Curcumin and Inflammatory Bowel Disease: Potential and Limits of Innovative Treatments


Abstract
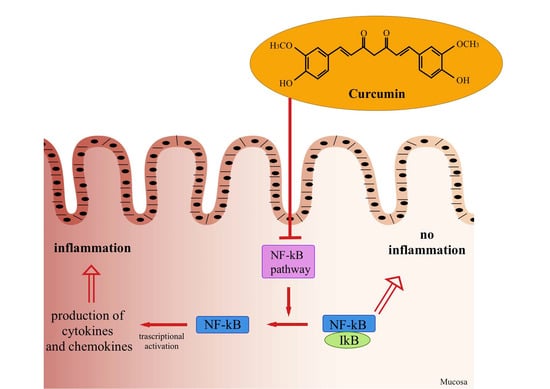
:
1. Introduction
2. Generalities on Inflammatory Bowel Disease

{kind=link}
{kind=link}
| Classification | Montreal | Paris |
|---|---|---|
| Age at Diagnosis | A1 <17 year A2 17–40 year A3 above 40 year | A1a 0–10 year A1b 10–17 year A2 17–40 year A3 > 40 year |
| Location | L1 terminal ileal ± limited cecal disease L2 colonic L3 ileocolonic L4 only upper disease | L1 distal 1/3 ileum ± limited cecal disease L2 colonic L3 ileocolonic L4a upper disease proximal to ligament of Treitz L4b upper disease distal to ligament of Treitz and proximal to distal 1/3 ileum |
| Behavior | B1 non stricturingnon penetrating B2 stricturing B3 penetrating P perianal disease modifier | B1 non stricturingnon penetrating B2 stricturing B3 penetrating B2B3 both penetrating and stricturing disease, either at the same or different times P perianal disease modifier |
| Growth | not classified | G0 no evidence G1 growth delay |
2.1. Environmental Exposure and Lifestyle
2.2. Genetic Involvement in IBD
2.3. Immune Response and IBD
3. IBD and Pharmacological Treatments
Adverse Effects of IBD Treatments
4. Curcumin: Potential and Limits
| Localization | Symptoms | Cytokine Inflammation | TH1/TH2 | Treatments | |
|---|---|---|---|---|---|
| Crohn’s Disease (CD) | Deep layers of the intestinal wall, the ileum, the first part of the colon, esophagus, stomach and duodenum | Pain in the abdomen, diarrhoea, weight loss, rectal bleeding and fever | Interferon gamma (IFN-Y), Interlukin 12 (IL-12), Tumor Necrosis Factor (TNF) | TH1 disease | Anti-inflammation drugs, corticosteroids, immunomodulators and biologic treatments |
| Ulcerative Colitis (UC) | Inner lining of the colon (large interstine) and rectum | Diarrhoea, abdominal cramps, rectal bleeding, frequent fever and nausea | Interlukin 5 (IL-5), Interlukin 33/Interlukin 1, Receptor ST2 (IL-33/ST2) | TH2 disease | Aminosalicylates, corticosteroids, immunomodulators and biological treatments |
| Targets of Curcumin | Effects of Curcumin | Diseases Involved |
|---|---|---|
| Activation of redox-regulated transcription factor Nrf2 that induces heme oxygenase 1 (HO1) paraoxonase 1 (PON1) and GSH 3 [58,60,61,62] | Free-radical-scavenging activity | Chronic inflammatory diseases |
| Inhibition of DNA-binding of STAT3 3 [63] | Anti-inflammatory activity | Chronic inflammatory diseases |
| Reduced phosphorylation of cytosolic phospholipase A2 (cPLA2) limiting the arachidonic acid availability [64] | Anti-inflammatory activity | Chronic inflammatory diseases |
| Reduced phosphorylation of IκB [65] | Anti-inflammatory activity | Chronic inflammatory diseases Cancer |
| Inhibition of the transcription factor Nf-κB [65,66,67,68,69,70,71,72] | Anti-inflammatory activityAnti-oxidant activity Tumor suppressive activity | Chronic inflammatory diseases Cancer |
| Inhibition of mRNA levels of COX2 and iNOS [73,74,75,76] | Anti-inflammatory activity Tumor suppressive activity | Chronic inflammatory diseases Cancer |
| Inhibition of matrix metalloproteinases MMP-9 and MMP-2 [72,77,78, 79] | Tumor suppressive activity (Anti-inflammatory activity) | Cancer (Chronic inflammatory diseases) |
| Inhibition of histone deacetylases (HDACs) and acetyltransferases (HATs) activity [80,81,82,83,84,85,86,87,88] | Gene regulation | Cancer |
| Up-regulation and down-regulation of micro RNA (22, 199, 186, 203) [80,89,90,91] | Pro-apoptotic activity Tumor suppressive activity | Cancer |
| Activation of caspase 3, 7, 8 and caspase 9 [92,93,94] | Pro-apoptotic activity | Cancer |
| Increased cleavage of poly (ADP-ribose) polymerase (PARP) [67,95] | Pro-apoptotic activity | Cancer |
| Up-regulation of several tumor suppressor genes [95,96,97,98,99] | Tumor suppressive activity | Cancer |
| Up-regulation of different proapoptotic genes [100,101,102,103] | Pro-apoptotic activity Tumor suppressive activity | Cancer |
| Inactivation of several oncogenes [104,105,106,107,108,109,110,111,112] | Tumor suppressive activity | Cancer |
| Down-regulation of different antiapoptotic genes [113,114] | Pro-apoptotic activity Tumor suppressive activity | Cancer |
| Inhibition of angiogenesis suppressing VEGF, Akt and PI3K [65,115,116,117,118] | Tumor suppressive activity | Cancer |
| Inhibition of enzymes of phase I reactions [119,120,121] | Tumor suppressive activity | Cancer |
| Activation of enzymes of phase II reactions [122,123,124,125,126,127,128] | Tumor suppressive activity | Cancer |
| Down-regulation of androgen receptor (AR) [110,129,130,131] | Tumor suppressive activity | Cancer |
| Repressed N-methyl-D-aspartate (NMDA) receptor-mediated Ca2+ [132,133,134,135,136] | Protection from excitotoxicity | Neurodegenerative diseases |
| Reduced oxidative mitochondrial damage [137,138,139,140] | Antioxidative activity | Neurodegenerative diseases |
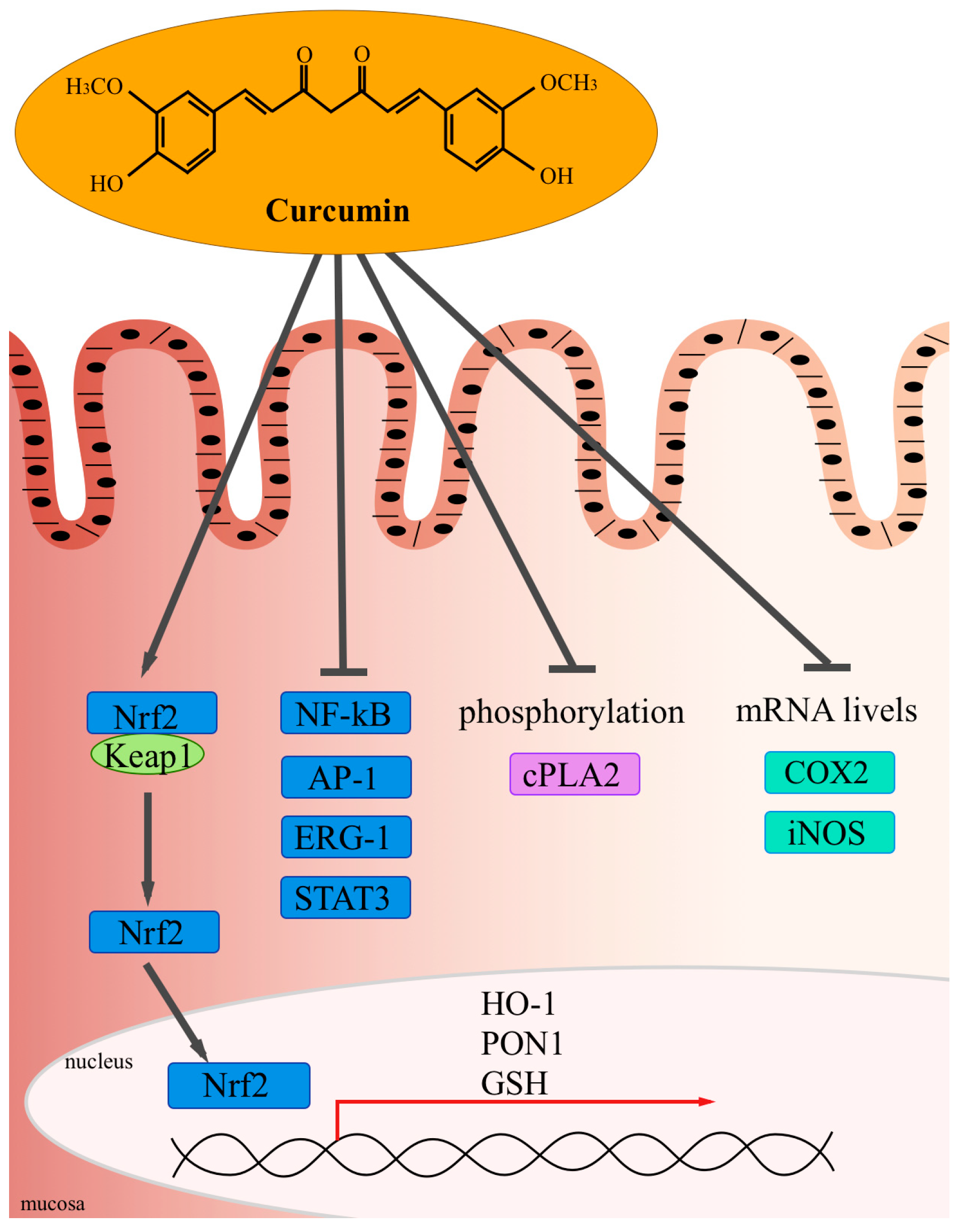
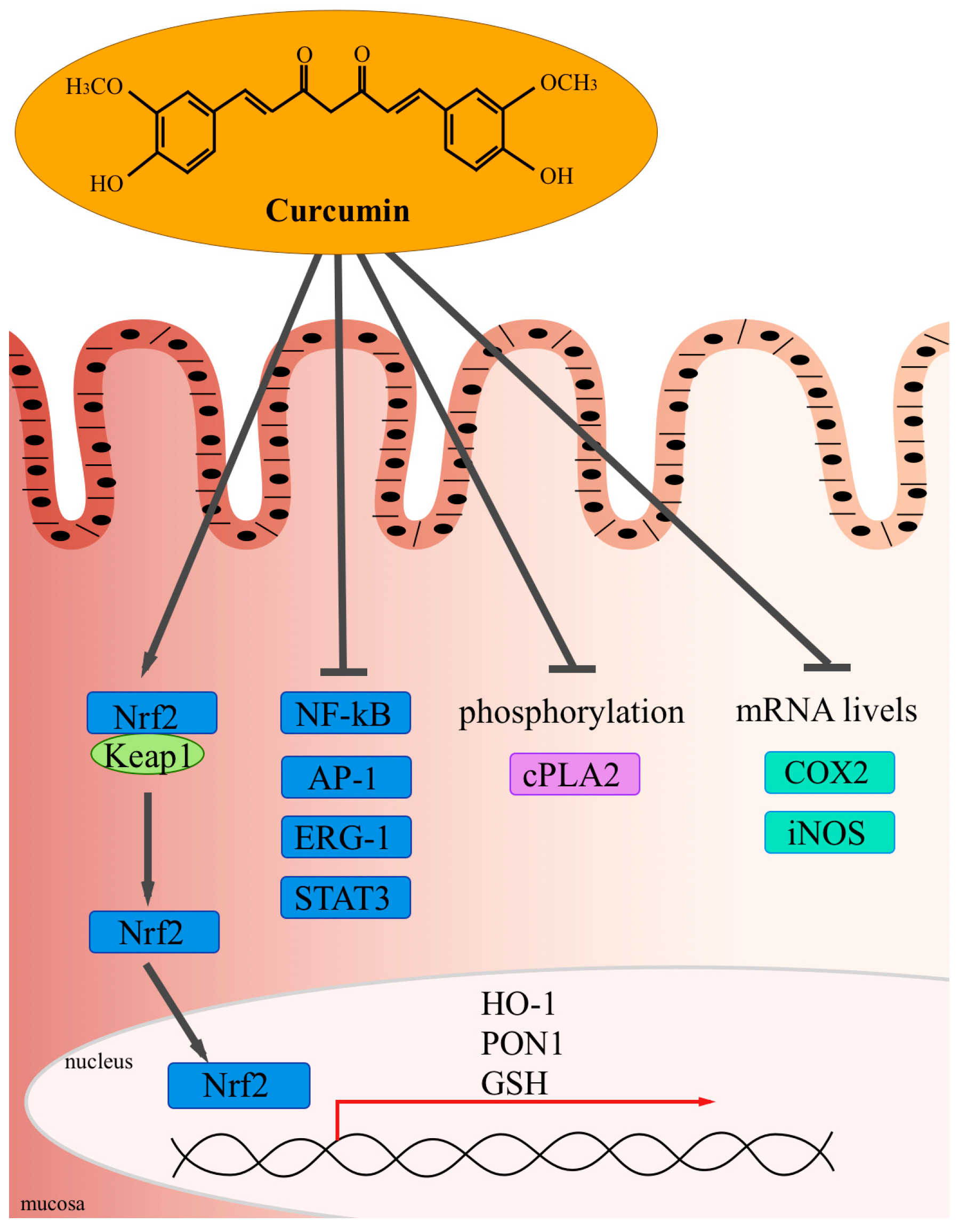
Experimental Studies and Clinical Trials
5. Curcumin and Inflammation
| ClinicalTrials.gov Identifier | Number of Patients (Age) | Disease | Doses of Curcumin | Phase |
|---|---|---|---|---|
| NCT01320436 | 50 (18 to 70 years) | Ulcerative colitis (Disease activity score of >5 and ≤13 according to the Simple clinical colitis activity index (SCCAI) | Patients allocated for this arm will receive 5ASA medication (as advised by their treating physician) + 3 capsules (820 mg each) curcumin twice daily after meals. | 3 |
| NCT00889161 | 11 (8 to 18 years) | Inflammatory bowel disease (mild disease or in clinical remission) [160] | Initial dosage of 500 mg twice a day for 3 weeks. Using the forced dose titration design, dose will be titrated up to 1 g twice a day at week 3 for a total of three weeks and then titrated again to 2 g twice a day at week 6 for three weeks | 1 |
| NCT00793130 | 30 (18 to 75 years) | Mild or moderate Ulcerative Colitis | Dietary Supplement: Coltect Two tablets twice daily (BID) during the 2 months of the study. Each tablet contains 500 mg Curcumin, 250 mg Green tea and 100 μg Selenomethionine. | Unknown |
| NCT01647412 | 40 (10 to 17 years) | Crohn’s Disease (Moderate to severely active CD, as defined by a PCDAI score >30 and | The experimental group will receive the exclusion diet and nutraceutical therapy (DNT) and daily subcutaneously administered recombinant human growth hormone (rhGH) for the first 26 weeks. After 26 weeks this group will continue on the exclusion diet nutraceutical therapy for the remaining 26 weeks of the study. | 2 |
