Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
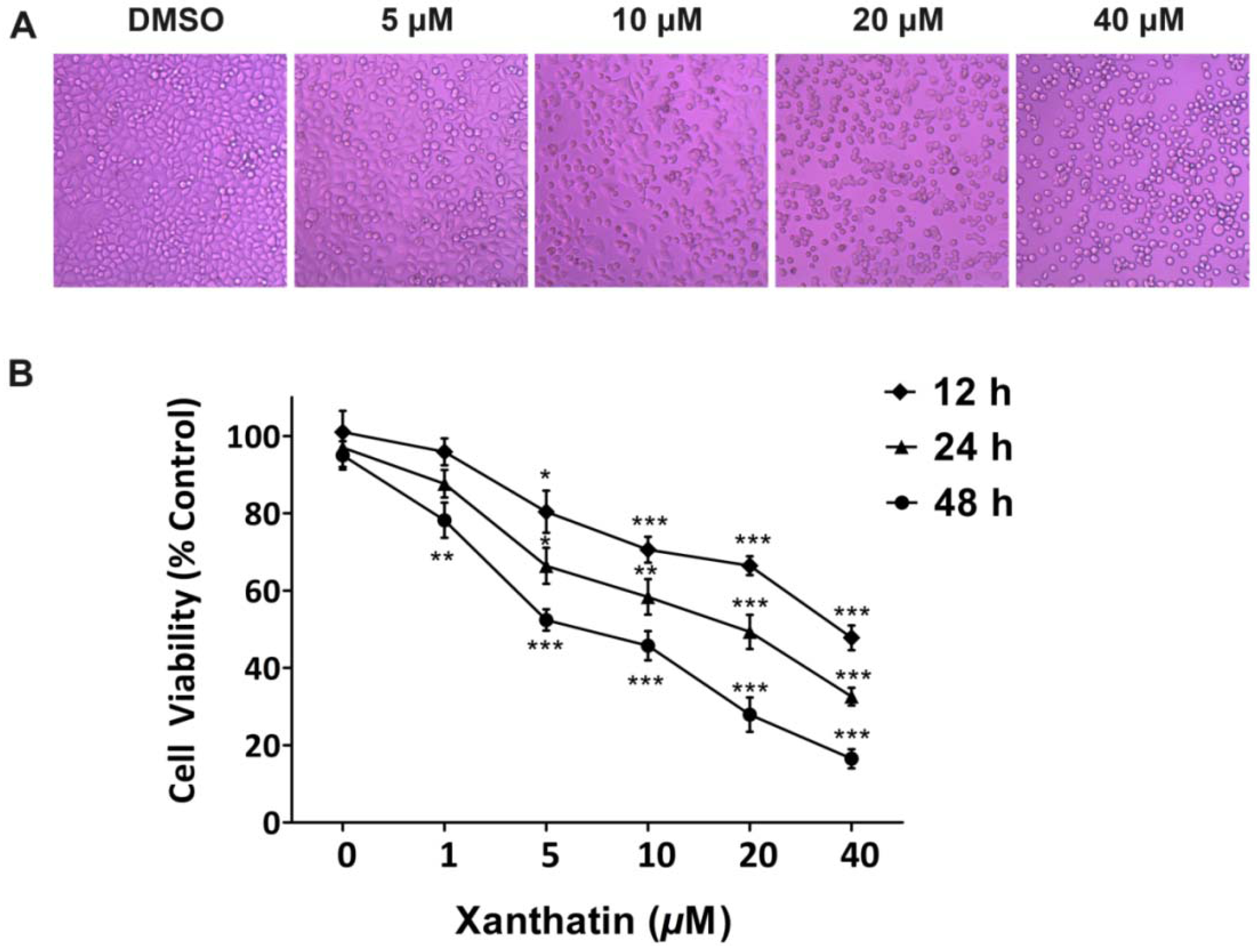
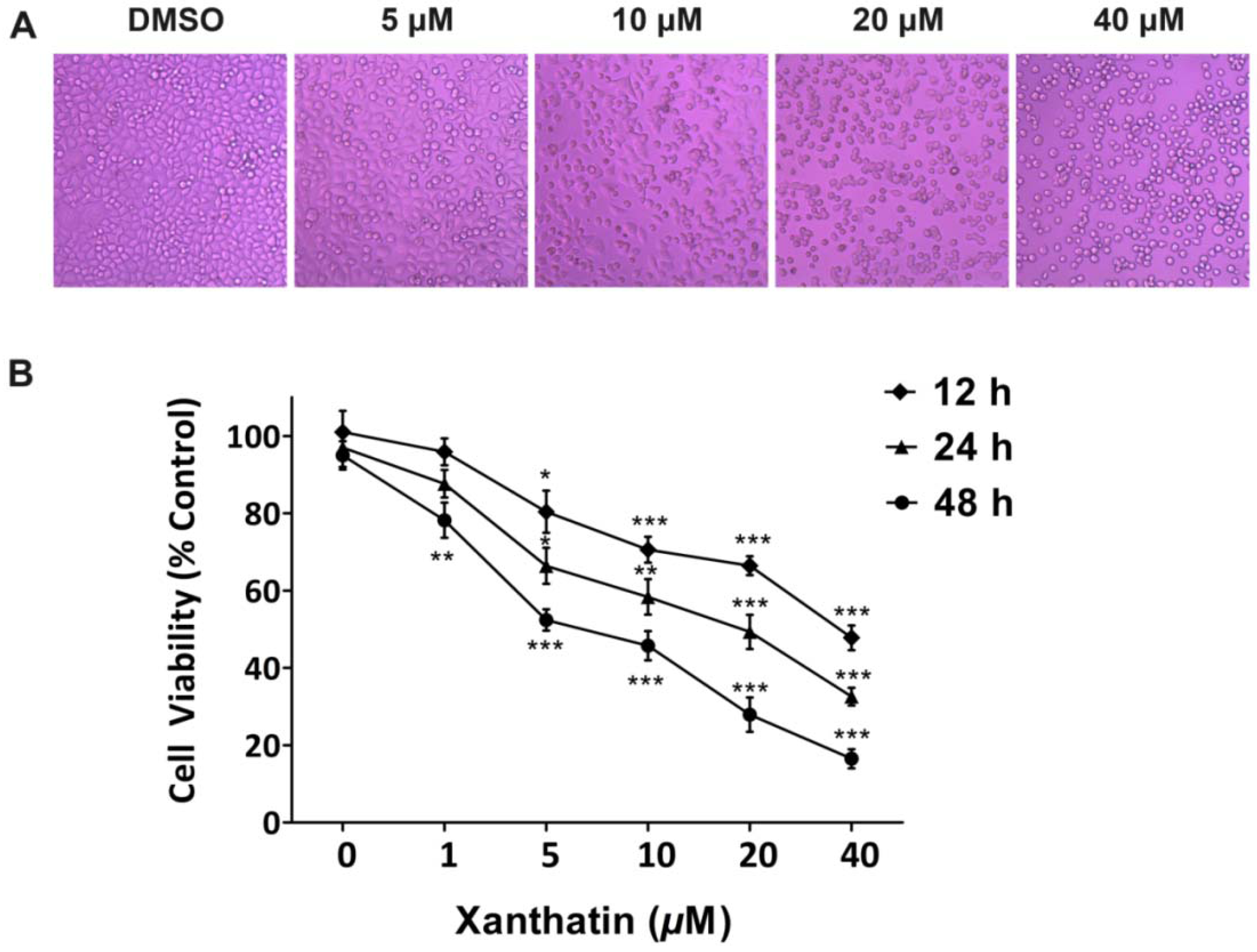
2.1.1. Xanthatin Inhibited A549 Cell Growth Dose-/Time-Dependently
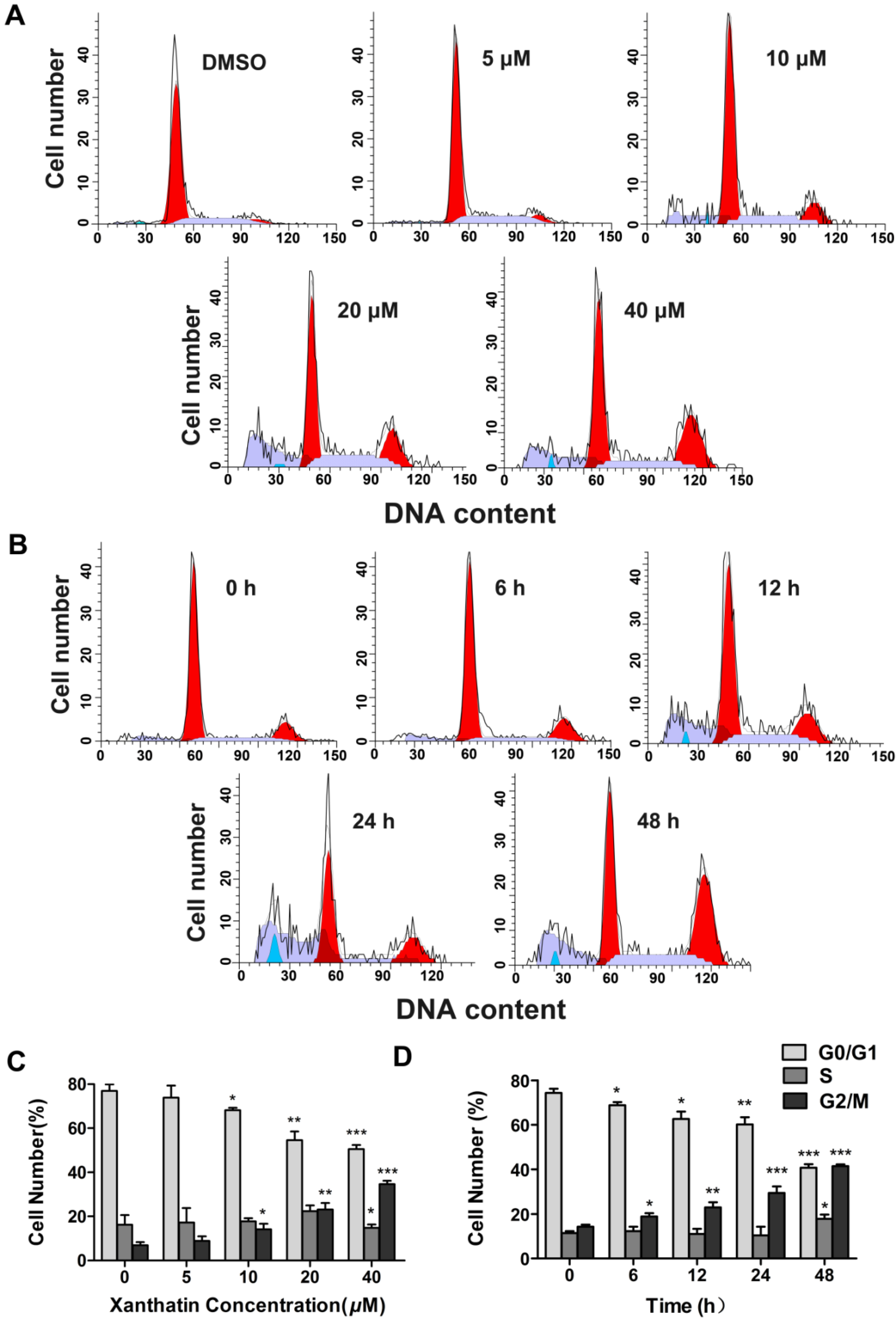
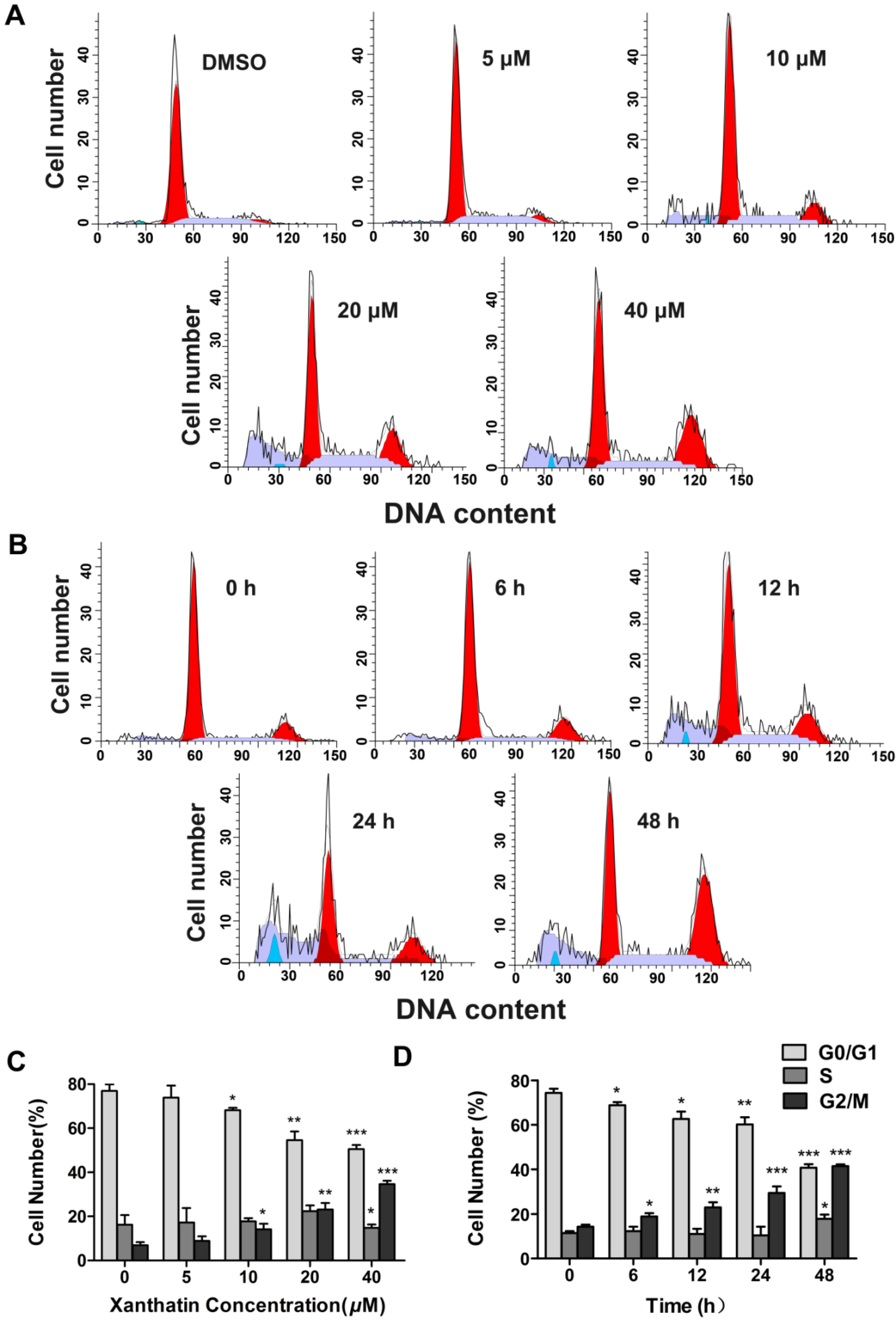
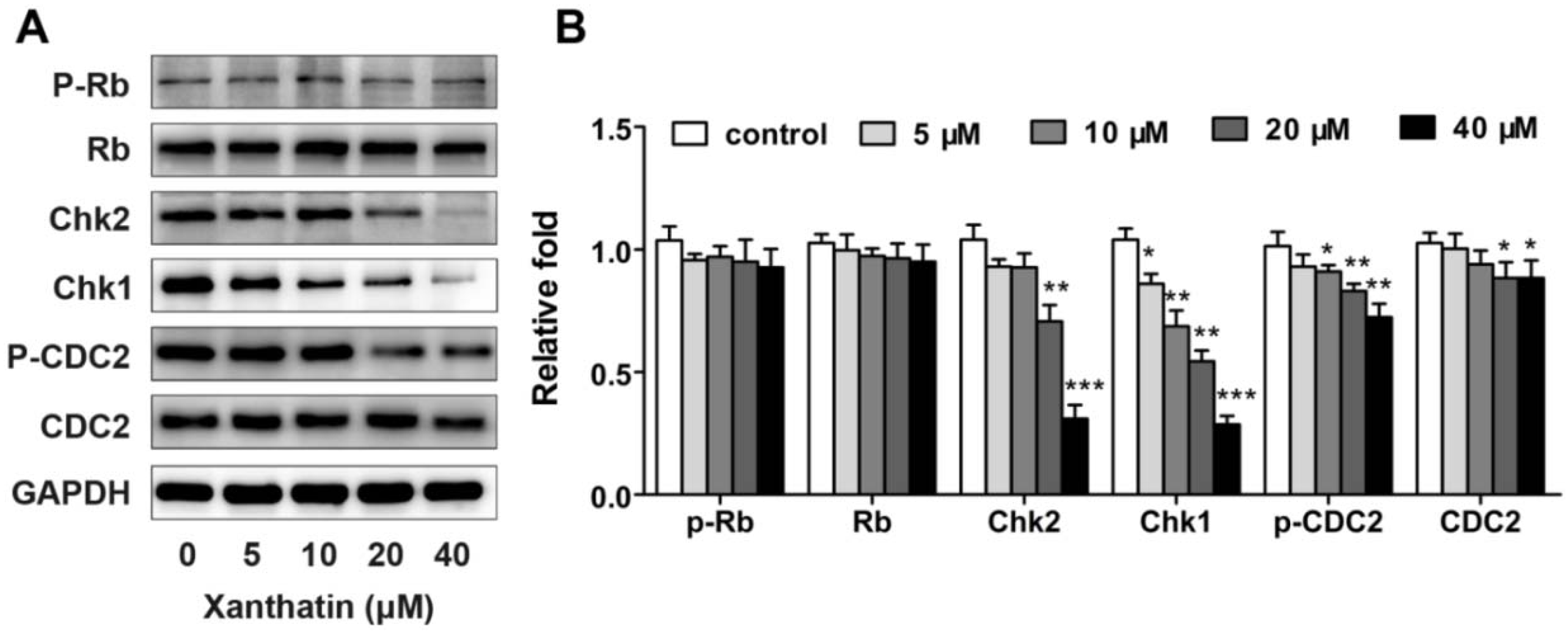
2.1.2. Xanthatin Induced Cell Cycle Arrest at G2 Phase in A549 Cells
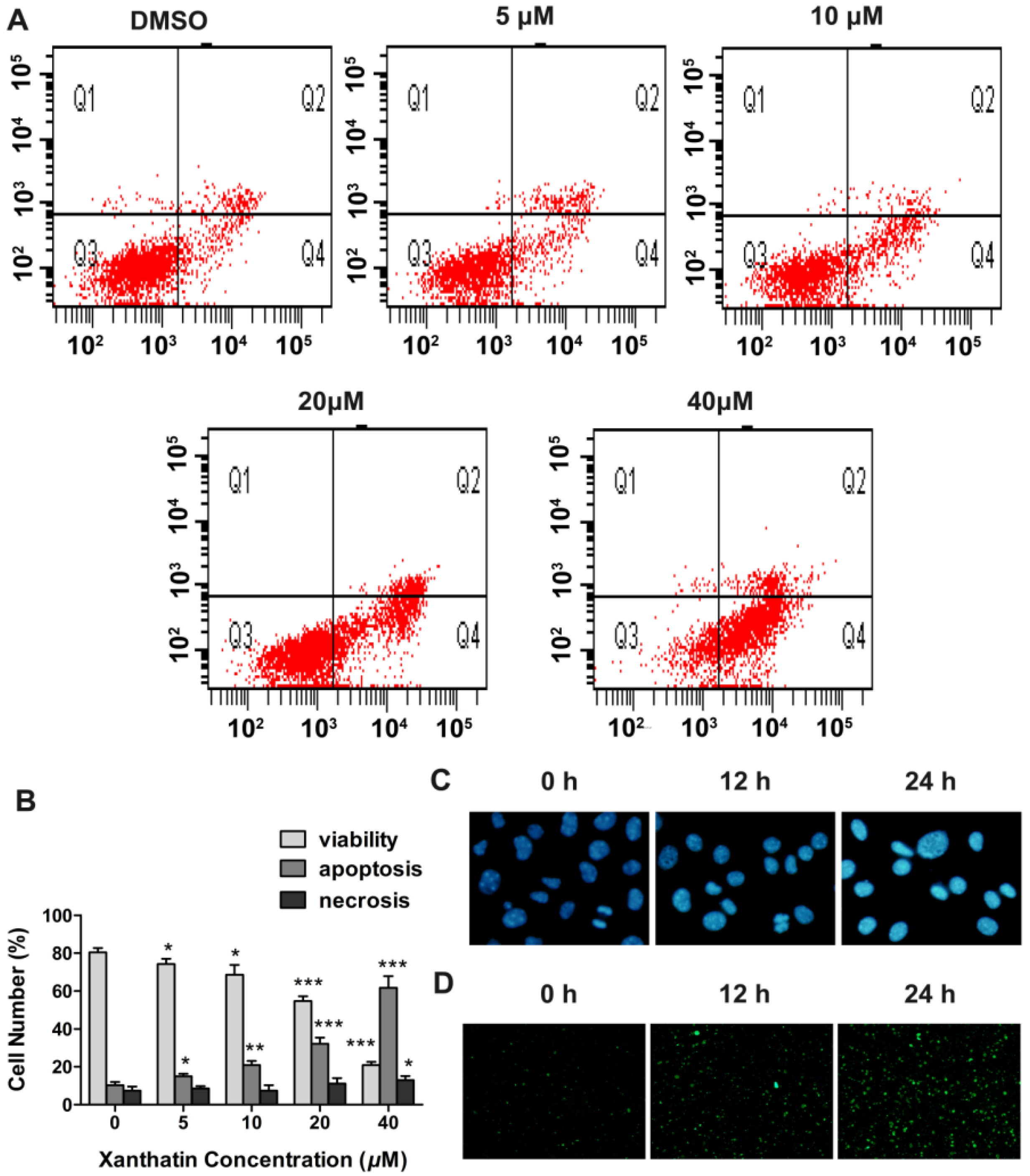
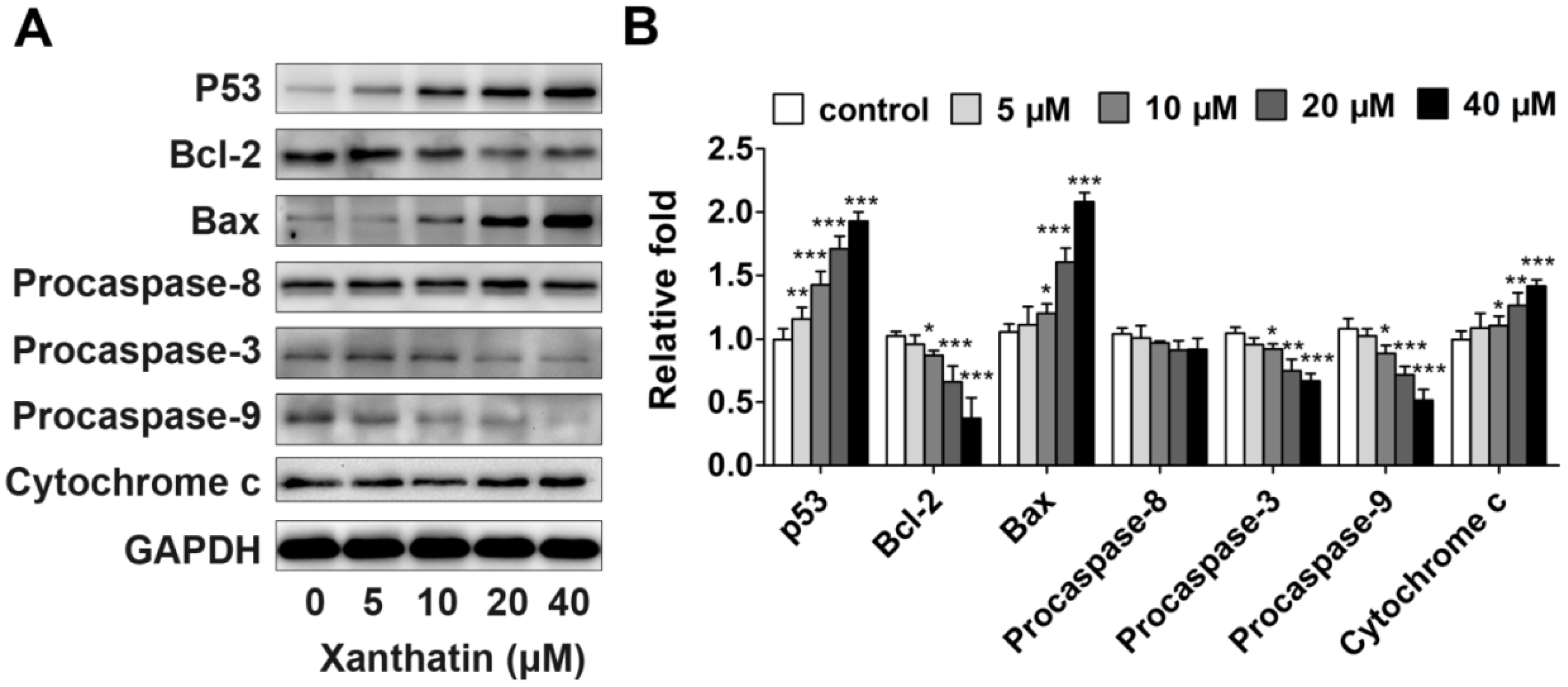
2.1.3. Xanthatin Induced Apoptosis via p53 Activation and Intrinsic Pathway in A549 Cells
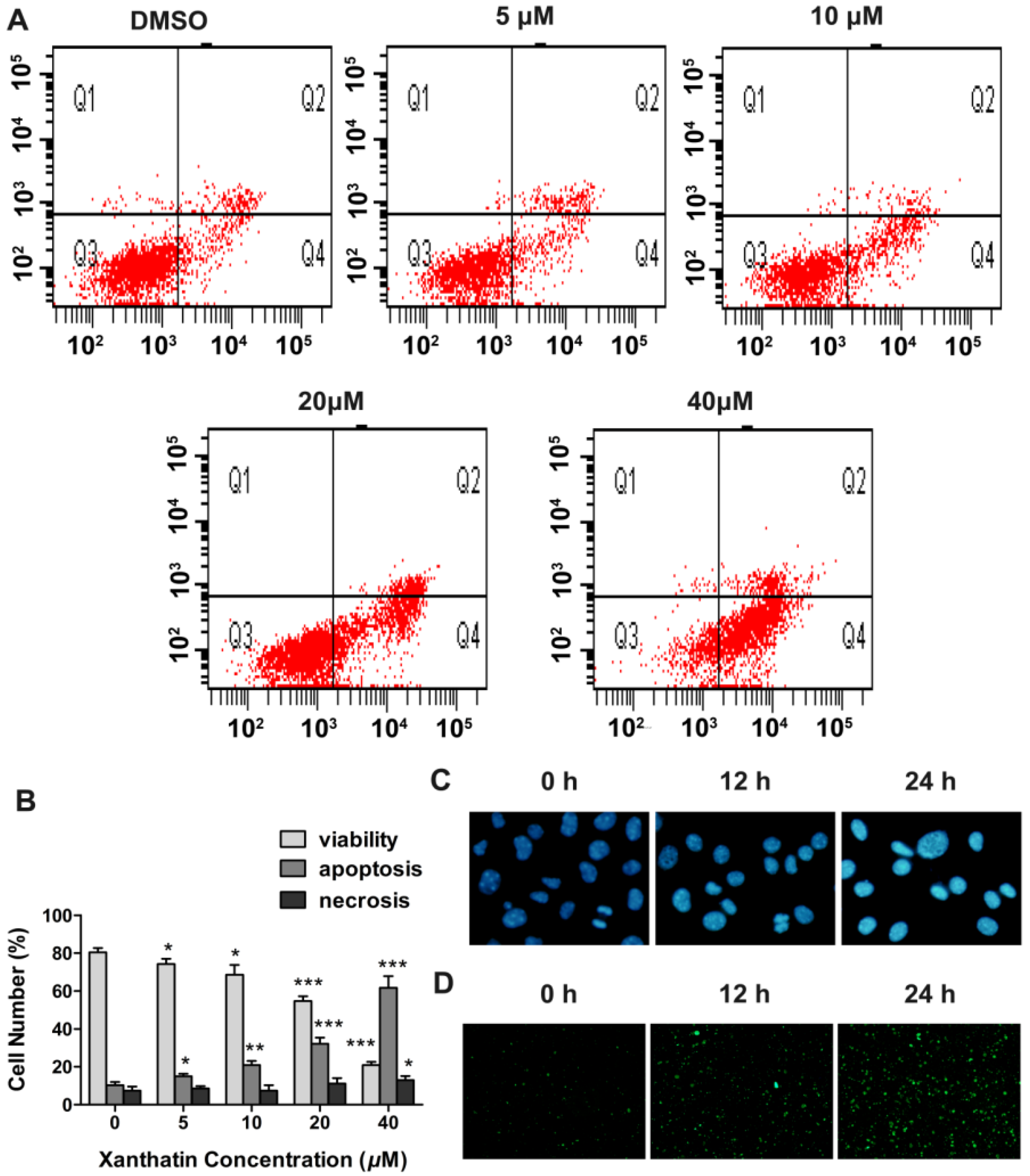
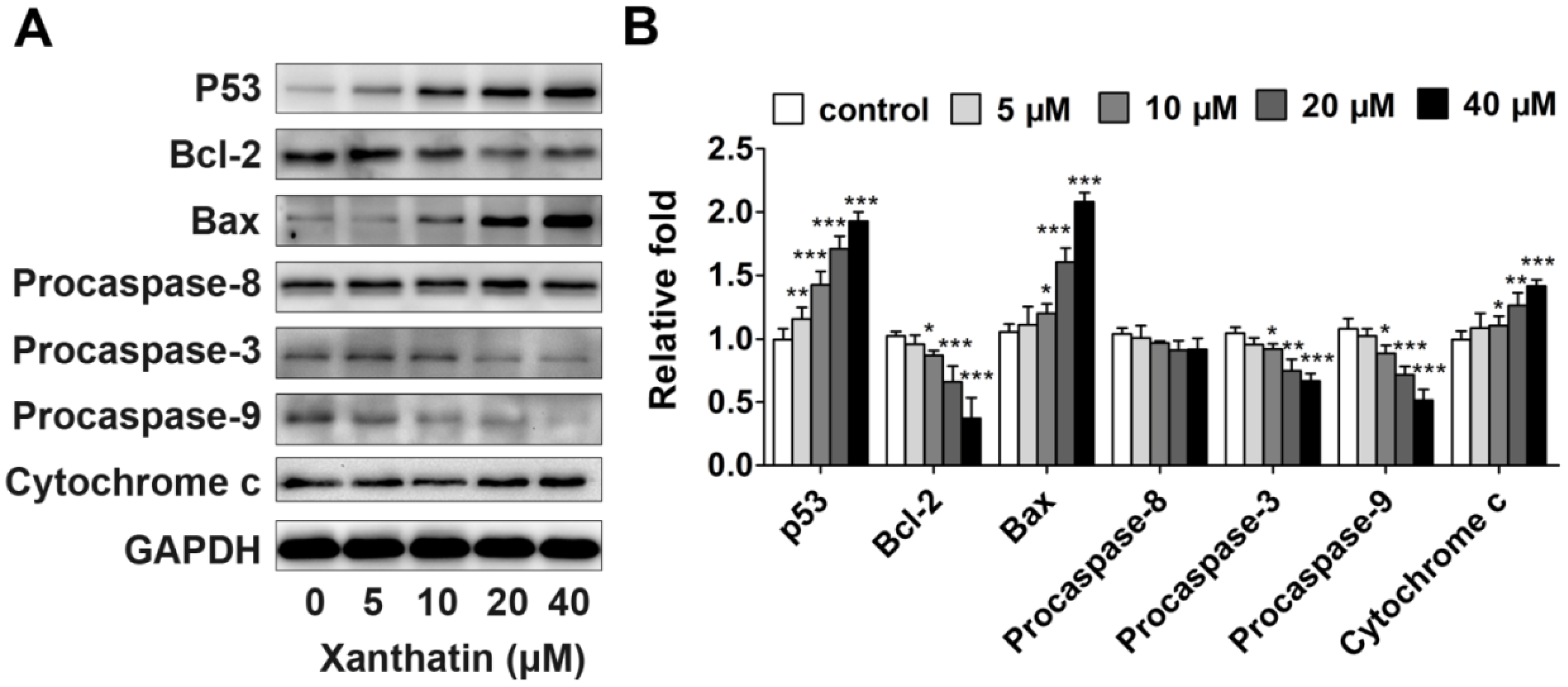
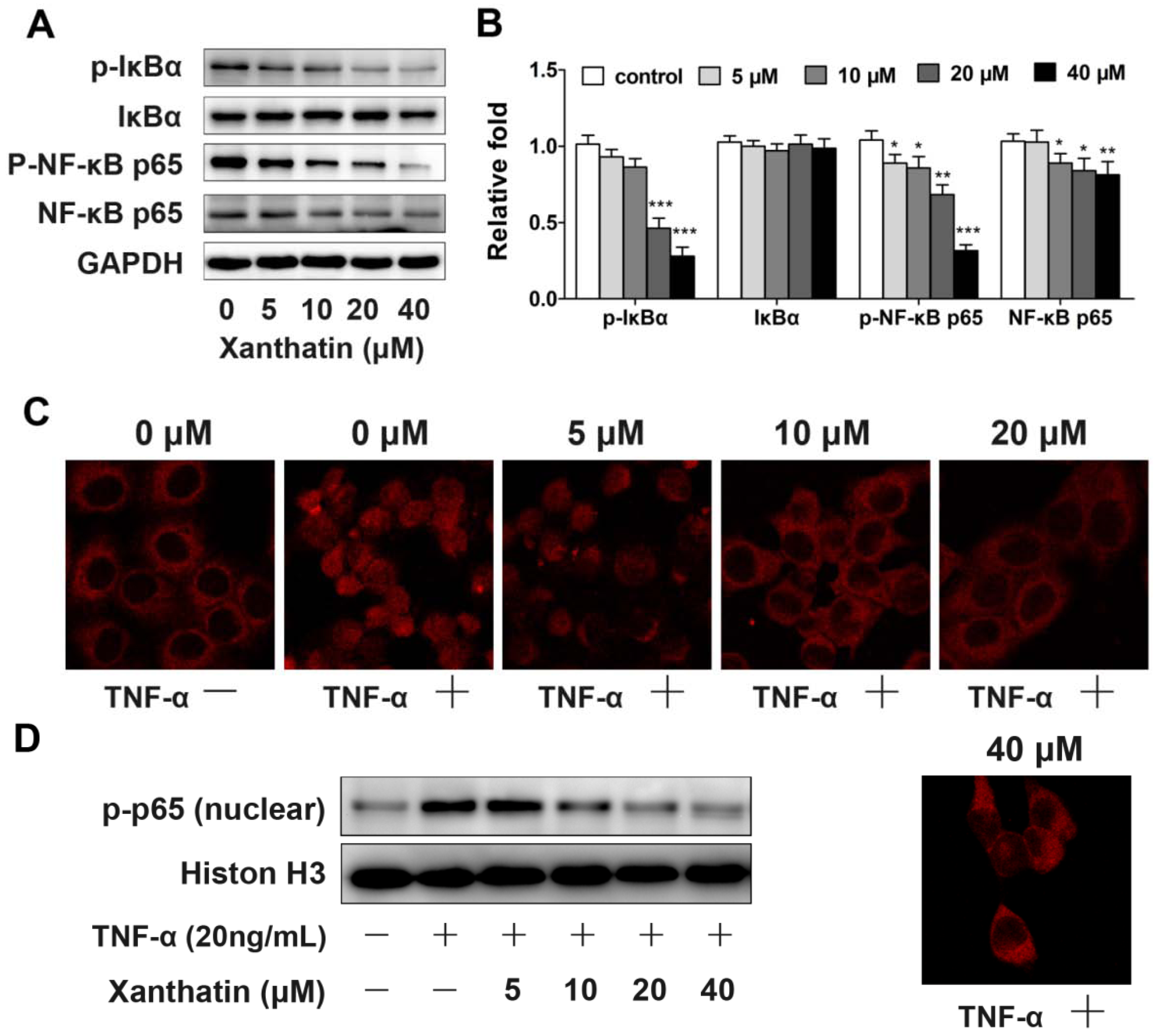
2.1.4. Xanthatin Disrupted NF-κB Signaling in A549 Cells
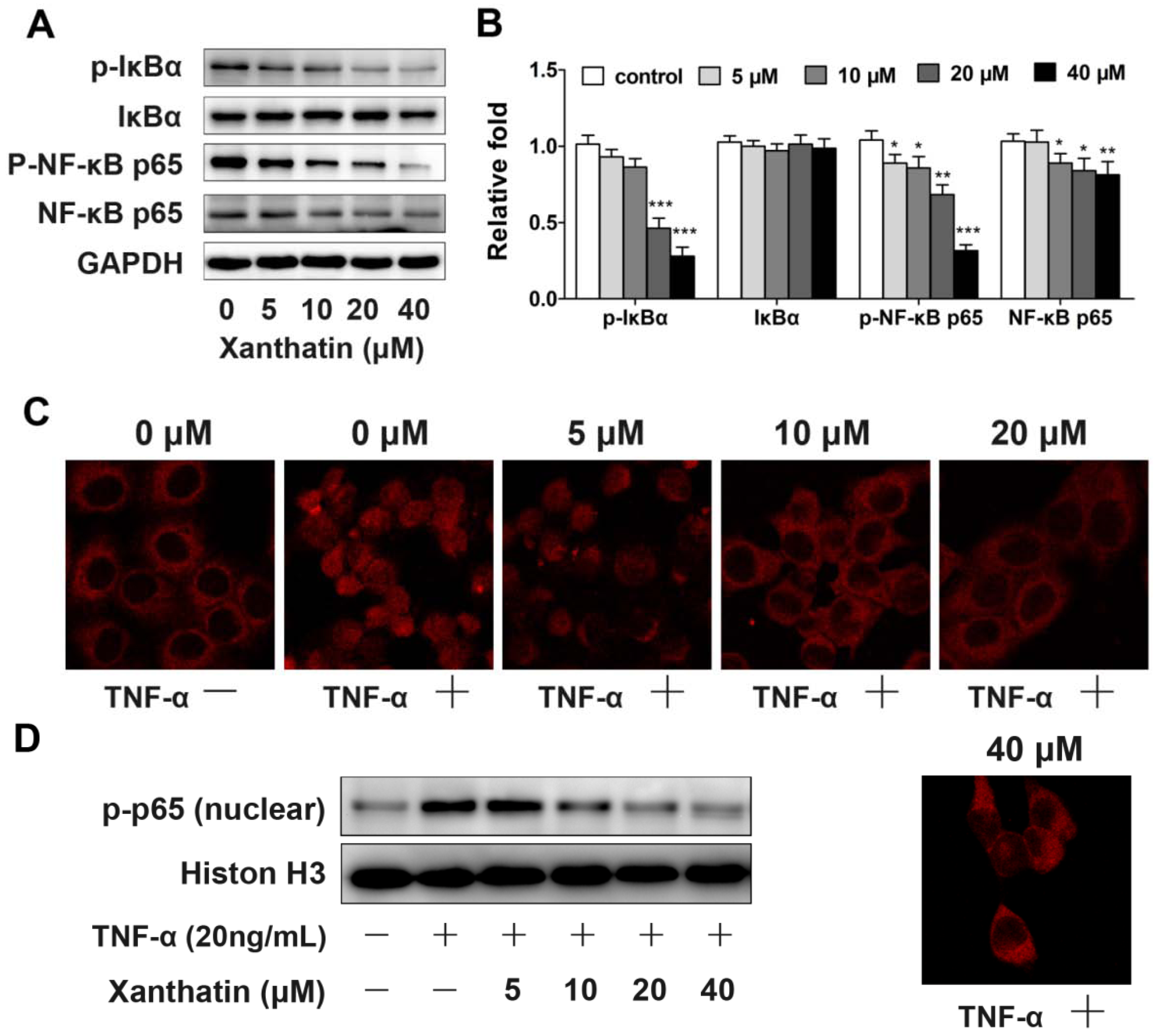
2.2. Discussion
3. Experimental
3.1. Xanthatin Preparation
3.2. Regents
3.3. Cell Culture
3.4. Cell Morphology Assessment
3.5. MTS Assay
3.6. Cell Cycle Analysis
3.7. Annexin-V/PI Double Staining Assay
