Fructus Gardenia Extract Ameliorates Oxonate-Induced Hyperuricemia with Renal Dysfunction in Mice by Regulating Organic Ion Transporters and mOIT3

Abstract
:1. Introduction
2. Results
2.1. Effects of FGE on Serum Uric Acid Levels, Renal Excretion of Uric Acid and Renal Function
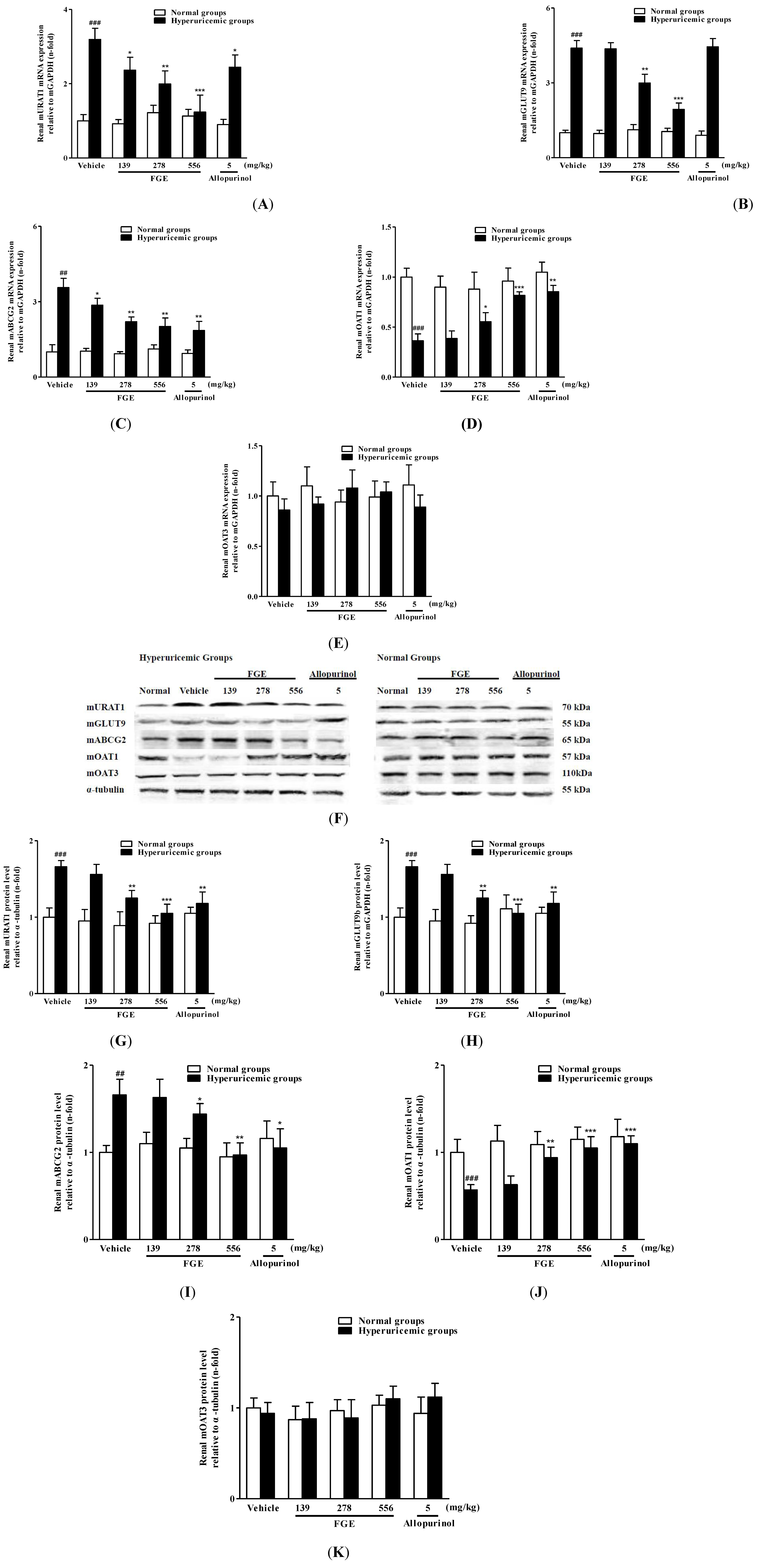
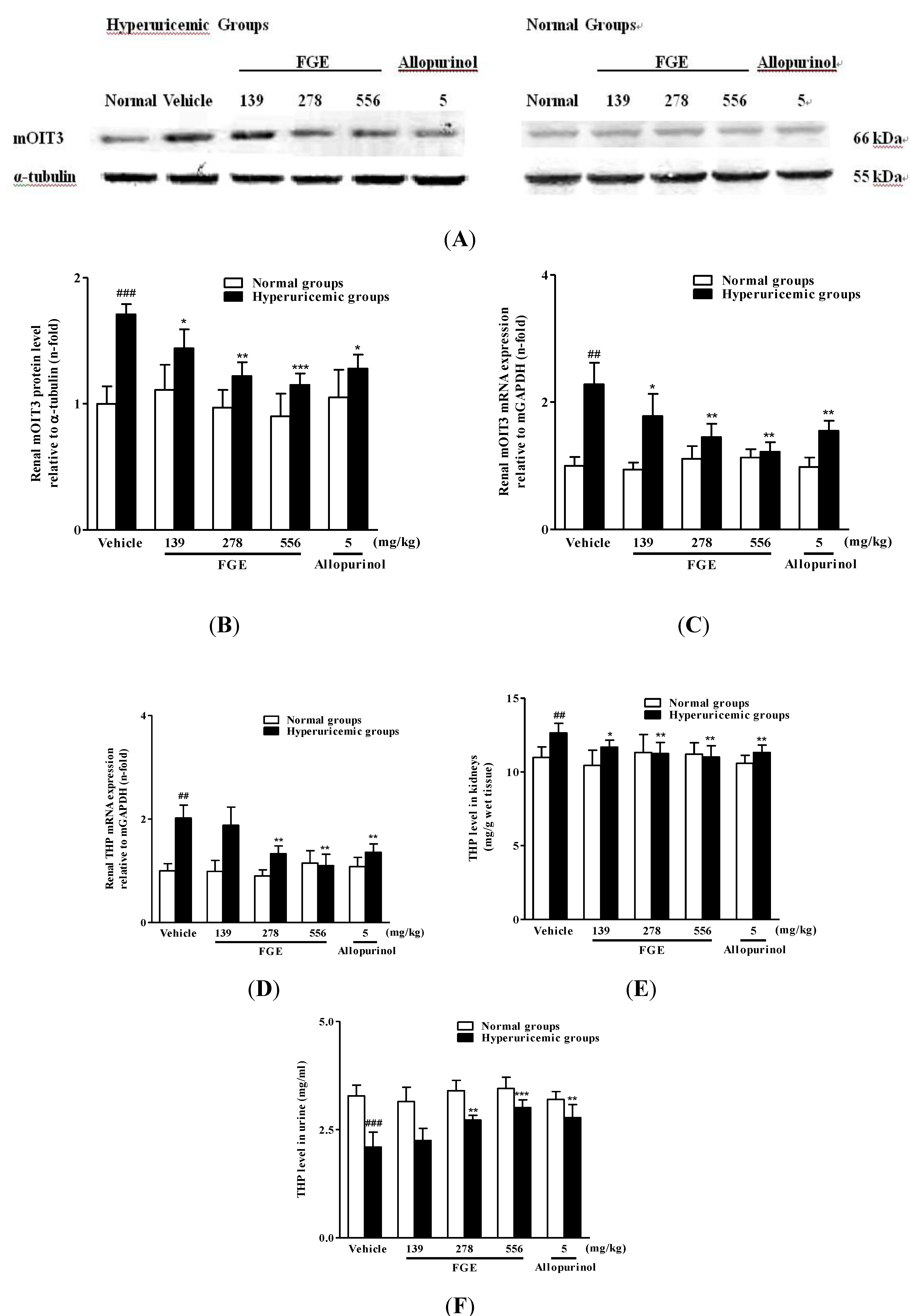
2.2. Effects of FGE on Renal mRNA and Protein Levels of Urate Transporters

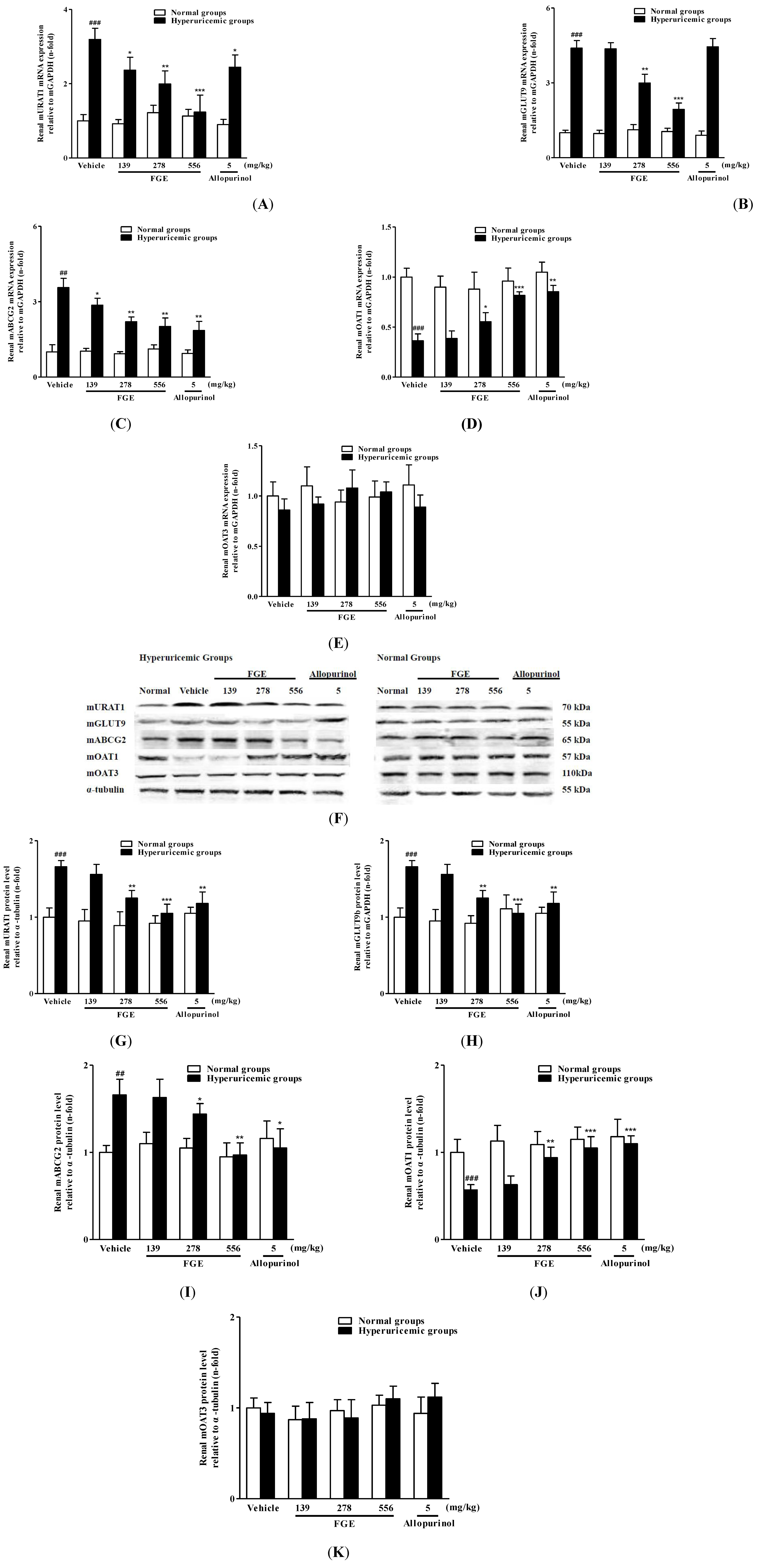{kind=link}
{kind=link}
{kind=link}
| Dose (mg/kg) | SUA (mg/dL) | SCr (mg/dL) | UUA (mg) | UCr (mg) | FEUA | BUN (mg/dL) | |
|---|---|---|---|---|---|---|---|
| Normal | |||||||
| Vehicle | - | 3.88 ± 0.14 | 0.92 ± 0.08 | 0.398 ± 0.017 | 0.55 ± 0.05 | 19.0 ± 3.3 | 13.3 ± 0.3 |
| FGE | 139 | 3.96 ± 0.18 | 0.88 ± 0.05 | 0.373 ± 0.028 | 0.30 ± 0.04 | 18.4 ± 2.4 | 13.1 ± 0.2 |
| 278 | 4.10 ± 0.15 | 0.95 ± 0.11 | 0.390 ± 0.016 | 0.38 ± 0.05 | 19.4 ± 2.5 | 14.0 ± 1.1 | |
| 556 | 3.77 ± 0.20 | 0.91 ± 0.07 | 0.351 ± 0.031 | 0.51 ± 0.07 | 18.2 ± 2.7 | 13.5 ± 0.4 | |
| 5 | 3.02 ± 0.12 *** | 0.95 ± 0.07 | 0.389 ± 0.024 | 0.52 ± 0.01 | 19.5 ± 3.1 | 12.9 ± 0.6 | |
| Hyperuricemia | |||||||
| Vehicle | 5.31 ± 0.21 ### | 1.25 ± 0.06 ### | 0.215 ± 0.045 ### | 0.28 ± 0.02 ### | 14.3 ± 5.6 ## | 16.5 ± 0.4 ### | |
| FGE | 139 | 4.36 ± 0.14 ** | 1.08 ± 0.06 * | 0.243 ± 0.008 | 0.30 ± 0.04 | 16.4 ± 2.8 * | 15.3 ± 0.2 * |
| 278 | 4.20 ± 0.12 *** | 1.05 ± 0.09 ** | 0.290 ± 0.026 ** | 0.38 ± 0.05 * | 17.0 ± 1.8 * | 14.9 ± 0.5 ** | |
| 556 | 4.01 ± 0.18 *** | 0.93 ± 0.02 *** | 0.371 ± 0.013 *** | 0.51 ± 0.07 ** | 18.2 ± 0.7 ** | 13.5 ± 0.1 *** | |
| AP | 5 | 3.58 ± 0.18 *** | 1.03 ± 0.05 ** | 0.359 ± 0.020 *** | 0.52 ± 0.01 *** | 19.1 ± 4.0 ** | 13.9 ± 0.3 *** |
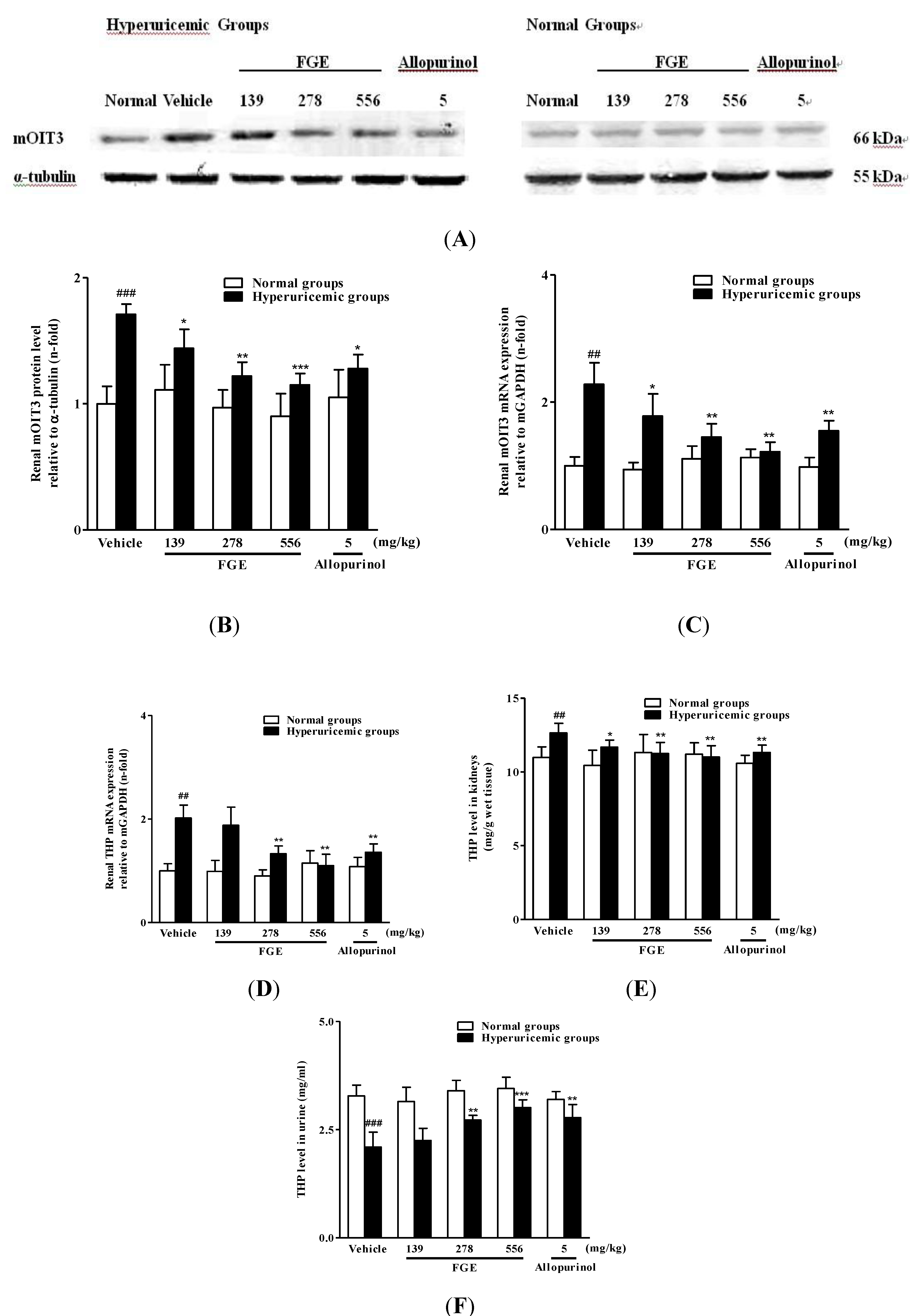
2.3. Effects of FGE on Renal mRNA and Protein Levels of mOIT3 and THP Concentrations
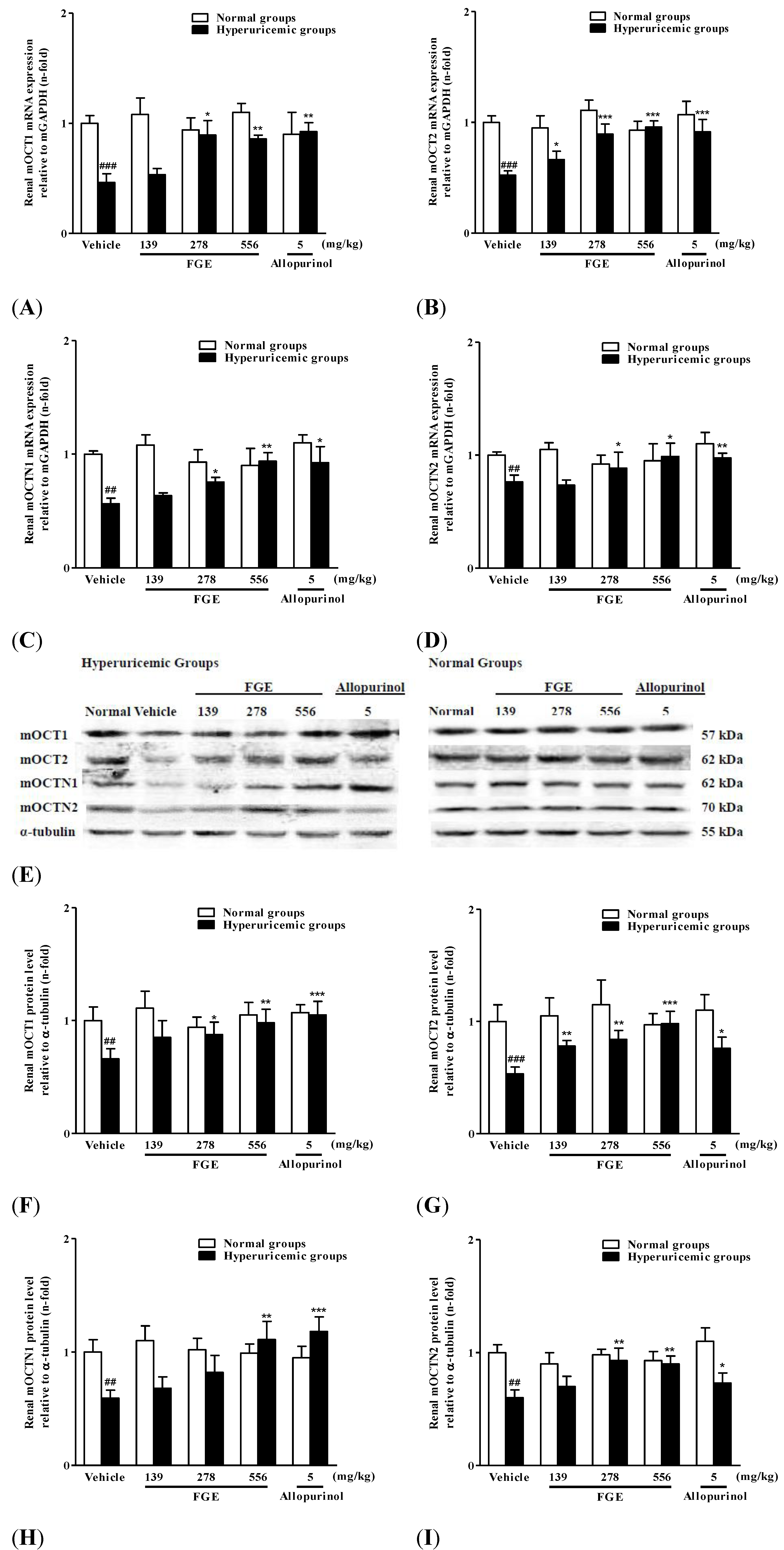
2.4. Effects of FGE on Renal mRNA and Protein Levels of Organic Cation/Carnitine Transporters

3. Discussion
4. Experimental
4.1. Reagents
4.2. Plant Material
4.3. Preparation of the Ethanol Extract
4.4. Animals
