Anti-Depressant-Like Effect of Kaempferitrin Isolated from Justicia spicigera Schltdl (Acanthaceae) in Two Behavior Models in Mice: Evidence for the Involvement of the Serotonergic System

Abstract
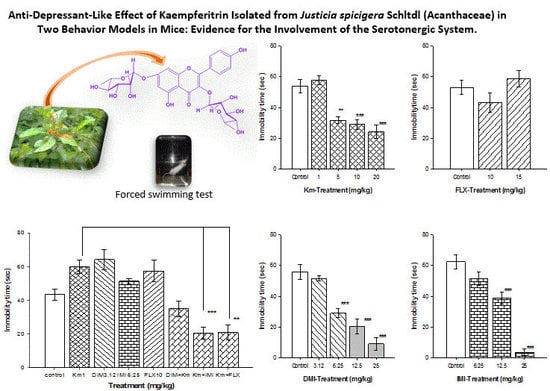
:
1. Introduction
2. Results and Discussion
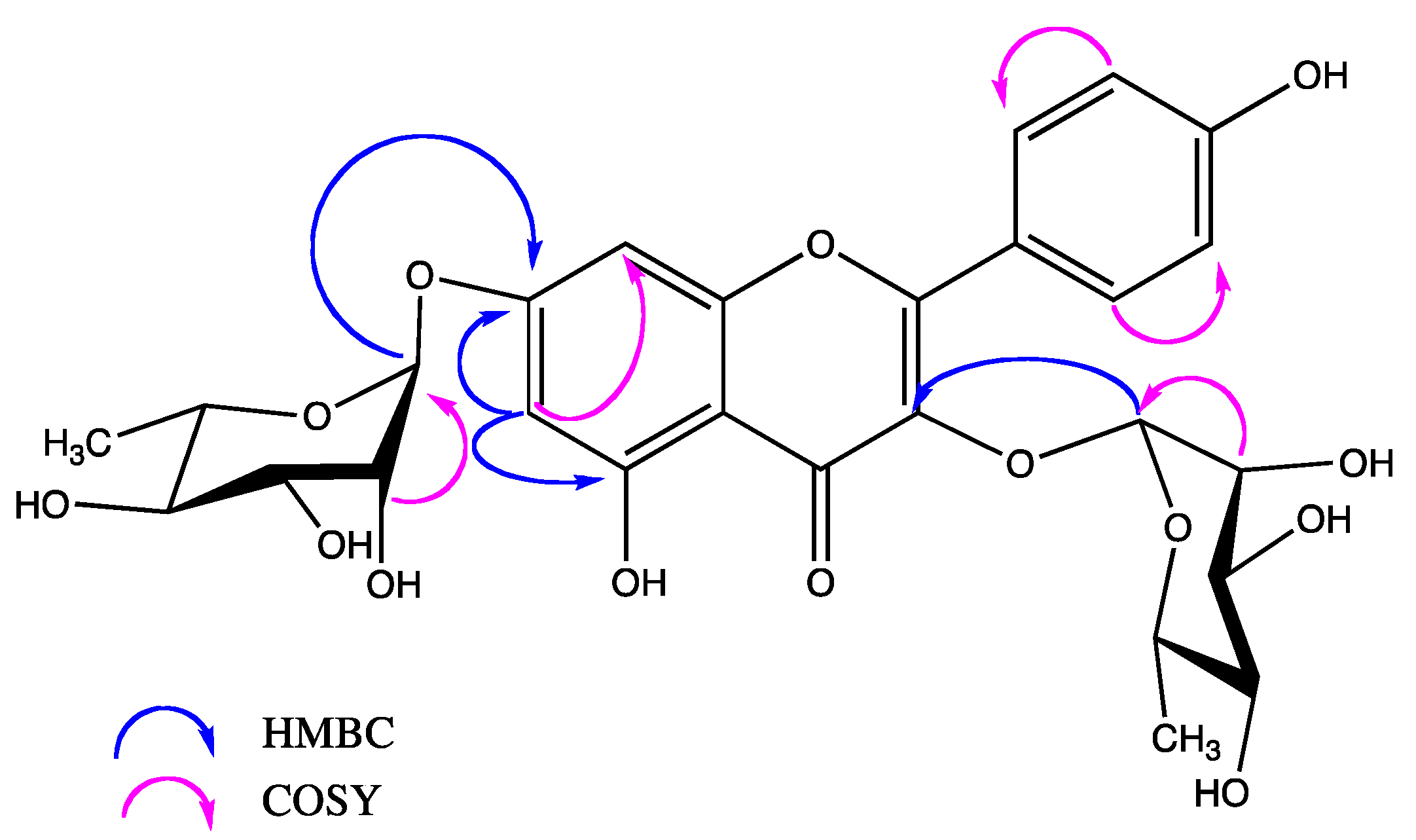
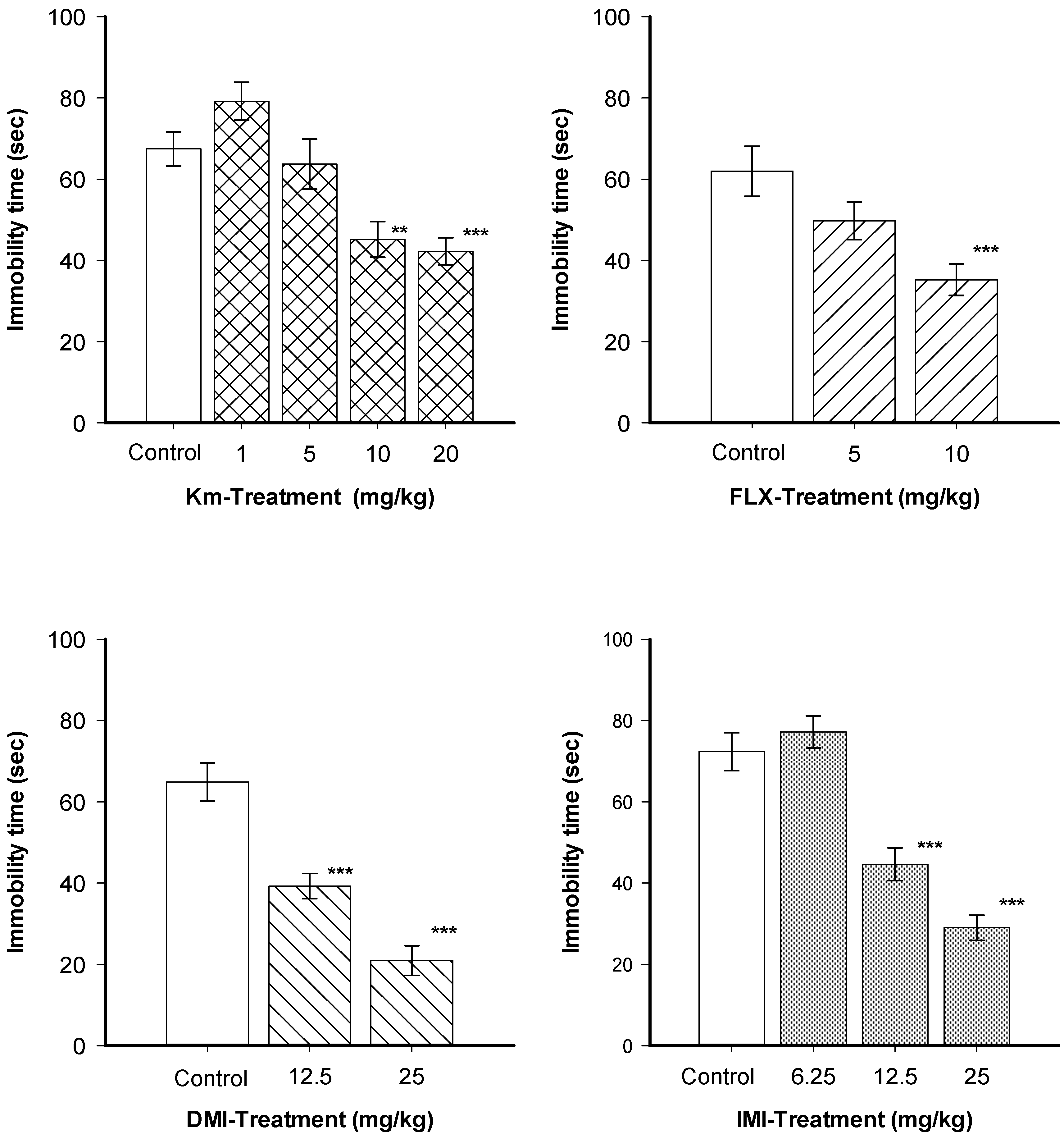
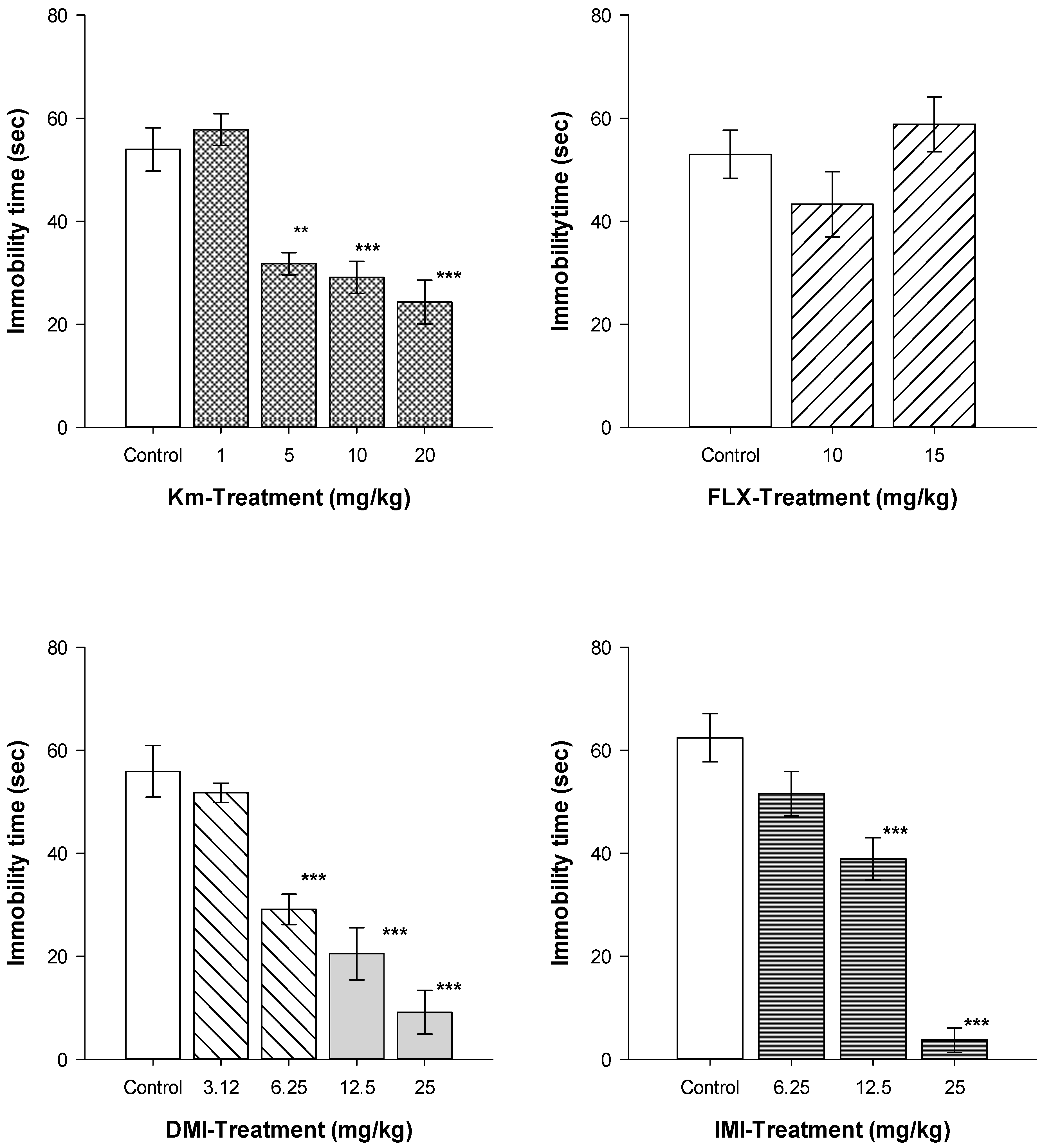

{kind=link}
{kind=link}
{kind=link}
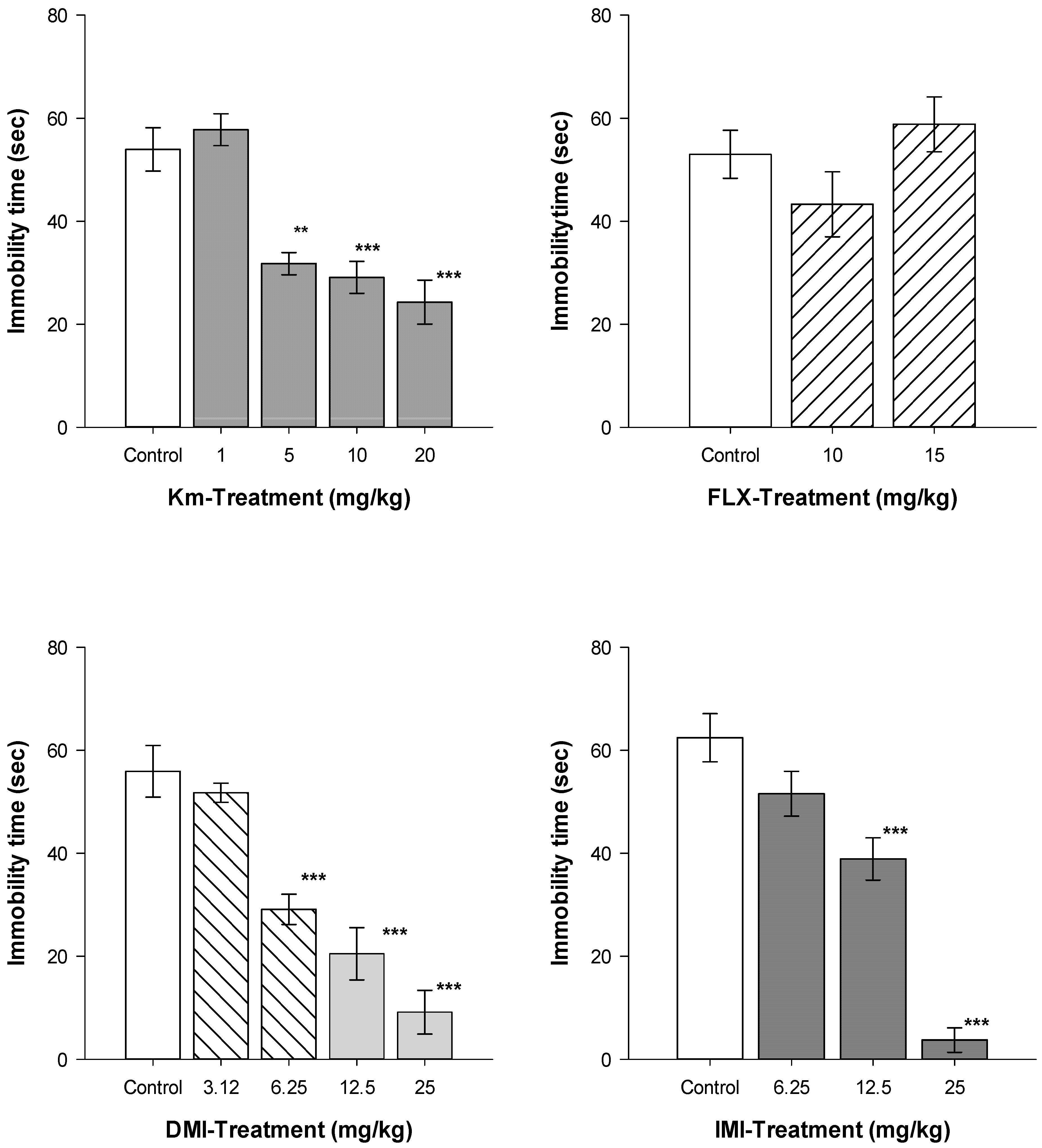{kind=link}
{kind=link}
| Treatment (mg/kg) | Ambulatory Activity (Count Number/5 min) | Rearing Number/5 min |
|---|---|---|
| Control | 31.7 ± 3.24 | 17.6 ± 1.46 |
| Km1 | 35.0 ± 3.31 | 13.0 ± 3.40 |
| Km5 | 29.0 ± 3.48 | 20.5 ± 3.90 |
| Km10 | 32.1 ± 3.64 | 13.8 ±3.23 |
| Km20 | 44.7 ± 0.2.29 * | 17.7 ± 2.80 |
| H = 10.72, df = 4, p = 0.030 | H = 3.18, df = 4, p = 0.527 | |
| Control | 41. 12 ±1.32 | 25.25 ± 3.63 |
| FLX10 | 37.00 ± 2.17 | 21.75 ± 3.37 |
| FLX 15 | 45.37 ± 3.13 | 28.12 ± 0.81 |
| H = 5.39, df = 2, p = 0.067 | H = 3.56, df = 2, p = 0.168 | |
| Control DMI 3.12 DMI 6.25 | 37.7 ± 3.24 35.50 ± 5.58 40.5 ± 6.33 | 29.3 ± 5.73 26.53 ± 3.82 26.8 ± 4.20 |
| DMI 12.5 | 47.8 ± 3.92 | 28.0 ± 3.53 |
