Plasticity of the Human Olfactory System: The Olfactory Bulb


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
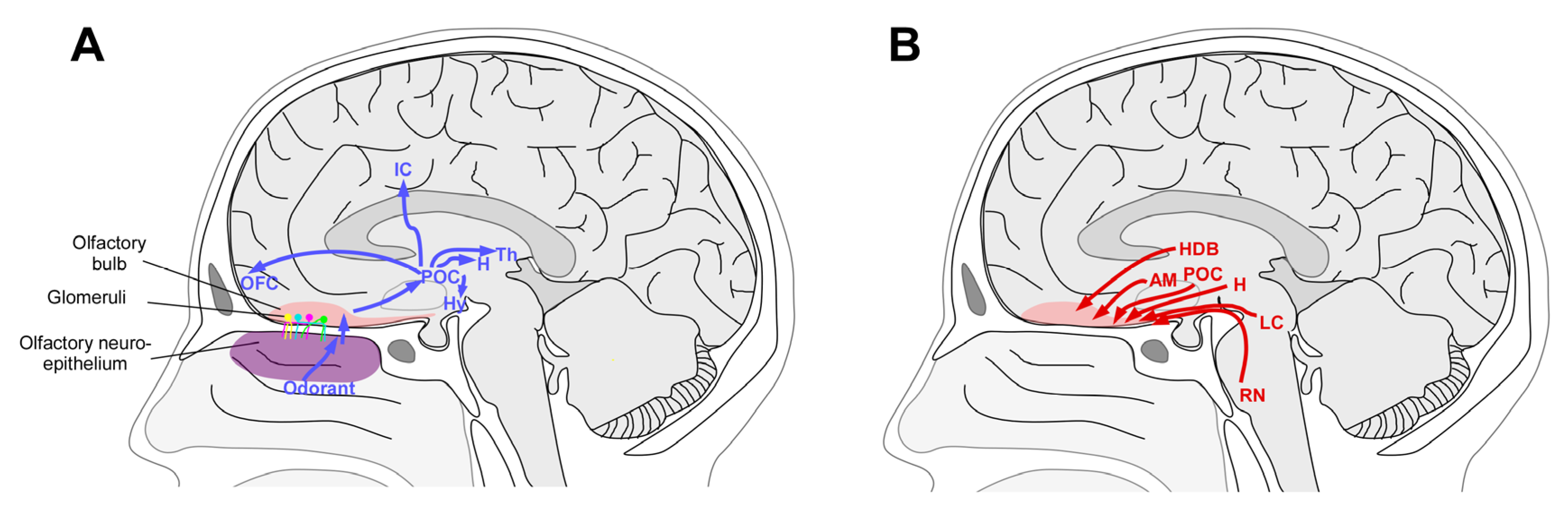
2. Anatomy and Physiology of the OB
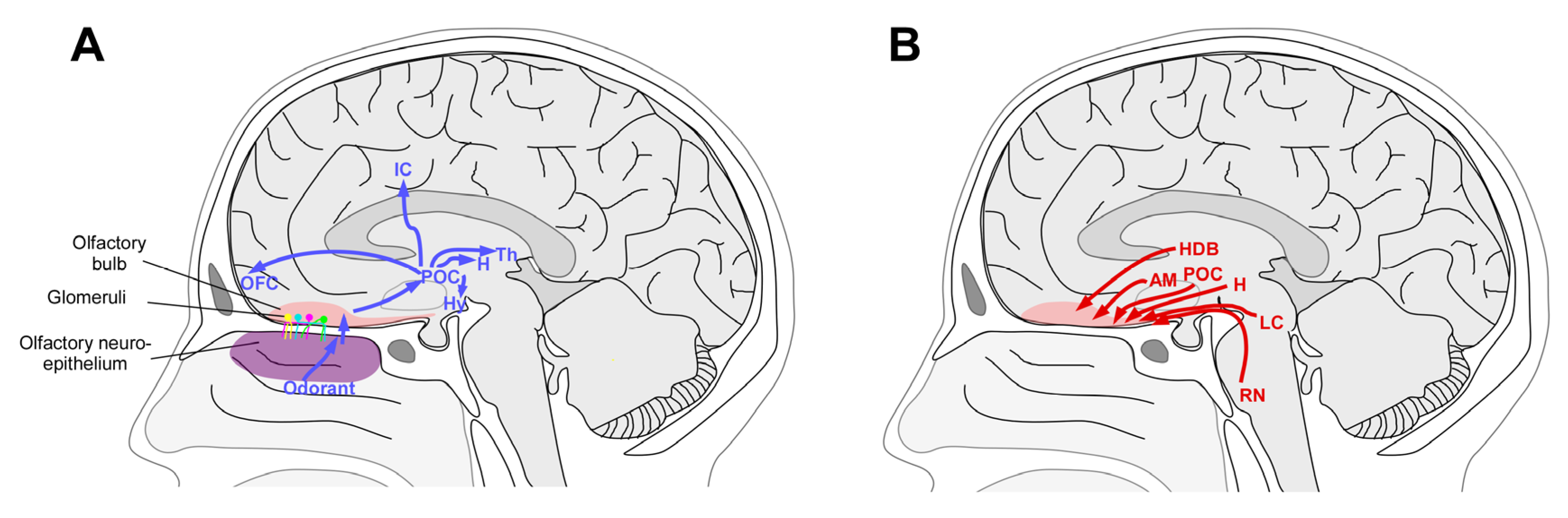
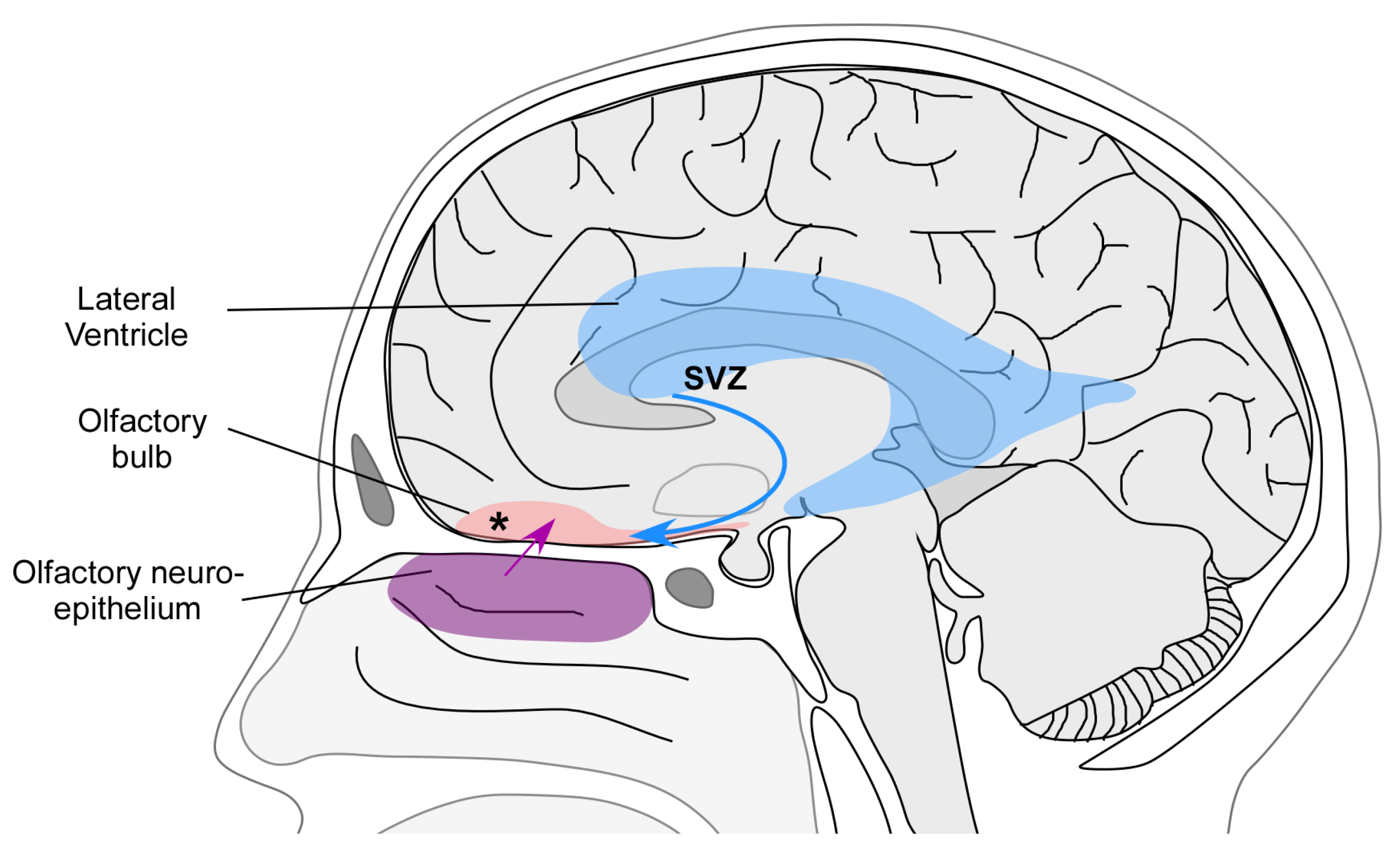
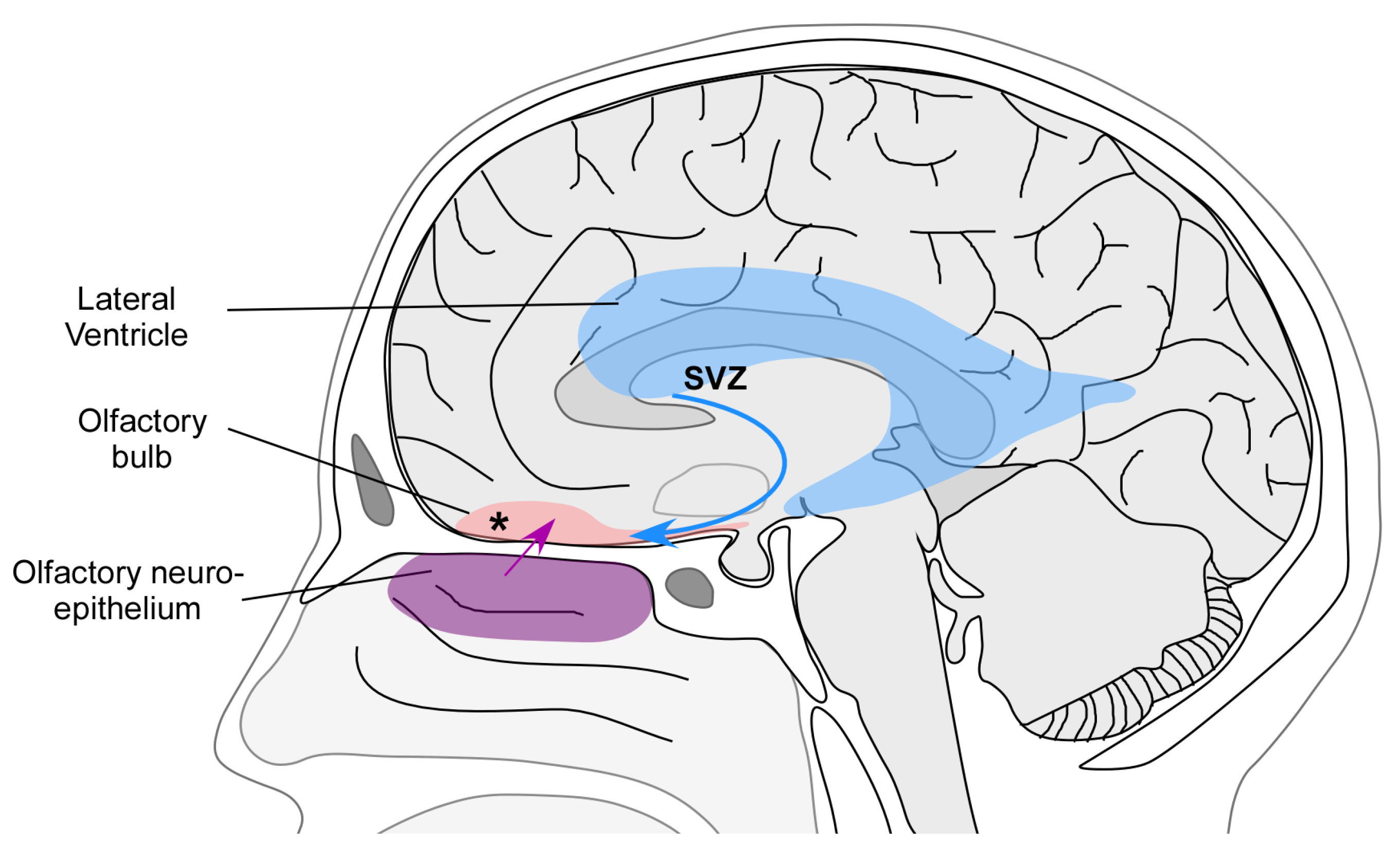
3. Mechanisms of Plasticity
4. Plasticity of the Human OB
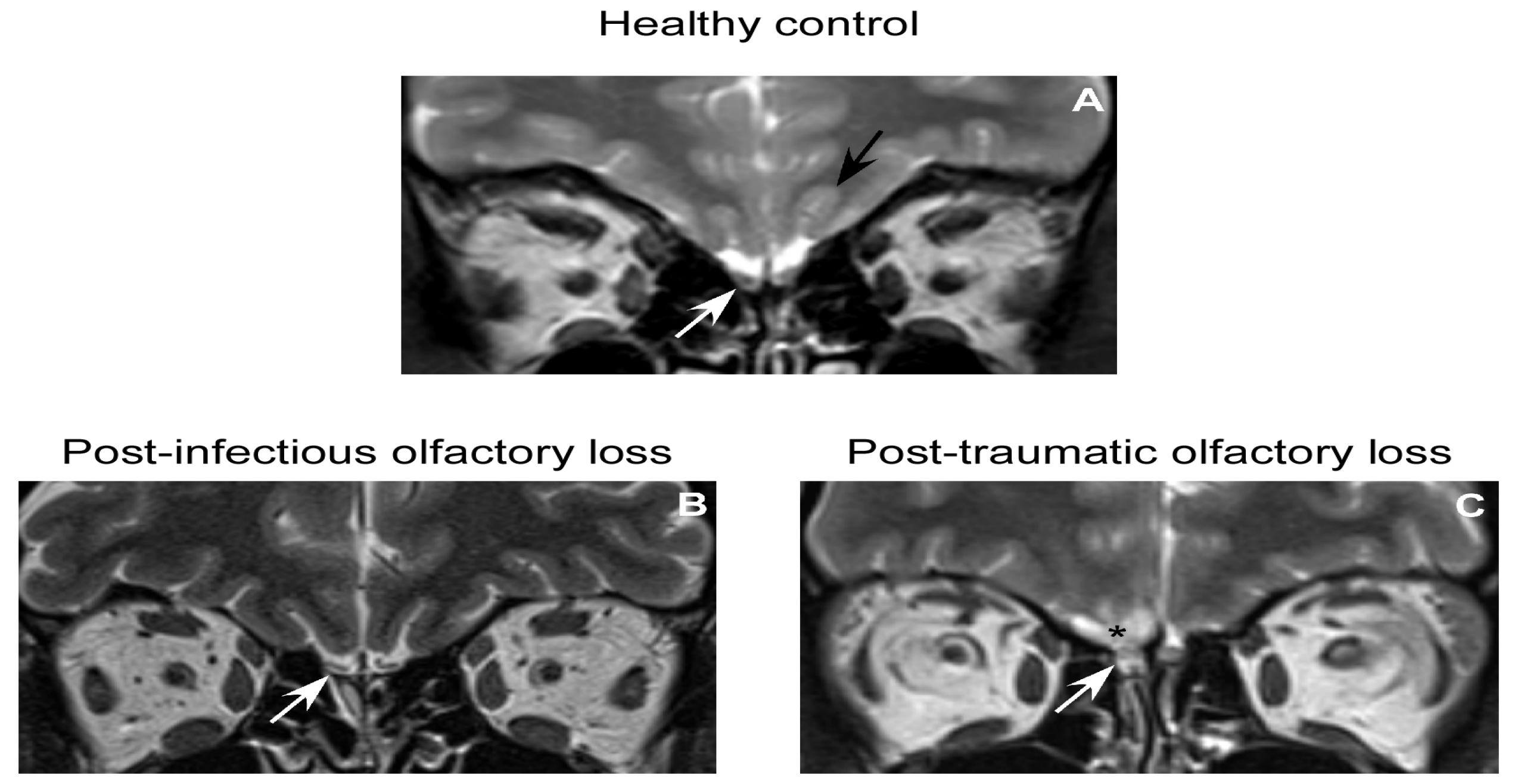
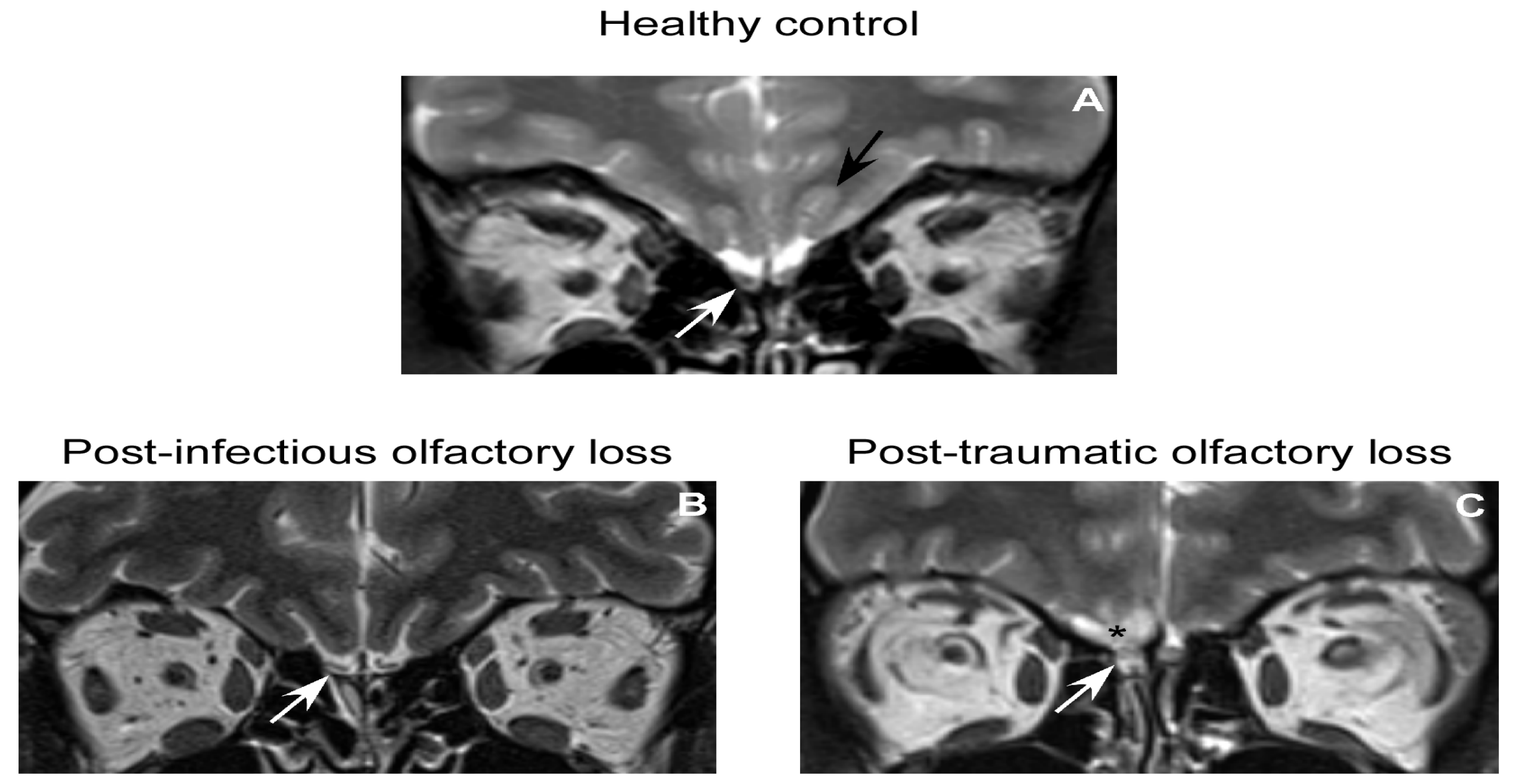
4.1. OB Volume as Measure of OB Function in Humans–Technical Details
4.2. OB in Healthy Subjects
4.3. OB in Patients
5. Conclusions
Conflicts of Interest
References
- Mueller, A.; Rodewald, A.; Reden, J.; Gerber, J.; von Kummer, R.; Hummel, T. Reduced olfactory bulb volume in post-traumatic and post-infectious olfactory dysfunction. Neuroreport 2005, 16, 475–478. [Google Scholar] [CrossRef]
- Rombaux, P.; Mouraux, A.; Bertrand, B.; Nicolas, G.; Duprez, T.; Hummel, T. Retronasal and orthonasal olfactory function in relation to olfactory bulb volume in patients with posttraumatic loss of smell. Laryngoscope 2006, 116, 901–905. [Google Scholar]
- Rombaux, P.; Mouraux, A.; Bertrand, B.; Nicolas, G.; Duprez, T.; Hummel, T. Olfactory function and olfactory bulb volume in patients with postinfectious olfactory loss. Laryngoscope 2006, 116, 436–439. [Google Scholar] [CrossRef]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; Kroger, H.; Doty, R.L. Posttraumatic smell loss: Relationship of psychophysical tests and volumes of the olfactory bulbs and tracts and the temporal lobes. Acad. Radiol. 1999, 6, 264–272. [Google Scholar] [CrossRef]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; McKeown, D.A.; Doty, R.L. Posttraumatic olfactory dysfunction: MR and clinical evaluation. AJNR. Am. J. Neuroradiol. 1996, 17, 1171–1179. [Google Scholar]
- Gudziol, V.; Buschhuter, D.; Abolmaali, N.; Gerber, J.; Rombaux, P.; Hummel, T. Increasing olfactory bulb volume due to treatment of chronic rhinosinusitis--a longitudinal study. Brain J. Neurol. 2009, 132, 3096–3101. [Google Scholar] [CrossRef]
- Lotsch, J.; Schaeffeler, E.; Mittelbronn, M.; Winter, S.; Gudziol, V.; Schwarzacher, S.W.; Hummel, T.; Doehring, A.; Schwab, M.; Ultsch, A. Functional genomics suggest neurogenesis in the adult human olfactory bulb. Brain Struct. Funct. 2013. [Google Scholar] [CrossRef]