| 89 (13 to 65 years) | Ulcerative Colitis (patients in remission of disease) [148] | Oral curcumin (2 g/day; 1 g morning and evening, after meals) | Concluded |
6. Curcumin Analogues and Nanoformulations
6.1. Curcumin Analogues
6.2. Nanoformulations
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gupta, S.C.; Kismali, G.; Aggarwal, B.B. Curcumin, a component of turmeric: From farm to pharmacy. Biofactors 2013, 39, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ahuja, A.; Ali, J.; Baboota, S. Conundrum and therapeutic potential of curcumin in drug delivery. Crit. Rev. Ther. Drug Carrier Syst. 2010, 27, 279–312. [Google Scholar] [CrossRef] [PubMed]
- Ammon, H.P.; Wahl, M.A. Pharmacology of Curcuma longa. Planta Med. 1991, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lev-Ari, S.; Strier, L.; Kazanov, D.; Elkayam, O.; Lichtenberg, D.; Caspi, D.; Arber, N. Curcumin synergistically potentiates the growth-inhibitory and pro-apoptotic effects of celecoxib in osteoarthritis synovial adherent cells. Rheumatology 2006, 45, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Neerati, P.; Devde, R.; Gangi, A.K. Evaluation of the effect of curcumin capsules on glyburide therapy in patients with Type-2 Diabetes Mellitus. Phytother. Res. 2014. [Google Scholar] [CrossRef]
- Kim, Y.S.; Young, M.R.; Bobe, G.; Colburn, N.H.; Milner, J.A. Bioactive food components, inflammatory targets, and cancer prevention. Cancer Prev. Res. (Phila) 2009, 2, 200–208. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.J.; Lee, K.W. Naturally occurring phytochemicals for the prevention of Alzheimer’s disease. J. Neurochem. 2010, 112, 1415–1430. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 19, 2066–2078. [Google Scholar] [CrossRef]
- De Bie, C.I.; Paerregaard, A.; Kolacek, S.; Ruemmele, F.M.; Koletzko, S.; Fell, J.M.; Escher, J.C.; EUROKIDS Porto IBD Working Group of ESPGHAN. Disease phenotype at diagnosis in pediatric Crohn’s disease: 5-Year analyses of the EUROKIDS Registry. Inflamm. Bowel Dis. 2013, 19, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, F.; Ansari, N.A.; Mazzamurro, F.; Civitelli, F.; Viola, F.; Cucchiara, S.; Catalano, C. Detection of Crohn Disease Lesions of the Small and Large Bowel in Pediatric Patients: Diagnostic Value of MR Enterography Versus Reference Examinations. Am. J. Roentgenol. 2014, 203, W533–W542. [Google Scholar] [CrossRef]
- Aloi, M.; Lionetti, P.; Barabino, A.; Guariso, G.; Costa, S.; Fontana, M.; Romano, C.; Lombardi, G.; Miele, E.; Alvisi, P.; et al. Phenotype and disease course of early-onset pediatric inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.; Griffiths, A.; Markowitz, J.; Wilson, D.C.; Turner, D.; Russell, R.K.; Fell, J.; Ruemmele, F.M.; Walters, T.; Sherlock, M.; et al. Pediatric modification of the Montreal classification for inflammatory bowel disease: the Paris classification. Inflamm. Bowel Dis. 2011, 17, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Ponder, A.; Long, M.D. A clinical review of recent findings in the epidemiology of inflammatory bowel disease. Clin. Epidemiol. 2013, 5, 237–247. [Google Scholar] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Bernstein, C.N.; Vatn, M.H.; Lakatos, P.L.; Loftus, E.V., Jr.; Tysk, C.; O’Morain, C.; Moum, B.; Colombel, J.F.; Epidemiology and Natural History Task Force of the International Organization of Inflammatory Bowel Disease (IOIBD). Geographical variability and environmental risk factors in inflammatory bowel disease. Gut 2013, 62, 630–649. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Kaplan, G.G. Environmental risk factors for inflammatory bowel disease. Gastroenterol. Hepatol. (N. Y.) 2010, 6, 339–346. [Google Scholar]
- Castiglione, F.; Diaferia, M.; Morace, F.; Labianca, O.; Meucci, C.; Cuomo, A.; Panarese, A.; Romano, M.; Sorrentini, I.; D’Onofrio, C.; et al. Risk factors for inflammatory bowel diseases according to the “hygiene hypothesis”: A case-control, multi-centre, prospective study in Southern Italy. J. Crohn’s Colitis 2012, 6, 324–329. [Google Scholar] [CrossRef]
- Baron, S.; Turck, D.; Leplat, C.; Merle, V.; Gower-Rousseau, C.; Marti, R.; Yzet, T.; Lerebours, E.; Dupas, J.L.; Debeugny, S.; et al. Environmental risk factors in paediatric inflammatory bowel diseases: A population based case control study. Gut 2005, 54, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Klement, E.; Cohen, R.V.; Boxman, J.; Joseph, A.; Reif, S. Breastfeeding and risk of inflammatory bowel disease: A systematic review with meta-analysis. Am. J. Clin. Nutr. 2004, 80, 1342–1352. [Google Scholar] [PubMed]
- Barclay, A.R.; Russell, R.K.; Wilson, M.L.; Gilmour, W.H.; Satsangi, J.; Wilson, D.C. Systematic review: The role of breastfeeding in the development of pediatric inflammatory bowel disease. J. Pediatr. 2009, 155, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Khalili, H.; Ananthakrishnan, A.N.; Higuchi, L.M.; Richter, J.M.; Fuchs, C.S.; Chan, A.T. Early life factors and risk of inflammatory bowel disease in adulthood. Inflamm. Bowel Dis. 2013, 19, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Kronman, M.P.; Zaoutis, T.E.; Haynes, K.; Feng, R.; Coffin, S.E. Antibiotic exposure and IBD development among children: A population-based cohort study. Pediatrics 2012, 130, 794–803. [Google Scholar] [CrossRef]
- Shaw, S.Y.; Blanchard, J.F.; Bernstein, C.N. Association between the use of antibiotics in the first year of life and pediatric inflammatory bowel disease. Am. J. Gastroenterol. 2010, 105, 2687–2692. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.Y.; Blanchard, J.F.; Bernstein, C.N. Association between the use of antibiotics and new diagnoses of Crohn’s disease and ulcerative colitis. Am. J. Gastroenterol. 2011, 106, 2133–2142. [Google Scholar] [CrossRef] [PubMed]