| DMI25 | 40.1 ± 4.76 | 18.7 ± 5.26 |
| H = 6.08, df = 4, p = 0.193 | H = 4.97, df = 4, p = 0.06 | |
| Control | 38.8 ± 1.29 | 25.2 ± 3.63 |
| IMI 6.25 | 44.5 ± 6.04 | 36.5 ± 4.67 |
| 12.5 | 30.2 ± 4.43 | 27.5 ± 2.77 |
| 25 | 41.12 ± 6.18 | 34.5 ±7.72 |
| H = 3.508, df = 3, p = 0.320 | H = 7.38, df = 3, p = 0.061 |
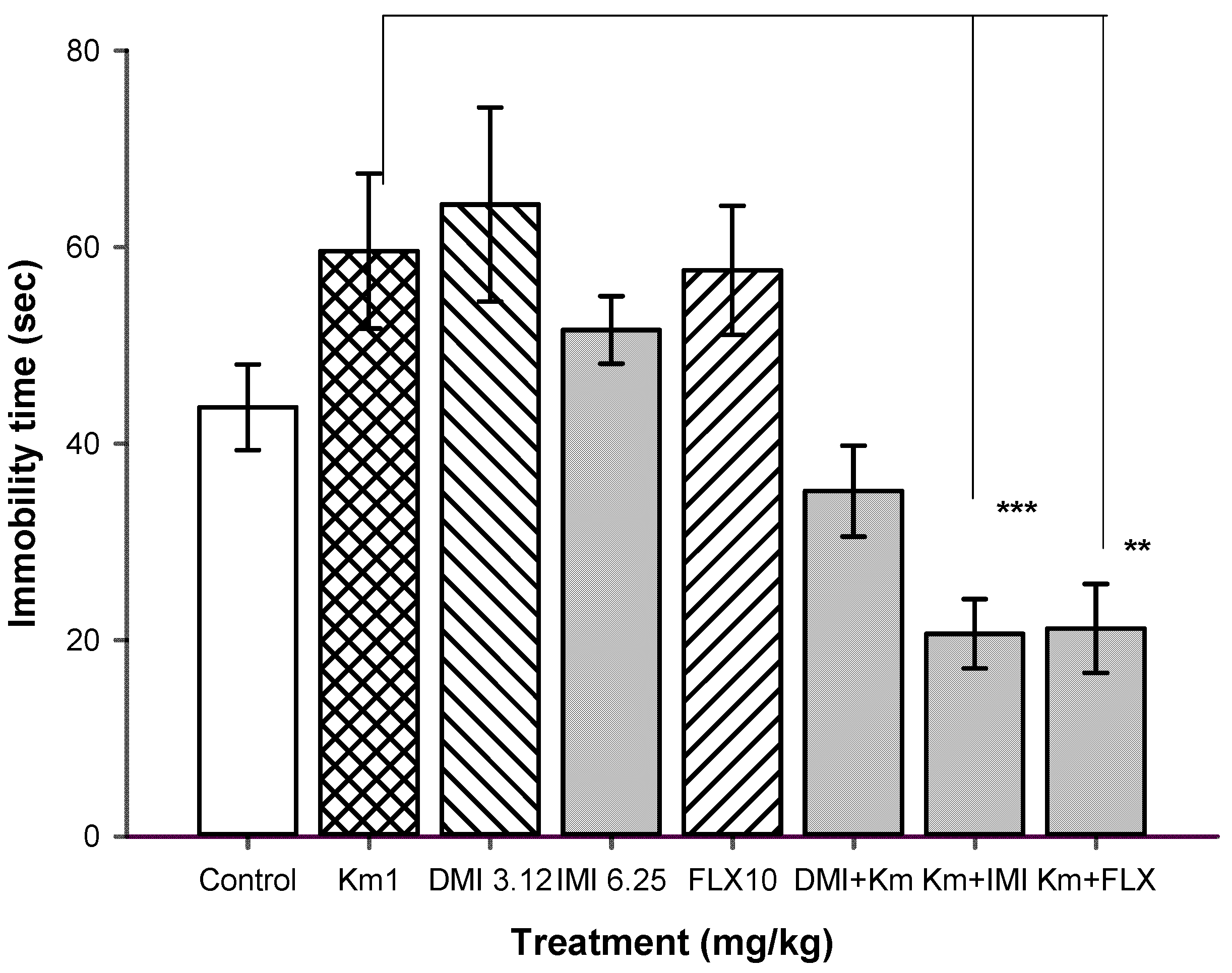
| Treatment 1 (mg/kg) | plus | Treatment 2 (mg/kg) | Immobility Time (sec) |
|---|---|---|---|
| Experiment I | |||
| Vehicle | vehicle | 57.93 ± 2.02 | |
| Vehicle | Km (10) | 27.73 ± 3.62 ** | |
| Pcpa (100) | vehicle | 50.36 ± 4.52 | |
| Pcpa (100) | Km(10) | 51.26 ± 6.24 | |
| H = 10.246, df = 3, p = 0.006 | |||
| Experiment II | |||
| Vehicle | vehicle | 57.93 ± 2.02 | |
| Pindolol (10) | Vehicle | 59.86 ± 7.83 ### | |
| Vehicle | Km (10) | 29.11 ± 3.08 *** | |
| Km (10) | Pindolol (10) | 13.10 ± 2.42 ***, ### | |
| H = 25.77, df = 3, p ≤ 0.001 | |||
| Experiment III | |||
| Vehicle | vehicle | 52.93 ± 2.02 | |
| Way100635 (0.03) | vehicle | 58.03 ± 5.49 | |
| Vehicle | Km (10) | 29.11 ± 3.08 *** | |
| Way(0.03) | Km(10) | 54.24 ± 6.85 | |
| H = 14.02, df = 3, p = 0.003 | |||
| Experiment IV | |||
| Vehicle | vehicle | 58.13 ± 2.31 | |
