorensen, E.S.; Barkholt, V.; Frokiaer, H. Sialic acid-containing milk proteins show differential immunomodulatory activities independent of sialic acid. J. Agric. Food Chem. 2005, 53, 7673–7680. [Google Scholar] [CrossRef] [PubMed]
- Zimecki, M.; Dawiskiba, J.; Zawirska, B.; Krawczyk, Z.; Kruzel, M. Bovine lactoferrin decreases histopathological changes in the liver and regulates cytokine production by splenocytes of obstructive jaundiced rats. Inflamm. Res. 2003, 52, 305–310. [Google Scholar] [PubMed]
- Mincheva-Nilsson, L.; Hammarstrom, S.; Hammarstrom, M.L. Activated human gamma delta t lymphocytes express functional lactoferrin receptors. Scand. J. Immunol. 1997, 46, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Morita, H.; Yoo, Y.C.; Kim, W.S.; Kumura, H.; Shimazaki, K. Detection of bovine lactoferrin binding protein on jurkat human lymphoblastic T cell line. J. Vet. Med. Sci. 2004, 66, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; van Berkel, P.H.; Salmon, V.; van Veen, H.A.; Slomianny, M.C.; Nuijens, J.H.; Spik, G. The N-terminal arg2, arg3 and arg4 of human lactoferrin interact with sulphated molecules but not with the receptor present on jurkat human lymphoblastic T-cells. Biochem. J. 1997, 327 Pt 3, 841–846. [Google Scholar] [PubMed]
- Zimecki, M.; Mazurier, J.; Machnicki, M.; Wieczorek, Z.; Montreuil, J.; Spik, G. Immunostimulatory activity of lactotransferrin and maturation of CD4- CD8- murine thymocytes. Immunol. Lett. 1991, 30, 119–123. [Google Scholar] [CrossRef]
- Frydecka, I.; Zimecki, M.; Bocko, D.; Kosmaczewska, A.; Teodorowska, R.; Ciszak, L.; Kruzel, M.; Wlodarska-Polinsk, J.; Kuliczkowski, K.; Kornafel, J. Lactoferrin-induced up-regulation of zeta (zeta) chain expression in peripheral blood T lymphocytes from cervical cancer patients. Anticancer Res. 2002, 22, 1897–1901. [Google Scholar] [PubMed]
- Zimecki, M.; Miedzybrodzki, R.; Mazurier, J.; Spik, G. Regulatory effects of lactoferrin and lipopolysaccharide on lfa-1 expression on human peripheral blood mononuclear cells. Arch. Immunol. Ther. Exp. 1999, 47, 257–264. [Google Scholar]
- Sfeir, R.M.; Dubarry, M.; Boyaka, P.N.; Rautureau, M.; Tomé, D. The mode of oral bovine lactoferrin administration influences mucosal and systemic immune responses in mice. J. Nutr. 2004, 134, 403–409. [Google Scholar] [PubMed]
- Wang, W.-P.; Iigo, M.; Sato, J.; Sekine, K.; Adachi, I.; Tsuda, H. Activation of intestinal mucosal immunity in tumor-bearing mice by lactoferrin. Cancer Sci. 2000, 91, 1022–1027. [Google Scholar] [CrossRef]
- Hashiguchi, K.; Takeshima, F.; Akazawa, Y.; Matsushima, K.; Minami, H.; Machida, H.; Yamaguchi, N.; Shiozawa, K.; Ohba, K.; Ohnita, K. Advantages of fecal lactoferrin measurement during granulocyte and monocyte adsorptive apheresis therapy in ulcerative colitis. Digestion 2015, 91, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Furlund, C.; Ulleberg, E.; Devold, T.; Flengsrud, R.; Jacobsen, M.; Sekse, C.; Holm, H.; Vegarud, G. Identification of lactoferrin peptides generated by digestion with human gastrointestinal enzymes. J. Dairy Sci. 2013, 96, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Troost, F.J.; Steijns, J.; Saris, W.H.; Brummer, R.J.M. Gastric digestion of bovine lactoferrin in vivo in adults. J. Nutr. 2001, 131, 2101–2104. [Google Scholar] [PubMed]
- Liu, Y.; Miyoshi, H.; Nakamura, M. Nanomedicine for drug delivery and imaging: A promising avenue for cancer therapy and diagnosis using targeted functional nanoparticles. Int. J. Cancer 2007, 120, 2527–2537. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, E.S.; Player, A. Nanotechnology, nanomedicine, and the development of new, effective therapies for cancer. Nanomedicine 2005, 1, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Lü, J.M.; Wang, X.; Marin-Muller, C.; Wang, H.; Lin, P.H.; Yao, Q.; Chen, C. Current advances in research and clinical applications of plga-based nanotechnology. Expert Rev. Mol. Diagn. 2009, 9, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.M.; Rubinstein, I.; Onyuksel, H. Role of nanotechnology in targeted drug delivery and imaging: A concise review. Nanomedicine 2005, 1, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Shapira, A.; Livney, Y.D.; Broxterman, H.J.; Assaraf, Y.G. Nanomedicine for targeted cancer therapy: Towards the overcoming of drug resistance. Drug Resist. Updates 2011, 14, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Nan, A.; Line, B.R.; Ghandehari, H. Nanocarriers for nuclear imaging and radiotherapy of cancer. Curr. Pharm. Des. 2006, 12, 4729–4749. [Google Scholar] [CrossRef] [PubMed]
