Mutagenicity of Flavonoids Assayed by Bacterial Reverse Mutation (Ames) Test

Abstract
:1. Introduction
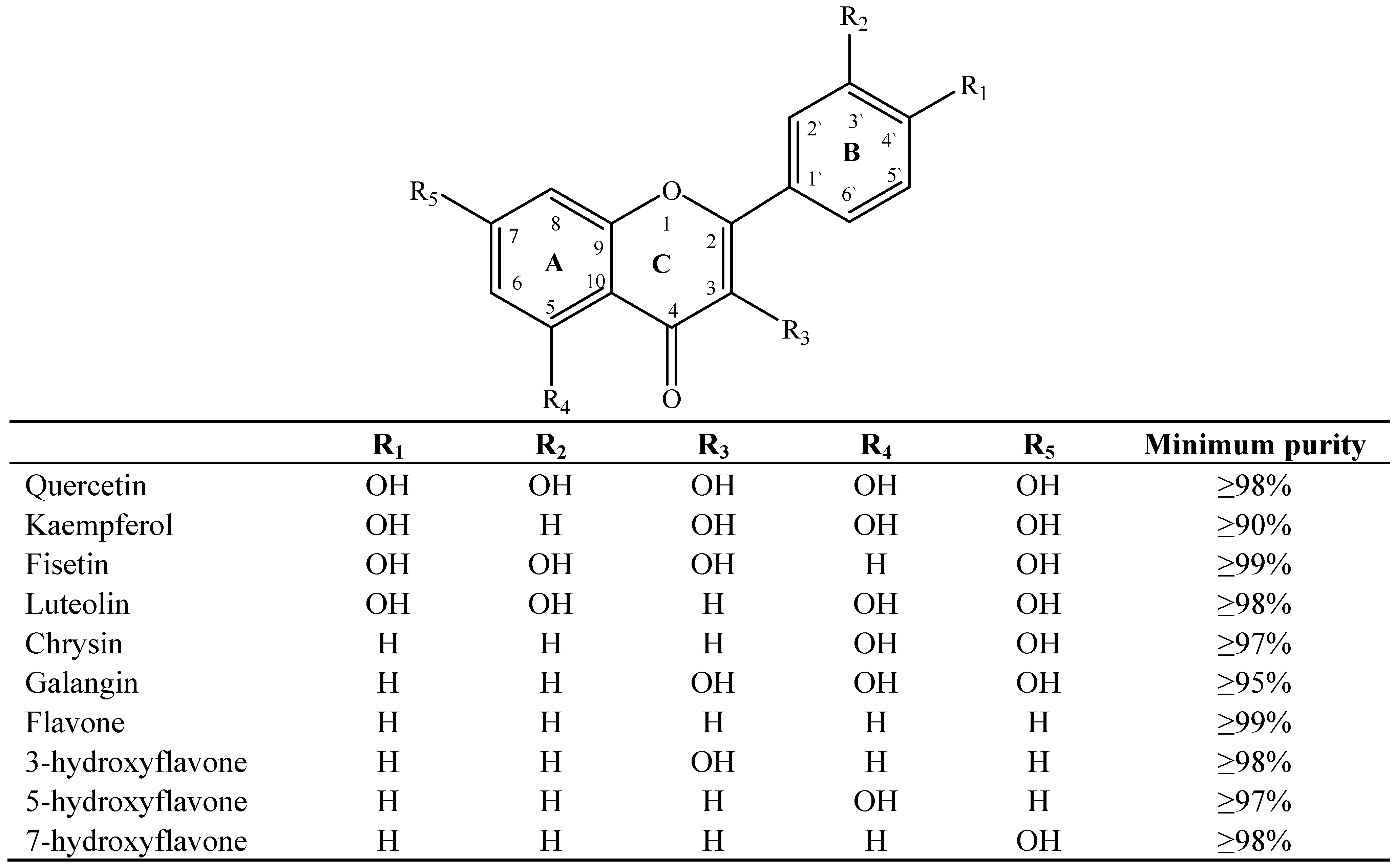
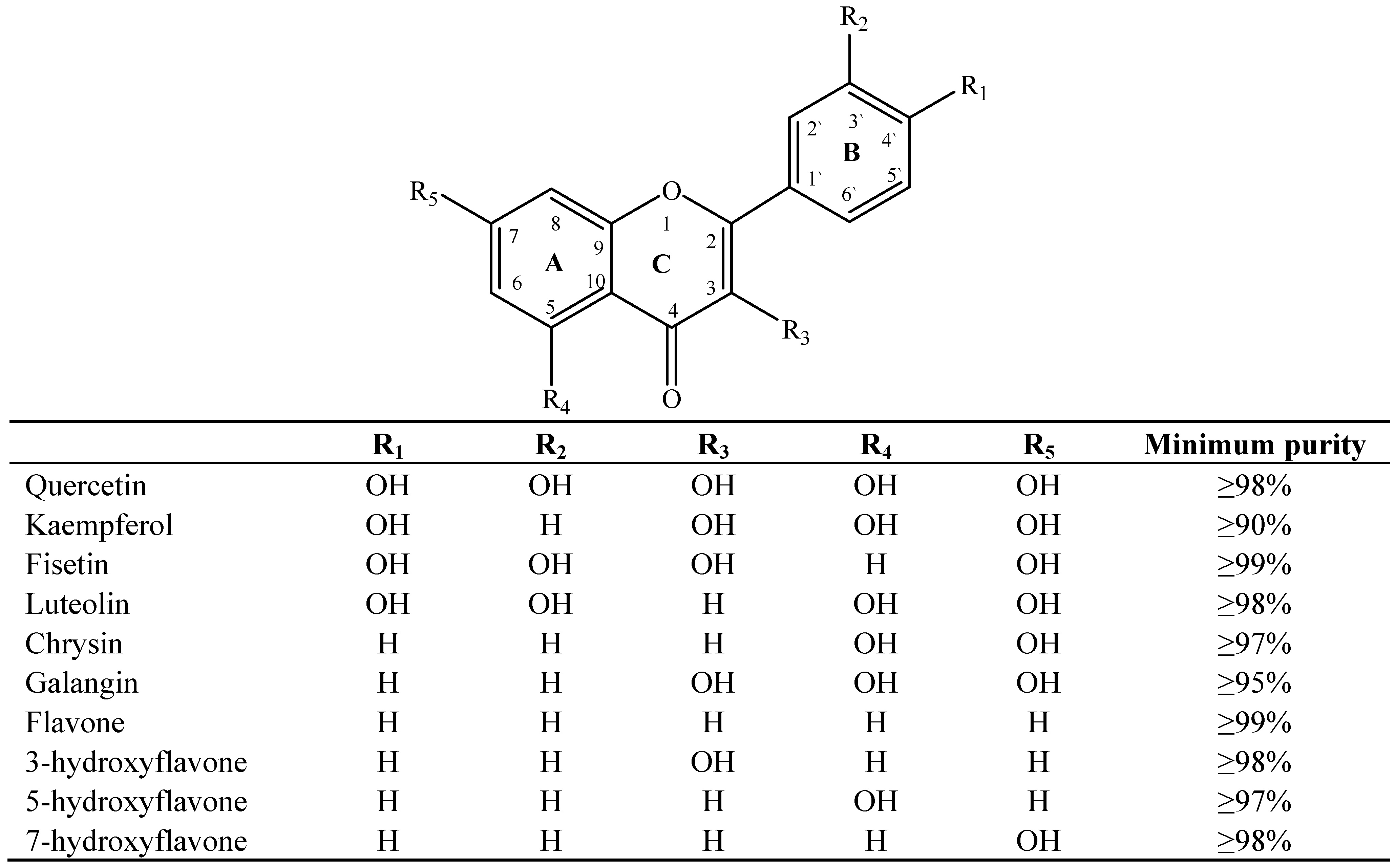
2. Results and Discussion
2.1. Results
2.2. Discussion

{kind=link}
| Treatments | Number of revertants∕plate in S. typhimurium strains (M ± SD) and (MI) | ||||||
|---|---|---|---|---|---|---|---|
| TA 98 | TA 100 | TA 102 | |||||
Quercetin  | nmol/plate | −S9 | +S9 | −S9 | +S9 | −S9 | +S9 |
| 0.0 a | 53 ± 8 | 39 ± 1 | 115 ± 3.5 | 206 ± 5 | 273 ± 15 | 248 ± 3 | |
| 12.1 | 81 ± 0.6 * (1.5) | 136 ± 5 ** (3.5) | 134 ± 8.5 (1.2) | 320 ± 2 * (1.5) | 279 ± 8 (1.0) | 305 ± 1 (1.2) | |
| 24.5 | 115 ± 6 ** (2.2) | 196 ± 4 ** (5.0) | 178 ± 6.1 (1.5) | 389 ± 4 ** (1.9) | 302 ± 4 (1.1) | 351 ± 13 (1.4) | |
| 49.1 | 172 ± 1 ** (3.3) | 393 ± 7 ** (10.0) | 202 ± 3.2 ** (1.7) | 492 ± 5 ** (2.4) | 331 ± 6 (1.2) | 446 ± 11 ** (1.8) | |
| 98.4 | 135 ± 11 ** (2.6) | 658 ± 10 ** (16.9) | 227 ± 2.5 ** (2.0) | 651 ± 4 ** (3.2) | 324 ± 3 (1.2) | 471 ± 1 ** (1.9) | |
| 147.8 | 197 ± 4 ** (3.8) | 794 ± 5 ** (20.4) | 212 ± 5.0 ** (1.8) | 642 ± 5 ** (3.1) | 310 ± 6 (1.1) | 524 ± 6 ** (2.1) | |
| Ctrol+ | 1347 ± 51 b | 1567 ± 47 e | 1582 ± 102 c | 1456 ± 67 e | 1656 ± 89 d | 1932 ± 93 f | |
Kaempferol 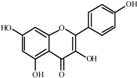 | 0.0 a | 58 ± 2 | 39 ± 1 | 106 ± 4 | 206 ± 53 | 235 ± 4 | 248 ± 3 |
| 14.3 | 54 ± 5 (0.9) | 141 ± 7** (3.6) | 131 ± 1 (1.2) | 286 ± 15 (1.4) | 287 ± 4 (1.2) | 336 ± 8 (1.3) | |