| Vehicle | Km (1) | 60.07 ± 3.82 | |
| 8OH-DPAT (0.05) | vehicle | 60.85 ± 3.85 ### | |
| 8OH-DPAT (0.5) | vehicle | 30.78 ± 3.48 *** | |
| 8OH-DPAT (0.05) | Km (1) | 34.91 ± 3.48 ***, ### | |
| H = 30.59, df = 5, p = 0.001 | |||
| Treatment (mg/kg) | Corticosterone Levels (nmol/L) |
|---|---|
| Naïve | 495.42 ± 58.98 |
| Saline solution | 891.42 ± 141.13 *** |
| Km 10 | 1000.52 ± 38.42 *** |
| Km 20 | 1271.51 ± 200.91 *** |
| IMI 25 | 925.25 ± 128.59*** |
| F(4,24) = 4.96, p = 0.05 |
3. Experimental
3.1. General Information
3.2. Plant Material
3.3. Isolation, Purification and Chemical Structure Determination of Kaempferitrin
3.4. Animals
3.5. Drugs
3.6. Tail Suspension Test (TST)
3.7. Forced Swimming Test (FST)
3.8. Combination of Km with IMI, DMI, or FLX in the FST
3.9. Combination of Km with PCPA, WAY 100635, Pindolol, or 8OH-DPAT in the FST
3.10. Open Field Test (OFT)
3.11. Corticosterone Levels Determination
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wong, M.L.; Lucino, J. Research and treatment approaches to depression. Nat. Rev. Neurosci. 2001, 2, 343–351. [Google Scholar] [PubMed]
- Wong, M.L.; Lucino, J. From monoamines to genomic targets: A paradigm shift for drug discovery in depression. Nat. Rev. Drugs Discov. 2004, 3, 136–153. [Google Scholar]