- Bergmann, O.; Liebl, J.; Bernard, S.; Alkass, K.; Yeung, M.S.; Steier, P.; Kutschera, W.; Johnson, L.; Landen, M.; Druid, H.; et al. The age of olfactory bulb neurons in humans. Neuron 2012, 74, 634–639. [Google Scholar] [CrossRef]
- Curtis, M.A.; Kam, M.; Faull, R.L. Neurogenesis in humans. Eur. J. Neurosci. 2011, 33, 1170–1174. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Zhang, X.; Firestein, S. The olfactory receptor gene superfamily of the mouse. Nat. Neurosci. 2002, 5, 124–133. [Google Scholar]
- Crasto, C.; Marenco, L.; Miller, P.; Shepherd, G. Olfactory Receptor Database: A metadata-driven automated population from sources of gene and protein sequences. Nucleic Acids Res. 2002, 30, 354–360. [Google Scholar] [CrossRef]
- Mombaerts, P.; Wang, F.; Dulac, C.; Chao, S.K.; Nemes, A.; Mendelsohn, M.; Edmondson, J.; Axel, R. Visualizing an olfactory sensory map. Cell 1996, 87, 675–686. [Google Scholar] [CrossRef]
- Berkowicz, D.A.; Trombley, P.Q.; Shepherd, G.M. Evidence for glutamate as the olfactory receptor cell neurotransmitter. J. Neurophysiol. 1994, 71, 2557–2561. [Google Scholar]
- Gutierrez-Mecinas, M.; Crespo, C.; Blasco-Ibanez, J.M.; Gracia-Llanes, F.J.; Marques-Mari, A.I.; Nacher, J.; Varea, E.; Martinez-Guijarro, F.J. Distribution of D2 dopamine receptor in the olfactory glomeruli of the rat olfactory bulb. Eur. J. Neurosci. 2005, 22, 1357–1367. [Google Scholar] [CrossRef]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobiol. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef]
- Kasa, P.; Rakonczay, Z.; Gulya, K. The cholinergic system in Alzheimer’s disease. Progr. Neurobiol. 1997, 52, 511–535. [Google Scholar] [CrossRef]
- Woolf, N.J. Cholinergic systems in mammalian brain and spinal cord. Progr. Neurobiol. 1991, 37, 475–524. [Google Scholar] [CrossRef]
- Brennan, P.; Kaba, H.; Keverne, E.B. Olfactory recognition: A simple memory system. Science 1990, 250, 1223–1226. [Google Scholar]
- Sullivan, R.M.; Zyzak, D.R.; Skierkowski, P.; Wilson, D.A. The role of olfactory bulb norepinephrine in early olfactory learning. Brain Res. Dev. Brain Res. 1992, 70, 279–282. [Google Scholar] [CrossRef]
- Varendi, H.; Porter, R.H.; Winberg, J. The effect of labor on olfactory exposure learning within the first postnatal hour. Behav. Neurosci. 2002, 116, 206–211. [Google Scholar] [CrossRef]
- Winberg, J.; Porter, R.H. Olfaction and human neonatal behaviour: Clinical implications. Acta Paediatr. 1998, 87, 6–10. [Google Scholar] [CrossRef]