| 29.0 | 56 ± 4 (1.0) | 172 ± 3** (4.4) | 146 ± 2 (1.4) | 313 ± 5 (1.5) | 258 ± 6 (1.1) | 367 ± 7 (1.5) | |
| 58.0 | 53 ± 1 (0.9) | 199 ± 8** (5.1) | 143 ± 4 (1.3) | 369 ± 4 ** (1.8) | 301 ± 9 (1.3) | 430 ± 5 ** (1.7) | |
| 116.4 | 58 ± 2 (1.0) | 205 ± 3** (5.3) | 130 ± 6 (1.2) | 384 ± 12 ** (1.9) | 305 ± 9 (1.3) | 418 ± 8 (1.4) | |
| 174.7 | 62 ± 3 (1.1) | 177 ± 4** (4.5) | 138 ± 3 (1.3) | 400 ± 18 ** (1.9) | 242 ± 9 (1.0) | 473 ± 13 ** (1.9) | |
| Ctrol+ | 1324 ± 61 b | 1567 ± 47 e | 1457 ± 53 c | 1456 ± 67 e | 1473 ± 119 d | 1932 ± 93 f | |
Fisetin 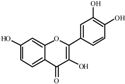 | 0.0 a | 50 ± 4 | 32 ± 1 | 146 ± 1 | 190 ± 1 | 264 ± 3 | 291 ± 8 |
| 14.3 | 50 ± 1 (1.0) | 41 ± 1 (1.3) | 157 ± 7 (1.1) | 209 ± 7 (1.1) | 341 ± 11 (1.3) | 333 ± 8 (1.1) | |
| 29.0 | 52 ± 3 (1.0) | 44 ± 2 (1.4) | 142 ± 1 (1.0) | 233 ± 6 (1.2) | 334 ± 2 (1.3) | 346 ± 10 (1.2) | |
| 58.0 | 57 ± 1 (1.1) | 40 ± 1 (1.2) | 175 ± 6 (1.2) | 232 ± 2 (1.2) | 372 ± 4 (1.4) | 348 ± 11 (1.2) | |
| 116.4 | 58 ± 2 (1.1) | 40 ± 2 (1.3) | 133 ± 3 (0.9) | 224 ± 8 (1.2) | 472 ± 16 ** (1.8) | 364 ± 15 (1.2) | |
| 174.7 | 60 ± 1 (1.2) | 36 ± 1 (1.1) | 131 ± 3 (0.9) | 232 ± 4 (1.2) | 339 ± 15 (1.3) | 387 ± 7 (1.3) | |
| Ctrol+ | 1425 ± 81 b | 1634 ± 88 e | 1325 ± 91 c | 1721 ± 72 e | 1689 ± 72 d | 1789 ± 90 f | |
Luteolin  | 0.0 a | 49 ± 3 | 39 ± 1 | 141 ± 2 | 206 ± 5 | 265 ± 2 | 248 ± 3 |
| 14.3 | 37 ± 2 (0.8) | 33 ± 2 (0.8) | 117 ± 3 (0.8) | 226 ± 8 (1.1) | 288 ± 6 (1.1) | 264 ± 2 (1.1) | |
