Endo-S-c-di-GMP Analogues-Polymorphism and Binding Studies with Class I Riboswitch

Abstract
:
1. Introduction
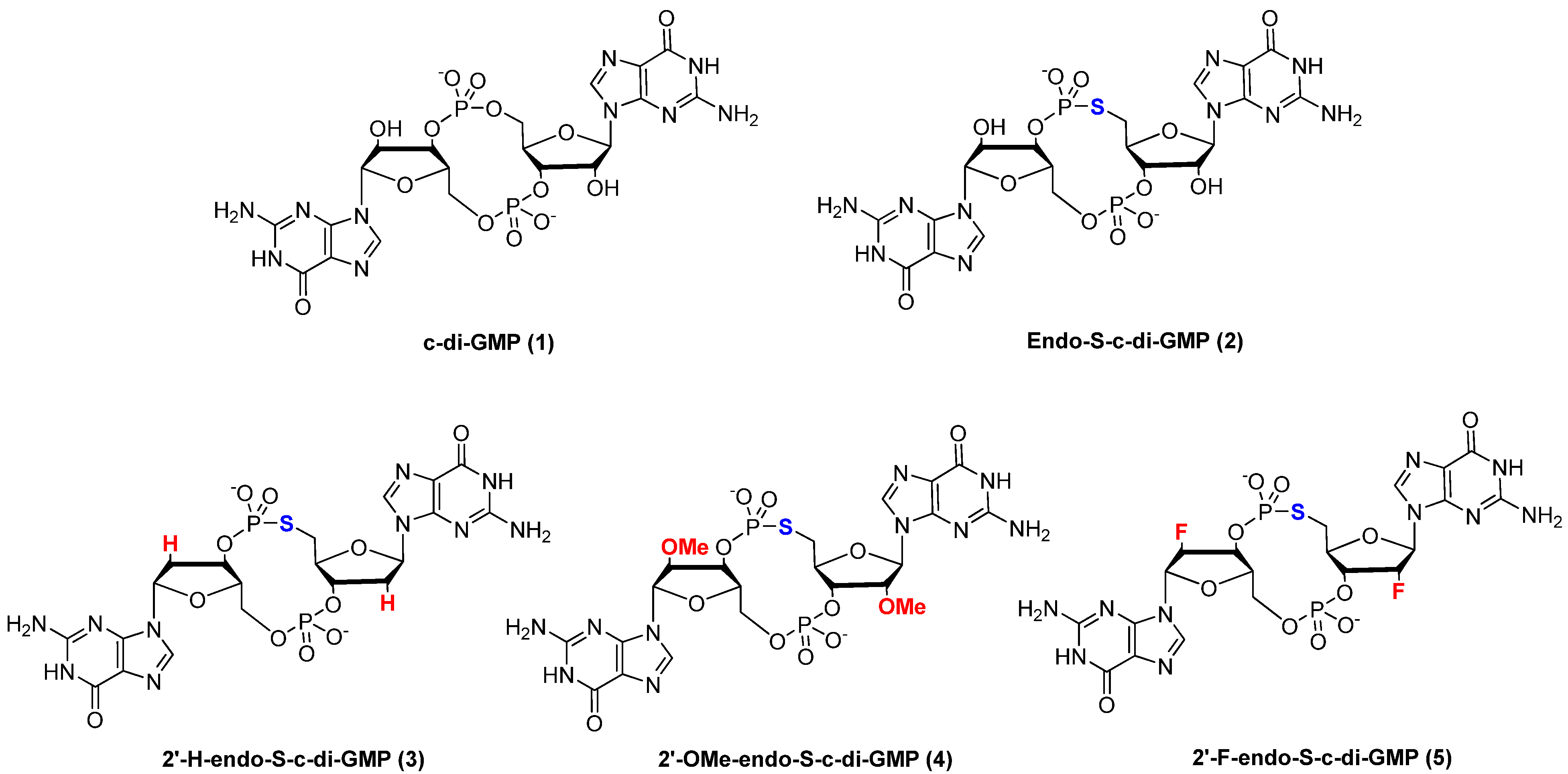
2. Results and Discussion
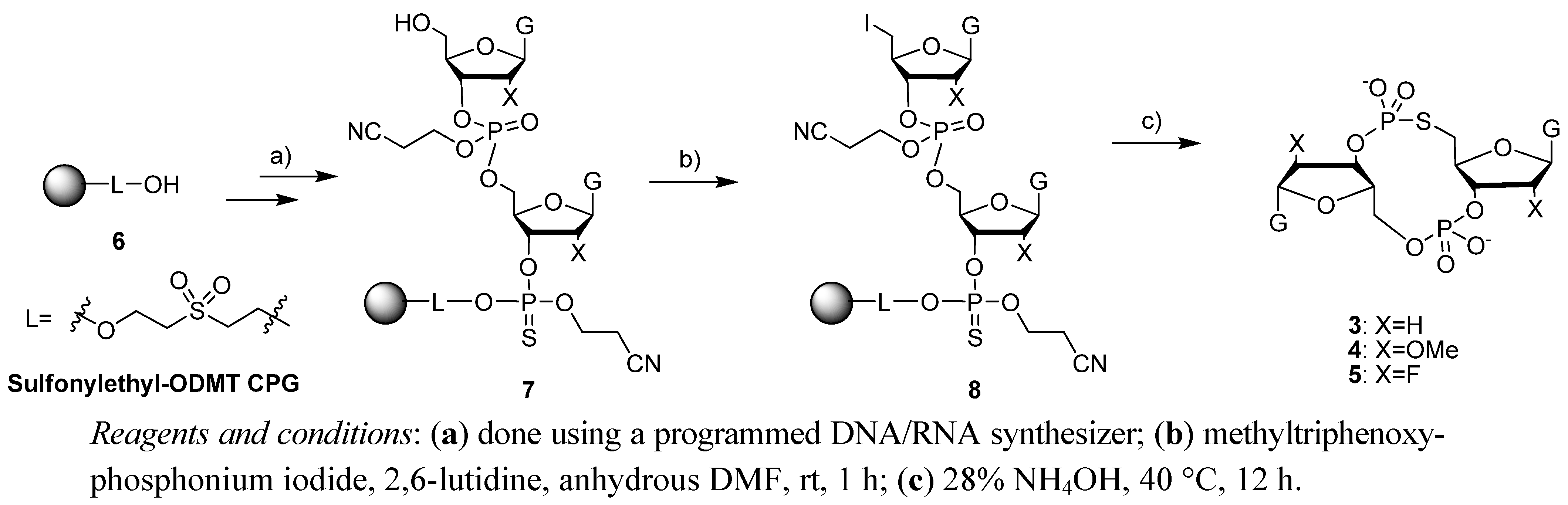
2.1. Synthesis of c-di-GMP Analogues