- Lazarini, F.; Lledo, P.M. Is adult neurogenesis essential for olfaction? Trends Neurosci. 2011, 34, 20–30. [Google Scholar] [CrossRef]
- Cleland, T.A.; Linster, C. Computation in the olfactory system. Chem. Senses 2005, 30, 801–813. [Google Scholar] [CrossRef]
- Schwob, J.E. Neural regeneration and the peripheral olfactory system. Anat. Record 2002, 269, 33–49. [Google Scholar] [CrossRef]
- Lois, C.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Chain migration of neuronal precursors. Science 1996, 271, 978–981. [Google Scholar]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef]
- Kornack, D.R.; Rakic, P. The generation, migration, and differentiation of olfactory neurons in the adult primate brain. Proc. Natl. Acad. Sci. USA 2001, 98, 4752–4757. [Google Scholar] [CrossRef]
- Alvarez-Buylla, A.; Kohwi, M.; Nguyen, T.M.; Merkle, F.T. The heterogeneity of adult neural stem cells and the emerging complexity of their niche. Cold Spring Harb. Symp. Quant. Biol. 2008, 73, 357–365. [Google Scholar] [CrossRef]
- Doetsch, F.; Caille, I.; Lim, D.A.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 1999, 97, 703–716. [Google Scholar] [CrossRef]
- Carlen, M.; Cassidy, R.M.; Brismar, H.; Smith, G.A.; Enquist, L.W.; Frisen, J. Functional integration of adult-born neurons. Curr. Biol. 2002, 12, 606–608. [Google Scholar] [CrossRef]
- Johansson, C.B.; Svensson, M.; Wallstedt, L.; Janson, A.M.; Frisen, J. Neural stem cells in the adult human brain. Exp. Cell Res. 1999, 253, 733–736. [Google Scholar] [CrossRef]
- Sanai, N.; Tramontin, A.D.; Quinones-Hinojosa, A.; Barbaro, N.M.; Gupta, N.; Kunwar, S.; Lawton, M.T.; McDermott, M.W.; Parsa, A.T.; Garcia-Verdugo, J.M.; et al. Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 2004, 427, 740–744. [Google Scholar] [CrossRef]
- Curtis, M.A.; Kam, M.; Nannmark, U.; Anderson, M.F.; Axell, M.Z.; Wikkelso, C.; Holtas, S.; van Roon-Mom, W.M.; Bjork-Eriksson, T.; Nordborg, C.; et al. Human neuroblasts migrate to the olfactory bulb via a lateral ventricular extension. Science 2007, 315, 1243–1249. [Google Scholar] [CrossRef]
- Guerrero-Cazares, H.; Gonzalez-Perez, O.; Soriano-Navarro, M.; Zamora-Berridi, G.; Garcia-Verdugo, J.M.; Quinones-Hinojosa, A. Cytoarchitecture of the lateral ganglionic eminence and rostral extension of the lateral ventricle in the human fetal brain. J. Comp. Neurol. 2011, 519, 1165–1180. [Google Scholar] [CrossRef]