| 29.0 | 45 ± 5 (0.9) | 38 ± 3 (1.0) | 129 ± 3 (0.9) | 249 ± 10 (1.2) | 282 ± 1 (1.1) | 310 ± 7 (1.2) | |
| 58.0 | 40 ± 3 (0.8) | 35 ± 2 (0.9) | 107 ± 3 (0.8) | 253 ± 11 (1.2) | 299 ± 16 (1.1) | 380 ± 4 (1.5) | |
| 116.4 | 45 ± 4 (0.9) | 31 ± 3 (0.8) | 150 ± 8 (1.1) | 188 ± 5 (0.9) | 340 ± 6 (1.3) | 387 ± 6 * (1.6) | |
| 174.7 | 48 ± 1 (1.0) | 27 ± 1 (0.7) | 148 ± 5 (1.0) | 198 ± 13 (1.0) | 288 ± 5 (1.1) | 404 ± 3 * (1.6) | |
| Ctrol+ | 1376 ± 67 b | 1567 ± 47 e | 1276 ± 72 c | 1456 ± 67 e | 1767 ± 41 d | 1932 ± 93 f | |
Chrysin 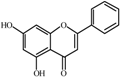 | 0.0 a | 49 ± 3 | 39 ± 1 | 156 ± 3 | 206 ± 5 | 235 ± 2 | 248 ± 3 |
| 16.1 | 34 ± 5 (0.7) | 34 ± 3 (0.9) | 154 ± 2 (1.0) | 222 ± 10 (1.1) | 253 ± 4 (1.1) | 305 ± 3 (1.2) | |
| 32.6 | 40 ± 2 (0.8) | 38 ± 2 (1.0) | 141 ± 3 (0.9) | 268 ± 7 (1.3) | 298 ± 8 (1.3) | 259 ± 2 (1.0) | |
| 65.3 | 37 ± 2 (0.7) | 33 ± 1 (0.8) | 144 ± 2 (0.9) | 274 ± 10 (1.3) | 265 ± 5 (1.1) | 233 ± 10 (0.9) | |
| 131.0 | 44 ± 1 (0.9) | 35 ± 3 (0.9) | 151 ± 4 (1.0) | 278 ± 15 (1.3) | 282 ± 2 (1.2) | 232 ± 6 (0.9) | |
| 196.7 | 49 ± 1 (1.0) | 32 ± 2 (0.8) | 124 ± 5 (0.8) | 254 ± 3 (1.2) | 230 ± 7 (1.0) | 231 ± 14 (0.9) | |
| Ctrol+ | 1376 ± 67 b | 1567 ± 47 e | 1579 ± 95 c | 1456 ± 67 e | 1377 ± 67 d | 1932 ± 93 f | |
Galangin 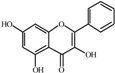 | 0.0 a | 22 ± 3 | 26 ± 1 | 208 ± 13 | 165 ± 1 | 144 ± 1 | 172 ± 5 |
