Real Time Anti-Toxoplasma gondii Activity of an Active Fraction of Eurycoma longifolia Root Studied by in Situ Scanning and Transmission Electron Microscopy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
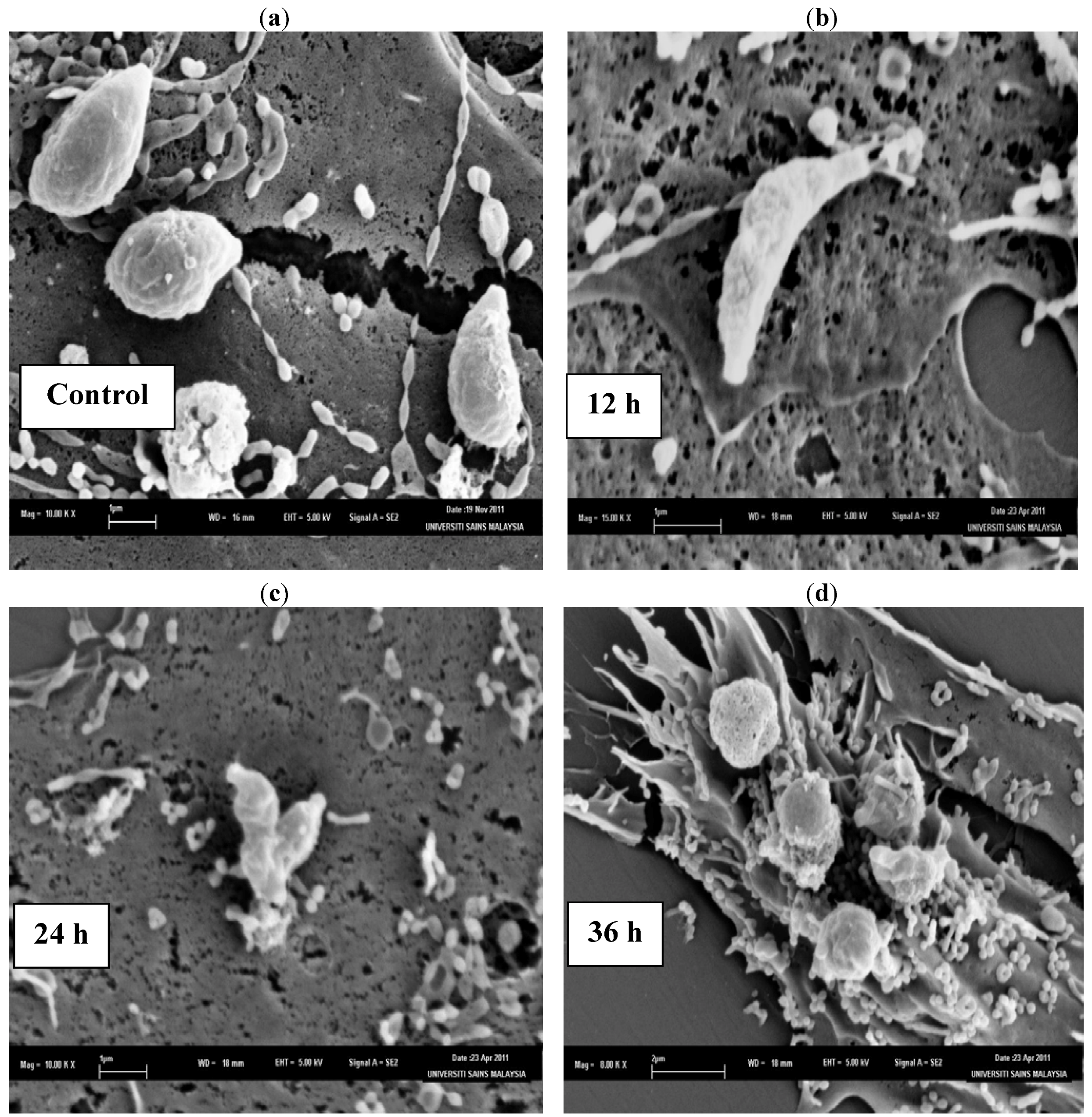
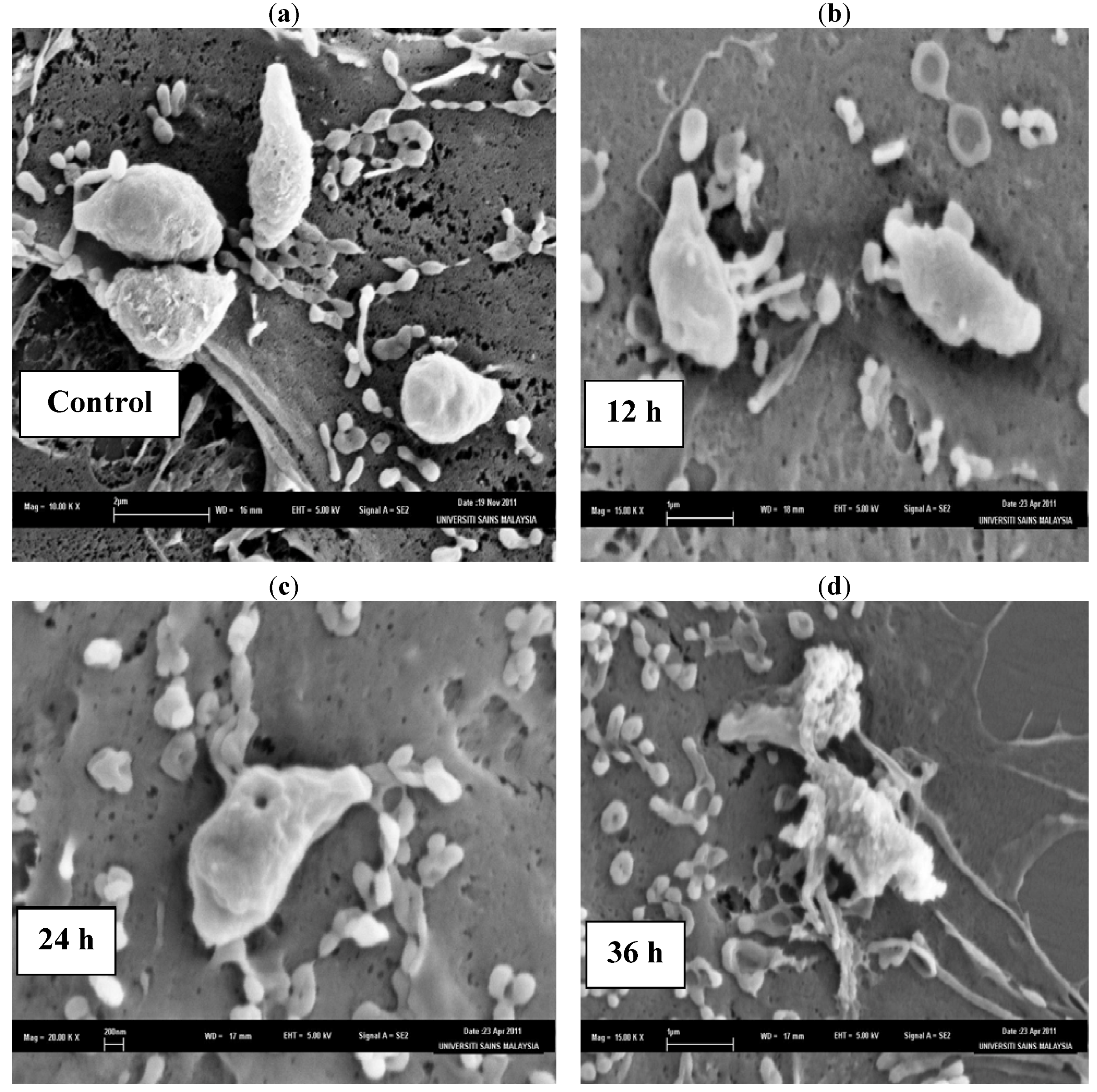
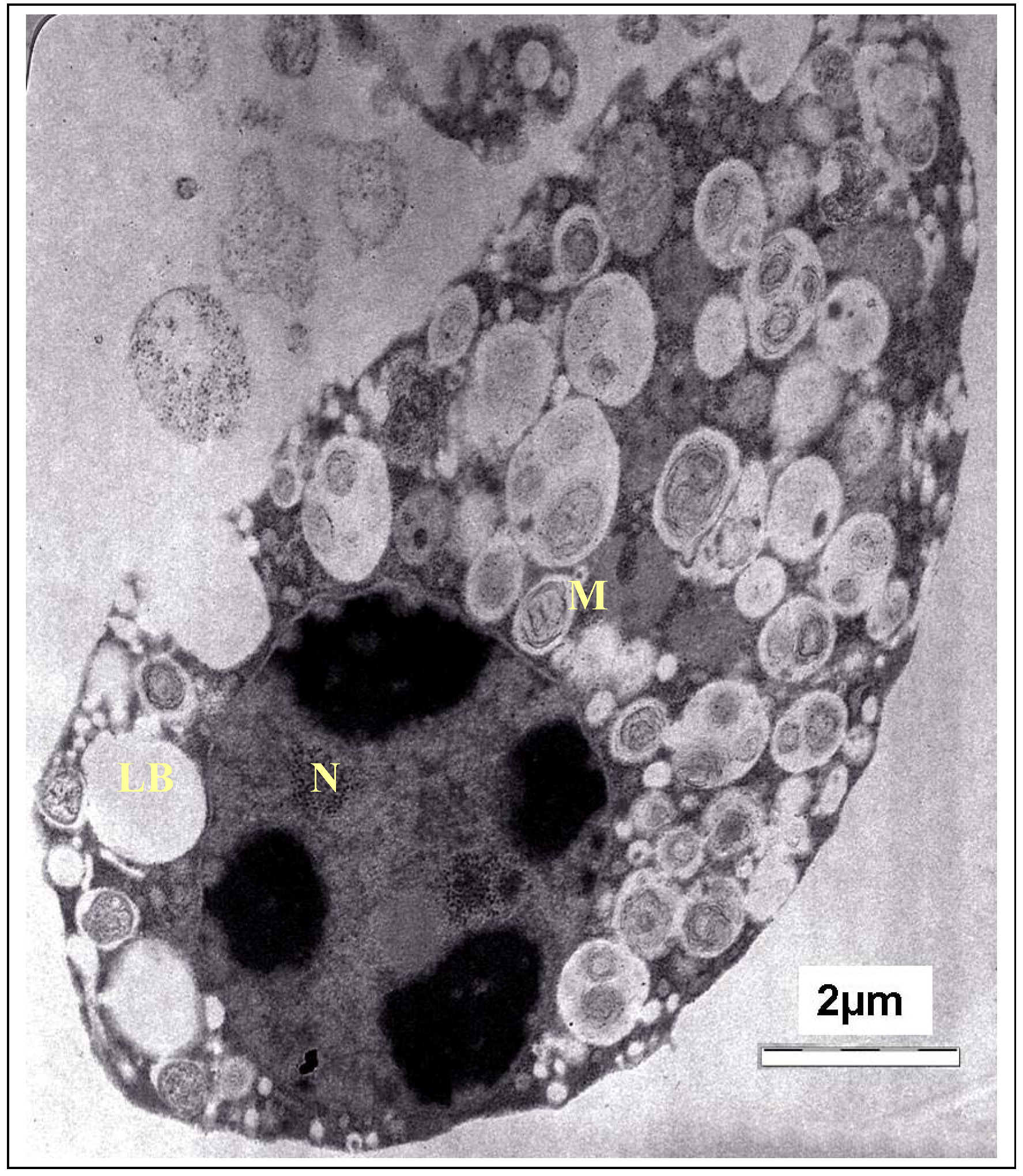
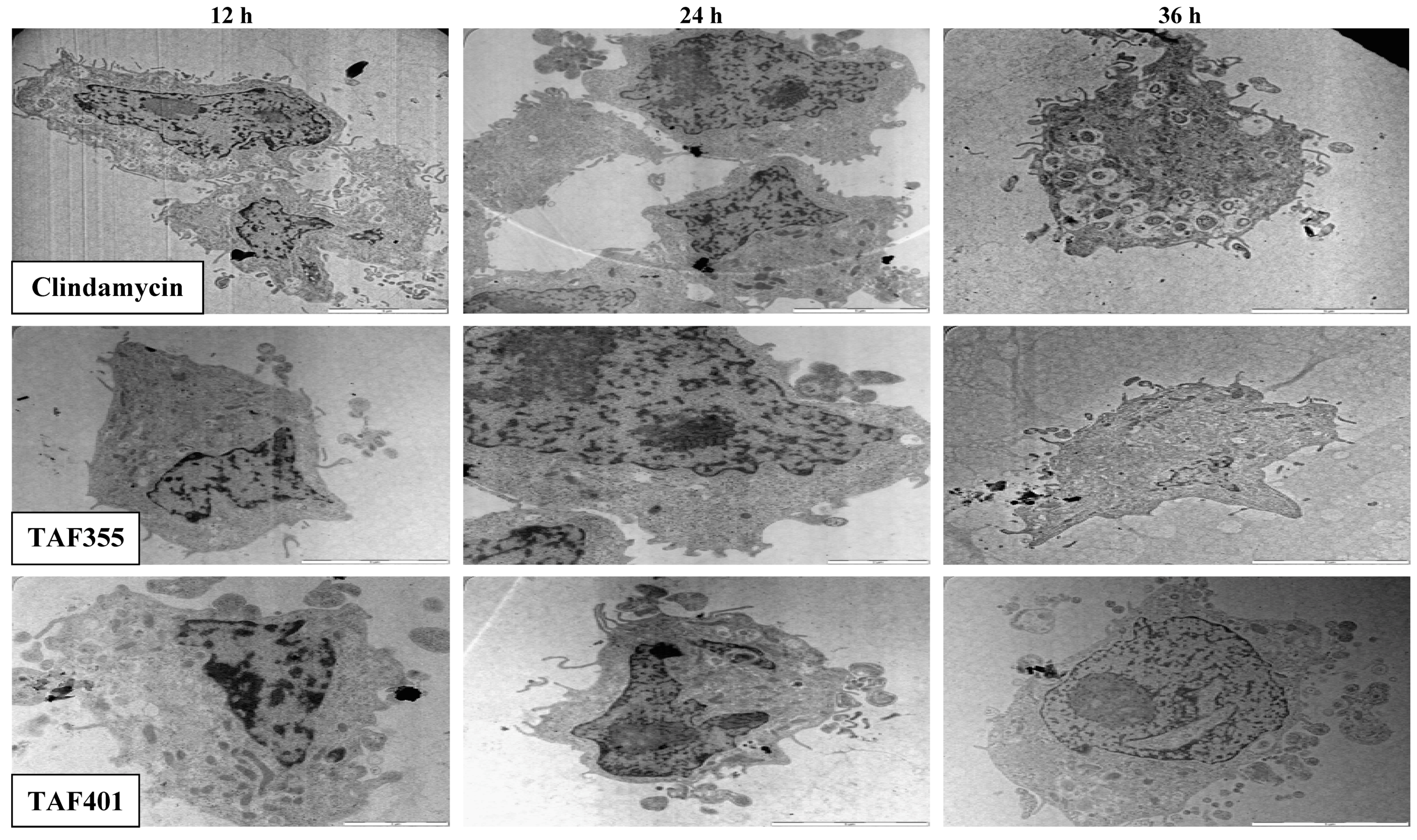
2. Results and Discussion

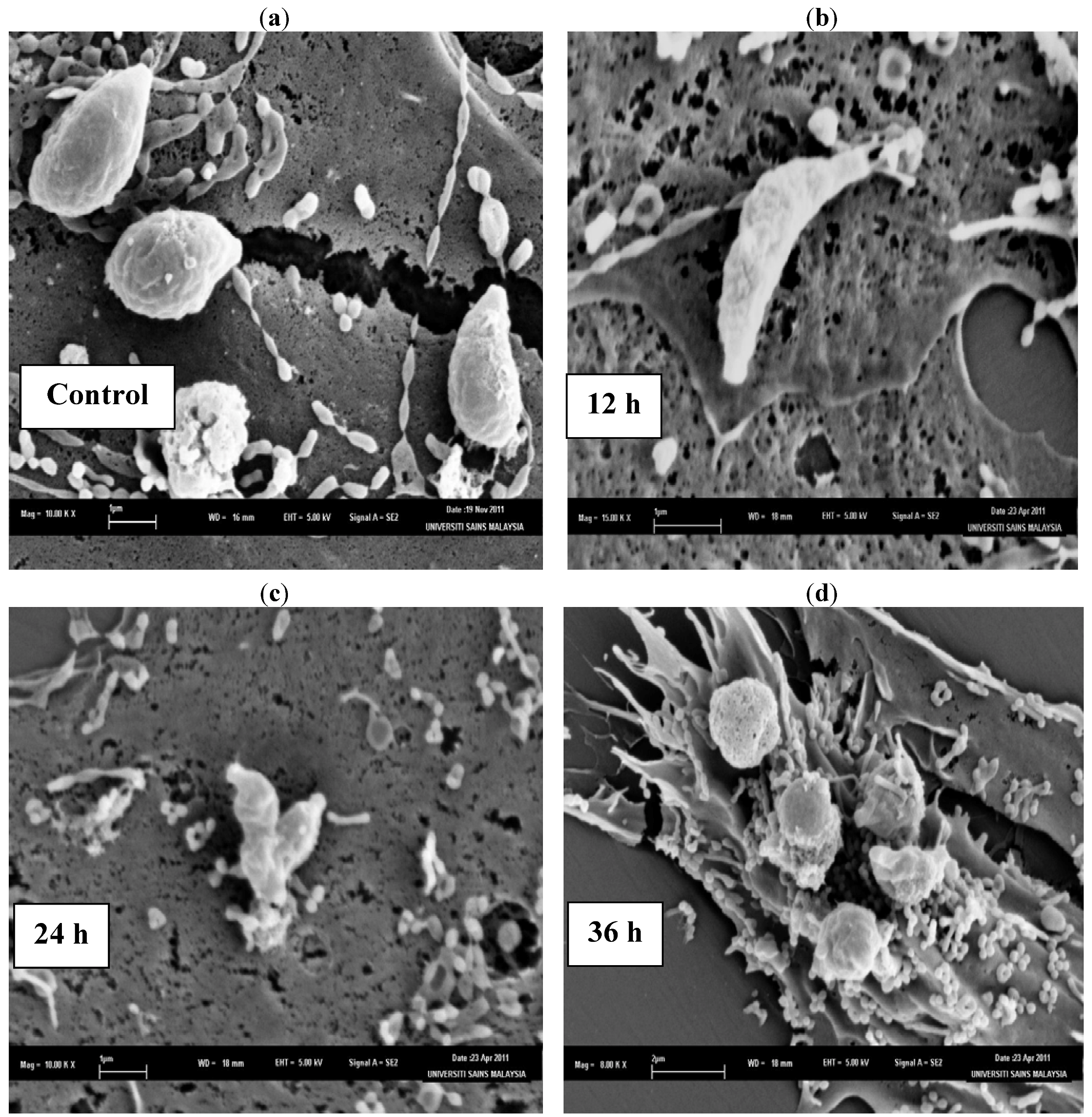
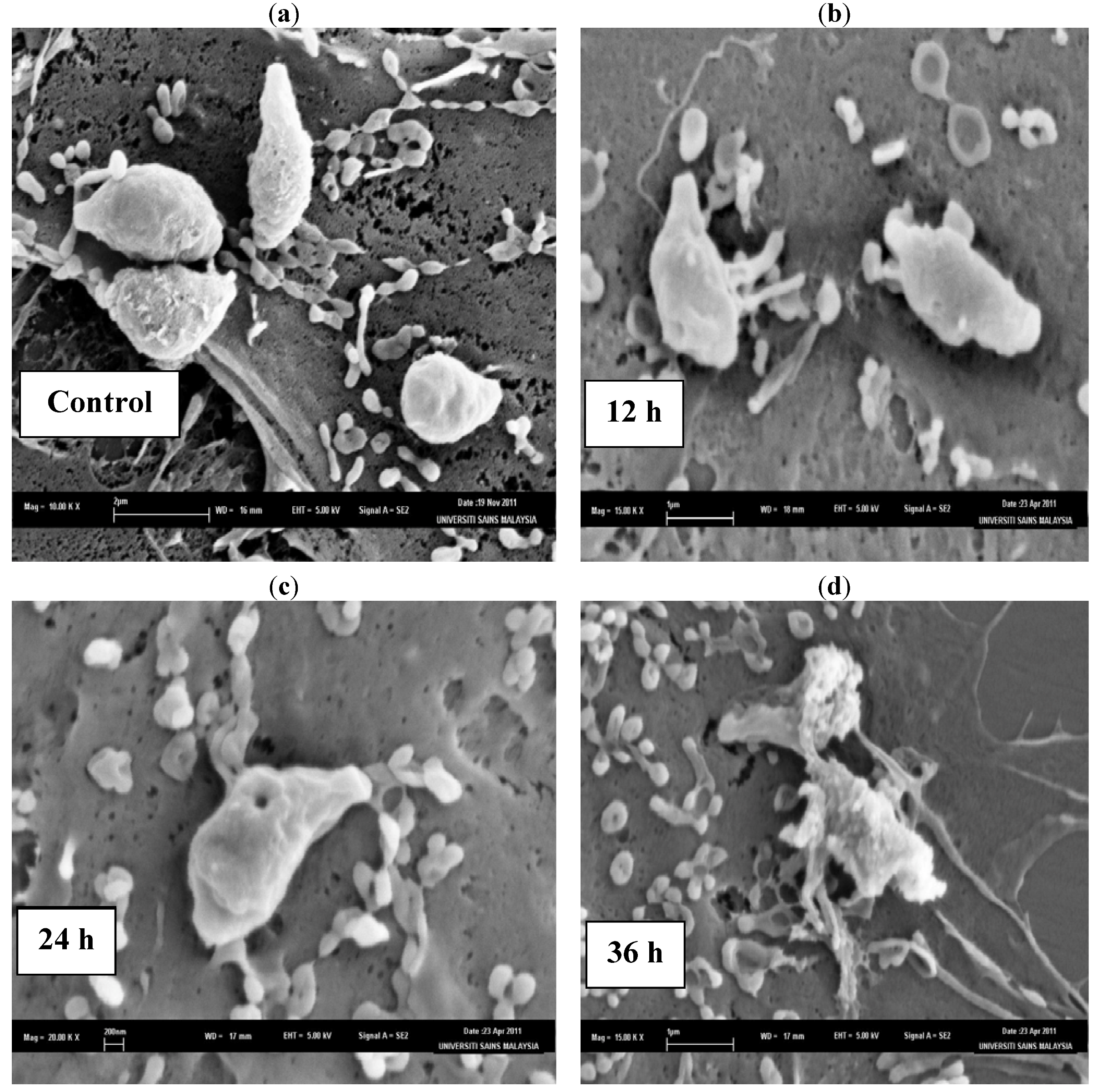
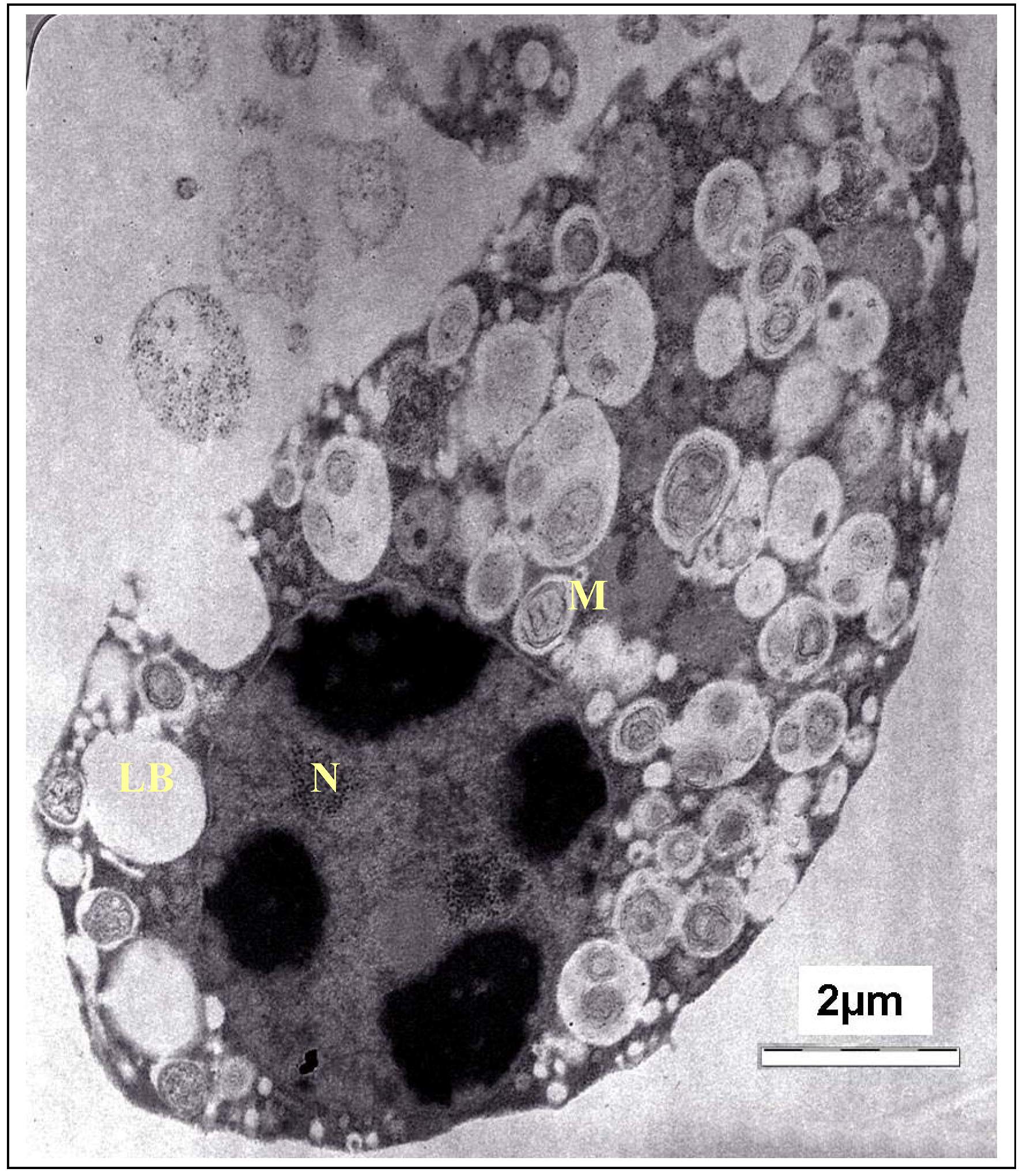