- Soon, I.S.; Molodecky, N.A.; Rabi, D.M.; Ghali, W.A.; Barkema, H.W.; Kaplan, G.G. The relationship between urban environment and the inflammatory bowel diseases: A systematic review and meta-analysis. BMC Gastroenterol. 2012, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G.; Hubbard, J.; Korzenik, J.; Sands, B.E.; Panaccione, R.; Ghosh, S.; Wheeler, A.J.; Villeneuve, P.J. The inflammatory bowel diseases and ambient air pollution: A novel association. Am. J. Gastroenterol. 2010, 105, 2412–2419. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.; Levy, E.; Mack, D.; Israel, D.; Lambrette, P.; Ghadirian, P.; Deslandres, C.; Morgan, K.; Seidman, E.G.; Amre, D.K. Dietary patterns and risk for Crohn’s disease in children. Inflamm. Bowel Dis. 2008, 14, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Chapman-Kiddell, C.A.; Davies, P.S.; Gillen, L.; Radford-Smith, G.L. Role of diet in the development of inflammatory bowel disease. Inflamm. Bowel Dis. 2010, 16, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Heyman, M.B.; Kirschner, B.S.; Gold, B.D.; Ferry, G.; Baldassano, R.; Cohen, S.A.; Winter, H.S.; Fain, P.; King, C.; Smith, T.; et al. Children with early-onset inflammatory bowel disease (IBD): analysis of a pediatric IBD consortium registry. J. Pediatr. 2005, 146, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.M.; Zanin, V.; Girardelli, M.; Magnolato, A.; Martelossi, S.; Tommasini, A.; Marcuzzi, A.; Crovella, S. A common genetic background could explain early-onset Crohn’s disease. Med. Hypotheses 2012, 78, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Cannioto, Z.; Berti, I.; Martelossi, S.; Bruno, I.; Giurici, N.; Crovella, S.; Ventura, A. IBD and IBD mimicking enterocolitis in children younger than 2 years of age. Eur. J. Pediatr. 2009, 168, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A.; et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, H.H. Monogenic diseases associated with intestinal inflammation: Implications for the understanding of inflammatory bowel disease. Gut 2013, 62, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.M.; Girardelli, M.; Vozzi, D.; Crovella, S.; Kleiner, G.; Marcuzzi, A. Mevalonate kinase deficiency and IBD: Shared genetic background. Gut 2014, 63, 1367–1368. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, H.H.; Schwerd, T.; Koletzko, S. The diagnostic approach to monogenic very early onset inflammatory bowel disease. Gastroenterology 2014, 147, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, L.; Garg, R.R.; Hoang, S.B.; Spina, L.; Mattioli, B.; Scarpa, M.; Fiocchi, C.; Vecchi, M.; Pizarro, T.T. Epithelial-derived IL-33 and its receptor ST2 are dysregulated in ulcerative colitis and in experimental Th1/Th2 driven enteritis. Proc. Natl. Acad. Sci. USA 2010, 107, 8017–8022. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, F. Cytokine-based therapies for Crohn’s disease-new paradigms. N. Engl. J. Med. 2004, 351, 2045–2048. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Martin, C.; Mishina, M.; Ross, W.G.; Rivera-Nieves, J.; Marini, M.; Cominelli, F. Proinflammatory effects of TH2 cytokines in a murine model of chronic small intestinal inflammation. Gastroenterology 2005, 128, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Wallace, K.L.; Zheng, L.B.; Kanazawa, Y.; Shih, D.Q. Immunopathology of inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Seidelin, J.B.; Coskun, M.; Kvist, P.H.; Holm, T.L.; Holgersen, K.; Nielsen, O.H. IL-33 promotes GATA-3 polarization of gut-derived T cells in experimental and ulcerative colitis. J. Gastroenterol. 2014. [Google Scholar] [CrossRef]
- Karatzas, P.S.; Gazouli, M.; Safioleas, M.; Mantzaris, G.J. DNA methylation changes in inflammatory bowel disease. Ann. Gastroenterol. 2014, 27, 125–132. [Google Scholar] [PubMed]
- Orel, R.; Kamhi Trop, T. Intestinal microbiota, probiotics and prebiotics in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 11505–11524. [Google Scholar] [CrossRef] [PubMed]
- Faubion, W.A., Jr.; Loftus, E.V., Jr.; Harmsen, W.S.; Zinsmeister, A.R.; Sandborn, W.J. The natural history of corticosteroid therapy for inflammatory bowel disease: A population-based study. Gastroenterology 2001, 121, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Present, D.H.; Meltzer, S.J.; Krumholz, M.P.; Wolke, A.; Korelitz, B.I. 6-Mercaptopurine in the management of inflammatory bowel disease: short- and long-term toxicity. Ann. Intern. Med. 1989, 111, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Ardizzone, S.; Bianchi Porro, G. Biologic therapy for inflammatory bowel disease. Drugs 2005, 65, 2253–2286. [Google Scholar] [CrossRef] [PubMed]
- Lazzerini, M.; Martelossi, S.; Magazzù, G.; Pellegrino, S.; Lucanto, M.C.; Barabino, A.; Calvi, A.; Arrigo, S.; Lionetti, P.; Lorusso, M.; et al. Effect of thalidomide on clinical remission in children and adolescents with refractory Crohn disease: A randomized clinical trial. JAMA 2013, 27, 2164–2173. [Google Scholar] [CrossRef]
- Renna, S.; Cottone, M.; Orlando, A. Optimization of the treatment with immunosuppressants and biologics in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 9675–9690. [Google Scholar] [CrossRef] [PubMed]
- Feagan, B.G.; Rochon, J.; Fedorak, R.N.; Irvine, E.J.; Wild, G.; Sutherland, L.; Steinhart, A.H.; Greenberg, G.R.; Gillies, R.; Hopkins, M. Methotrexate for the treatment of Crohn’s disease. The North American Crohn’s Study Group Investigators. N. Engl. J. Med. 1995, 332, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Felipez, L.M.; Gokhale, R.; Tierney, M.P.; Kirschner, B.S. Thalidomide use and outcomes in pediatric patients with Crohn disease refractory to infliximab and adalimumab. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 28–33. [Google Scholar] [PubMed]
- Denadaia, R.; Vieira Teixeiraa, F.; Steinwurzb, F.; Romitic, R.; Saad-Hossnea, R. Induction or exacerbation of psoriatic lesions during anti-TNF-α therapy for inflammatory bowel disease: A systematic literature review based on 222 cases. J. Crohn’s Colitis 2013, 7, 517–524. [Google Scholar] [CrossRef]
- Uyanikoglu, A.; Ermis, F.; Akyuz, F.; Pinarbasi, B.; Baran, B.; Aydogan, T.; Demir, K.; Besisik, F.; Kaymakoglu, S. Infliximab in inflammatory bowel disease: Attention to adverse events. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2337–2342. [Google Scholar] [PubMed]
- Lakatos, P.L.; Miheller, P. Is there an increased risk of lymphoma and malignancies under anti-TNF therapy in IBD? Curr. Drug Targets 2010, 11, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, A.; Hagel, S.; Bruns, T. Adverse effects of biologics used for treating IBD. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Van Assche, G.; D’haens, G.; Noman, M.; Vermeire, S.; Hiele, M.; Asnong, K.; Arts, J.; D’hoore, A.; Penninckx, F.; Rutgeerts, P. Randomized, double-blind comparison of 4 mg/kg versus 2 mg/kg intravenous cyclosporine in severe ulcerative colitis. Gastroenterology 2003, 125, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Rabizadeh, S.; Dubinsky, M. Update in pediatric inflammatory bowel disease. Rheum. Dis. Clin. North Am. 2013, 39, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.A.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin-from molecule to biological function. Angew. Chem. Int. Ed. 2012, 51, 5308–5332. [Google Scholar] [CrossRef]