| 2.6 | 22 ± 2 (1.0) | 34 ± 5 (1.3) | 238 ± 2 (1.1) | 209 ± 4 (1.3) | 155 ± 4 (1.1) | 144 ± 9 (0.8) | |
| 5.1 | 23 ± 2 (1.0) | 43 ± 2 * (1.6) | 235 ± 3 (1.1) | 207 ± 4 (1.3) | 150 ± 4 (1.0) | 162 ± 3 (0.9) | |
| 10.2 | 23 ± 2 (1.0) | 55 ± 3 ** (2.1) | 233 ± 9 (1.1) | 196 ± 5 (1.2) | 148 ± 7 (1.0) | 158 ± 6 (0.9) | |
| 20.5 | 25 ± 4 (1.1) | 57 ± 3 ** (2.1) | 209 ± 9 (1.0) | 177 ± 7 (1.1) | 143 ± 1 (1.0) | 151 ± 3 (0.9) | |
| 30.7 | 26 ± 2 (1.2) | 58 ± 3 ** (2.2) | 199 ± 7 (1.0) | 161 ± 9 (1.0) | 127 ± 2 (0.9) | 112 ± 7 (0.6) | |
| Ctrol+ | 1329 ± 66 b | 1422 ± 81 e | 1498 ± 45 c | 1667 ± 39 e | 1733 ± 77 d | 1980 ± 91 f | |
Flavone 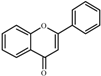 | 0.0 a | 35 ± 2 | 32 ± 1 | 173 ± 5 | 190 ± 1 | 313 ± 6 | 291 ± 8 |
| 18.4 | 36 ± 2 (1.0) | 38 ± 2 (1.2) | 182 ± 2 (1.0) | 221 ± 1 (1.2) | 282 ± 3 (0.9) | 350 ± 4 (1.2) | |
| 37.3 | 34 ± 3 (1.0) | 37 ± 2 (1.2) | 174 ± 2 (1.0) | 196 ± 4 (1.0) | 251 ± 4 (0.8) | 369 ± 7 (1.3) | |
| 74.7 | 38 ± 1 (1.1) | 36 ± 1 (1.1) | 179 ± 1 (1.0) | 208 ± 2 (1.1) | 212 ± 7 (0.7) | 414 ± 3 (1.4) | |
| 149.8 | 35 ± 3 (1.0) | 31 ± 2 (1.0) | 144 ± 5 (0.8) | 205 ± 4 (1.1) | 136 ± 2 (0.4) | 470 ± 8 * (1.6) | |
| 225.0 | 46 ± 2 (1.3) | 23 ± 2 (0.7) | 106 ± 4 (0.6) | 207 ± 5 (1.1) | 109 ± 3 (0.3) | 525 ± 6 ** (1.8) | |
3-
Hydroxyflavone 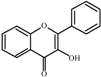 | 0.0 a | 50 ± 2 | 32 ± 1 | 194 ± 8 | 190 ± 1 | 235 ± 13 | 291 ± 8 |
| 17.2 | 50 ± 4 (1.0) | 39 ± 3 (1.2) | 196 ± 6 (1.0) | 235 ± 3 (1.2) | 216 ± 11 (0.9) | 308 ± 5 (1.1) | |