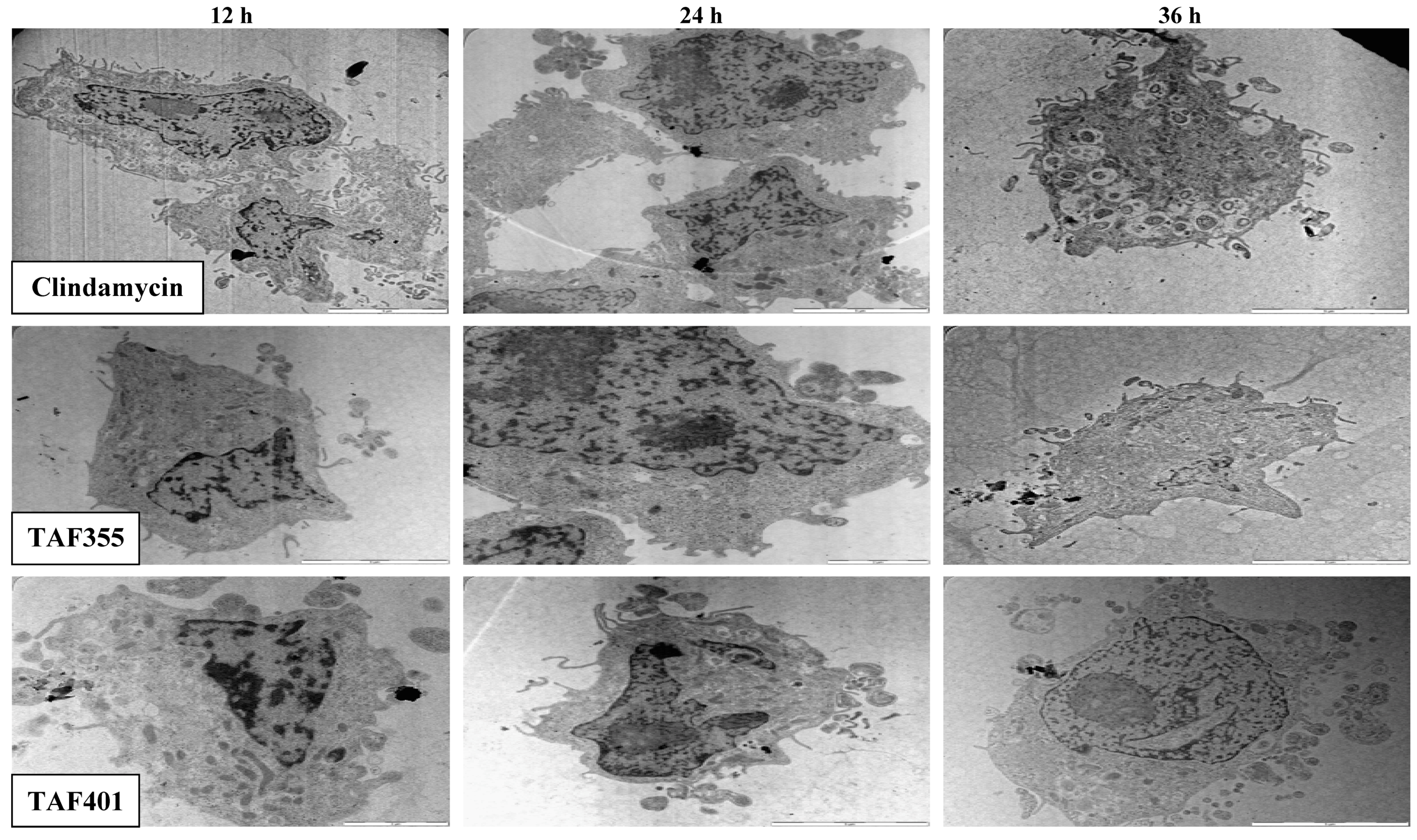
3. Experimental
3.1. Plant Collection
3.2. Preparation of Fraction
3.3. Toxoplasma Gondii Strain
3.4. Host Cells
3.5. Ultrastructural Observations
3.5.1. Scanning Electron Microscopy (SEM) Observation of the Tachyzoites in Vero Cells
3.5.2. Transmission Electron Microscopy (TEM) Observation of the Tachyzoites in Vero Cells
4. Conclusions
Acknowledgements
References
- Gimlette, J.D.; Thomson, H.W. A Dictionary of Malayan Medicine; Oxford University Press: Kuala Lumpur, Malaysia, 1977; p. 183. [Google Scholar]