4.5. Hyperuricemic Mice and Drug Administration
4.6. Blood, Urine and Tissue Sample Collection
4.7. Determination of Uric Acid and Creatinine Levels and Renal Urate Handling Parameters
4.8. Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
| Description | Genebank | primer (5′→ 3′) | Product size (bp) | Tm (°C) |
|---|---|---|---|---|
| mURAT1 | NM_009203 | GCTACCAGAATCGGCACGCT CACCGGGAAGTCCACAATCC | 342 | 58 |
| mGLUT9 | NM_001102414 | GAGATGCTCATTGTGGGACG GTGCTACTTCGTCCTCGGT | 316 | 58 |
| mABCG2 | NM_011920 | TAAATGGAGCACCTCAACCT GAGATGCCACGGATAAACTG | 238 | 58 |
| mOIT3 | NM_010959 | GCGCCATTGAAGTGAGTGTC CAGGTTGGGCACGTATCCTT | 305 | 58 |
| mOAT1 | NM_008766 | GCCTATGTGGGCACCTTGAT CTTGTTTCCCGTTGATGCGG | 238 | 58 |
| mOAT3 | NM_031194 | AAGAACATCTCTGTGAGGGTG GGCAAGATGAACCAAAACTGG | 297 | 58 |
| mOCT1 | NM_009202 | ACATCCATGTTGCTCTTTCG TTGCTCCATTATCCTTACCG | 315 | 58 |
| mOCT2 | NM_013667 | ACAGGTTTGGGCGGAAGT CACCAGAAATAGAGCAGGAAG | 331 | 58 |
| mOCTN1 | NM_019687 | AGGAGAGGTGGAAACATGCG TCCTTCGTCTCCAAGGGGAT | 233 | 58 |
| mOCTN2 | NM_011396 | CTTATTCCCATACGGGCGCT TTTCTGAGGCACCTGTCGTC | 279 | 58 |
| mGAPDH | NM_008084 | TGAGGCCGGTGCTGAGTATGT CAGTCTTCTGGGTGGCAGTGAT | 299 | 58 |
4.9. Western Blot Analysis of mURAT1, mABCG2, mGLUT9, mOIT3, mOAT1, mOCT1, mOCT2, mOCTN1, mOCTN2
| Company | Description | Catalog number |
|---|---|---|
| SaiChi Biotech | Rabbit murat1 antibody | 001052-R |
| (Beijing, China) | Rabbit mglut9 antibody | 001046-R |
| Rabbit moat1 antibody | 001019-R | |
| Rabbit moat3 antibody | 001020-R | |
| Rabbit moct1 antibody | 001017-R | |
| Rabbit moct2 antibody | 001018-R | |
| Alpha Diagnostic International | Rabbit moctn1 antibody | OCTN11-A |
| Inc. (San Antonio, TX, USA) | Rabbit moctn2 antibody | OCTN21-A |
| Cell Signaling Technology Inc. | ||
| (Boston, MA, USA) | Rabbit α-tubulin antibody | #2148 |
| Epitomics Inc. | Rabbit moit3 antibody | 6862-1 |
| (Hangzhou, China) | Goat anti-rabbit igg HRP | 3056-1 |
4.10. Determination of THP Levels
4.11. Statistical Analysis
5. Conclusion
Acknowledgments
Conflicts of Interest
References