- Monroe, S.M.; Mark, W.; Reid, M.W. Mistreating Psychology in the Decades of the Brain. Perspect. Psychol. Sci. 2010, 1, 716–743. [Google Scholar]
- Medina-Mora, M.E.; Borges, G.; Benjet, C.; Lara, C.; Berglund, P. Psychiatric disorders in Mexico: Lifetime prevalence in a nationally representative sample. Br. J. Psychiatry 2007, 190, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Holmes, A. The ascent of mouse: Advances in modelling human depression and anxiety. Nat. Rev. Drug Discov. 2005, 4, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, M.; Chelminski, I.; McDermut, W. Major depressive disorder and axis I diagnostic comorbidity. J. Clin. Psychiatry 2002, 63, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Gorman, M.D.; Jack, M. Comorbid depression and anxiety spectrum disorders. Depress. Anxiety 1997, 4, 160–168. [Google Scholar] [CrossRef]
- Zhang, Z. Therapeutic effects of herbal extracts and constituents in animal models of psychiatric disorders. Life Sci. 2004, 75, 1659–1699. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.G.; Kaster, M.P.; Binfaré, R.W.; Dias, M.; Santos, A.R.S.; Pizzolatti, M.G.; Brighentee, I.M.; Rodrigues, A.L.S. Antidepressant-like effect of the extract from leaves of Schinus molle L. in mice: Evidence for the involvement of the monoaminergic system. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Yi, L.T.; Pan, Y.; Wang, X.; Li, Y.C.; Li, J.M.; Kong, L.D. Antidepressant-like effects of the mixture of honokiol and magnolol from the barks of Magnolia officinalis in stressed rodents. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 715–725. [Google Scholar] [PubMed]
- Ji, H.-F.; Li, X.-J.; Zhang, H.-Y. Natural products and drug discovery. EMBO Rep. 2009, 10, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M. Las Plantas Medicinales de México; Editorial Botas: Mexico City, Mexico, 2005; Volumen I, pp. 225–226. [Google Scholar]
- Martinez, A.M.A. Medicinal plants used in a totonac community of the sierra norte de puebla: Tuzamapan de gale ana, puebla, Mexico. J. Ethnopharmacol. 1984, 11, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Alva Real, G.; Laboratorio de Fitofarmacología, Dirección de Investigaciones en Neurociencias, INPRFM, Mexcio, D.F. Mexico. Personal communication as a result of her ethnobotanical field research, 2008.