- Rahmani, A.H.; al Zohairy, M.A.; Aly, S.M.; Khan, M.A. Curcumin: A potential candidate in prevention of cancer via modulation of molecular pathways. Biomed. Res. Int. 2014, 2014, 761608. [Google Scholar] [CrossRef] [PubMed]
- Balogun, E.; Hoque, M.; Gong, P.; Killeen, E.; Green, C.J.; Foresti, R.; Alam, J.; Motterlini, R. Curcumin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive element. Biochem. J. 2003, 371, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schiborr, C.; Frank, J.; Rimbach, G. Curcumin induces paraoxonase 1 in cultured hepatocytes in vitro but not in mouse liver in vivo. Br. J. Nutr. 2011, 105, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I. Antioxidant therapeutic advances in COPD. Ther. Adv. Respir. Dis. 2008, 2, 351–374. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Zhong, X.; Yum, H.W.; Lee, H.J.; Kundu, J.K.; Na, H.K.; Surh, Y.J. Curcumin Inhibits STAT3 Signaling in the Colon of Dextran Sulfate Sodium-treated Mice. J. Cancer Prev. 2013, 18, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Bose, M.; Ju, J.; Ryu, J.H.; Chen, X.; Sang, S.; Lee, M.J.; Yang, C.S. Modulation of arachidonic acid metabolism by curcumin and related beta-diketone derivatives: Effects on cytosolic phospholipase A(2), cyclooxygenases and 5-lipoxygenase. Carcinogenesis 2004, 25, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Aggarwal, B.B. Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane) [corrected]. J. Biol. Chem. 1995, 270, 24995–5000. [Google Scholar] [CrossRef] [PubMed]
- Plummer, S.M.; Holloway, K.A.; Manson, M.M.; Munks, R.J.; Kaptein, A.; Farrow, S.; Howells, L. Inhibition of cyclo-oxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of NF-kappaB activation via the NIK/IKK signalling complex. Oncogene 1999, 18, 6013–6020. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.C.; Donato, N.; Singh, S.; Aggarwal, B.B. Curcumin (diferuloylmethane) down-regulates the constitutive activation of nuclear factor-kappa B and IkappaBalpha kinase in human multiple myeloma cells, leading to suppression of proliferation and induction of apoptosis. Blood 2003, 101, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.K. Suppression of protein kinase C and nuclear oncogene expression as possible action mechanisms of cancer chemoprevention by Curcumin. Arch. Pharm. Res. 2004, 27, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Amin, H.M.; Lai, R.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive NF-kappaB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem. Pharmacol. 2005, 70, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Ichikawa, H.; Takada, Y.; Sandur, S.K.; Shishodia, S.; Aggarwal, B.B. Curcumin (diferuloylmethane) down-regulates expression of cell proliferation and antiapoptotic and metastatic gene products through suppression of IkappaBalpha kinase and Akt activation. Mol. Pharmacol. 2006, 69, 195–206. [Google Scholar] [PubMed]
- Mackenzie, G.G.; Queisser, N.; Wolfson, M.L.; Fraga, C.G.; Adamo, A.M.; Oteiza, P.I. Curcumin induces cell-arrest and apoptosis in association with the inhibition of constitutively active NF-kappaB and STAT3 pathways in Hodgkin’s lymphoma cells. Int. J. Cancer 2008, 123, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S.; Takada, Y.; Banerjee, S.; Newman, R.A.; Bueso-Ramos, C.E.; Price, J.E. Curcumin suppresses the paclitaxel-induced nuclear factor NF-kappaB pathway in breast cancer cells and inhibits lung metastasis of human breast cancer in nude mice. Clin. Cancer Res. 2005, 11, 7490–7498. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Zhao, Y.X.; Zhang, S.; Liu, G.D.; Kang, W.Y.; Tang, H.D.; Ding, J.Q.; Chen, S.D. PPAR gamma agonist curcumin reduces the amyloid-beta-stimulated inflammatory responses in primary astrocytes. J. Alzheimer Dis. 2010, 20, 1189–1199. [Google Scholar]
- Jin, C.Y.; Lee, J.D.; Park, C.; Choi, Y.H.; Kim, G.Y. Curcumin attenuates the release of pro-inflammatory cytokines in lipopolysaccharide-stimulated BV2 microglia. Acta Pharmacol. Sin. 2007, 28, 645–651. [Google Scholar]
- Goel, A.; Boland, C.R.; Chauhan, D.P. Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett. 2001, 172, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer potential of curcumin: Preclinical and clinical studies. Anticancer Res. 2003, 23, 363–398. [Google Scholar] [PubMed]
- Woo, M.S.; Jung, S.H.; Kim, S.Y.; Hyun, J.W.; Ko, K.H.; Kim, W.K.; Kim, H.S. Curcumin suppresses phorbol ester-induced matrix metalloproteinase-9 expression by inhibiting the PKC to MAPK signaling pathways in human astroglioma cells. Biochem. Biophys. Res. Commun. 2005, 335, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.K.; Daghestani, M.H. Curcumin effect onMMPs and TIMPs genes in a breast cancer cell line. Asian Pac. J. Cancer Prev. 2012, 13, 3259–3264. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kim, J.H.; Lee, H.J.; Surh, Y.J. Curcumin inhibits phorbol ester-induced up-regulation of cyclooxygenase-2 and matrix metalloproteinase-9 by blocking ERK1/2 phosphorylation and NF-kappaB transcriptional activity in MCF10A human breast epithelial cells. Antioxid. Redox Signal. 2005, 7, 1612–1620. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Park, B.; Goel, A.; Aggarwal, B.B. Epigenetic changes induced by curcumin and other natural compounds. Genes Nutr. 2011, 6, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Bora-Tatar, G.; Dayangaç-Erden, D.; Demir, A.S.; Dalkara, S.; Yelekçi, K.; Erdem-Yurter, H. Molecular modifications on carboxylic acid derivatives as potent histone deacetylase inhibitors: Activity and docking studies. Bioorg. Med. Chem. 2009, 17, 5219–5228. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Chen, Y.; Cui, G.H.; Zhou, J.F. Curcumin, a potent anti-tumor reagent, is a novel histone deacetylase inhibitor regulating B-NHL cell line Raji proliferation. Acta Pharmacol. Sin. 2005, 26, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shu, W.; Chen, W.; Wu, Q.; Liu, H.; Cui, G. Curcumin, both histone deacetylase and p300/CBP-specific inhibitor, represses the activity of nuclear factor kappa B and Notch 1 in Raji cells. Basic Clin. Pharmacol. Toxicol. 2007, 101, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Krauthauser, C.; Maduskuie, V.; Fawcett, P.T.; Olson, J.M.; Rajasekaran, S.A. Curcumin-induced HDAC inhibition and attenuation of medulloblastoma growth in vitro and in vivo. BMC Cancer 2011, 11, 144. [Google Scholar] [CrossRef] [PubMed]