- Sanai, N.; Nguyen, T.; Ihrie, R.A.; Mirzadeh, Z.; Tsai, H.H.; Wong, M.; Gupta, N.; Berger, M.S.; Huang, E.; Garcia-Verdugo, J.M.; et al. Corridors of migrating neurons in the human brain and their decline during infancy. Nature 2011, 478, 382–386. [Google Scholar] [CrossRef]
- Wang, C.; Liu, F.; Liu, Y.Y.; Zhao, C.H.; You, Y.; Wang, L.; Zhang, J.; Wei, B.; Ma, T.; Zhang, Q.; et al. Identification and characterization of neuroblasts in the subventricular zone and rostral migratory stream of the adult human brain. Cell Res. 2011, 21, 1534–1550. [Google Scholar] [CrossRef]
- Bedard, A.; Parent, A. Evidence of newly generated neurons in the human olfactory bulb. Brain Res. Dev. Brain Res. 2004, 151, 159–168. [Google Scholar] [CrossRef]
- Kaneko, N.; Marin, O.; Koike, M.; Hirota, Y.; Uchiyama, Y.; Wu, J.Y.; Lu, Q.; Tessier-Lavigne, M.; Alvarez-Buylla, A.; Okano, H.; et al. New neurons clear the path of astrocytic processes for their rapid migration in the adult brain. Neuron 2010, 67, 213–223. [Google Scholar] [CrossRef]
- Gritti, A.; Bonfanti, L.; Doetsch, F.; Caille, I.; Alvarez-Buylla, A.; Lim, D.A.; Galli, R.; Verdugo, J.M.; Herrera, D.G.; Vescovi, A.L. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 437–445. [Google Scholar]
- Pagano, S.F.; Impagnatiello, F.; Girelli, M.; Cova, L.; Grioni, E.; Onofri, M.; Cavallaro, M.; Etteri, S.; Vitello, F.; Giombini, S.; et al. Isolation and characterization of neural stem cells from the adult human olfactory bulb. Stem Cells 2000, 18, 295–300. [Google Scholar] [CrossRef]
- Brennan, P.A.; Kendrick, K.M.; Keverne, E.B. Neurotransmitter release in the accessory olfactory bulb during and after the formation of an olfactory memory in mice. Neuroscience 1995, 69, 1075–1086. [Google Scholar] [CrossRef]
- Brennan, P.A.; Keverne, E.B. Neural mechanisms of mammalian olfactory learning. Progr. Neurobiol. 1997, 51, 457–481. [Google Scholar] [CrossRef]
- Rombaux, P.; Grandin, C.; Duprez, T. How to measure olfactory bulb volume and olfactory sulcus depth? B-ENT 2009, 5, 53–60. [Google Scholar]
- Duprez, T.P.; Rombaux, P. Imaging the olfactory tract (cranial nerve #1). Eur. J. Radiol. 2010, 74, 288–298. [Google Scholar] [CrossRef]
- Buschhuter, D.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Witt, M.; Abolmaali, N.D.; Hummel, T. Correlation between olfactory bulb volume and olfactory function. NeuroImage 2008, 42, 498–502. [Google Scholar] [CrossRef]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; Doty, R.L. Olfactory bulb and tract and temporal lobe volumes. Normative data across decades. Ann. N.Y. Acad. Sci. 1998, 855, 546–555. [Google Scholar] [CrossRef]