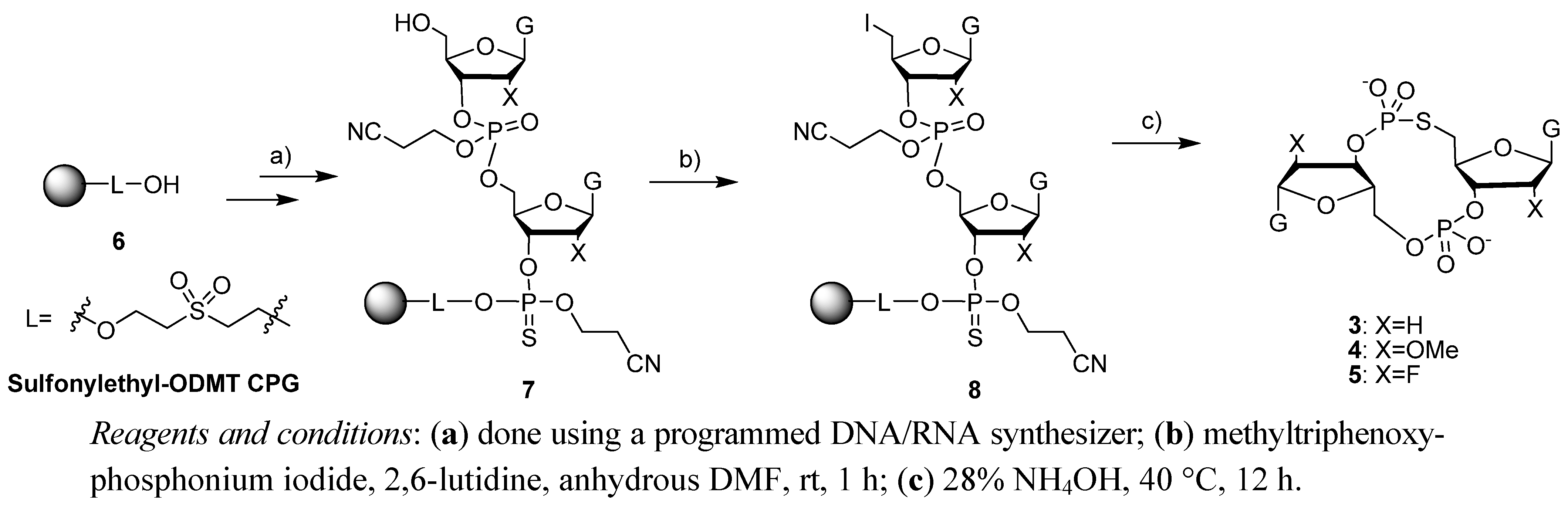
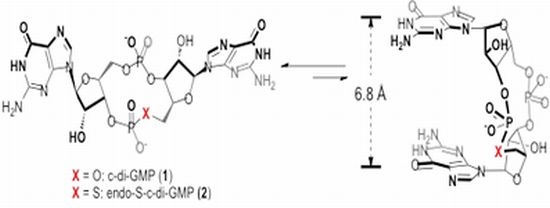
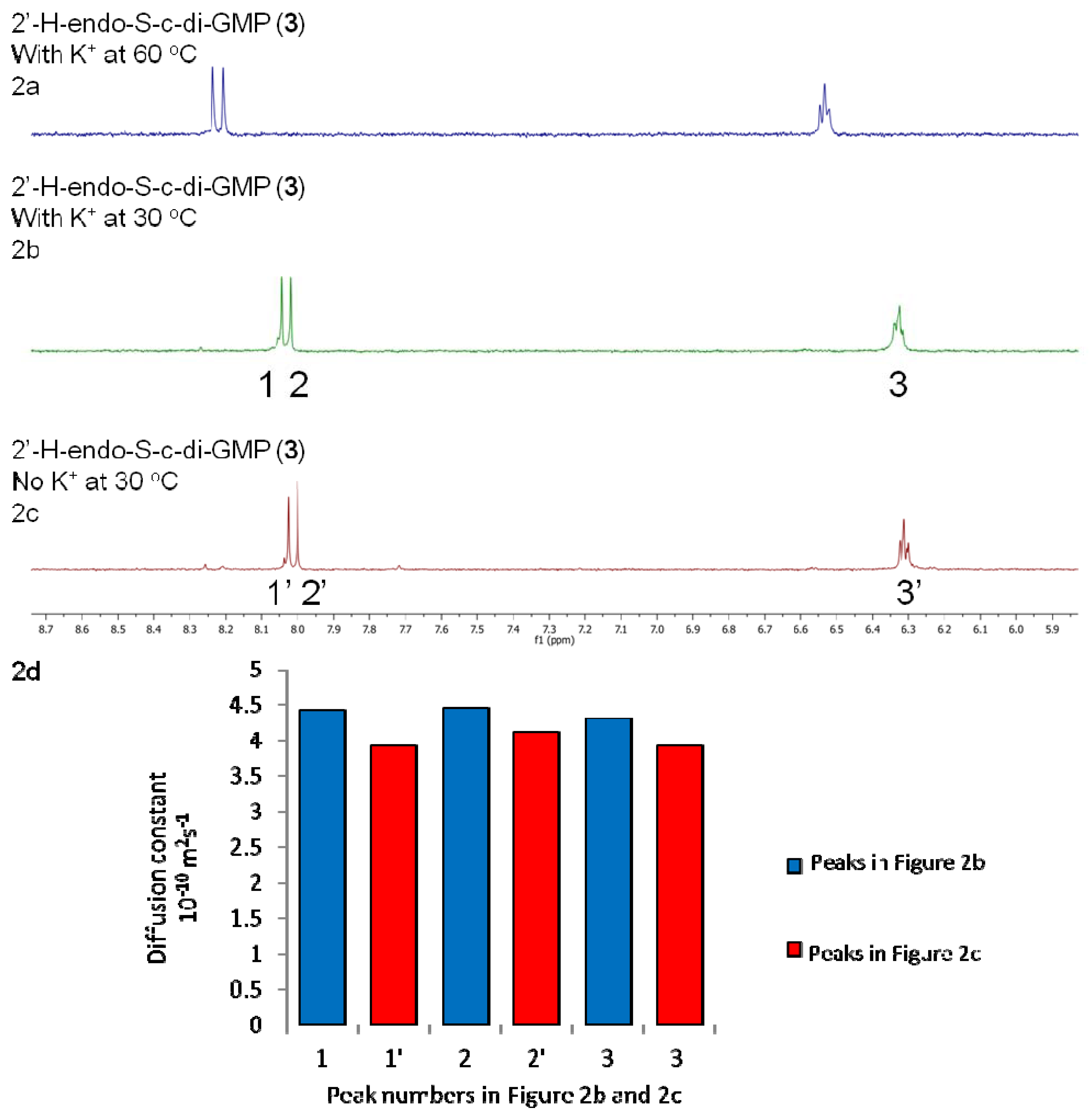
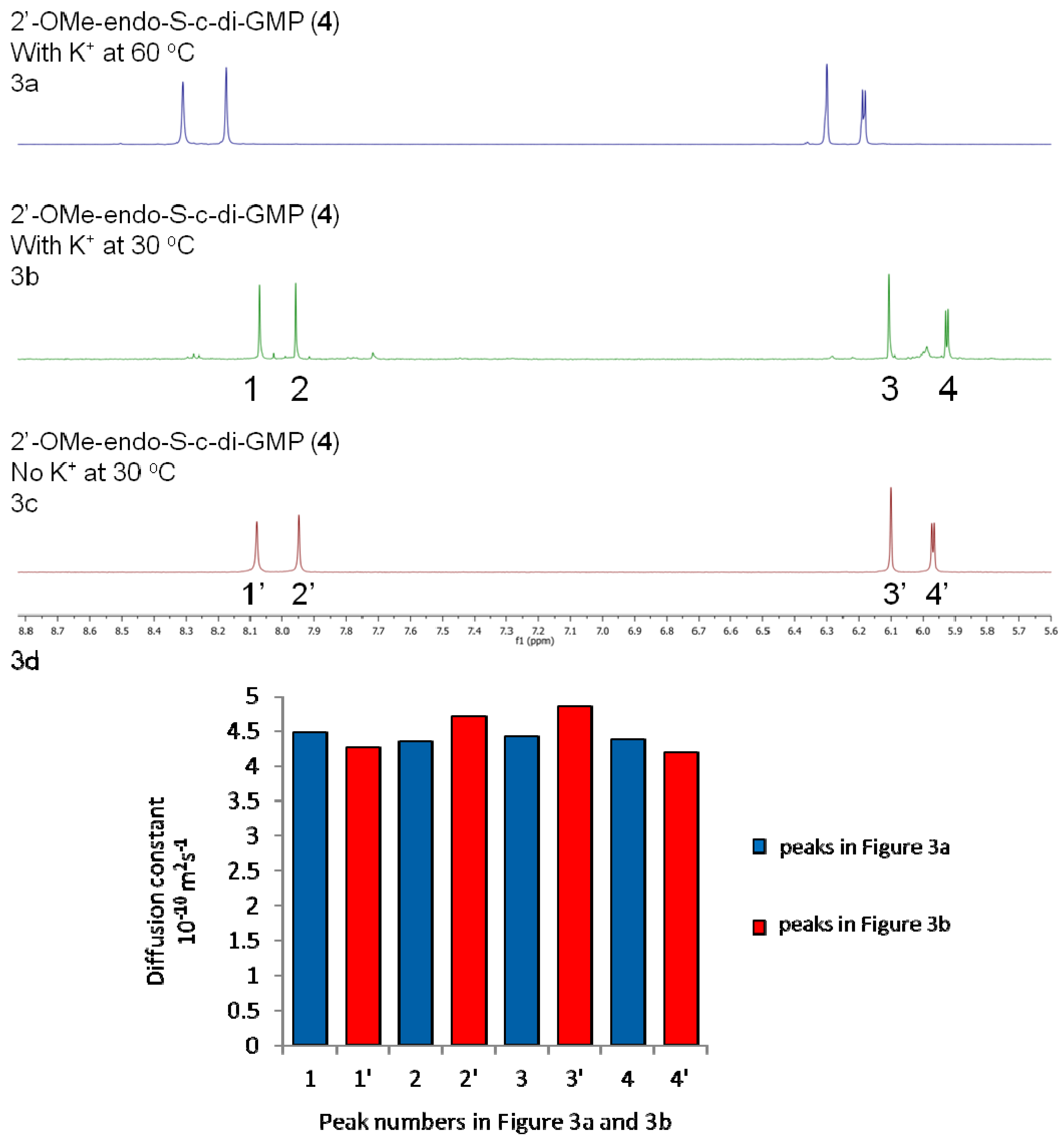
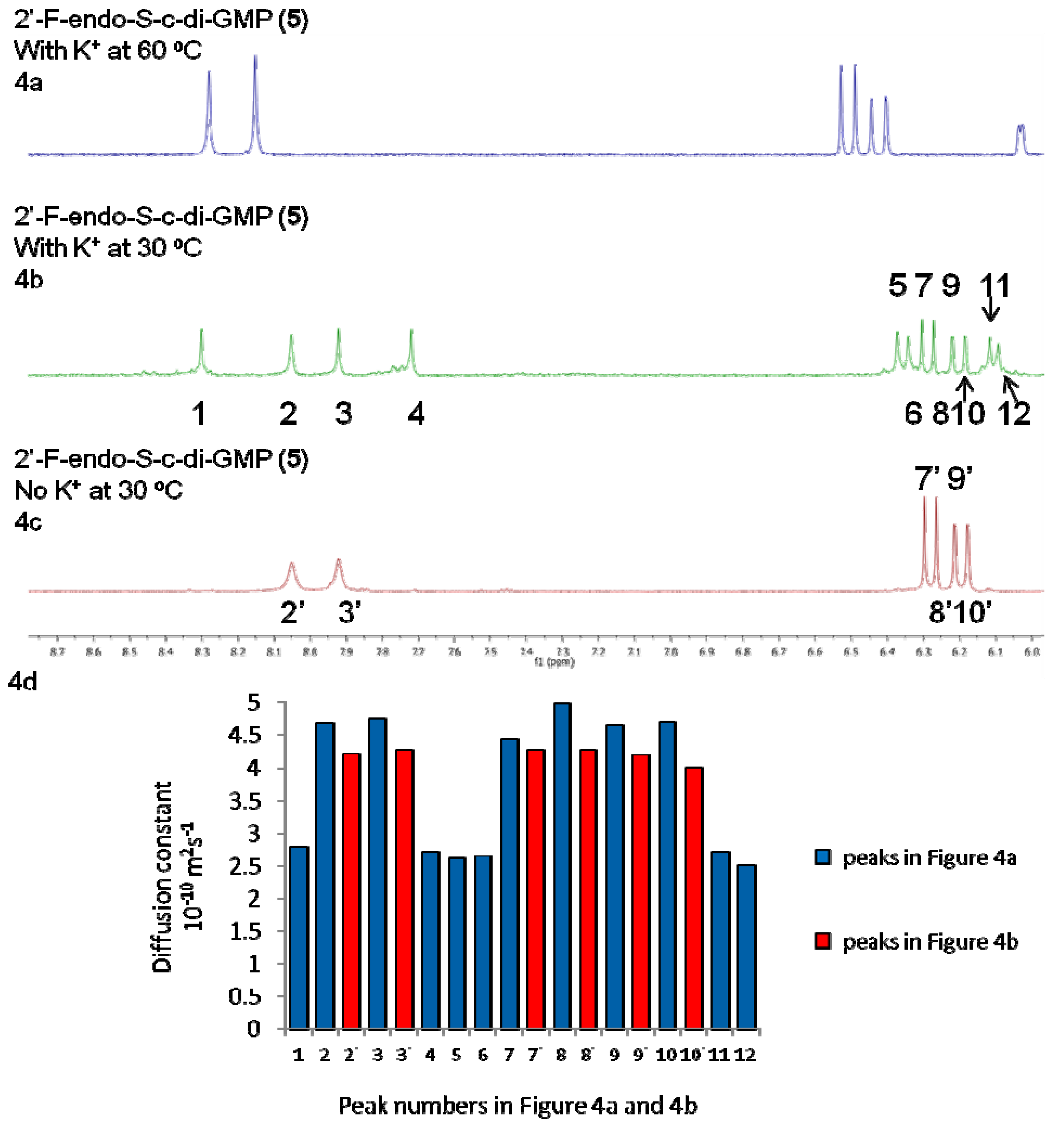
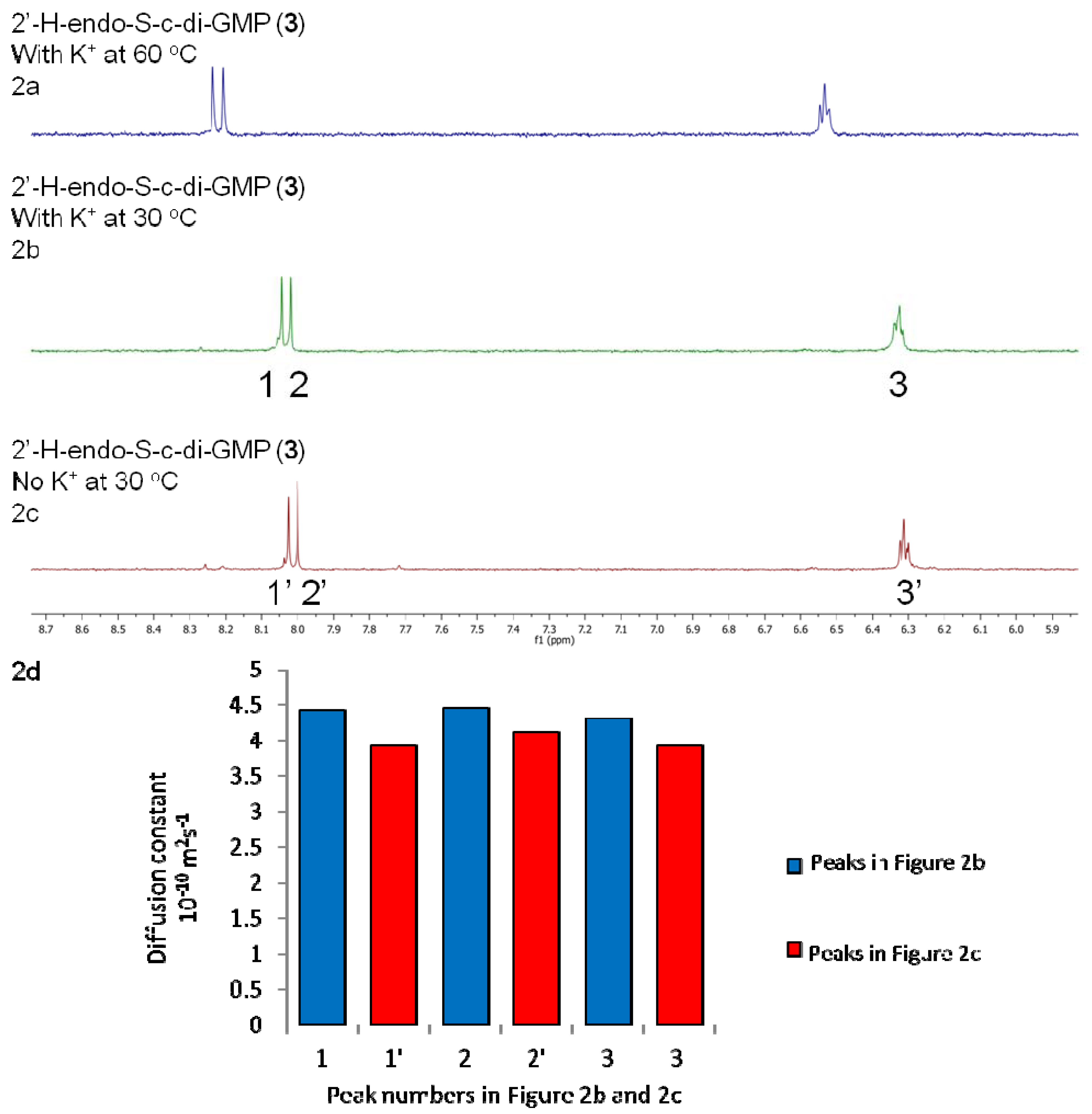