| 34.8 | 43 ± 6 (0.8) | 36 ± 3 (1.1) | 207 ± 5 (1.1) | 196 ± 2 (1.0) | 207 ± 19 (0.8) | 308 ± 7 (1.1) | |
| 69.7 | 38 ± 7 (0.8) | 34 ± 2 (1.1) | 192 ± 5 (1.0) | 192 ± 8 (1.0) | 214 ± 8 (0.9) | 334 ± 4 (1.1) | |
| 139.8 | 48 ± 10 (0.9) | 34 ± 2 (1.1) | 217 ± 2 (1.1) | 204 ± 6 (1.1) | 212 ± 10 (0.9) | 348 ± 3 (1.2) | |
| 209.9 | 53 ± 1 (1.0) | 27 ± 3 (0.8) | 196 ± 5 (1.0) | 212 ± 12 (1.1) | 222 ± 16 (0.9) | 348 ± 7 (1.2) | |
| Ctrol+ | 1337 ± 92 b | 1634 ± 88 e | 1523 ± 85 c | 1721 ± 72 e | 1384 ± 82 d | 1789 ± 90 f | |
5-
Hydroxyflavone 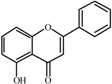 | 0.0 a | 32 ± 1 | 32 ± 1 | 177 ± 2 | 190 ± 1 | 306 ± 8 | 291 ± 8 |
| 17.2 | 32 ± 2 (1.0) | 26 ± 1 (0.8) | 166 ± 3 (0.9) | 202 ± 10 (1.1) | 290 ± 7 (0.9) | 353 ± 4 (1.2) | |
| 34.8 | 36 ± 2 (1.1) | 28 ± 2 (0.9) | 156 ± (0.9) | 208 ± 17 (1.1) | 303 ± 5 (1.0) | 361 ± 8 (1.2) | |
| 69.7 | 30 ± 2 (0.9) | 29 ± 3 (0.9) | 185 ± 3 (1.0) | 203 ± 6 (1.1) | 296 ± 4 (1.0) | 391 ± 8 (1.3) | |
| 139.8 | 44 ± 2 (1.4) | 31 ± 1 (1.0) | 166 ± 3 (0.9) | 206 ± 1 (1.1) | 285 ± 4 (0.9) | 405 ± 10 (1.4) | |
| 209.9 | 34 ± 2 (1.1) | 31 ± 4 (1.0) | 142 ± 4 (0.8) | 193 ± 8 (1.0) | 265 ± 6 (0.9) | 438 ± 6 (1.5) | |
| Ctrol+ | 1237 ± 61 b | 1634 ± 88 e | 1211 ± 72 c | 1721 ± 72 e | 1877 ± 52 d | 1789 ± 90 f | |
7-
Hydroxyflavone 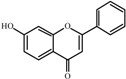 | 0.0 a | 50 ± 2 | 32 ± 1 | 136 ± 8 | 190 ± 1 | 229 ± 7 | 291 ± 8 |
| 17.2 | 57 ± 4 (1.1) | 30 ± 4 (0.9) | 124 ± 3 (0.9) | 211 ± 5 (1.1) | 246 ± 25 (1.1) | 344 ± 6 (1.2) | |