- Zrazhevskiy, P.; Sena, M.; Gao, X. Designing multifunctional quantum dots for bioimaging, detection, and drug delivery. Chem. Soc. Rev. 2010, 39, 4326–4354. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Lee, S.; Chen, X. Nanoparticle-based theranostic agents. Adv. Drug Deliv. Rev. 2010, 62, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Yoncheva, K.; Momekov, G. Antiangiogenic anticancer strategy based on nanoparticulate systems. Expert Opin. Drug Deliv. 2011, 8, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Mamot, C.; Drummond, D.C.; Greiser, U.; Hong, K.; Kirpotin, D.B.; Marks, J.D.; Park, J.W. Epidermal growth factor receptor (egfr)-targeted immunoliposomes mediate specific and efficient drug delivery to egfr- and egfrviii-overexpressing tumor cells. Cancer Res. 2003, 63, 3154–3161. [Google Scholar] [PubMed]
- Pan, X.; Lee, R.J. Construction of anti-egfr immunoliposomes via folate-folate binding protein affinity. Int. J. Pharm. 2007, 336, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.Y.; Kim, I.S.; Kwon, I.C.; Kim, Y.H. Tumor targetability and antitumor effect of docetaxel-loaded hydrophobically modified glycol chitosan nanoparticles. J. Control. Release 2008, 128, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Benny, O.; Fainaru, O.; Adini, A.; Cassiola, F.; Bazinet, L.; Adini, I.; Pravda, E.; Nahmias, Y.; Koirala, S.; Corfas, G.; et al. An orally delivered small-molecule formulation with antiangiogenic and anticancer activity. Nat. Biotechnol. 2008, 26, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, Y.S.; Park, K.; Kang, E.; Lee, S.; Nam, H.Y.; Kim, K.; Park, J.H.; Chi, D.Y.; Park, R.W.; et al. Self-assembled glycol chitosan nanoparticles for the sustained and prolonged delivery of antiangiogenic small peptide drugs in cancer therapy. Biomaterials 2008, 29, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, M.; Guo, R.; Jiang, Z.; Liu, Y.; Jiang, X.; Yang, C. Chitosan surface-modified hydroxycamptothecin loaded nanoparticles with enhanced transport across caco-2 cell monolayer. J. Nanosci. Nanotechnol. 2006, 6, 2912–2920. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, P.; Lee, B.J.; Oh, D.H.; Kim, J.O.; Lee, Y.I.; Kim, D.D.; Jee, J.P.; Lee, Y.B.; Woo, J.S.; Yong, C.S.; et al. Enhanced oral bioavailability of coenzyme Q10 by self-emulsifying drug delivery systems. Int. J. Pharm. 2009, 374, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Li, S. Hydroxycamptothecin-loaded nanoparticles enhance target drug delivery and anticancer effect. BMC Biotechnol. 2008, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Howard, K.A.; Paludan, S.R.; Behlke, M.A.; Besenbacher, F.; Deleuran, B.; Kjems, J. Chitosan/sirna nanoparticle-mediated tnf-[alpha] knockdown in peritoneal macrophages for anti-inflammatory treatment in a murine arthritis model. Mol. Ther. 2008, 17, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; van Vlerken, L.; Little, S.; Amiji, M. Evaluations of combination mdr-1 gene silencing and paclitaxel administration in biodegradable polymeric nanoparticle formulations to overcome multidrug resistance in cancer cells. Cancer Chemother. Pharmacol. 2009, 63, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lee, S.; Gan, C.; Feng, S.S. In vitro and in vivo investigation on pla–tpgs nanoparticles for controlled and sustained small molecule chemotherapy. Pharm. Res. 2008, 25, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.S.; Mei, L.; Anitha, P.; Gan, C.W.; Zhou, W. Poly(lactide)-vitamin E derivative/montmorillonite nanoparticle formulations for the oral delivery of docetaxel. Biomaterials 2009, 30, 3297–3306. [Google Scholar] [CrossRef] [PubMed]