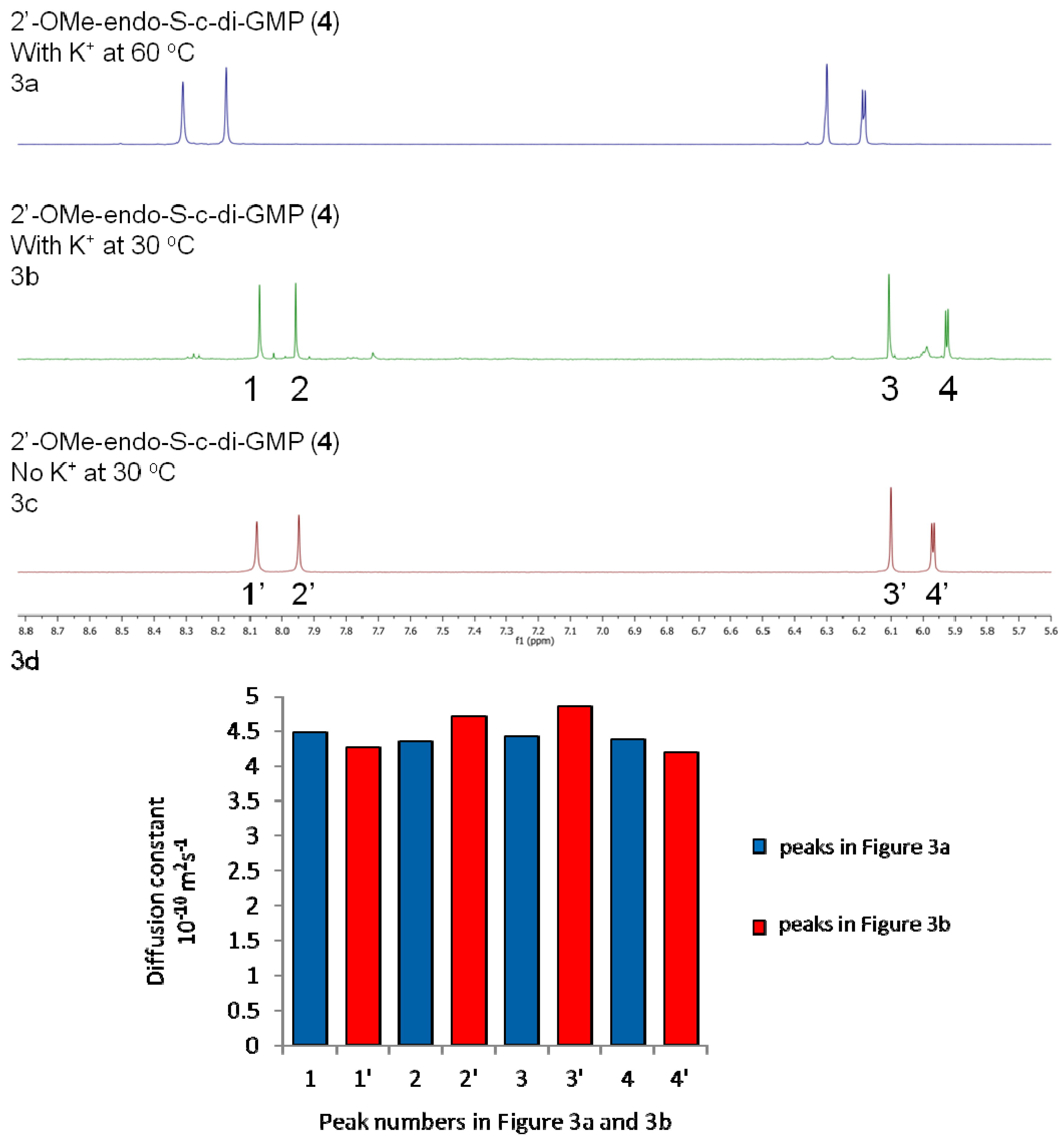
2.2. Polymorphism of c-di-GMP Analogues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ΔEsol(open–closed) a | ratio (open:closed) b | |
|---|---|---|
| c-di-GMP (1) | 1.9 | 1:25 |
| endo-S-c-di-GMP (2) | 1.3 | 1:9 |