| 34.8 | 56 ± 3 (1.1) | 35 ± 3 (1.1) | 121 ± 5 (0.9) | 190 ± 9 (1.0) | 270 ± 22 (1.2) | 344 ± 5 (1.2) | |
| 69.7 | 55 ± 8 (1.1) | 36 ± 1 (1.1) | 145 ± 6 (1.1) | 205 ± 4 (1.1) | 249 ± 13 (1.1) | 345 ± 4 (1.2) | |
| 139.8 | 65 ± 2 (1.3) | 36 ± 1 (1.1) | 143 ± 4 (1.0) | 197 ± 8 (1.0) | 246 ± 16 (1.1) | 351 ± 13 (1.2) | |
| 209.9 | 54 ± 4 (1.1) | 39 ± 3 (1.2) | 132 ± 18 (1.0) | 177 ± 2 (0.9) | 244 ± 16 (1.1) | 369 ± 2 (1.3) | |
| Ctrol+ | 1337 ± 92 b | 1634 ± 88 e | 1398 ± 51 c | 1721 ± 72 e | 1277 ± 57 d | 1789 ± 90 f | |
| Treatments | TA 98 | TA 100 | TA 102 | |||
|---|---|---|---|---|---|---|
| −S9 | +S9 | −S9 | +S9 | −S9 | +S9 | |
| Quercetin | + | + | + | + | - | + |
| Kaempferol | - | + | - | +∕- | - | +∕- |
| Fisetin | - | - | - | - | +∕- | - |
| Luteolin | - | - | - | - | - | +∕- |
| Chrysin | - | - | - | - | - | - |
| Galangin | - | + | - | - | - | - |
| Flavone | - | - | - | - | - | +∕- |
| 3-Hydroxyflavone | - | - | - | - | - | - |
| 5-Hydroxyflavone | - | - | - | - | - | - |
| 7-Hydroxyflavone | - | - | - | - | - | - |
3. Experimental
3.1. Chemicals and Culture Media
3.2. Metabolic Activation System (S9 Mixture)
3.3. Salmonella Mutagenic Assay
4. Conclusions
Acknowledgements
Conflict of Interest
- Sample Availability:Samples of the compounds quercetin, kaempferol, luteolin, fisetin, chrysin, galangin, flavone, 3-hydroxyflavone, 5-hydroxyflavone and 7-hydroxyflavone are available from the authors.