- Peinado, I.; Lesmes, U.; Andrés, A.; McClements, J.D. Fabrication and morphological characterization of biopolymer particles formed by electrostatic complexation of heat treated lactoferrin and anionic polysaccharides. Langmuir 2010, 26, 9827–9834. [Google Scholar] [CrossRef] [PubMed]
- Bengoechea, C.; Jones, O.G.; Guerrero, A.; McClements, D.J. Formation and characterization of lactoferrin/pectin electrostatic complexes: Impact of composition, pH and thermal treatment. Food Hydrocolloids 2011, 25, 1227–1232. [Google Scholar] [CrossRef]
- David-Birman, T.; Mackie, A.; Lesmes, U. Impact of dietary fibers on the properties and proteolytic digestibility of lactoferrin nano-particles. Food Hydrocoll. 2013, 31, 33–41. [Google Scholar] [CrossRef]
- Bengoechea, C.; Peinado, I.; McClements, D.J. Formation of protein nanoparticles by controlled heat treatment of lactoferrin: Factors affecting particle characteristics. Food Hydrocoll. 2011, 25, 1354–1360. [Google Scholar] [CrossRef]
- Shimoni, G.; Levi, C.S.; Tal, S.L.; Lesmes, U. Emulsions stabilization by lactoferrin nano-particles under in vitro digestion conditions. Food Hydrocoll. 2013, 33, 264–272. [Google Scholar] [CrossRef]
- Duarte, D.C.; Nicolau, A.; Teixeira, J.A.; Rodrigues, L.R. The effect of bovine milk lactoferrin on human breast cancer cell lines. J. Dairy Sci. 2011, 94, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Paliwal, S.R.; Paliwal, R.; Agrawal, G.P.; Vyas, S.P. Liposomal nanomedicine for breast cancer therapy. Nanomedicine 2011, 6, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Pamujula, S.; Hazari, S.; Bolden, G.; Graves, R.A.; Chinta, D.D.; Dash, S.; Kishore, V.; Mandal, T.K. Cellular delivery of pegylated plga nanoparticles. J Pharm. Pharmacol. 2012, 64, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, S.; Verderio, P.; Sommaruga, S.; Colombo, M.; Salvadè, A.; Corsi, F.; Galeffi, P.; Tortora, P.; Prosperi, D. Multiple presentation of scfv800e6 on silica nanospheres enhances targeting efficiency toward her-2 receptor in breast cancer cells. Bioconj. Chem. 2011, 22, 2296–2303. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Kozu, T.; Iinuma, G.; Ohashi, Y.; Saito, Y.; Saito, D.; Akasu, T.; Alexander, D.; Futakuchi, M.; Fukamachi, K.; et al. Cancer prevention by bovine lactoferrin: From animal studies to human trial. Biometals 2010, 23, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Banno, Y.; Kato, Y.; Nozawa, Y.; Kawaguchi, M. Pepsin-digested bovine lactoferrin induces apoptotic cell death with jnk/sapk activation in oral cancer cells. J. Pharmacol. Sci. 2005, 98, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Sekine, K.; Fujita, K.I.; Iigo, M. Cancer prevention by bovine lactoferrin and underlying mechanisms—A review of experimental and clinical studies. Biochem. Cell Biol. 2002, 80, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Li, J.; Shen, Y.; Lu, W.; Gao, X.; Zhang, Q.; Jiang, X. Lactoferrin-conjugated PEG–PLA nanoparticles with improved brain delivery: In vitro and in vivo evaluations. J. Control. Release 2009, 134, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ke, W.; Liu, Y.; Jiang, C.; Pei, Y. The use of lactoferrin as a ligand for targeting the polyamidoamine-based gene delivery system to the brain. Biomaterials 2008, 29, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Zhu, Y.; Jiang, W.; Zhou, Q.; Yang, H.; Gu, N.; Zhang, Y.; Xu, H.; Xu, H.; Yang, X. Lactoferrin-conjugated superparamagnetic iron oxide nanoparticles as a specific MRI contrast agent for detection of brain glioma in vivo. Biomaterials 2010, 32, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Feng, L.; Hua, R.; Chen, J.; Gao, H.; Pan, S.; Jiang, X.; Zhang, P. Lactoferrin-conjugated biodegradable polymersome holding doxorubicin and tetrandrine for chemotherapy of glioma rats. Mol. Pharm. 