- Marcu, M.G.; Jung, Y.J.; Lee, S.; Chung, E.J.; Lee, M.J.; Trepel, J.; Neckers, L. Curcumin is an inhibitor of p300 histone acetylatransferase. Med. Chem. 2006, 2, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Chen, J.; Shi, Y.; Jia, J.; Zhang, Y. Curcumin-induced histone hypoacetylation: The role of reactive oxygen species. Biochem. Pharmacol. 2005, 69, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanyam, K.; Varier, R.A.; Altaf, M.; Swaminathan, V.; Siddappa, N.B.; Ranga, U.; Kundu, T.K. Curcumin, a novel p300/CREB-binding protein-specific inhibitor of acetyltransferase, represses the acetylation of histone/nonhistone proteins and histone acetyltransferase-dependent chromatin transcription. J. Biol. Chem. 2004, 279, 51163–51171. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, T.; Sunagawa, Y.; Kawamura, T.; Takaya, T.; Wada, H.; Nagasawa, A.; Komeda, M.; Fujita, M.; Shimatsu, A.; Kita, T.; et al. The dietary compound curcumin inhibits p300 histone acetyltransferase activity and prevents heart failure in rats. J. Clin. Investig. 2008, 118, 868–878. [Google Scholar] [PubMed]
- Saini, S.; Arora, S.; Majid, S.; Shahryari, V.; Chen, Y.; Deng, G.; Yamamura, S.; Ueno, K.; Dahiya, R. Curcumin modulates microRNA-203-mediated regulation of the Src-Akt axis in bladder cancer. Cancer Prev. Res. (Phila) 2011, 4, 1698–1709. [Google Scholar] [CrossRef]
- Sun, M.; Estrov, Z.; Ji, Y.; Coombes, K.R.; Harris, D.H.; Kurzrock, R. Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Mol. Cancer Ther. 2008, 7, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, T.; Ti, X.; Shi, J.; Wu, C.; Ren, X.; Yin, H. Curcumin promotes apoptosis in A549/DDP multidrug-resistant human lung adenocarcinoma cells through an miRNA signaling pathway. Biochem. Biophys. Res. Commun. 2010, 399, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.J.; Jiang, G.; Li, L.T.; Zheng, J.N. Curcumin induces apoptosis through mitochondrial pathway and caspases activation in human melanoma cells. Mol. Biol. Rep. 2014. [Google Scholar] [CrossRef]
- Ravindran, J.; Prasad, S.; Aggarwal, B.B. Curcumin and cancer cells: How many ways can curry kill tumor cells selectively? AAPS J. 2009, 11, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Mezzanotte, L.; An, N.; Mol, I.M.; Löwik, C.W.; Kaijzel, E.L. A new multicolor bioluminescence imaging platform to investigate NF-κB activity and apoptosis in human breast cancer cells. PLoS One 2014, 9, e85550. [Google Scholar] [CrossRef] [PubMed]
- Sa, G.; Das, T. Anticancer effects of curcumin: Cycle of life and death. Cell Div. 2008, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Kim, E.H.; Park, I.C.; Lee, H.C.; Woo, S.H.; Lee, J.Y.; Hong, Y.J.; Rhee, C.H.; Choi, S.H.; Shim, B.S.; et al. Curcumin inhibits cell cycle progression of immortalized human umbilical vein endothelial (ECV304) cells by up-regulating cyclin-dependent kinase inhibitor, p21WAF1/CIP1, p27KIP1 and p53. Int. J. Oncol. 2002, 21, 379–383. [Google Scholar] [PubMed]
- Weinberg, R.A. The retinoblastoma protein and cell cycle control. Cell 1995, 81, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Chen, Q.; Siddiqui, I.; Sarva, K.; Shankar, S. Linkage of curcumin-induced cell cycle arrest and apoptosis by cyclin-dependent kinase inhibitor p21/WAF1/CIP1. Cell Cycle 2007, 6, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Banerjee, S.; Stafford, L.J.; Xia, C.; Liu, M.; Aggarwal, B.B. Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene 2002, 21, 8852–8861. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Srivastava, R.K. Involvement of Bcl-2 family members, phosphatidylinositol 3'-kinase/AKT and mitochondrial p53 in curcumin (diferulolylmethane)-induced apoptosis in prostate cancer. Int. J. Oncol. 2007, 30, 905–918. [Google Scholar] [PubMed]
- Tourkina, E.; Gooz, P.; Oates, J.C.; Ludwicka-Bradley, A.; Silver, R.M.; Hoffman, S. Curcumin-induced apoptosis in scleroderma lung fibroblasts: Role of protein kinase cepsilon. Am. J. Respir. Cell Mol. Biol. 2004, 31, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhou, X.; He, X.; Dai, M.; Zhang, Q. Curcumin induces apoptosis involving bax/bcl-2 in human hepatoma SMMC-7721 cells. Asian Pac. J. Cancer Prev. 2011, 12, 1925–1929. [Google Scholar] [PubMed]
- Choudhuri, T.; Pal, S.; Agwarwal, M.L.; Das, T.; Sa, G. Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. FEBS Lett. 2002, 512, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Elamin, M.H.; Shinwari, Z.; Hendrayani, S.F.; Al-Hindi, H.; Al-Shail, E.; Khafaga, Y.; Al-Kofide, A.; Aboussekhra, A. Curcumin inhibits the sonic hedgehog signaling pathway and triggers apoptosis in medulloblastoma cells. Mol. Carcinog. 2010, 49, 302–314. [Google Scholar] [PubMed]
- Limtrakul, P.; Anuchapreeda, S.; Lipigorngoson, S.; Dunn, F.W. Inhibition of carcinogen induced c-Ha-ras and cfos proto-oncogenes expression by dietary curcumin. BMC Cancer 2001, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Seol, D.W.; Chen, Q.; Zarnegar, R. Transcriptional activation of the hepatocyte growth factor receptor (c-met) gene by its ligand (hepatocyte growth factor) is mediated through AP-1. Oncogene 2000, 19, 1132–1137. [Google Scholar] [CrossRef] [PubMed]
- Bangaru, M.L.Y.; Chen, S.; Woodliff, J.; Kansra, S. Curcumin (diferuloylmethane) induces apoptosis and blocks migration of humanmedulloblastoma cells. Anticancer Res. 2010, 30, 499–504. [Google Scholar] [PubMed]
- Chen, A.; Xu, J.; Johnson, A.C. Curcumin inhibits human colon cancer cell growth by suppressing gene expression of epidermal growth factor receptor through reducing the activity of the transcription factor Egr-1. Oncogene 2006, 25, 278–287. [Google Scholar] [PubMed]
- Korutla, L.; Cheung, J.Y.; Mendelsohn, J.; Kumar, R. Inhibition of ligand-induced activation of epidermal growth factor receptor tyrosine phosphorylation by curcumin. Carcinogenesis 1995, 16, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Dorai, T.; Gehani, N.; Katz, A. Therapeutic potential of curcumin in human prostate cancer. II. Curcumin inhibits tyrosine kinase activity of epidermal growth factor receptor and depletes the protein. Mol. Urol. 2000, 4, 1–6. [Google Scholar] [PubMed]