3.8. Hoechst 33258 Staining Assay
3.9. Western Blot Analysis
3.10. NF-κB (p65) Translocation
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Demain, A.L.; Vaishnav, P. Natural products for cancer chemotherapy. Microb. Biotechnol. 2011, 4, 687–699. [Google Scholar]
- Vasas, A.; Hohmann, J. Xanthane sesquiterpenoids: Structure, synthesis and biological activity. Nat. Prod. Rep. 2011, 28, 824–842. [Google Scholar]
- Zhang, S.; Won, Y.K.; Ong, C.N.; Shen, H.M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem. Anticancer Agents 2005, 5, 239–249. [Google Scholar]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Saliba, N.A.; Darwiche, N. What made sesquiterpene lactones reach cancer clinical trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef]
- Ramirez-Erosa, I.; Huang, Y.; Hickie, R.A.; Sutherland, R.G.; Barl, B. Xanthatin and xanthinosin from the burs of Xanthium strumarium L. as potential anticancer agents. Can. J. Physiol. Pharmacol. 2007, 85, 1160–1172. [Google Scholar] [CrossRef]
- Kovacs, A.; Vasas, A.; Forgo, P.; Rethy, B.; Zupko, I.; Hohmann, J. Xanthanolides with antitumour activity from Xanthium italicum. Z. Naturforsch. C 2009, 64, 343–349. [Google Scholar]
- Maddika, S.; Ande, S.R.; Panigrahi, S.; Paranjothy, T.; Weglarczyk, K.; Zuse, A.; Eshraghi, M.; Manda, K.D.; Wiechec, E.; Los, M. Cell survival, cell death and cell cycle pathways are interconnected: Implications for cancer therapy. Drug Resist. Updat. 2007, 10, 13–29. [Google Scholar]
- Wu, Y.; Zhou, B.P. Inflammation: A driving force speeds cancer metastasis. Cell Cycle 2009, 8, 3267–3273. [Google Scholar]
- Yoon, J.H.; Lim, H.J.; Lee, H.J.; Kim, H.D.; Jeon, R.; Ryu, J.H. Inhibition of lipopolysaccharide-induced inducible nitric oxide synthase and cyclooxygenase-2 expression by xanthanolides isolated from Xanthium strumarium. Bioorg. Med. Chem. Lett. 2008, 18, 2179–2182. [Google Scholar] [CrossRef]