- Gibson, T. Hyperuricemia, gout and the kidney. Curr. Opin. Rheumatol. 2012, 24, 127–131. [Google Scholar] [CrossRef]
- Simão, A.N.; Lozovoy, M.A.; Dichi, I. The uric acid metabolism pathway as a therapeutic target in hyperuricemia related to metabolic syndrome. Expert. Opin. Ther. Targets 2012, 16, 1175–1187. [Google Scholar] [CrossRef]
- Hikita, M.; Ohno, I.; Hosoya, T. Hyperuricemia and gout. Nihon. Rinsho. 2011, 69, 416–420. [Google Scholar]
- Kanai, Y. Molecular mechanisms of urate transport in renal tubules: localization and function of urate transporters. Nihon. Rinsho. 2008, 66, 659–666. [Google Scholar]
- Yamauchi, T.; Ueda, T. Primary hyperuricemia due to decreased renal uric acid excretion. Nihon. Rinsho. 2008, 66, 679–681. [Google Scholar]
- Bobulescu, I.A.; Moe, O.W. Renal transport of uric acid: evolving concepts and uncertainties. Adv. Chronic Kidney Dis. 2012, 19, 358–371. [Google Scholar] [CrossRef]
- Sato, M.; Wakayama, T.; Mamada, H.; Shirasaka, Y.; Nakanishi, T.; Tamai, I. Identification and functional characterization of uric acid transporter Urat1 (Slc22a12) in rats. Biochim. Biophys. Acta. 2011, 1808, 1441–1447. [Google Scholar] [CrossRef]
- Vitart, V.; Rudan, I.; Hayward, C.; Gray, N.K.; Floyd, J. SLC2A9 is a newly identified urate transporter influencing serum urate concentration, urate excretion and gout. Nat. Genet. 2008, 40, 437–442. [Google Scholar] [CrossRef]
- Keembiyehetty, C.; Augustin, R.; Carayannopoulos, M.O.; Steer, S.; Manolescu, A.; Cheeseman, C.I.; Moley, K.H. Mouse glucose transporter 9 splice variants are expressed in adult liver and kidney and are up-regulated in diabetes. Mol. Endocrinol. 2006, 20, 686–697. [Google Scholar]
- Augustin, R.; Carayannopoulos, M.O.; Dowd, L.O.; Phay, J.E.; Moley, J.F.; Moley, K.H. Identification and characterization of human glucose transporter-like protein-9 (GLUT9): Alternative splicing alters trafficking. J. Biol. Chem. 2004, 279, 16229–16236. [Google Scholar]
- Bibert, S.; Hess, S.K.; Firsov, D.; Thorens, B.; Geering, K.; Horisberger, J.D.; Bonny, O. Mouse GLUT9: evidences for a urate uniporter. Am. J. Physiol. Renal Physiol. 2009, 297, 612–619. [Google Scholar] [CrossRef]