| 2'-H-endo-S-c-di-GMP (3) | −0.3 | 2:1 |
| 2'-OMe-endo-S-c-di-GMP (4) | −2.2 | 39:1 |
| 2'-F-endo-S-c-di-GMP (5) | 0.5 | 1:3 |
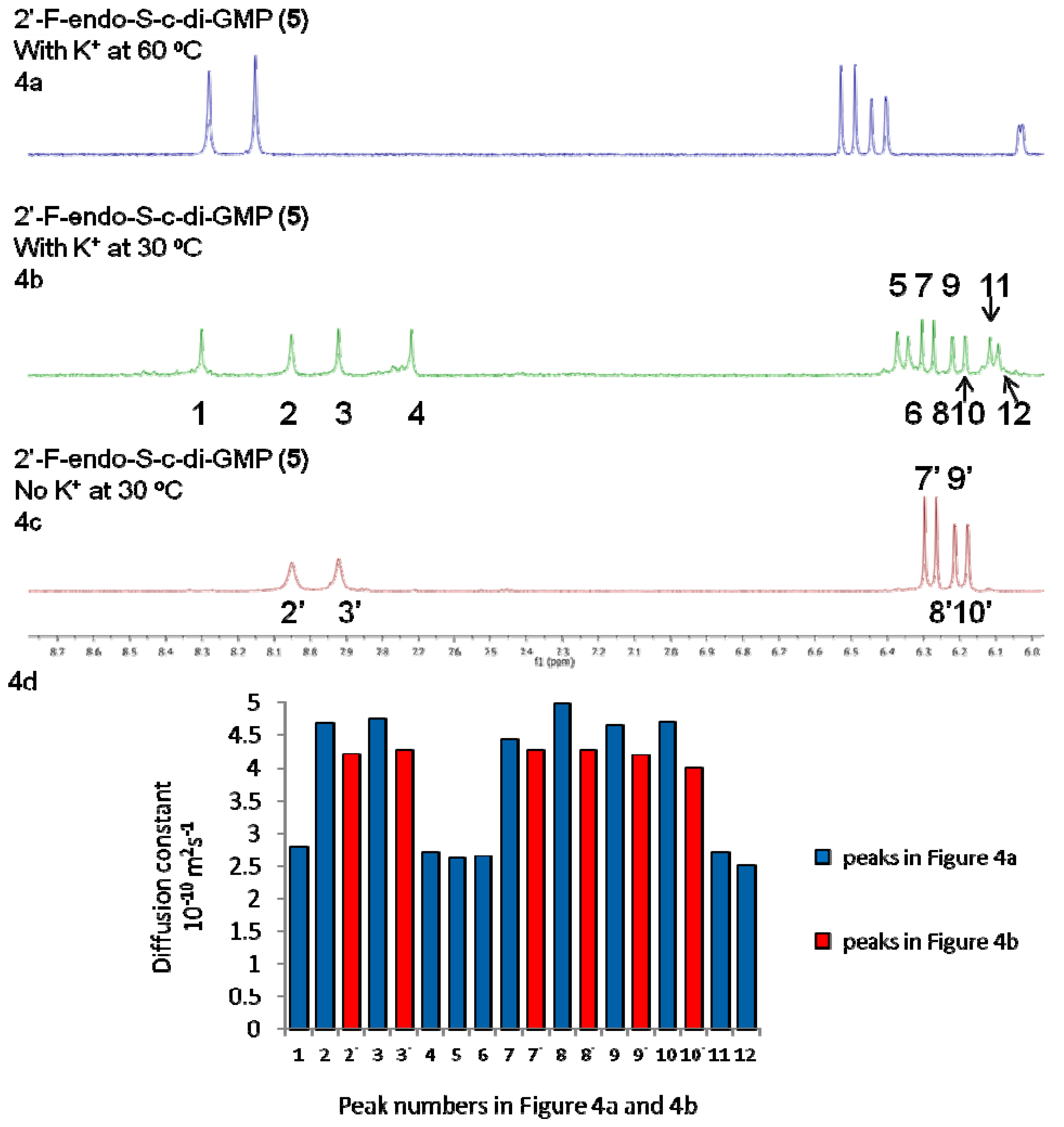
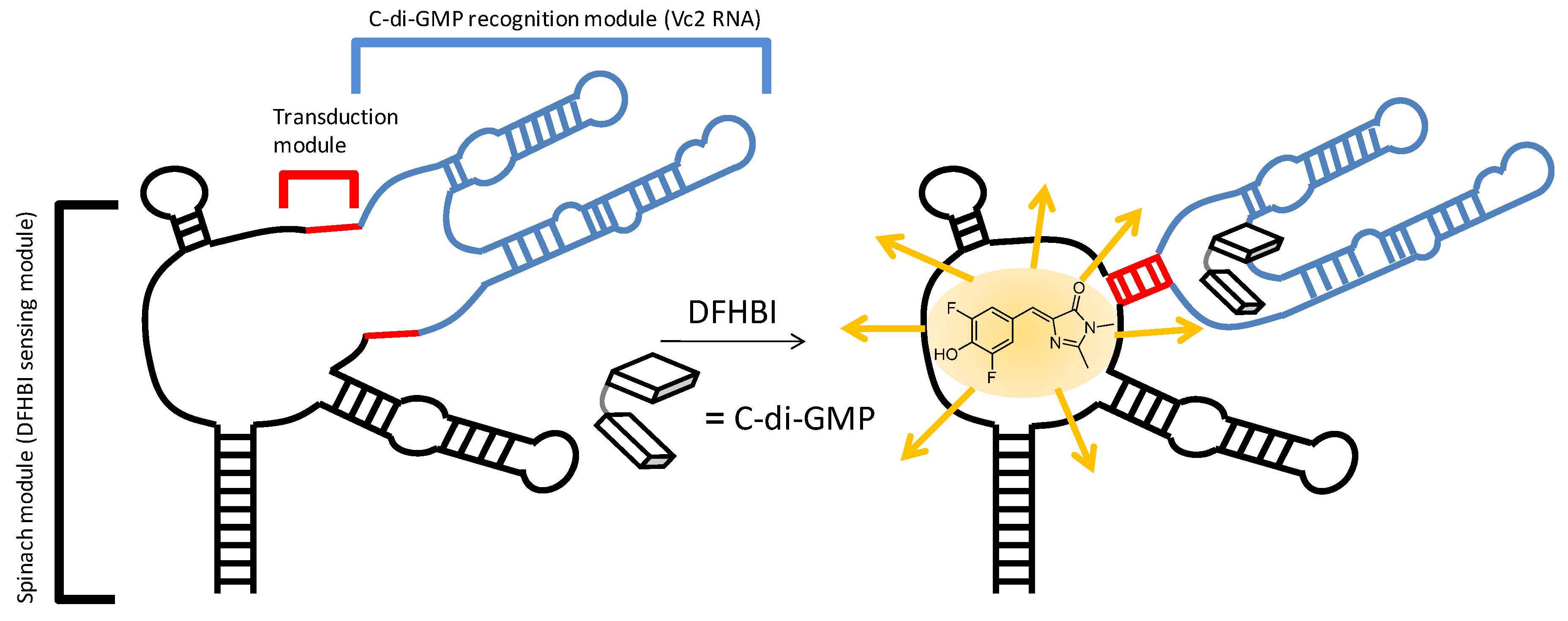
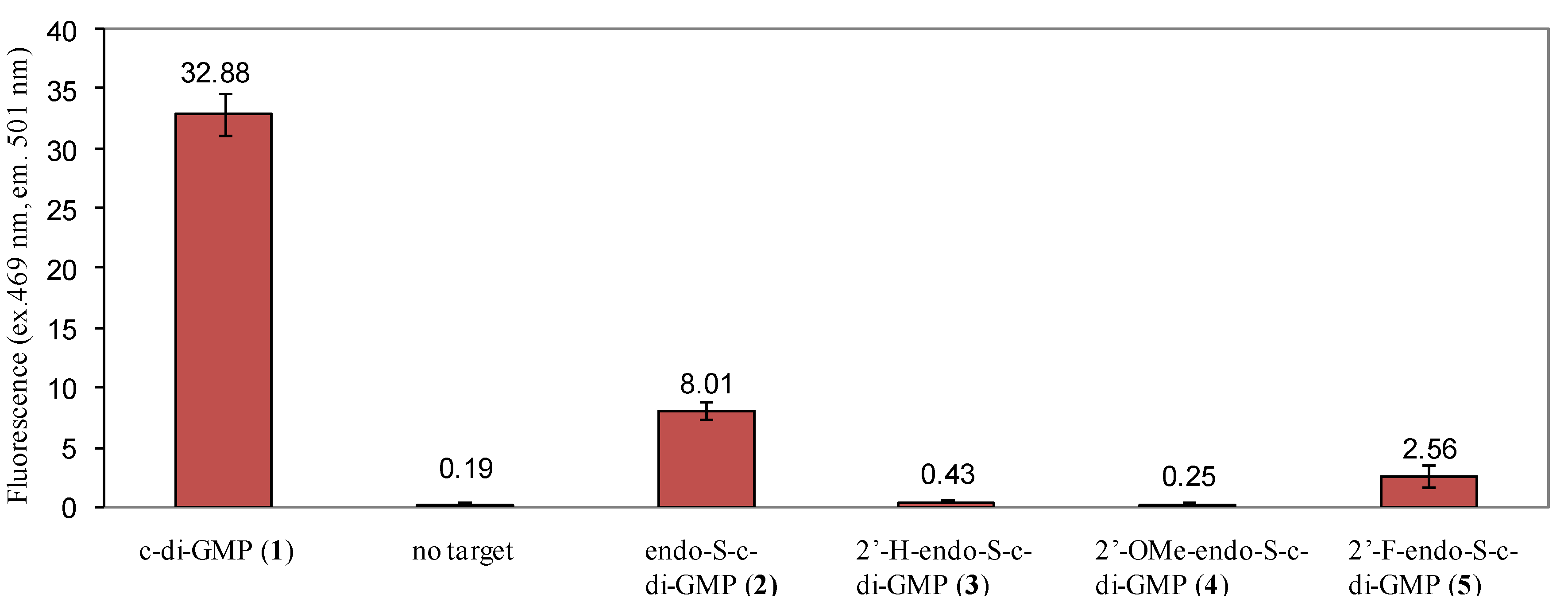