- Hummel, T.; Kobal, G.; Gudziol, H.; Mackay-Sim, A. Normative data for the “Sniffin’ Sticks” including tests of odor identification, odor discrimination, and olfactory thresholds: An upgrade based on a group of more than 3,000 subjects. Eur. Arch. Otorhinolaryngol. 2007, 264, 237–243. [Google Scholar] [CrossRef]
- Murphy, C.; Schubert, C.R.; Cruickshanks, K.J.; Klein, B.E.; Klein, R.; Nondahl, D.M. Prevalence of olfactory impairment in older adults. JAMA 2002, 288, 2307–2312. [Google Scholar] [CrossRef]
- Rawson, N.E. Olfactory loss in aging. Sci. Ag. Knowl. Environ. 2006, 2006, pe6. [Google Scholar] [CrossRef]
- Naessen, R. An enquiry on the morphological characteristics and possible changes with age in the olfactory region of man. Acta Oto-Laryngol. 1971, 71, 49–62. [Google Scholar] [CrossRef]
- Conley, D.B.; Robinson, A.M.; Shinners, M.J.; Kern, R.C. Age-related olfactory dysfunction: Cellular and molecular characterization in the rat. Am. J. Rhinol. 2003, 17, 169–175. [Google Scholar]
- Paik, S.I.; Lehman, M.N.; Seiden, A.M.; Duncan, H.J.; Smith, D.V. Human olfactory biopsy. The influence of age and receptor distribution. Arch. Otolaryngol.--Head Neck Surg. 1992, 118, 731–738. [Google Scholar] [CrossRef]
- Chen, Y.; Getchell, T.V.; Sparks, D.L.; Getchell, M.L. Patterns of adrenergic and peptidergic innervation in human olfactory mucosa: Age-related trends. J. Comp. Neurol. 1993, 334, 104–116. [Google Scholar] [CrossRef]
- Bhatnagar, K.P.; Kennedy, R.C.; Baron, G.; Greenberg, R.A. Number of mitral cells and the bulb volume in the aging human olfactory bulb: A quantitative morphological study. Anat. Record 1987, 218, 73–87. [Google Scholar] [CrossRef]
- Meisami, E.; Mikhail, L.; Baim, D.; Bhatnagar, K.P. Human olfactory bulb: Aging of glomeruli and mitral cells and a search for the accessory olfactory bulb. Ann. N. Y. Acad. Sci. 1998, 855, 708–715. [Google Scholar] [CrossRef]
- Kovacs, T.; Cairns, N.J.; Lantos, P.L. beta-amyloid deposition and neurofibrillary tangle formation in the olfactory bulb in ageing and Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 1999, 25, 481–491. [Google Scholar] [CrossRef]
- Hoogland, P.V.; van den Berg, R.; Huisman, E. Misrouted olfactory fibres and ectopic olfactory glomeruli in normal humans and in Parkinson and Alzheimer patients. Neuropathol. Appl. Neurobiol. 2003, 29, 303–311. [Google Scholar] [CrossRef]