- Nakayama, A.; Matsuo, H.; Takada, T.; Ichida, K.; Nakamura, T.; Ikebuchi, Y.; Ito, K.; Hosoya, T.; Kanai, Y.; Suzuki, H.; Shinomiya, N. ABCG2 is a high-capacity urate transporter and its genetic impairment increases serum uric acid levels in humans. Nucleo. Nucleot. Nucleic Acids 2011, 30, 1091–1097. [Google Scholar] [CrossRef]
- Eraly, S.A.; Vallon, V.; Rieg, T.; Gangoiti, J.A.; Wikoff, W.R.; Siuzdak, G.; Barshop, B.A.; Nigam, S.K. Multiple organic anion transporters contribute to net renal excretion of uric acid. Physiol. Genomics 2008, 33, 180–192. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Z.Z.; Huang, L.Y.; Shen, H.L.; Han, Z.G. OIT3 deficiency impairs uric acid reabsorption in renal tubule. FEBS Lett. 2012, 586, 760–765. [Google Scholar] [CrossRef]
- Urakami, Y.; Okuda, M.; Masuda, S.; Akazawa, M.; Saito, H.; Inui, K. Distinct characteristics of organic cation transporters, OCT1 and OCT2, in the basolateral membrane of renal tubules. Pharm. Res. 2011, 18, 1528–1534. [Google Scholar]
- Burckhardt, G.; Wolff, N.A. Structure of renal organic anion and cation transporters. Am. J. Physiol. Renal Physiol. 2000, 278, 853–866. [Google Scholar]
- Komazawa, H.; Yamaguchi, H.; Hidaka, K.; Ogura, J.; Kobayashi, M.; Iseki, K. Renal uptake of substrates for organic anion transporters Oat1 and Oat3 and organic cation transporters Oct1 and Oct2 is altered in rats with adenine-induced chronic renal failure. J. Pharm. Sci. 2013, 102, 1086–1094. [Google Scholar] [CrossRef]
- Ulu, R.; Dogukan, A.; Tuzcu, M.; Gencoglu, H.; Ulas, M.; Ilhan, N.; Muqbil, I.; Mohammad, R. M.; Kucuk, O.; Sahin, K. Regulation of renal organic anion and cation transporters by thymoquinone in cisplatin induced kidney injury. Food Chem. Toxicol. 2012, 50, 1675–1679. [Google Scholar] [CrossRef]
- Aleksunes, L.M.; Augustine, L.M.; Scheffer, G.L.; Cherrington, N.J.; Manautou, J.E. Renal xenobiotic transporters are differentially expressed in mice following cisplatin treatment. Toxicology 2008, 250, 82–88. [Google Scholar] [CrossRef]