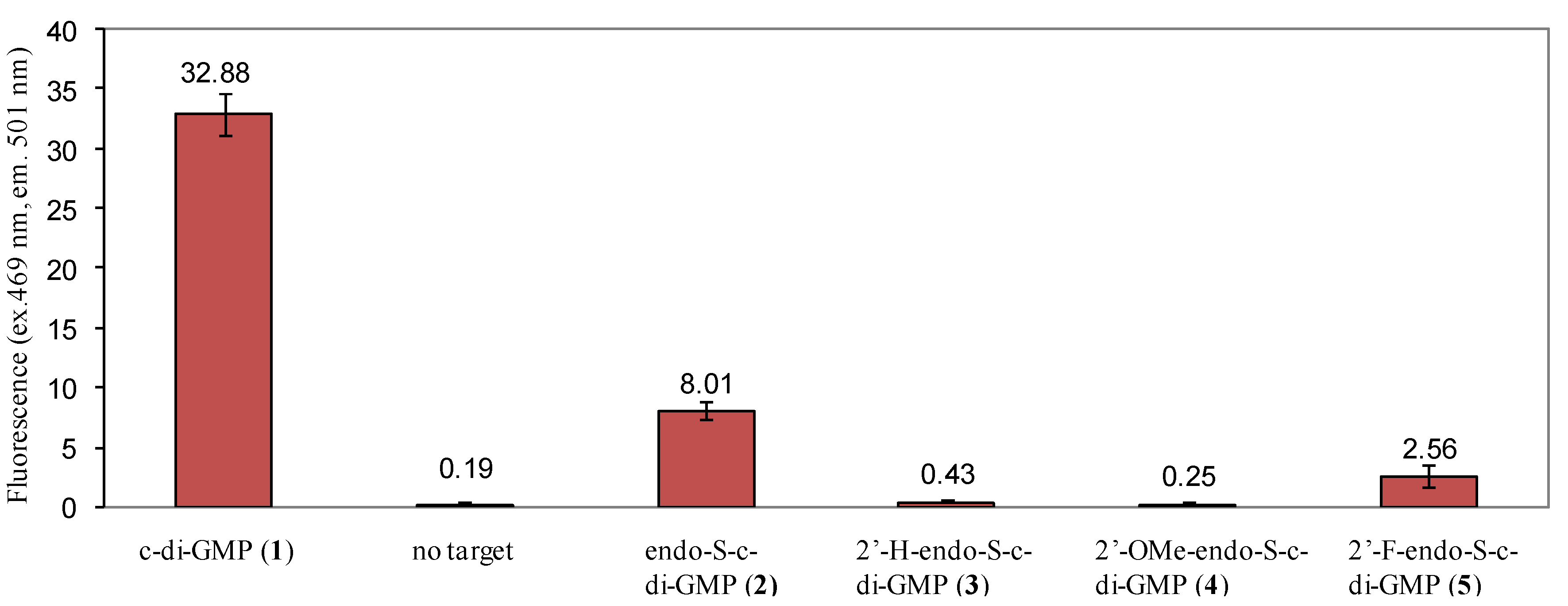
2.3. Binding of c-di-GMP and Analogues (2–5) to Vc2 c-di-GMP Riboswitch
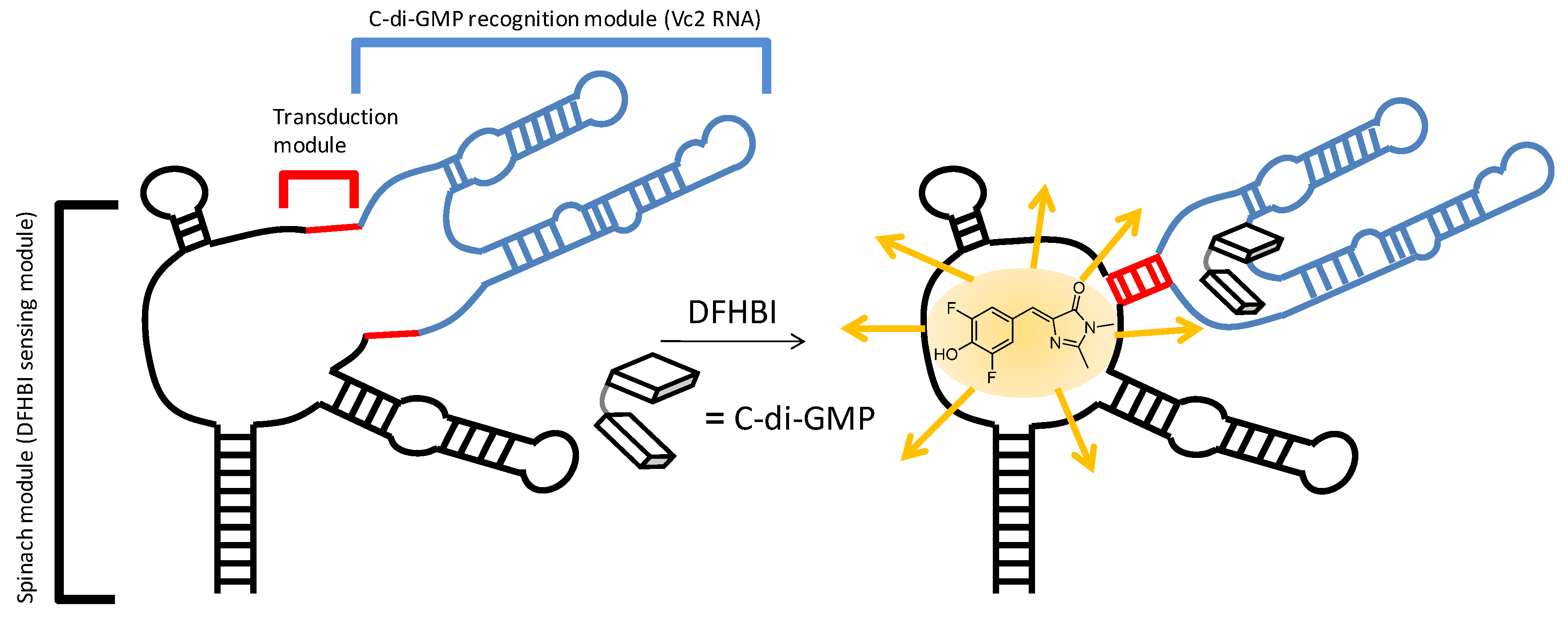
3. Experimental
3.1. General
3.2. Synthesis
3.3. Sample Preparation for Spectrometric Measurements
3.4. DOSY NMR Experiments and T1/T2 Relaxation Analysis
3.5. Sample Preparation of Fluorescence Measurement
3.6. Fluorescence Measurement
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the materials are available from the authors.