- Attems, J.; Lintner, F.; Jellinger, K.A. Olfactory involvement in aging and Alzheimer’s disease: An autopsy study. J. Alzheimer’s Dis. JAD 2005, 7, 149–157; discussion 173–180. [Google Scholar]
- Hummel, T.; Heilmann, S.; Murphy, C. Age-Related Changes of Chemosensory Functions. In Olfaction, Taste, and Cognition; Rouby, C., Schaal, B., Dubois, D., Gervais, R., Holley, A., Eds.; Cambridge University Press: New York, NY, USA, 2002; pp. 441–456. [Google Scholar]
- Hornung, D.E.; Kurtz, D.B.; Bradshaw, C.B.; Seipel, D.M.; Kent, P.F.; Blair, D.C.; Emko, P. The olfactory loss that accompanies an HIV infection. Physiol. Behav. 1998, 64, 549–556. [Google Scholar] [CrossRef]
- Jones-Gotman, M.; Zatorre, R.J. Olfactory identification deficits in patients with focal cerebral excision. Neuropsychologia 1988, 26, 387–400. [Google Scholar] [CrossRef]
- Moberg, P.J.; Agrin, R.; Gur, R.E.; Gur, R.C.; Turetsky, B.I.; Doty, R.L. Olfactory dysfunction in schizophrenia: A qualitative and quantitative review. Neuropsychopharmacol. Off. Public. Am. Coll. Neuropsychopharmacol. 1999, 21, 325–340. [Google Scholar] [CrossRef]
- Smitka, M.; Abolmaali, N.; Witt, M.; Gerber, J.C.; Neuhuber, W.; Buschhueter, D.; Puschmann, S.; Hummel, T. Olfactory bulb ventricles as a frequent finding in magnetic resonance imaging studies of the olfactory system. Neuroscience 2009, 162, 482–485. [Google Scholar] [CrossRef]
- Burmeister, H.P.; Bitter, T.; Baltzer, P.A.; Dietzel, M.; Guntinas-Lichius, O.; Gudziol, H.; Kaiser, W.A. Olfactory bulb ventricles as a frequent finding--a myth or reality? Evaluation using high resolution 3 Tesla magnetic resonance imaging. Neuroscience 2011, 172, 547–553. [Google Scholar] [CrossRef]
- Burmeister, H.P.; Bitter, T.; Heiler, P.M.; Irintchev, A.; Frober, R.; Dietzel, M.; Baltzer, P.A.; Schad, L.R.; Reichenbach, J.R.; Gudziol, H.; et al. Imaging of lamination patterns of the adult human olfactory bulb and tract: In vitro comparison of standard- and high-resolution 3T MRI, and MR microscopy at 9.4 T. NeuroImage 2012, 60, 1662–1670. [Google Scholar] [CrossRef]
- Rombaux, P.; Potier, H.; Bertrand, B.; Duprez, T.; Hummel, T. Olfactory bulb volume in patients with sinonasal disease. Am. J. Rhinol. 2008, 22, 598–601. [Google Scholar]