- Rødevand, E.; Sletvold, O.; Kvande, K.T. Side effects off allopurinol. Tidsskr. Nor. Laegeforen 2004, 124, 2618–2619. [Google Scholar]
- Lee, M.H.; Graham, G.G.; Williams, K.M.; Day, R.O. A benefit-risk assessment of benzbromarone in the treatment of gout. Was its withdrawal from the market in the best interest of patients? Drug Saf. 2008, 31, 643–665. [Google Scholar] [CrossRef]
- Zhu, J.X.; Zhu, Y.Y.; Luo, G.M.; Zeng, J.X. Research on the effect of fructus gardenia extract on reducing uric acid in hyperuricemia mice and its action mechanism. J. Anhui Agri. Sci. 2012, 39, 22317–22318. [Google Scholar]
- Doshi, M.; Takiue, Y.; Saito, H.; Hosoyamada, M. The increased protein level of URAT1 was observed in obesity/metabolic syndrome model mice. Nucleos. Nucleot. Nucleic Acids 2011, 30, 1290–1294. [Google Scholar] [CrossRef]
- Dinour, D.; Gray, N.K.; Campbell, S.; Shu, X.; Sawyer, L.; Richardson, W.; Rechavi, G.; Amariglio, N.; Ganon, L.; Sela, B.A.; et al. Homozygous SLC2A9 mutations cause severe renal hypouricemia. J. Am. Soc. Nephrol. 2010, 21, 64–72. [Google Scholar] [CrossRef]
- Ichida, K.; Hosoyamada, M.; Hisatome, I.; Enomoto, A.; Hikita, M.; Endou, H.; Hosoya, T. Clinical and molecular analysis of patients with renal hypouricemia in Japan-influence of URAT1 gene on urinary urate excretion. J. Am. Soc. Nephrol. 2004, 15, 164–173. [Google Scholar] [CrossRef]
- Habu, Y.; Yano, I.; Takeuchi, A.; Saito, H.; Okuda, M.; Fukatsu, A.; Inui, K. Decreased activity of basolateral organic ion transports in hyperuricemic rat kidney: roles of organic ion transporters, rOAT1, rOAT3 and rOCT2. Biochem. Pharmacol. 2003, 66, 1107–1114. [Google Scholar] [CrossRef]
- Shi, Y.W.; Wang, C.P.; Wang, X.; Zhang, Y.L.; Liu, L.; Wang, R.W.; Ye, J.F.; Hu, L.S.; Kong, L.D. Uricosuric and nephroprotective properties of Ramulus Mori ethanol extract in hyperuricemic mice. J. Ethnopharmacol. 2012, 143, 896–904. [Google Scholar] [CrossRef]
- Hu, Q.H.; Jiao, R.Q.; Wang, X.; Lv, Y.Z.; Kong, L.D. Simiao pill ameliorates urate underexcretion and renal dysfunction in hyperuricemic mice. J. Ethnopharmacol. 2010, 128, 685–692. [Google Scholar] [CrossRef]