- Ortiz-Andrade, R.; Cabañas-Wuan, A.; Arana-Argaez, V.E.; Alonso-Castro, A.J.; Zapata-Bustos, R.; Salazar-Olivo, L.A.; Dominguez, F.; Chavez, M.; Carranza-Alvarez, C.; Garcia-Carrancá, A. Antidiabetic effects of Justicia spicigera Schltdl (Acanthaceae). J. Ethnopharmacol. 2012, 143, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, G.M.; de C. Alcântara, A.F. Chemical constituents and biological activities of species of Justicia—A review. Rev. Bras. Farmacogn. Braz. J. Pharmacogn. 2012, 22, 220–238. [Google Scholar] [CrossRef]
- Fernández, L.; Mas, R.; Pérez, H.; Biscay, R.T.; Galan, L. Evaluación preliminar de los efectos neurofarmacologicos de J. pectoralis. Rev. Cuba. Farmacol. 1989, 23, 161–166. [Google Scholar]
- Domínguez, X.A.; Achenbach, H.; González, C.; Ferré-D´Amore, A. Estudio químico del “muicle” (Justicia spicigera). Rev. Quím. Latinoam. 1990, 21, 142–143. [Google Scholar]
- Euler, K.L.; Alam, M. Isolation of Kaempferitrin from Justicia spicigera. J. Nat. Prod. 1982, 45, 220–221. [Google Scholar] [CrossRef]
- Fernández, S.; Wasowski, C.; Paladini, A.C.; Marder, M. Sedative and sleep-enhancing properties of linarin, a flavonoid-isolated from Valeriana officinalis. Pharmacol. Biochem. Behav. 2004, 77, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.G.; Bettio, L.E.B.; Cunha, M.P.; Santos, A.R.S.; Pizzolatti, M.G.; Brighente, I.M.C.; Rodrigues, A.L.S. Antidepressant-like effect of rutin isolated from the ethanolic extract from Schinus molle L. in mice: Evidence for the involvement of the serotonergic and noradrenergic systems. Eur. J. Pharmacol. 2008, 587, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Cassani, J.; Araujo, A.G.; Martinez-Vazquez, M.; Manjarrez, N.; Moreno, J.; Estrada-Reyes, R. Anxiolytic-like and antinociceptive effects of 2(S)-neoponcirin in mice. Molecules 2013, 18, 7584–7599. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.C.; Fernández, S.P.; Loscalzo, L.M.; Wasowski, C.; Paladini, A.C.; Marder, M.; Medina, J.H.; Viola, H. Hesperidin, a flavonoid glycoside with sedative effect, decreases brain Perk1/2 levels in mice. Pharmacol. Biochem. Behav. 2009, 92, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Sagud, M.; Nela Pivac, N.; Mück-Seler, D.; Jakovljevic, M.M.; Mihaljevic-Peles, A.; KorsI, M. Effects of sertraline treatment on plasma cortisol, prolactin and thyroid hormones in female depressed patients. Neuropsychobiology 2002, 45, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Swaab, D.F.; Bao, A.-M.; Lucassen, P.J. The stress system in the human brain in depression and neurodegeneration. Ageing Res. Rev. 2005, 4, 141–194. [Google Scholar] [CrossRef] [PubMed]
- Groenink, L.; Dirks, A.; Verdouw, M.P.; Schipholt, M.I.; Veening, J.G.; Gugten, J.V.D.; Olivier, B. HPA Axis Dysregulation in Mice Overexpressing Corticotropin Releasing Hormone. Biol. Psychiatry 2002, 51, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Aisa, B.; Tordera, R.; Lasheras, B.; Del Río, J.; Ramírez, M.J. Cognitive impairment associated to HPA axis hyperactivity after maternal separation in rats. Psychoneuroendocrinology 2007, 32, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Dziurkowska, E.; Wesolowski, M. Salivary cortisol in women with major depressive disorder under selective serotonin reuptake inhibitors therapy. Arch. Womens Ment. Health 2013, 16, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Vincent, M.Y.; Hussain, R.J.; Zampi, M.E.; Sheeran, K.; Solomon, M.B.; Herman, J.P.; Khan, A.; Jacobson, L. Sensitivity of depression-like behavior to glucocorticoids and antidepressants is independent of forebrain glucocorticoid receptors. Brain Res. 2013, 1525, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sazali, H.A.; Nordin, H.L.; Sargent, M.V. Kaempferitrin from the Leaves of Hedyotis verticillata and its Biological Activity. Planta Med. 1994, 60, 388–389. [Google Scholar] [PubMed]
- Yerra, K.R.; Madamanchi, G.; Hsiao-Sung, C.; Wen-Shi, W.; Yew-Min, T. High-performance liquid chromatographic determination of kaempferol glycosides in Cinnamomum osmophloeum leaves. Int. J. Appl. Sci. Eng. 2009, 7, 1–9. [Google Scholar]
- Venâncio, E.T.; Rocha, N.F.M.; Rios, E.R.V.; Feitosa, M.L.; Linhares, M.I.; Melo, F.H.C.; Matias, M.S.; Fonseca, F.N.; Sousa, F.C.F.; Leal, L.K.A.M.; et al. Anxiolytic-like effects of standardized extract of Justicia pectoralis (SEJP) in mice: Involvement of GABA/benzodiazepine in receptor. Phytother. Res. 2011, 25, 444–450. [Google Scholar] [PubMed]