- Perry, L.M. Medicinal Plants for East and Southeast Asia. Attributed Properties and Uses; M.I.T. Press: Cambridge, MA, USA, 1980; p. 389. [Google Scholar]
- Itokawa, H.; Kishi, E.; Morita, H.; Takeya, K. Cytotoxic quassinoids and tirucallane-type triterpenes from the woods of Eurycoma longifolia. Chem. Pharm. Bull. 1992, 40, 1053–1055. [Google Scholar] [CrossRef]
- Chan, K.L.; O’Neill, M.J.; Phillipson, J.D.; Warhurst, D.C. Plants as sources of antimalarial drugs. Part 3. Eurycoma longifolia Jack. Planta Med. 1986, 52, 105–107. [Google Scholar] [CrossRef]
- Chan, K.L.; Lee, S.P.; Sam, T.W.; Han, B.H. A quassinoid glycoside from the roots of Eurycoma longifolia. Phytochemistry 1989, 28, 2857–2859. [Google Scholar] [CrossRef]
- Chan, K.L.; Lee, S.P.; Sam, T.W.; Tan, S.C.; Noguchi, H.; Sankawa, U. 13β,18-dihydroeurycomanol, a quassinoid from Eurycoma longifolia. Phytochemistry 1991, 30, 3138–3141. [Google Scholar]
- Ang, H.H.; Chan, K.L.; Mak, J.W. Effect of 7-day daily replacement of culture medium containing Eurycoma longifolia