- Polager, S.; Ginsberg, D. p53 and E2f: Partners in life and death. Nat. Rev. Cancer 2009, 9, 738–748. [Google Scholar]
- Hu, W.; Kavanagh, J.J. Anticancer therapy targeting the apoptotic pathway. Lancet Oncol. 2003, 4, 721–729. [Google Scholar]
- Coultas, L.; Strasser, A. The role of the Bcl-2 protein family in cancer. Semin. Cancer Biol. 2003, 13, 115–123. [Google Scholar]
- Lavrik, I.N.; Golks, A.; Krammer, P.H. Caspases: Pharmacological manipulation of cell death. J. Clin. Invest. 2005, 115, 2665–2672. [Google Scholar]
- Penissi, A.B.; Vera, M.E.; Mariani, M.L.; Rudolph, M.I.; Cenal, J.P.; de Rosas, J.C.; Fogal, T.H.; Tonn, C.E.; Favier, L.S.; Giordano, O.S.; et al. Novel anti-ulcer alpha,beta-unsaturated lactones inhibit compound 48/80-induced mast cell degranulation. Eur. J. Pharmacol. 2009, 612, 122–130. [Google Scholar] [CrossRef]
- Roussakis, C.; Chinou, I.; Vayas, C.; Harvala, C.; Verbist, J.F. Cytotoxic activity of xanthatin and the crude extracts of Xanthium strumarium. Planta Med. 1994, 60, 473–474. [Google Scholar] [CrossRef]