References and Notes
- Ross, P.; Weinhouse, H.; Aloni, Y.; Michaeli, D.; Weinberger-Ohana, P.; Mayer, R.; Braun, S.; de Vroom, E.; van der Marel, G.A.; van Boom, H.H.; et al. Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature 1987, 325, 279–281. [Google Scholar]
- Römling, U.; Gomelsky, M.; Galperin, M.Y. C-di-GMP: The dawning of a novel bacterial signalling system. Mol. Microbiol. 2005, 57, 629–639. [Google Scholar] [CrossRef]
- Sondermann, H.; Shikuma, N.J.; Yildiz, F.H. You’ve come a long way: C-di-GMP signaling. Curr. Opin. Microbiol. 2012, 15, 140–146. [Google Scholar] [CrossRef]
- Ryan, R.P.; Tolker-Nielsen, T.; Dow, J.M. When the PilZ don’t work: Effectors for cyclic di-GMP action in bacteria. Trends Microbiol. 2012, 20, 235–242. [Google Scholar] [CrossRef]
- Krasteva, P.V.; Giqlio, K.M.; Sondermann, H. Sensing the messenger: The diverse ways that bacteria signal through c-di-GMP. Protein Sci. 2012, 21, 929–948. [Google Scholar] [CrossRef]
- Römling, U. Cyclic di-GMP, an established secondary messenger still speeding up. Environ. Microbiol. 2012, 14, 1817–1829. [Google Scholar] [CrossRef]
- Chan, C.; Paul, R.; Samoray, D.; Amiot, N.C.; Giese, B.; Jenal, U.; Schirmer, T. Structural basis of activity and allosteric control of diguanylate cyclase. Proc. Natl. Acad. Sci. USA 2004, 101, 17084–17089. [Google Scholar]
- Yang, C.Y.; Chin, K.H.; Chuah, M.L.; Liang, Z.X.; Wang, A.H.; Chou, S.H. The structure and inhibition of a GGDEF diguanylate cyclase complexed with (c-di-GMP)2 at the active site. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 997–1008. [Google Scholar] [CrossRef]
- Minasov, G.; Padavattan, S.; Shuvalova, L.; Brunzelle, J.S.; Miller, D.J.; Baslé, A.; Massa, C.; Collart, F.R.; Schirmer, T.; Anderson, W.F. Crystal structures of YkuI and its complex with second messenger cyclic di-GMP suggest catalytic mechanism of phosphodiester bond cleavage by EAL domains. J. Biol. Chem. 2009, 284, 13174–13184. [Google Scholar]
- Tchigvintsev, A.; Xu, X.; Singer, A.; Chang, C.; Brown, G.; Proudfoot, M.; Cui, H.; Flick, R.; Anderson, W.F.; Joachimiak, A.; et al. Structural insight into the mechanism of c-di-GMP hydrolysis by EAL domain phosphodiesterases. J. Mol. Biol. 2010, 402, 524–538. [Google Scholar]
- Ko, J.; Ryu, K.S.; Kim, H.; Shin, J.S.; Lee, J.O.; Cheong, C.; Choi, B.S. Structure of PP4397 reveals the molecular basis for different c-di-GMP binding modes by Pilz domain proteins. J. Mol. Biol. 2010, 398, 97–110. [Google Scholar] [CrossRef]
- Benach, J.; Swaminathan, S.S.; Tamayo, R.; Handelman, S.K.; Folta-Stogniew, E.; Ramos, J.E.; Forouhar, F.; Neely, H.; Seetharaman, J.; Camilli, A.; et al. The structural basis of cyclic diguanylate signal transduction by PilZ domains. EMBO J. 2007, 26, 5153–5166. [Google Scholar] [CrossRef]
- Habazettl, J.; Allan, M.G.; Jenal, U.; Grzesiek, S. Solution structure of the PilZ domain protein PA4608 complex with cyclic di-GMP identifies charge clustering as molecular readout. J. Biol. Chem. 2011, 286, 14304–14314. [Google Scholar] [CrossRef]
- Hickman, J.W.; Harwood, C.S. Identification of FleQ from Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol. Microbiol. 2008, 69, 376–389. [Google Scholar] [CrossRef]
- Baraquet, C.; Murakami, K.; Parsek, M.R.; Harwood, C.S. The FleQ protein from Pseudomonas aeruginosa functions as both a repressor and an activator to control gene expression from the pel operon promoter in response to c-di-GMP. Nucleic Acids Res. 2012, 40, 7207–7218. [Google Scholar] [CrossRef]
- Sudarsan, N.; Lee, E.R.; Weinberg, Z.; Moy, R.H.; Kim, J.N.; Link, K.H.; Breaker, R.R. Riboswitches in eubacteria sense the second messenger cyclic di-GMP. Science 2008, 321, 411–413. [Google Scholar] [CrossRef]
- Smith, K.D.; Shanahan, C.A.; Moore, E.L.; Simon, A.C.; Strobel, S.A. Structural basis of differential ligand recognition by two classes of bis-(3'-5')-cyclic dimeric guanosine monophosphate-binding riboswitches. Proc. Natl. Acad. Sci. USA 2011, 108, 7757–7762. [Google Scholar]
- Cheek, M.A.; Dobrikov, M.I.; Wennefors, C.K.; Xu, Z.; Hashmi, S.N.; Shen, X.; Shaw, B.R. Synthesis and properties of (alpha-P-borano)-nucleoside 5'-triphosphate analogues as potential antiviral agents. Nucleic Acids Symp. Ser. (Oxf.) 2008, 52, 81–82. [Google Scholar] [CrossRef]
- Wennefors, C.K.; Dobrikov, M.I.; Xu, Z.; Li, P.; Shaw, B.R. Stereospecificity, substrate, and inhibitory properties of nucleoside diphosphate analogs for creatine and pyruvate kinases. Bioorg. Chem. 2008, 36, 169–177. [Google Scholar] [CrossRef]
- Hamm, M.L.; Cholera, R.; Hoey, C.L.; Gill, T.J. Oligonucleotide incorporation of 8-thio-2'-deoxyguanosine. Org. Lett. 2004, 6, 3817–3820. [Google Scholar] [CrossRef]
- Hamm, M.L.; Gill, T.J.; Nicolson, S.C.; Summers, M.R. Substrate specificity of Fpg (MutM) and hOGG1, two repair glycosylases. J. Am. Chem. Soc. 2007, 129, 7724–7725. [Google Scholar] [CrossRef]
- Wauchope, O.R.; Johnson, C.; Krishnamoorthy, P.; Andrei, G.; Snoeck, R.; Balzarini, J.; Seley-Radtke, K.L. Synthesis and biological evaluation of a series of thieno-expanded tricyclic purine 2'-deoxy nucleoside analogues. Bioorg. Med. Chem. 2012, 20, 3009–3015. [Google Scholar] [CrossRef]
- Novikov, M.S.; Ivanova, O.N.; Ivanov, A.V.; Ozerov, A.A.; Valuev-Elliston, V.T.; Temburnikar, K.; Gurskaya, G.V.; Kochetkov, S.N.; Pannecouque, C.; Balzarini, J.; et al. 1-[2-(2-Benzoyl- and 2-benzylphenoxy)ethyl]uracils as potent anti-HIV-1 agents. Bioorg. Med. Chem. 2011, 19, 5794–5802. [Google Scholar]