Jack constituents on the Malaysian Plasmodium falciparum isolates. J. Ethnopharmacol. 1995, 49, 171–175. [Google Scholar] [CrossRef]
- Ang, H.H.; Chan, K.L.; Wah Mak, J. In vitro antimalarial activity of quassinoids from Eurycoma longifolia against Malaysian chloroquine-resistant Plasmodium falciparum isolates. Planta Med. 1995, 61, 177–178. [Google Scholar] [CrossRef]
- Kardono, L.B.S.; Angerhofer, C.K.; Tsauri, S.; Padmawinata, K.; Pezzuto, J.M.; Kinghorn, A.D. Cytotoxic and antimalarial constituents of the roots of Eurycoma longifolia. J. Nat. Prod. 1991, 54, 1360–1367. [Google Scholar] [CrossRef]
- Morita, H.; Kishi, E.; Takeya, K.; Itokawa, H.; Iitaka, Y. New quassinoids from the roots of Eurycoma longifolia. Chem. Lett. 1990, 5, 749–752. [Google Scholar]
- Itokawa, H.; Qin, X.R.; Morita, H.; Takeya, K.; Iitaka, Y. Novel quassinoids from Eurycoma longifolia. Chem. Pharm. Bull. 1993, 41, 403–405. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.L.; Choo, C.Y.; Noor, R.A.; Zakiah, I. Antiplasmodial studies of Eurycoma longifolia Jack using the lactate dehydrogenase assay of Plasmodium falciparum. J. Ethnopharmacol. 2004, 92, 223–227. [Google Scholar] [CrossRef]
- Luder, C.G.; Bohne, W.; Soldati, D. Toxoplasmosis: A persisting challenge. Trends. Parasitol. 2001, 17, 460–463. [Google Scholar] [CrossRef]
- Toxoplasmosis Infection. Harrison’s Principles of Internal Medicine, Chapter 214; AccessMedicine, McGraw-Hill Companies. Available online: http://www.accessmedicine.com/content.aspx?aID=9125477&searchStr=toxoplasmosis#9125477 (accessed on 31 July 2012).