- Li, J.M.; Zhang, X.; Wang, X.; Xie, Y.C.; Kong, L.D. Protective effects of cortex fraxini coumarines against oxonate-induced hyperuricemia and renal dysfunction in mice. Eur. J. Pharmacol. 2011, 666, 192–204. [Google Scholar]
- Kemter, E.; Rathkolb, B.; Rozman, J.; Hans, W.; Schrewe, A.; Landbrecht, C.; Klaften, M.; Ivandic, B.; Fuchs, H.; Gailus-Durner, V.; et al. Novel missense mutation of uromodulin in mice causes renal dysfunction with alterations in urea handling, energy, and bone metabolism. Am. J. Physiol. Renal Physiol. 2009, 297, 1391–1398. [Google Scholar] [CrossRef]
- Dahan, K.; Devuyst, O.; Smaers, M.; Vertommen, D.; Loute, G.; Poux, J.M.; Viron, B.; Jacquot, C.; Gagnadoux, M.F.; Chauveau, D.; et al. A cluster of mutations in the UMOD gene causes familial juvenile hyperuricemic nephropathy with abnormal expression of uromodulin. J. Am. Soc. Nephrol. 2003, 14, 2883–2893. [Google Scholar] [CrossRef]
- Hart, T.C.; Gorry, M.C.; Hart, P.S.; Woodard, A.S.; Shihabi, Z.; Sandhu, J.; Shirts, B.; Xu, L.; Zhu, H.; Barmada, M.M.; et al. Mutations of the UMOD gene are responsible for medullary cystic kidney disease 2 and familial juvenile hyperuricaemic nephropathy. J. Med. Genet. 2002, 39, 882–892. [Google Scholar] [CrossRef]
- Cheng, Y.; Vapurcuyan, A.; Shahidullah, M.; Aleksunes, L.M.; Pelis, R.M. Expression of organic anion transporter 2 in the human kidney and its potential role in the tubular secretion of guanine-containing antiviral drugs. Drug Metab. Dispos. 2012, 40, 617–624. [Google Scholar] [CrossRef]
- Ekaratanawong, S.; Anzai, N.; Jutabha, P.; Miyazaki, H.; Noshiro, R.; Takeda, M.; Kanai, Y.; Sophasan, S.; Endou, H. Human organic anion transporter 4 is a renal apical organic anion/dicarboxylate exchanger in the proximal tubules. J. Pharmacol. Sci. 2004, 94, 297–304. [Google Scholar] [CrossRef]
- Burckhardt, G. Drug transport by Organic Anion Transporters (OATs). Pharmacol. Ther. 2012, 136, 106–130. [Google Scholar] [CrossRef]
- Cha, S.H.; Sekine, T.; Kusuhara, H.; Yu, E.; Kim, J.Y.; Kim, D.K.; Sugiyama, Y.; Kanai, Y.; Endou, H. Molecular cloning and characterization of multispecific organic anion transporter 4 expressed in the placenta. J. Biol. Chem. 2000, 275, 4507–4512. [Google Scholar] [CrossRef]