- Sepúlveda-Jimenez, G.; Reyna-Aquino, C.; Chaires-Martínez, L.; Bermúdez-Torres, K.; Rodríguez-Monroy, M. Antioxidant activity and content of phenolic compounds and flavonoids from Justicia spicigera. J. Biol. Sci. 2009, 9, 629–632. [Google Scholar] [CrossRef]
- Joseph, H.; Gleye, J.; Moulis, C.; Fouraste, I.; Stanislas, E. O-Methoxylated C-glycosylflavones from Justicia pectoralis. J. Nat. Prod. 1988, 51, 804–805. [Google Scholar] [CrossRef] [PubMed]
- David, D.J.; Bourin, M.; Hascoet, M.; Colombel, M.C.; Baker, G.B.; Jolliet, P. Comparison of antidepressant activity in 4- and 40-week-old male mice in the forced swimming test: Involvement of 5-HT1A and 5-HT1B receptors in old mice. Psychopharmacology 2001, 153, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Le Pinchon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: A primary screening test for antidepressants. Achieves Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Venna, V.R.; Deplanque, D.; Allet, C.; Belarbi, K.; Hamdane, M.; Bordet, R. PUFA induce antidepressant-like effects in parallel to structural and molecular changes in the hippocampus. Psychoneuroendocrinology 2009, 34, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vázquez, M.; Estrada-Reyes, R.; Araujo Escalona, A.G.; Ledesma Velázquez, I.; Martínez-Mota, L.; Moreno, J.; Heinze, G. Antidepressant-like effects of an alkaloid extract of the aerial parts of Annona cherimolia in mice. J. Ethnopharmacol. 2012, 139, 164–170. [Google Scholar] [CrossRef] [PubMed]
- López-Rubalcava, C.; Mostalac-Preciado, C.; Estrada-Camarena, E. The rat forced swimming test: An animal model for the study of antidepressant drugs. In Models of Neuropharmacology; Rocha-Arrieta, L.L., Grandos-Soto, V., Eds.; Transworld Res Network: Kenin, India, 2009; pp. 149–168. [Google Scholar]
- Estrada-Reyes, R.; Lopez-Rubalcava, C.; Ferreyra-Cruz, O.A.; Dorantes-Barron, A.M.; Heinze, G.; Moreno, A.J.; Martinez-Vazquez, M. Central nervous system effects and chemical composition of two subspecies of Agastache mexicana; an ethnomedicine of Mexico. J. Ethnopharmacol. 2014, 153, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.-C.; Ko, Y.-J.; Cheng, H.-Y.; Chang, C.-W.; Lin, Y.-C.; Cheng, Y.-H.; Hsieh, M.-T.; Peng, W.H. Antidepressant-Like activity of the Ethanolic Extract from Uncaria lanosa Wallich var. appendiculata Ridsd in the Forced Swimming Test and in the Tail Suspension Test in Mice. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Martínez-Mota, L.; Estrada-Camarena, E.; López-Rubalcava, C. Indorenate produces antidepressant-like actions in the rat forced swimming test via 5-HT1A receptors. Psychopharmacology 2002, 165, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Zhen, L.; Zhu, J.; Zhao, X.; Huang, W.; An, Y.; Li, S.; Du, X.; Lin, M.; Wang, Q.; Xu, Y.; et al. The antidepressant-like effect of fisetin involves the serotonergic and noradrenergic system. Behav. Brain Res. 2012, 17, 359–366. [Google Scholar] [CrossRef]
- Benkert, O.; Gründer, G.; Wetzel, H. Is there an advantage to venlafaxine in comparison with other antidepressants? Hum. Psychopharmacol. 1997, 12, 53–64. [Google Scholar] [CrossRef]
- Dhingra, D.; Sharma, A. Antidepressant-like activity of Glycyrrhiza glabra L. in mouse models of immobility tests. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Zomkowski, A.D.E.; Rosa, A.O.; Lin, J.; Santos, A.R.S.; Calixto, J.B.; Severo, R.A.L. Evidence for serotonin receptor subtypes involvement in agmatine antidepressant like-effect in the mouse forced swimming test. Brain Res. 2004, 1023, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Capra, J.C.; Cunha, M.P.; Machado, D.G.; Zomkowski, A.D.E.; Mendes, B.G.; Santos, A.R.S.; Pizzolatti, M.G.; Rodrigues, A.L.S. Antidepressant-like effect of scopoletin, a coumarin isolated from Polygala sabulosa (Polygalaceae) in mice: Evidence for the involvement of monoaminergic systems. Eur. J. Pharmacol. 2010, 643, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.-T.; Li, Y.-C.; Pan, Y.; Li, J.-M.; Xu, Q.; MO, S.-F.; Qiao, C.-F.; Jiang, F.-X.; Xu, H.X.; Lu, X.