References and notes
- Chebil, L.; Anthoni, J.; Humeau, C.; Gerardin, C.; Engasser, J.M.; Ghoul, M. Enzymatic acylation of flavonoids: Effect of the nature of the substrate, origin of lipase, and operating conditions on conversion yield and regioselectivity. J. Agric. Food Chem. 2007, 55, 9496–9502. [Google Scholar]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biotechnol. 2011, 91, 949–956. [Google Scholar] [CrossRef]
- Taylor, L.P.; Grotewold, E. Flavonoids as developmental regulators. Curr. Opin. Plant Biol. 2005, 8, 317–323. [Google Scholar] [CrossRef]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: Modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J.Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Spencer, J.P.E.; Vauzour, D.; Rendeiro, C. Flavonoids and cognition: The molecular mechanisms underlying their behavioural effects. Arch. Biochem. Biophys. 2009, 492, 1–9. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Benigni, R.; Bossa, C. Structure alerts for carcinogenicity, and the Salmonella assay system: A novel insight through the chemical relational databases technology. Mutat. Res. 2008, 659, 248–261. [Google Scholar] [CrossRef]
- Boersma, M.G.; Vervoort, J.; Szymusiak, H.; Lemanska, K.; Tyrakowska, B.; Cenas, N.; Segura-Aguilar, J.; Rietjens, I.M.C.M. Regioselectivity and reversibility of the glutathione conjugation of quercetin quinone methide. Chem. Res. Toxicol. 2000, 13, 185–191. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Duarte Silva, I.; Gaspar, J.; Gomes da Costa, G.; Rodrigues, A.S.; Laires, A.; Rueff, J. Chemical features of flavonols affecting their genotoxicity. Potential implications in their use as therapeutical agents. Chem. Biol. Interact. 2000, 124, 29–51. [Google Scholar] [CrossRef]
- Mazzatorta, P.; Tran, L.A.; Schilter, B.; Grigorov, M. Integration of structure-activity relationship and artificial intelligence systems to improve in silico prediction of Ames test mutagenicity. J. Chem. Inf. Model. 2007, 47, 34–38. [Google Scholar] [CrossRef]
- Snijman, P.W.; Swanevelder, S.; Elizabeth Joubert, E.; Green, I.R.; Gelderblom, W.C.A. The antimutagenic activity of the major flavonoids of rooibos (Aspalathus linearis): Some dose-response effects on mutagen activation-flavonoid interactions. Mutat. Res. 2007, 631, 111–123. [Google Scholar] [CrossRef]
- Macgregor, J.T. Mutagenic and carcinogenic effects of flavonoids. In Plant Flavonoids in Biology and Medicine: Biochemical Pharmacological and Structure-Activity Relationships; Cody, V., Middlenton, E., Harbone, J.B., Eds.; Alan R. Liss: New York, NY, USA, 1986; pp. 411–424. [Google Scholar]
- Skibola, C.F.; Smith, M.T. Potential health impacts of excessive flavonoid intake. Free Radic. Biol. Med. 2000, 29, 375–383. [Google Scholar] [CrossRef]
- Sahu, R.K.; Basu, R.; Sharma, A. Genetic toxicological testing of some plant flavonoids by the micronucleus test. Mutat. Res. 1981, 89, 69–74. [Google Scholar] [CrossRef]
- Das, A.; Wang, J.H.; Lien, E.J. Carcinogenicity, mutagenicity and cancer preventing activities of flavonoids: A structure-system activity relationship (SSAR) analysis. Prog. Drug Res. 1994, 42, 133–167. [Google Scholar]
- Rietjens, I.M.C.M.; Boersma, M.G.; van der Woude, H.; Jeurissen, S.M.F.; Schutte, M.E.; Alink, G.M. Flavonoids and alkenylbenzenes: Mechanisms of mutagenic action and carcinogenic risk. Mutat. Res. 2005, 574, 124–138. [Google Scholar] [CrossRef]