- Tamai, I. Pharmacological and pathophysiological roles of carnitine/organic cation transporters (OCTNs: SLC22A4, SLC22A5 and Slc22a21). Biopharm. Drug Dispos. 2013, 34, 29–44. [Google Scholar] [CrossRef]
- Ciarimboli, G.; Deuster, D.; Knief, A.; Sperling, M.; Holtkamp, M.; Edemir, B.; Pavenstädt, H.; Lanvers-Kaminsky, C.; am Zehnhoff-Dinnesen, A.; Schinkel, A. H.; et al. Organic cation transporter 2 mediates cisplatin-induced oto- and nephrotoxicity and is a target for protective interventions. Am. J. Pathol. 2010, 176, 1169–1180. [Google Scholar] [CrossRef]
- Jonker, J.W.; Wagenaar, E.; Van Eijl, S.; Schinkel, A.H. Deficiency in the organic cation transporters 1 and 2 (Oct1/Oct2 [Slc22a1/Slc22a2]) in mice abolishes renal secretion of organic cations. Mol. Cell Biol. 2003, 23, 7902–7908. [Google Scholar] [CrossRef]
- Wang, J.; Pan, Y.; Hong, Y.; Zhang, Q.Y.; Wang, X.N.; Kong, L.D. Quercetin Protects against Cadmium-Induced Renal Uric Acid Transport System Alteration and Lipid Metabolism Disorder in Rats. Evid. Based Complement. Alternat. Med. 2012, 2012, 548430. [Google Scholar]
- Fei, Y.; Zhou, Y.; Zhu, D.P.; Liu, P. Protective effect of Gardeniae fructus on diabetic nephropathy in rats. Chin. J. Gerontol. 2012, 1, 115–118. [Google Scholar]
- Wang, Y.; Yan, J.; Xi, L.; Qian, Z.; Wang, Z.; Yang, L. Protective effect of crocetin on hemorrhagic shock-induced acute renal failure in rats. Shock 2012, 38, 63–67. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Wu, W.K.; Li, F.; Song, W.; He, Y.; Yang, M. Study on liver protection, cholagogic effect and hepatotoxicity of Gardeniae Fructus. China J. Chin. Mater. Med. 2011, 19, 2610–2614. [Google Scholar]
- Imaoka, T.; Kusuhara, H.; Adachi-Akahane, S.; Hasegawa, M.; Morita, N.; Endou, H.; Sugiyama, Y. The renal-specific transporter mediates facilitative transport of organic anions at the brush border membrane of mouse renal tubules. J. Am. Soc. Nephrol. 2004, 15, 2012–2022. [Google Scholar] [CrossRef]