-B.; et al. Antidepressant-like effect of psoralidin isolated from the seeds of Psoralea Corylifolia in the forced swimming test in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Redrobe, P.J.; MacSweeney, C.P.; Bourin, M. The role of 5-HT1A and 5-HT B receptors in antidepressant drug actions in the mouse forced swimming test. Eur. J. Pharmacol. 1996, 318, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.-T.; Li, J.-M.; Li, Y.-C.; Pang, Y.; Xu, Q.; Kong, L.-D. Antidepressant-like behavioral and neurochemical effects of the citrus-associated chemical apigenin. Life Sci. 2008, 82, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, Y.; Wu, H.-L.; Li, Y.-B.; Li, Y.-H.; Guo, J.-B.; Li, X.-J. The antidepressant effects of curcumin in the forced swimming test involve 5-HT1 and 5-HT2 receptors. Eur. J. Pharmacol. 2008, 578, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Borsini, F. Role of the Serotonergic System in the Forced Swimming Test. Neurosci. Biobehav. Rev. 1995, 19, 377–395. [Google Scholar] [CrossRef] [PubMed]
- Zhen, W.; Jianhua, G.; Xueer, W.; Kai, X.; Qinsong, L.; Nianqing, W.; Qun, Z.; Hong, J.; Dexiang, L. Antidepressant-like activity of resveratrol treatment in the forced swim test and tail suspension test in mice: The HPA axis, BDNF expression and phosphorylation of ERK. Pharmacol. Biochem. Behav. 2013, 112, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Kaster, M.P.; Santos, A.R.S.; Rodrigues, A.L.S. Involvement of 5-HT1A receptors in the antidepressant-like effect of adenosine in the mouse forced swimming test. Brain Res. Bull. 2005, 67, 53–61. [Google Scholar] [CrossRef] [PubMed]
- National Institutes of Health. Guide for the Care and Use of Laboratory Animals; NIH Publication No. 85-23; National Academies Press: Washington, DC, USA, 1985. [Google Scholar]
- Rogóz, Z.; Kabzinski, M.; Sadaj, W.; Rachwalswa, P.; Gądek-Michalska, A. Effect of co-treatment with fluoxetine or mirtazapine and risperidone on the active bahaviors in rats subjected to the forced swim test. Pharmacol. Rep. 2012, 64, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Connor, T.J.; Kelliher, P.; Harkin, A.; Kelly, J.P.; Leonard, B.E. Reboxetine attenuates forced swim test-induced behavioural and neurochemical alterations in the rat. Eur. J. Pharmacol. 1999, 379, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-X.; Jing, Z.L.; Zhang, L.-M.; Xue, R.; Xu, X.-D.; Zhao, N.; Qiu, Z.-K.; Wang, X.-W.; Zhang, Y.-Z.; Yang, R.-F.; et al. Antidepressant-like acticity of YL-0919: A novel combined selective serotonin reuptake inhibitor and 5-HT1A receptor agonist. PLoS One 2013, 8, e83271. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.A.L.; da Silva, L.G.; Mateussi, S.A.; Fernandes, S.E.; Miguel, G.O.; Yunes, A.R.; Calixto, B.J.; Santos, S.R.A. Ivolvement of monoaminergic system in the antidepressant-like effect of hydroalcoholic extract of Siphocampylus verticillatus. Life Sci. 2002, 70, 1347–1358. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds kaempferitrin not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassani, J.; Dorantes-Barrón, A.M.; Novales, L.M.; Real, G.A.; Estrada-Reyes, R. Anti-Depressant-Like Effect of Kaempferitrin Isolated from Justicia spicigera Schltdl (Acanthaceae) in Two Behavior Models in Mice: Evidence for the Involvement of the Serotonergic System. Molecules 2014, 19, 21442-21461. https://doi.org/10.3390/molecules191221442
Cassani J, Dorantes-Barrón AM, Novales LM, Real GA, Estrada-Reyes R. Anti-Depressant-Like Effect of Kaempferitrin Isolated from Justicia spicigera Schltdl (Acanthaceae) in Two Behavior Models in Mice: Evidence for the Involvement of the Serotonergic System. Molecules. 2014; 19(12):21442-21461. https://doi.org/10.3390/molecules191221442
Chicago/Turabian StyleCassani, Julia, Ana María Dorantes-Barrón, Lilian Mayagoitia Novales, Guadalupe Alva Real, and Rosa Estrada-Reyes. 2014. "Anti-Depressant-Like Effect of Kaempferitrin Isolated from Justicia spicigera Schltdl (Acanthaceae) in Two Behavior Models in Mice: Evidence for the Involvement of the Serotonergic System" Molecules 19, no. 12: 21442-21461. https://doi.org/10.3390/molecules191221442
APA StyleCassani, J., Dorantes-Barrón, A. M., Novales, L. M., Real, G. A., & Estrada-Reyes, R. (2014). Anti-Depressant-Like Effect of Kaempferitrin Isolated from Justicia spicigera Schltdl (Acanthaceae) in Two Behavior Models in Mice: Evidence for the Involvement of the Serotonergic System. Molecules, 19(12), 21442-21461. https://doi.org/10.3390/molecules191221442