- Rombaux, P.; Potier, H.; Markessis, E.; Duprez, T.; Hummel, T. Olfactory bulb volume and depth of olfactory sulcus in patients with idiopathic olfactory loss. Eur. Arch. Oto-Rhino-Laryngol. Off. J. Eur. Feder. Oto-Rhino-Laryngol. Soc. 2012, 267, 1551–1556. [Google Scholar]
- Wang, J.; You, H.; Liu, J.F.; Ni, D.F.; Zhang, Z.X.; Guan, J. Association of olfactory bulb volume and olfactory sulcus depth with olfactory function in patients with Parkinson disease. AJNR Am. J. Neuroradiol. 2011, 32, 677–681. [Google Scholar] [CrossRef]
- Thomann, P.A.; Dos Santos, V.; Toro, P.; Schonknecht, P.; Essig, M.; Schroder, J. Reduced olfactory bulb and tract volume in early Alzheimer’s disease--a MRI study. Neurobiol. Ag. 2009, 30, 838–841. [Google Scholar] [CrossRef]
- Mueller, A.; Abolmaali, N.D.; Hakimi, A.R.; Gloeckler, T.; Herting, B.; Reichmann, H.; Hummel, T. Olfactory bulb volumes in patients with idiopathic Parkinson’s disease a pilot study. J. Neural Transm. 2005, 112, 1363–1370. [Google Scholar] [CrossRef]
- Negoias, S.; Croy, I.; Gerber, J.; Puschmann, S.; Petrowski, K.; Joraschky, P.; Hummel, T. Reduced olfactory bulb volume and olfactory sensitivity in patients with acute major depression. Neuroscience 2010, 169, 415–421. [Google Scholar] [CrossRef]
- Veyseller, B.; Aksoy, F.; Yildirim, Y.S.; Bayraktar, F.G.; Gurbuz, D.; Savas, Y.; Ozturan, O. Reduced olfactory bulb volume in total laryngectomy patients: A magnetic resonance imaging study. Rhinology 2011, 49, 112–116. [Google Scholar]
- Haehner, A.; Rodewald, A.; Gerber, J.C.; Hummel, T. Correlation of olfactory function with changes in the volume of the human olfactory bulb. Arch. Otolaryngol.--Head Neck Surg. 2008, 134, 621–624. [Google Scholar] [CrossRef]
- Shiga, H.; Taki, J.; Washiyama, K.; Yamamoto, J.; Kinase, S.; Okuda, K.; Kinuya, S.; Watanabe, N.; Tonami, H.; Koshida, K.; et al. Assessment of olfactory nerve by SPECT-MRI image with nasal thallium-201 administration in patients with olfactory impairments in comparison to healthy volunteers. PLoS One 2013, 8, e57671. [Google Scholar] [CrossRef]
- Hummel, T.; Haehner, A.; Hummel, C.; Croy, I.; Iannilli, E. Lateralized differences in olfactory bulb volume relate to lateralized differences in olfactory function. Neuroscience 2013, 237, 51–55. [Google Scholar] [CrossRef]
- Benson, T.E.; Ryugo, D.K.; Hinds, J.W. Effects of sensory deprivation on the developing mouse olfactory system: A light and electron microscopic, morphometric analysis. J. Neurosci. Off. J. Soc. Neurosci. 1984, 4, 638–653. [Google Scholar]