- Cassera, M.B.; Zhang, Y.; Hazleton, K.Z.; Schramm, V.L. Purine and pyrimidine pathways as targets in Plasmodium falciparum. Curr. Top. Med. Chem. 2011, 11, 2103–2115. [Google Scholar] [CrossRef]
- Chemama, M.; Fonvielle, M.; Villet, R.; Arthur, M.; Valéry, J.M.; Etheve-Quelquejeu, M. Stable analogues of aminoacyl-tRNA for inhibition of an essential step of bacterial cell-wall synthesis. J. Am. Chem. Soc. 2007, 129, 12642–12643. [Google Scholar]
- Shukla, S.; Sumaria, C.S.; Pradeepkumar, P.I. Exploring chemical modifications for siRNA therapeutics: A structural and functional outlook. ChemMedChem 2010, 5, 328–349. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.; Donaldson, G.P.; Nakayama, S.; Yan, L.; Lam, Y.F.; Lee, V.T.; Sintim, H.O. Conservative change to the phosphate moiety of cyclic diguanylic monophosphate remarkably affects its polymorphism and ability to bind DGC, PDE, and PilZ proteins. J. Am. Chem. Soc. 2011, 133, 9320–9330. [Google Scholar]
- Shanahan, C.A.; Gaffney, B.L.; Jones, R.A.; Strobel, S.A. Differential analogue binding by two classes of c-di-GMP riboswitches. J. Am. Chem. Soc. 2011, 133, 15578–15592. [Google Scholar]
- Smith, K.D.; Strobel, S.A. Interactions of the c-di-GMP riboswitch with its second messenger ligand. Biochem. Soc. Trans. 2011, 39, 647–651. [Google Scholar] [CrossRef]
- Furukawa, K.; Gu, H.; Sudarsan, N.; Hayakawa, Y.; Hyodo, M.; Breaker, R.R. Identification of ligand analogues that control c-di-GMP riboswitches. ACS Chem. Biol. 2012, 7, 1436–1443. [Google Scholar] [CrossRef]
- Ching, S.M.; Tan, W.J.; Chua, K.L.; Lam, Y. Synthesis of cyclic di-nucleotidic acids as potential inhibitors targeting diguanylate cyclase. Bioorg. Med. Chem. 2010, 18, 6657–6665. [Google Scholar] [CrossRef]
- Sharma, I.M.; Dhanaraman, T.; Mathew, R.; Chatterji, D. Synthesis and characterization of a fluorescent analogue of cyclic di-GMP. Biochemistry 2012, 51, 5443–5453. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, J.; Watt, S.K.; Lee, V.T.; Dayie, T.K.; Sintim, H.O. Differential binding of 2'-biotinylated analogs of c-di-GMP with c-di-GMP riboswitches and binding proteins. Mol. Biosyst. 2012, 8, 772–778. [Google Scholar] [CrossRef]
- Smietana, M.; Kool, E.T. Efficient and simple solid-phase synthesis of short cyclic oligodeoxynucleotides bearing a phosphorothioate linkage. Angew. Chem. Int. Ed. Engl. 2002, 41, 3704–3707. [Google Scholar] [CrossRef]
- Chin, K.H.; Kuo, W.T.; Yu, Y.J.; Liao, Y.T.; Yang, M.T.; Chou, S.H. Structural polymorphism of c-di-GMP bound to an EAL domain and in complex with a type II PilZ-domain protein. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 1380–1392. [Google Scholar] [CrossRef]
- Zhang, Z.; Kim, S.; Gaffney, B.L.; Jones, R.A. Polymorphism of the signaling molecule c-di-GMP. J. Am. Chem. Soc. 2006, 128, 7015–7024. [Google Scholar] [CrossRef]
- Zhang, Z.; Gaffney, B.L.; Jones, R.A. C-di-GMP displays a monovalent metal ion-dependent polymorphism. J. Am. Chem. Soc. 2004, 126, 16700–16701. [Google Scholar] [CrossRef]
- Gaussian 09, Revision A02; Software for technical computation; Gaussian Inc.: Wallingford, CT, USA, 2009.
- Gentner, M.; Allan, M.G.; Zaehringer, F.; Schirmer, T.; Grzesiek, S. Oligomer formation of the bacterial second messenger c-di-GMP: Reaction rates and equilibrium constants indicate a monomeric state at physiological concentrations. J. Am. Chem. Soc. 2012, 134, 1019–1029. [Google Scholar]
- Kulshina, N.; Baird, N.J.; Ferré-D’Amaré, A.R. Recognition of the bacterial second messenger cyclic diguanylate by its cognate riboswitch. Nat. Struct. Mol. Biol. 2009, 16, 1212–1217. [Google Scholar]
- Smith, K.D.; Lipchock, S.V.; Ames, T.D.; Wang, J.; Breaker, R.R.; Strobel, S.A. Structural basis of ligand binding by a c-di-GMP riboswitch. Nat. Struct. Mol. Biol. 2009, 16, 1218–1223. [Google Scholar]
- Smith, K.D.; Lipchock, S.V.; Livingston, A.L.; Shanahan, C.A.; Strobel, S.A. Structural and biochemical determinants of ligand binding by the c-di-GMP riboswitch. Biochemistry 2010, 49, 7351–7359. [Google Scholar]
- Nakayama, S.; Luo, Y.; Zhou, J.; Dayie, T.K.; Sintim, H.O. Nanomolar fluorescent detection of c-di-GMP using a modular aptamer strategy. Chem. Commun. (Camb.) 2012, 48, 9059–9061. [Google Scholar]
- Patnaik, S.; Kumar, P.; Garg, B.S.; Gandhi, R.P.; Gupta, K.C. Photomodulation of PS-modified oligonucleotides containing azobenzene substituent at pre-selected positions in phosphate backbone. Bioorg. Med. Chem. 2007, 15, 7840–7849. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhou, J.; Sayre, D.A.; Wang, J.; Pahadi, N.; Sintim, H.O. Endo-S-c-di-GMP Analogues-Polymorphism and Binding Studies with Class I Riboswitch. Molecules 2012, 17, 13376-13389. https://doi.org/10.3390/molecules171113376
Zhou J, Sayre DA, Wang J, Pahadi N, Sintim HO. Endo-S-c-di-GMP Analogues-Polymorphism and Binding Studies with Class I Riboswitch. Molecules. 2012; 17(11):13376-13389. https://doi.org/10.3390/molecules171113376
Chicago/Turabian StyleZhou, Jie, David A. Sayre, Jingxin Wang, Nirmal Pahadi, and Herman O. Sintim. 2012. "Endo-S-c-di-GMP Analogues-Polymorphism and Binding Studies with Class I Riboswitch" Molecules 17, no. 11: 13376-13389. https://doi.org/10.3390/molecules171113376
APA StyleZhou, J., Sayre, D. A., Wang, J., Pahadi, N., & Sintim, H. O. (2012). Endo-S-c-di-GMP Analogues-Polymorphism and Binding Studies with Class I Riboswitch. Molecules, 17(11), 13376-13389. https://doi.org/10.3390/molecules171113376