- Takeda, S.; Matsuo, K.; Yaji, K.; Okajima-Miyazaki, S.; Harada, M.; Miyoshi, H.; Okamoto, Y.; Amamoto, T.; Shindo, M.; Omiecinski, C.J.; et al. (-)-anthatin selectively induces GADD45gamma and stimulates caspase-independent cell death in human breast cancer MDA-MB-231 cells. Chem. Res. Toxicol. 2011, 24, 855–865. [Google Scholar] [CrossRef]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar]
- Poonepalli, A.; Banerjee, B.; Ramnarayanan, K.; Palanisamy, N.; Putti, T.C.; Hande, M.P. Telomere-mediated genomic instability and the clinico-pathological parameters in breast cancer. Genes Chromosomes Cancer 2008, 47, 1098–1109. [Google Scholar]
- Siegel, R.M. Caspases at the crossroads of immune-cell life and death. Nat. Rev. Immunol. 2006, 6, 308–317. [Google Scholar]
- Rowinsky, E.K. Targeted induction of apoptosis in cancer management: The emerging role of tumor necrosis factor-related apoptosis-inducing ligand receptor activating agents. J. Clin. Oncol. 2005, 23, 9394–9407. [Google Scholar]
- Kawabe, T. G2 checkpoint abrogators as anticancer drugs. Mol. Cancer Ther. 2004, 3, 513–519. [Google Scholar]
- Abraham, R.T. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 2001, 15, 2177–2196. [Google Scholar]
- Bartek, J.; Lukas, J. Chk1 and Chk2 kinases in checkpoint control and cancer. Cancer Cell 2003, 3, 421–429. [Google Scholar]
- Bucher, N.; Britten, C.D. G2 checkpoint abrogation and checkpoint kinase-1 targeting in the treatment of cancer. Br. J. Cancer 2008, 98, 523–528. [Google Scholar]
- Udayakumar, T.; Shareef, M.M.; Diaz, D.A.; Ahmed, M.M.; Pollack, A. The E2F1/Rb and p53/MDM2 pathways in DNA repair and apoptosis: Understanding the crosstalk to develop novel strategies for prostate cancer radiotherapy. Semin. Radiat. Oncol. 2010, 20, 258–266. [Google Scholar]
- Leu, J.I.; Dumont, P.; Hafey, M.; Murphy, M.E.; George, D.L. Mitochondrial p53 activates Bak and causes disruption of a Bak-Mcl1 complex. Nat. Cell Biol. 2004, 6, 443–450. [Google Scholar]