- Hou, D.X.; Kumamoto, T. Flavonoids as protein kinase inhibitors for cancer chemoprevention: Direct binding and molecular modeling. Antioxid. Redox Signal. 2010, 13, 691–719. [Google Scholar] [CrossRef]
- Brown, J.P.; Dietrich, P.S. Mutagenicity of plant flavonoids in the Salmonella/mammalian microsome test. Activation of flavonol glycosides by mixed glycosides from rat fecal bacteria and other sources. Mutat. Res. 1979, 66, 223–240. [Google Scholar] [CrossRef]
- Duarte Silva, I.; Rodrigues, A.S.; Gaspar, J.; Laires, A.; Rueff, J. Metabolism of galangin by rat cytochromes P450: Relevance to the genotoxicity of galangin. Mutat. Res. 1997, 393, 247–257. [Google Scholar] [CrossRef]
- van der Woude, H.; Alink, G.M.; van Rossum, B.E.J.; Walle, K.; van Steeg, H.; Walle, T.; Rietjens, I.M.C.M. Formation of transient covalent protein and DNA adducts by quercetin in cells with and without oxidative enzyme activity. Chem. Res. Toxicol. 2005, 18, 1907–1916. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Boots, A.W.; Li, H.; Schins, R.P.; Duffin, R.; Heemskerk, J.W.; Bast, A.; Haenen, G.R. The quercetin paradox. Toxicol. Appl. Pharmacol. 2007, 222, 89–96. [Google Scholar] [CrossRef]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Myers, L.N.; Adams, N.; Kier, T.K.; Rao, B.; Shaw, B.; Williams, L. Microcomputer software for data management and statistical analyses of the Ames∕Salmonella. In Statistical Methods in Toxicological Research; Krewski, D., Ed.; Gordon and Brech: New York, NY, USA, 1991; pp. 265–279. [Google Scholar]
- Bernstein, L.; Kaldor, J.; Mccann, J.; Pike, M.C. An empirical approach to the statistical analysis of mutagenesis data from the Salmonella test. Mutat. Res. 1982, 97, 267–281. [Google Scholar] [CrossRef]
- Santos, F.V.; Colus, I.M.S.; Silva, M.A.; Vilegas, W.; Varanda, E.A. Assessment of DNA damage induced by extracts and fractions of Strychnos pseudoquina, a Brazilian medicinal plant with antiulcerogenic activity. Food Chem. Toxicol. 2006, 44, 1585–1589. [Google Scholar] [CrossRef]
- McGeorge, L.S.; Louis, J.B.; Altrerholt, T.B.; Mcgarrity, G.J. Mutagenicity analysis of industrial effluents: Results and consideration for integration into water pollution control programs. In Short Term Bioassays in the Analysis of Complex Environmental Mixtures; Waters, M.D., Claxton, L., Lewtas, J., Nesnow, S., Sandhu, S.S., Strauss, G., Eds.; Plenum: New York, NY, USA, 1985; Volume 4, pp. 247–267. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Resende, F.A.; Vilegas, W.; Dos Santos, L.C.; Varanda, E.A. Mutagenicity of Flavonoids Assayed by Bacterial Reverse Mutation (Ames) Test. Molecules 2012, 17, 5255-5268. https://doi.org/10.3390/molecules17055255
Resende FA, Vilegas W, Dos Santos LC, Varanda EA. Mutagenicity of Flavonoids Assayed by Bacterial Reverse Mutation (Ames) Test. Molecules. 2012; 17(5):5255-5268. https://doi.org/10.3390/molecules17055255
Chicago/Turabian StyleResende, Flavia Aparecida, Wagner Vilegas, Lourdes Campaner Dos Santos, and Eliana Aparecida Varanda. 2012. "Mutagenicity of Flavonoids Assayed by Bacterial Reverse Mutation (Ames) Test" Molecules 17, no. 5: 5255-5268. https://doi.org/10.3390/molecules17055255
APA StyleResende, F. A., Vilegas, W., Dos Santos, L. C., & Varanda, E. A. (2012). Mutagenicity of Flavonoids Assayed by Bacterial Reverse Mutation (Ames) Test. Molecules, 17(5), 5255-5268. https://doi.org/10.3390/molecules17055255