2010, 7, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qin, Y.; Zhang, Q.; Jiang, W.; Tang, L.; Liu, J.; He, Q. Lactoferrin modified doxorubicin-loaded procationic liposomes for the treatment of gliomas. Eur. J. Pharm. Sci. 2011, 44, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Elfinger, M.; Maucksch, C.; Rudolph, C. Characterization of lactoferrin as a targeting ligand for nonviral gene delivery to airway epithelial cells. Biomaterials 2007, 28, 3448–3455. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ke, W.; Liu, Y.; Wu, D.; Feng, L.; Jiang, C.; Pei, Y. Gene therapy using lactoferrin-modified nanoparticles in a rotenone-induced chronic parkinson model. J. Neurol. Sci. 2010, 290, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H.; Koyama, K.; Sakata, O.; Machida, Y. Preparation of chitosan/alginate/calcium complex microparticles loaded with lactoferrin and their efficacy on carrageenan-induced edema in rats. Drug Dev. Ind. Pharm. 2010, 36, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ke, W.; Han, L.; Liu, Y.; Shao, K.; Ye, L.; Lou, J.; Jiang, C.; Pei, Y. Brain-targeting mechanisms of lactoferrin-modified DNA-loaded nanoparticles. J. Cereb. Blood Flow Metabol. 2009, 29, 1914–1923. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Mahidhara, G.; Kanwar, R.K. Novel alginate-enclosed chitosan-calcium phosphate-loaded iron-saturated bovine lactoferrin nanocarriers for oral delivery in colon cancer therapy. Nanomedicine 2012, 7, 1521–1550. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Gajbhiye, K.R.; Soni, V. Lactoferrin-appended solid lipid nanoparticles of paclitaxel for effective management of bronchogenic carcinoma. Drug Deliv. 2015, 22, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Chen, Y.C. Targeting delivery of etoposide to inhibit the growth of human glioblastoma multiforme using lactoferrin-and folic acid-grafted poly (lactide-co-glycolide) nanoparticles. Inter. J. Pharm. 2015, 479, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Amini, A.A.; Kan, H.M.; Cui, Z.; Maye, P.; Nair, L.S. Enzymatically cross-linked bovine lactoferrin as injectable hydrogel for cell delivery. Tissue Eng. Part A 2014, 20, 2830–2839. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Wakabayashi, H.; Shin, K.; Yamauchi, K.; Yaeshima, T.; Iwatsuki, K. Twenty-five years of research on bovine lactoferrin applications. Biochimie 2009, 91, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Paesano, R.; Berlutti, F.; Pietropaoli, M.; Goolsbee, W.; Pacifici, E.; Valenti, P. Lactoferrin efficacy versus sulphate in curing iron disorders in pregnant and non-pregnant women. Int. J. Immunopathol. Pharmacol. 2010, 23, 577–587. [Google Scholar] [PubMed]
- Tanaka, M.S.; Tojima, T.; Dousako, S.; Tatsumi, K. Method of Manufacturing Iron-Fortified Beverage. U.S. Patent 509,872, 24 March 1992. [Google Scholar]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanwar, J.R.; Roy, K.; Patel, Y.; Zhou, S.-F.; Singh, M.R.; Singh, D.; Nasir, M.; Sehgal, R.; Sehgal, A.; Singh, R.S.; et al. Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions. Molecules 2015, 20, 9703-9731. https://doi.org/10.3390/molecules20069703
Kanwar JR, Roy K, Patel Y, Zhou S-F, Singh MR, Singh D, Nasir M, Sehgal R, Sehgal A, Singh RS, et al. Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions. Molecules. 2015; 20(6):9703-9731. https://doi.org/10.3390/molecules20069703
Chicago/Turabian StyleKanwar, Jagat R., Kislay Roy, Yogesh Patel, Shu-Feng Zhou, Manju Rawat Singh, Deependra Singh, Muhammad Nasir, Rakesh Sehgal, Alka Sehgal, Ram Sarup Singh, and et al. 2015. "Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions" Molecules 20, no. 6: 9703-9731. https://doi.org/10.3390/molecules20069703
APA StyleKanwar, J. R., Roy, K., Patel, Y., Zhou, S.-F., Singh, M. R., Singh, D., Nasir, M., Sehgal, R., Sehgal, A., Singh, R. S., Garg, S., & Kanwar, R. K. (2015). Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions. Molecules, 20(6), 9703-9731. https://doi.org/10.3390/molecules20069703