- Hong, R.L.; Spohn, W.H.; Hung, M.C. Curcumin inhibits tyrosine kinase activity of p185neu and also depletes p185neu. Clin. Cancer Res. 1999, 5, 1884–1891. [Google Scholar] [PubMed]
- Camacho-Barquero, L.; Villegas, I.; Sáànchez-Calvo, J.M.; Talero, E.; Sánchez-Fidalgo, S.; Motilva, V.; Alarcón de la Lastra, C. Curcumin, a Curcuma longa constituent, acts on MAPK p38 pathway modulating COX-2 and iNOS expression in chronic experimental colitis. Int. Immunopharmacol. 2007, 7, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Catz, S.D.; Johnson, J.L. Transcriptional regulation of bcl-2 by nuclear factor kappaB and its significance in prostate cancer. Oncogene 2001, 20, 7342–7351. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.L.; Briones, F., Jr.; Brisbay, S.; Logothetis, C.J.; McDonnell, T.J. Prostate carcinoma cell death resulting from inhibition of proteasome activity is independent of functionalBcl-2 and p53. Oncogene 1998, 17, 2889–2899. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Guha, S.; Krishnan, S.; Diagaradjane, P.; Gelovani, J.; Aggarwal, B.B. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res. 2007, 67, 3853–3861. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.C.; Hamdy, R.C.; Chua, B.H.L. Mechanism of transforming growth factor-beta1-induced expression of vascular endothelial growth factor in murine osteoblastic MC3T3-E1 cells. Biochim. Biophys. Acta 2000, 1497, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chadalapaka, G.; Jutooru, I.; Chintharlapalli, S.; Papineni, S.; Smith, R., 3rd; Li, X.; Safe, S. Curcumin decreases specificity protein expression in bladder cancer cells. Cancer Res. 2008, 68, 5345–5354. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Shen, G.; Khor, T.O.; Kim, J.H.; Kong, A.N.T. Curcumin inhibits Akt/mammalian target of rapamycin signaling through protein phosphatase-dependent mechanism. Mol. Cancer Ther. 2008, 7, 2609–2620. [Google Scholar] [CrossRef] [PubMed]
- Thapliyal, R.; Maru, G.B. Inhibition of cytochrome P450 isozymes by curcumins in vitro and in vivo. Food Chem. Toxicol. 2001, 39, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Firozi, P.F.; Aboobaker, V.S.; Bhattacharya, R.K. Action of curcumin on the cytochrome P450-system catalyzing the activation of aflatoxin B1. Chem. Biol. Interact. 1996, 100, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Ciolino, H.P.; Daschner, P.J.; Wang, T.T.Y.; Yeh, G.C. Effect of curcumin on the aryl hydrocarbon receptor and cytochrome P450 1A1 inMCF-7 human breast carcinoma cells. Biochem. Pharmacol. 1998, 56, 197–206. [Google Scholar]
- Singh, S.V.; Hu, X.; Srivastava, S.K.; Singh, M.; Xia, H.; Orchard, J.L.; Zaren, H.A. Mechanism of inhibition of benzo[a]pyrene-induced forestomach cancer in mice by dietary curcumin. Carcinogenesis 1998, 19, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Thapliyal, R.; Deshpande, S.S.; Maru, G.B. Mechanism(s) of turmeric-mediated protective effects against benzo(a)pyrenederived DNA adducts. Cancer Lett. 2002, 175, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Ireson, C.R.; Verschoyle, R.D.; Hill, K.A.; Williams, M.L.; Leuratti, C.; Manson, M.M.; Marnett, L.J.; Steward, W.P.; et al. Effects of dietary curcumin on glutathione S-transferase and malondialdehyde-DNA adducts in rat liver and colon mucosa: Relationship with drug levels. Clin. Cancer Res. 2001, 7, 1452–1458. [Google Scholar] [PubMed]
- Nishinaka, T.; Ichijo, Y.; Ito, M.; Kimura, M.; Katsuyama, M.; Iwata, K.; Miura, T.; Terada, T.; Yabe-Nishimura, C. Curcumin activates human glutathione S-transferase P1 expression through antioxidant response element. Toxicol. Lett. 2007, 170, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Piper, J.T.; Singhal, S.S.; Salameh, M.S.; Torman, R.T.; Awasthi, Y.C.; Awasthi, S. Mechanisms of anticarcinogenic properties of curcumin: the effect of curcumin on glutathione linked detoxification enzymes in rat liver. Int. J. Biochem. Cell Biol. 1998, 30, 445–456. [Google Scholar] [CrossRef]
- Valentine, S.P.; le Nedelec, M.J.; Menzies, A.R.; Scandlyn, M.J.; Goodin, M.G.; Rosengren, R.J. Curcumin modulates drug metabolizing enzymes in the female SwissWebster mouse. Life Sci. 2006, 78, 2391–2398. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Gupta, S.; Maru, G.B. Dietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: Mechanism of its anti-initiating action. Carcinogenesis 2008, 29, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xu, Y.M.; Ye, Z.Q.; Yu, J.H.; Hu, X.Y. Curcumin induces cell cycle arrest and apoptosis of prostate cancer cells by regulating the expression of IkappaBalpha, c-Jun and androgen receptor. Pharmazie 2013, 68, 431–434. [Google Scholar] [PubMed]
- Nakamura, K.; Yasunaga, Y.; Segawa, T.; Ko, D.; Moul, J.W.; Srivastava, S.; Rhim, J.S. Curcumin downregulates AR gene expression and activation in prostate cancer cell lines. Int. J. Oncol. 2002, 21, 825–380. [Google Scholar] [PubMed]
- Tsui, K.H.; Feng, T.H.; Lin, C.M.; Chang, P.L.; Juang, H.H. Curcumin blocks the activation of androgen and interlukin-6 on prostate-specific antigen expression in human prostatic carcinoma cells. J. Androl. 2008, 29, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Grant, R.; Adams, S.; Guillemin, G.J. Neuroprotective effects of naturally occurring polyphenols on quinolinic acid-induced excitotoxicity in human neurons. FEBS J. 2010, 277, 368–382. [Google Scholar] [CrossRef] [PubMed]
- Matteucci, A.; Cammarota, R.; Paradisi, S.; Varano, M.; Balduzzi, M.; Leo, L.; Bellenchi, G.C.; de Nuccio, C.; Carnovale-Scalzo, G.; Scorcia, G.; et al. Curcumin protects against NMDA-induced toxicity: A possible role for NR2A subunit. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1070–1077. [Google Scholar] [CrossRef]
- Matteucci, A.; Frank, C.; Domenici, M.R.; Balduzzi, M.; Paradisi, S.; Carnovale-Scalzo, G.; Scorcia, G.; Malchiodi-Albedi, F. Curcumin treatment protects rat retinal neurons against excitotoxicity: Effect on N-methyl-D: -aspartate-induced intracellular Ca(2+) increase. Exp. Brain Res. 2005, 167, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, Y.B.; Li, Y.H.; Xu, Y.; Wu, H.L.; Li, X.J. Curcumin protects against glutamate excitotoxicity in rat cerebral cortical neurons by increasing brain-derived neurotrophic factor level and activating TrkB. Brain Res. 2008, 1210, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, K.; Kihara, T.; Shen, H.; Shimmyo, Y.; Niidome, T.; Sugimoto, H. Distinct mechanisms underlie distinct polyphenol-induced neuroprotection. FEBS Lett. 2006, 580, 6623–6628. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Chen, X.C.; Chen, Z.Z.; Zeng, Y.Q.; Shi, G.B.; Su, Y.H.; Peng, X. Curcumin protects mitochondria from oxidative damage and attenuates apoptosis in cortical neurons. Acta Pharmacol. Sin. 2004, 25, 1606–1612. [Google Scholar] [PubMed]