- Cummings, D.M.; Brunjes, P.C. The effects of variable periods of functional deprivation on olfactory bulb development in rats. Exp. Neurol. 1997, 148, 360–366. [Google Scholar] [CrossRef]
- Von Gudden, B. Experimentaluntersuchungen ueber das periphere und zentrale Nervensystem. Archiv f Psychiatrie u Nervenkrankheiten 1870, 21, 693–723. [Google Scholar] [CrossRef]
- McLean, J.H.; Darby-King, A.; Bonnell, W.S. Neonatal olfactory sensory deprivation decreases BDNF in the olfactory bulb of the rat. Brain Res. Dev. Brain Res. 2001, 128, 17–24. [Google Scholar] [CrossRef]
- Lewin, G.R.; Barde, Y.A. Physiology of the neurotrophins. Ann. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef]
- Hummel, T.; Henkel, S.; Negoias, S.; Galvan, J.R.; Bogdanov, V.; Hopp, P.; Hallmeyer-Elgner, S.; Gerber, J.; Reuner, U.; Haehner, A. Olfactory bulb volume in patients with temporal lobe epilepsy. J. Neurol. 2013, 260, 1004–1008. [Google Scholar] [CrossRef]
- Goektas, O.; Schmidt, F.; Bohner, G.; Erb, K.; Ludemann, L.; Dahlslett, B.; Harms, L.; Fleiner, F. Olfactory bulb volume and olfactory function in patients with multiple sclerosis. Rhinology 2011, 49, 221–226. [Google Scholar]
- Mineur, Y.S.; Belzung, C.; Crusio, W.E. Functional implications of decreases in neurogenesis following chronic mild stress in mice. Neuroscience 2007, 150, 251–259. [Google Scholar] [CrossRef]
- Rombaux, P.; Huart, C.; de Volder, A.G.; Cuevas, I.; Renier, L.; Duprez, T.; Grandin, C. Increased olfactory bulb volume and olfactory function in early blind subjects. Neuroreport 2010, 21, 1069–1073. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huart, C.; Rombaux, P.; Hummel, T. Plasticity of the Human Olfactory System: The Olfactory Bulb. Molecules 2013, 18, 11586-11600. https://doi.org/10.3390/molecules180911586
Huart C, Rombaux P, Hummel T. Plasticity of the Human Olfactory System: The Olfactory Bulb. Molecules. 2013; 18(9):11586-11600. https://doi.org/10.3390/molecules180911586
Chicago/Turabian StyleHuart, Caroline, Philippe Rombaux, and Thomas Hummel. 2013. "Plasticity of the Human Olfactory System: The Olfactory Bulb" Molecules 18, no. 9: 11586-11600. https://doi.org/10.3390/molecules180911586
APA StyleHuart, C., Rombaux, P., & Hummel, T. (2013). Plasticity of the Human Olfactory System: The Olfactory Bulb. Molecules, 18(9), 11586-11600. https://doi.org/10.3390/molecules180911586