- Sasidharan, S.; Kavitha, N.; Noordin, R.; Chan, K.L. Anti-Toxoplasma Gondii activity of root extract/fraction of Eurycoma Longifolia Jack in vitro. In Functional Food Components in Health and Disease, Science and Pratice; Martirosyan, D.M., Nicolson, G.L., Eds.; University of San Diego: San Diego, CA, USA, 2011; pp. 155–156. [Google Scholar]
- Sasidharan, S.; Yoga Latha, L.; Angeline, T. Imaging In vitro anti-biofilm activity to visualize the ultrastructural changes. In Microscopy: Science, Technology, Applications and Education; Méndez-Vilas, A., Díaz, J., Badajoz, D.J., Eds.; FORMATEX: Badajoz, Spain, 2010; pp. 622–626. [Google Scholar]
- Kovacs, J.A.; Hiemenz, J.W.; Macher, A.M.; Stover, D.; Murray, H.W.; Shelhamer, J.; Lane, H.C.; Urmacher, C.; Honig, C.; Longo, D.L.; et al. Pneumocystis carinli pneumonia: A comparison between patients with the acquired immunodeficiency syndrome and patients with other immunodeficiencies. Ann. Intern. Med. 1984, 100, 663–671. [Google Scholar]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar]
- Strobl, J.S.; Cassell, M.; Mitchell, S.M.; Reilly, C.M.; Lindsay, D.S. Scriptaid and suberoylanilide hydroxamic acid are histone deacetylase inhibitors with potent anti-Toxoplasma gondii activity. in vitro. J. Parasitol. 2007, 93, 694–700. [Google Scholar]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants and ageing. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef]
- Wang, W.; Fang, H.; Groom, L.; Cheng, A.; Zhang, W.; Liu, J.; Wang, X.; Li, K.; Han, P.; Zheng, M.; et al. Superoxide flashes in single mitochondria. Cell 2008, 134, 279–290. [Google Scholar] [CrossRef]
- Kavitha, N.; Noordin, R.; Chan, K.L.; Sasidharan, S. Cytotoxicity activity of root extract/fractions of Eurycoma longifolia Jack root against vero and Hs27cells. J. Med. Plants Res. 2010, 4, 2383–2387. [Google Scholar]
- Dantas-Leite, L.; Urbina, J.A.; de Souza, W.; Vommaro, R.C. Selective anti-Toxoplasma gondii activities of azasterols. Int. J. Antimicrob. Agents 2004, 23, 620–626. [Google Scholar] [CrossRef]
- Hemphill, A.; Croft, S.L. Electron microscopy in parasitology. In Analytical Parasitology; Rogan, M., Ed.; Springer Verlag: Heidelberg, Germany, 1997; pp. 227–268. [Google Scholar]
- Sample Availability: Samples of the extract of E. longifolia are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kavitha, N.; Noordin, R.; Kit-Lam, C.; Sasidharan, S. Real Time Anti-Toxoplasma gondii Activity of an Active Fraction of Eurycoma longifolia Root Studied by in Situ Scanning and Transmission Electron Microscopy. Molecules 2012, 17, 9207-9219. https://doi.org/10.3390/molecules17089207
Kavitha N, Noordin R, Kit-Lam C, Sasidharan S. Real Time Anti-Toxoplasma gondii Activity of an Active Fraction of Eurycoma longifolia Root Studied by in Situ Scanning and Transmission Electron Microscopy. Molecules. 2012; 17(8):9207-9219. https://doi.org/10.3390/molecules17089207
Chicago/Turabian StyleKavitha, Nowroji, Rahmah Noordin, Chan Kit-Lam, and Sreenivasan Sasidharan. 2012. "Real Time Anti-Toxoplasma gondii Activity of an Active Fraction of Eurycoma longifolia Root Studied by in Situ Scanning and Transmission Electron Microscopy" Molecules 17, no. 8: 9207-9219. https://doi.org/10.3390/molecules17089207
APA StyleKavitha, N., Noordin, R., Kit-Lam, C., & Sasidharan, S. (2012). Real Time Anti-Toxoplasma gondii Activity of an Active Fraction of Eurycoma longifolia Root Studied by in Situ Scanning and Transmission Electron Microscopy. Molecules, 17(8), 9207-9219. https://doi.org/10.3390/molecules17089207