- Sarkar, F.H.; Li, Y. NF-kappaB: A potential target for cancer chemoprevention and therapy. Front. Biosci. 2008, 13, 2950–2959. [Google Scholar]
- Hehner, S.P.; Heinrich, M.; Bork, P.M.; Vogt, M.; Ratter, F.; Lehmann, V.; Schulze-Osthoff, K.; Droge, W.; Schmitz, M.L. Sesquiterpene lactones specifically inhibit activation of NF-kappa B by preventing the degradation of I kappa B-alpha and I kappa B-beta. J. Biol. Chem. 1998, 273, 1288–1297. [Google Scholar]
- Zhang, W.; Han, W.; Li, Y.; Zhang, S.; Zhao, D. Chemical constituents from Xanthium mongolicum. Zhongguo Zhong Yao Za Zhi 2009, 34, 1687–1689. [Google Scholar]
- Coward, P.; Wada, H.G.; Falk, M.S.; Chan, S.D.; Meng, F.; Akil, H.; Conklin, B.R. Controlling signaling with a specifically designed Gi-coupled receptor. Proc. Natl. Acad. Sci. USA 1998, 95, 352–357. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, L.; Ruan, J.; Yan, L.; Li, W.; Wu, Y.; Tao, L.; Zhang, F.; Zheng, S.; Wang, A.; Lu, Y. Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells. Molecules 2012, 17, 3736-3750. https://doi.org/10.3390/molecules17043736
Zhang L, Ruan J, Yan L, Li W, Wu Y, Tao L, Zhang F, Zheng S, Wang A, Lu Y. Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells. Molecules. 2012; 17(4):3736-3750. https://doi.org/10.3390/molecules17043736
Chicago/Turabian StyleZhang, Lei, Junshan Ruan, Linggeng Yan, Weidong Li, Yu Wu, Li Tao, Feng Zhang, Shizhong Zheng, Aiyun Wang, and Yin Lu. 2012. "Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells" Molecules 17, no. 4: 3736-3750. https://doi.org/10.3390/molecules17043736
APA StyleZhang, L., Ruan, J., Yan, L., Li, W., Wu, Y., Tao, L., Zhang, F., Zheng, S., Wang, A., & Lu, Y. (2012). Xanthatin Induces Cell Cycle Arrest at G2/M Checkpoint and Apoptosis via Disrupting NF-κB Pathway in A549 Non-Small-Cell Lung Cancer Cells. Molecules, 17(4), 3736-3750. https://doi.org/10.3390/molecules17043736