- Wang, Q.; Sun, A.Y.; Simonyi, A.; Jensen, M.D.; Shelat, P.B.; Rottinghaus, G.E.; MacDonald, R.S.; Miller, D.K.; Lubahn, D.E.; Weisman, G.A.; et al. Neuroprotective mechanisms of curcumin against cerebral ischemia-induced neuronal apoptosis and behavioral deficits. J. Neurosci. Res. 2005, 82, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, M.; Ojha, R.P.; Rajamanickam, G.V.; Agrawal, A.; Aggarwal, A.; Dubey, G.P. Curcuminoids modulates oxidative damage and mitochondrial dysfunction in diabetic rat brain. Free Radical Res. 2008, 42, 999–1005. [Google Scholar] [CrossRef]
- Sood, P.K.; Nahar, U.; Nehru, B. Curcumin attenuates aluminum-induced oxidative stress and mitochondrial dysfunction in rat brain. Neurotoxic. Res. 2011, 20, 351–361. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Jha, A.; Youssef, D.; Rupasinghe, H.P. Curcumin and its carbocyclic analogs: Structure-activity in relation to antioxidant and selected biological properties. Molecules 2013, 18, 5389–5404. [Google Scholar] [CrossRef] [PubMed]
- Somchit, M.; Changtam, C.; Kimseng, R.; Utaipan, T.; Lertcanawanichakul, M.; Suksamrarn, A.; Chunglok, W. Demethoxycurcumin from Curcuma longa rhizome suppresses iNOS induction in an in vitro inflamed human intestinal mucosa model. Asian Pac. J. Cancer Prev. 2014, 15, 1807–1810. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.B.; Gao, J.L.; Zhong, Z.F.; Hoi, P.M.; Lee, S.M.; Wang, Y.T. Bisdemethoxycurcumin suppresses MCF-7 cells proliferation by inducing ROS accumulation and modulating senescence-related pathways. Pharmacol. Rep. 2013, 65, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Beevers, C.S.; Huang, S. The targets of curcumin. Curr. Drug Targets 2011, 12, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Kurien, B.T.; Singh, A.; Matsumoto, H.; Scofield, R.H. Improving the solubility and pharmacological efficacy of curcumin by heat treatment. Assay Drug Dev. Technol. 2007, 5, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Chignell, C.F.; Bilski, P.; Reszka, K.J.; Motten, A.G.; Sik, R.H.; Dahl, T.A. Spectral and photochemical properties of curcumin. Photochem. Photobiol. 1994, 59, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, S.A. Therapeutic potential of curcumin in gastrointestinal diseases. World J. Gastrointest. Pathophysiol. 2011, 15, 1–14. [Google Scholar] [CrossRef]
- Taylor, R.A.; Leonard, M.C. Curcumin for inflammatory bowel disease: A review of human studies. Altern. Med. Re. 2011, 16, 152–156. [Google Scholar]
- Hanai, H.; Sugimoto, K. Curcumin has bright prospects for the treatment of inflammatory bowel disease. Curr. Pharm. Des. 2009, 15, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Iida, T.; Takeuchi, K.; Watanabe, F.; Maruyama, Y.; Andoh, A.; Tsujikawa, T.; Fujiyama, K.; Mitsuyama, K.; Sata, M.; et al. Curcumin maintenance therapy for ulcertive colitis: Randomized, multicenter, double-blind, placebo-controlled trial. Clin. Gastroenterol. Hepatol. 2006, 4, 1502–1506. [Google Scholar] [CrossRef] [PubMed]
- Billerey-Larmonier, C.; Uno, J.K.; Larmonier, N.; Midura, A.J.; Timmermann, B.; Ghishan, F.K.; Kiela, P.R. Protective effects of dietary curcumin in mouse model of chemically induced colitis are strain dependent. Inflamm. Bowel Dis. 2008, 14, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.; Sanderson, I.R.; MacDonald, T.T. Curcumin as a therapeutic agent: The evidence from in vitro animal and human studies. Br. J. Nutr. 2010, 103, 407–412. [Google Scholar]
- Epstein, J.; Docena, G.; MacDonald, T.T.; Sanderson, I.R. Curcumin suppresses p38 mitogen-activated protein kinase activation, reduces IL-1B and matrix metalloproteinase-3 and anhances IL-10 in the mucosa of children and adults with inflammatory bowel disease. Br. J. Nutr. 2010, 103, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.R.; Katz, S.; Kirshoff, R. Curcumin therapy in inflammatory bowel disease: A pilot study. Dig. Dis. Sci. 2005, 50, 2191–2193. [Google Scholar] [CrossRef] [PubMed]
- Salh, B.; Assi, K.; Templeman, V.; Parhar, K.; Owen, D.; Gómez-Muñoz, A.; Jacobson, K. Curcumin attenuates DNB-induced murine colitis. Am. J. Physiol. Gastrointest. Liver. Physiol. 2003, 285, G235–G243. [Google Scholar] [PubMed]
- Ukil, A.; Maity, S.; Karmakar, S.; Datta, N.; Vedasiromoni, J.R.; Das, P.K. Curcumin, the major component of food flavour turmetic, reduces mucosal injury in trinitrobenzene sulphonic acid-induced colitis. Br. J. Pharmacol. 2003, 139, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Nones, K.; Dommels, Y.E.; Martell, S.; Butts, C.; McNabb, W.C.; Park, Z.A.; Zhu, S.; Hedderley, D.; Barnett, M.P.; Roy, N.C. The effects of dietary curcumin and rutin on colonic inflammation and gene expression in multidrug resistance gene-deficient (mdrla-/-) mice, a model of inflammatory bowel diseases. Br. J. Nutr. 2009, 101, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.Y.; Song, Y.J.; Kim, K.M.; Choe, Y.K.; Hwang, S.Y.; Kim, T.S. Curcumin inhibits Th1 cytokine profile in CD4 T cells by suppressing interleukin-12 production in macrophages. Br. J. Pharmacol. 1999, 128, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Deng, C.S.; Zheng, J.J.; Xia, J. Curcumin regulated shift from Th1 to Th2 in trinitrobenzene sulphonic acid-induced chronic colitis. Acta Pharmacol. Sin. 2006, 27, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Suskind, D.L.; Wahbeh, G.; Burpee, T.; Cohen, M.; Christie, D.; Webwe, W. Tolerability of curcumin in pediatric inflammatory bowel disease: A forced-dose titration study. J. Pediatr. Gastroneterol. Nutr. 2013, 56, 277–279. [Google Scholar] [CrossRef]