- Hosoyamada, M.; Ichida, K.; Enomoto, A.; Hosoya, T.; Endou, H. Function and localization of urate transporter 1 in mouse kidney. J. Am. Soc. Nephrol. 2004, 15, 261–268. [Google Scholar] [CrossRef]
- Hu, Q.H.; Zhang, X.; Wang, X.; Jiao, R.Q.; Kong, L.D. Quercetin regulates organic ion transporter and uromodulin expression and improves renal function in hyperuricemic mice. Eur. J. Nutr. 2012, 51, 593–606. [Google Scholar] [CrossRef]
- Carroll, J.J.; Coburn, H.; Douglass, R.; Babson, A.L. A simplified alkaline phosphotungstate assay for uric acid in serum. Clin. Chem. 1971, 17, 158–160. [Google Scholar]
- Perez-Ruiz, F.; Calabozo, M.; Erauskin, G.G.; Ruibal, A.; Herrero-Beites, A.M. Renal underexcretion of uric acid is present in patients with apparent high urinary uric acid output. Arthritis Rheum. 2002, 47, 610–613. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds Fructus Gardenia Extract are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hu, Q.-H.; Zhu, J.-X.; Ji, J.; Wei, L.-L.; Miao, M.-X.; Ji, H. Fructus Gardenia Extract Ameliorates Oxonate-Induced Hyperuricemia with Renal Dysfunction in Mice by Regulating Organic Ion Transporters and mOIT3. Molecules 2013, 18, 8976-8993. https://doi.org/10.3390/molecules18088976
Hu Q-H, Zhu J-X, Ji J, Wei L-L, Miao M-X, Ji H. Fructus Gardenia Extract Ameliorates Oxonate-Induced Hyperuricemia with Renal Dysfunction in Mice by Regulating Organic Ion Transporters and mOIT3. Molecules. 2013; 18(8):8976-8993. https://doi.org/10.3390/molecules18088976
Chicago/Turabian StyleHu, Qing-Hua, Ji-Xiao Zhu, Jing Ji, Lin-Lin Wei, Ming-Xing Miao, and Hui Ji. 2013. "Fructus Gardenia Extract Ameliorates Oxonate-Induced Hyperuricemia with Renal Dysfunction in Mice by Regulating Organic Ion Transporters and mOIT3" Molecules 18, no. 8: 8976-8993. https://doi.org/10.3390/molecules18088976
APA StyleHu, Q.-H., Zhu, J.-X., Ji, J., Wei, L.-L., Miao, M.-X., & Ji, H. (2013). Fructus Gardenia Extract Ameliorates Oxonate-Induced Hyperuricemia with Renal Dysfunction in Mice by Regulating Organic Ion Transporters and mOIT3. Molecules, 18(8), 8976-8993. https://doi.org/10.3390/molecules18088976