- Lahiff, C.; Moss, A.C.M. Curcumin for clinical and endoscopic remission in ulcerative colitis. Inflamm. Bowel Dis. 2011, 17, E66. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, C.; Celeghini, C.; Monasta, L.; Zauli, G. NF-κB pathways in hematological malignancies. Cell. Mol. Life Sci. 2014, 71, 2083–2102. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Karin, M. Missing pieces in the NF-kappaB puzzle. Cell 2002, 109, S81–S96. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti-inflammatory roles for NF-kappaB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Gilroy, D.W.; Colville-Nash, P.R.; Willoughby, D.A. Possible new role for NF-kappaB in the resolution of inflammation. Nat. Med. 2001, 7, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Mobasheri, A.; Matis, U.; Shakibaei, M. Curcumin mediated suppression of nuclear factor-κB promotes chondrogenic differentiation of mesenchymal stem cells in a high-density co-culture microenvironment. Arthritis Res. Ther. 2010, 12, R127. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; John, T.; Schulze-Tanzil, G.; Lehmann, I.; Mobasheri, A. Suppression of NF-kappaB activation by curcumin leads to inhibition of expression of cyclo-oxygenase-2 and matrix metalloproteinase-9 in human articular chondrocytes: Implications for the treatment of osteoarthritis. Biochem. Pharmacol. 2007, 73, 1434–1445. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Mobasheri, A.; Busch, F.; Aldinger, C.; Stahlmann, R.; Montaseri, A.; Shakibaei, M. Curcumin modulates nuclear factor kappaB (NF-kappaB)-mediated inflammation in human tenocytes in vitro: Role of the phosphatidylinositol 3-kinase/Akt pathway. J. Biol. Chem. 2011, 286, 28556–28566. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H. Discovery and development of natural product-derived chemotherapeutic agents based on a medicinal chemistry approach. J. Nat. Prod. 2010, 73, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Zhongfa, L.; Chiu, M.; Wang, J.; Chen, W.; Yen, W.; Fan-Havard, P.; Yee, LD.; Chan, K.K. Enhancement of curcumin oral absorption and pharmacokinetics of curcuminoids and curcumin metabolites in mice. Cancer Chemother. Pharmacol. 2012, 69, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Anticancer and carcinogenic properties of curcumin: considerations for its clinical development as a cancer chemopreventive and chemotherapeutic agent. Mol. Nutr. Food Res. 2008, 52, S103–S127. [Google Scholar] [PubMed]
- Xia, Y.Q.; Wei, X.Y.; Li, W.L.; Kanchana, K.; Xu, C.C.; Chen, D.H.; Chou, P.H.; Jin, R.; Wu, J.Z.; Liang, G. Curcumin Analogue A501 induces G2/M Arrest and Apoptosis in Non-small Cell Lung Cancer Cells. Asian Pac. J. Cancer Prev. 2014, 15, 6893–6898. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Lee, J.H.; Cheong, Y.K.; Chung, H.T.; Pae, H.O. Antidiabetic potential of the heme oxygenase-1 inducer curcumin analogues. Biomed. Res. Int. 2013, 2013, 918039. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Li, H.; Bai, H.; Su, Z.; Xiang, Q.; Wang, C.; Zhao, B.; Zhang, Y.; Zhang, Q.; Chu, Y.; et al. Synthesis of novel curcumin analogues for inhibition of 11β-hydroxysteroid dehydrogenase type 1 with anti-diabetic properties. Eur. J. Med. Chem. 2014, 77, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Anthwal, A.; Thakur, B.K.; Rawat, M.S.; Rawat, D.S.; Tyagi, A.K.; Aggarwal, B.B. Synthesis, Characterization and in Vitro Anticancer Activity of C-5 Curcumin Analogues with Potential to Inhibit TNF-α-Induced NF-κB Activation. BioMed Res. Int. 2014, 2014, 524161. [Google Scholar] [CrossRef] [PubMed]
- Sardjiman, S.S.; Reksohadiprodjo, M.S.; Hakim, L.; van der Goot, H.; Timmerman, H. 1,5-Diphenyl-1,4-pentadiene-3-ones and cyclic analogues as antioxidative agents. Synthesis and structure-activity relationship. Eur. J. Med. Chem. 1997, 32, 625–630. [Google Scholar]
- Di Bello, M.G.; Masini, E.; Ioannides, C.; Ndisang, J.F.; Raspanti, S.; Sacchi, T.B.; Mannaioni, P.F. Histamine release from rat mast cells induced by the metabolic activation of drugs of abuse into free radicals. Inflamm. Res. 1998, 47, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Mannaioni, P.F.; Bello, M.G.D.; Raspanti, S.; Mugnai, L.; Romano, V.; Masini, E. Free radical mediated release of histamine from rat mast cells induced by drugs of abuse. Inflamm. Res. 1996, 45, S25–S26. [Google Scholar] [CrossRef] [PubMed]
- Nugroho, A.E.; Ikawati, Z.; Sardjiman; Maeyama, K. Effects of benzylidenecyclopentanone analogues of curcumin on histamine release from mast cells. Biol. Pharm. Bull. 2009, 32, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Naksuriya, O.; Okonogi, S.; Schiffelers, R.M.; Hennink, W.E. Curcumin nanoformulations: A review of pharmaceutical properties and preclinical studies and clinical data related to cancer treatment. Biomaterials 2014, 35, 3365–3383. [Google Scholar] [CrossRef] [PubMed]
- Ghalandarlaki, N.; Alizadeh, A.M.; Ashkani-Esfahani, S. Nanotechnology-applied curcumin for different diseases therapy. BioMed Res. Int. 2014, 2014, 394264. [Google Scholar] [CrossRef] [PubMed]
- Beloqui, A.; Coco, R.; Memvanga, P.B.; Ucakar, B.; des Rieux, A.; Préat, V. pH-sensitive nanoparticles for colonic delivery of curcumin in inflammatory bowel disease. Int. J. Pharm. 2014, 473, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Gugulothu, D.; Kulkarni, A.; Patravale, V.; Dandekar, P. pH-sensitive nanoparticles of curcumin-celecoxib combination: Evaluating drug synergy in ulcerative colitis model. J. Pharm. Sci. 2014, 103, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Suresh, S.; Devi, K.; Yadav, S. Effect of cyclodextrin complexation of curcumin on its solubility and antiangiogenic and anti-inflammatory activity in rat colitis model. AAPS PharmSciTech 2009, 10, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples are not available from authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vecchi Brumatti, L.; Marcuzzi, A.; Tricarico, P.M.; Zanin, V.; Girardelli, M.; Bianco, A.M. Curcumin and Inflammatory Bowel Disease: Potential and Limits of Innovative Treatments. Molecules 2014, 19, 21127-21153. https://doi.org/10.3390/molecules191221127
Vecchi Brumatti L, Marcuzzi A, Tricarico PM, Zanin V, Girardelli M, Bianco AM. Curcumin and Inflammatory Bowel Disease: Potential and Limits of Innovative Treatments. Molecules. 2014; 19(12):21127-21153. https://doi.org/10.3390/molecules191221127
Chicago/Turabian StyleVecchi Brumatti, Liza, Annalisa Marcuzzi, Paola Maura Tricarico, Valentina Zanin, Martina Girardelli, and Anna Monica Bianco. 2014. "Curcumin and Inflammatory Bowel Disease: Potential and Limits of Innovative Treatments" Molecules 19, no. 12: 21127-21153. https://doi.org/10.3390/molecules191221127
APA StyleVecchi Brumatti, L., Marcuzzi, A., Tricarico, P. M., Zanin, V., Girardelli, M., & Bianco, A. M. (2014). Curcumin and Inflammatory Bowel Disease: Potential and Limits of Innovative Treatments. Molecules, 19(12), 21127-21153. https://doi.org/10.3390/molecules191221127