Inhibitory Effect and Possible Mechanism of Action of Patchouli Alcohol against Influenza A (H2N2) Virus

Abstract
:1. Introduction
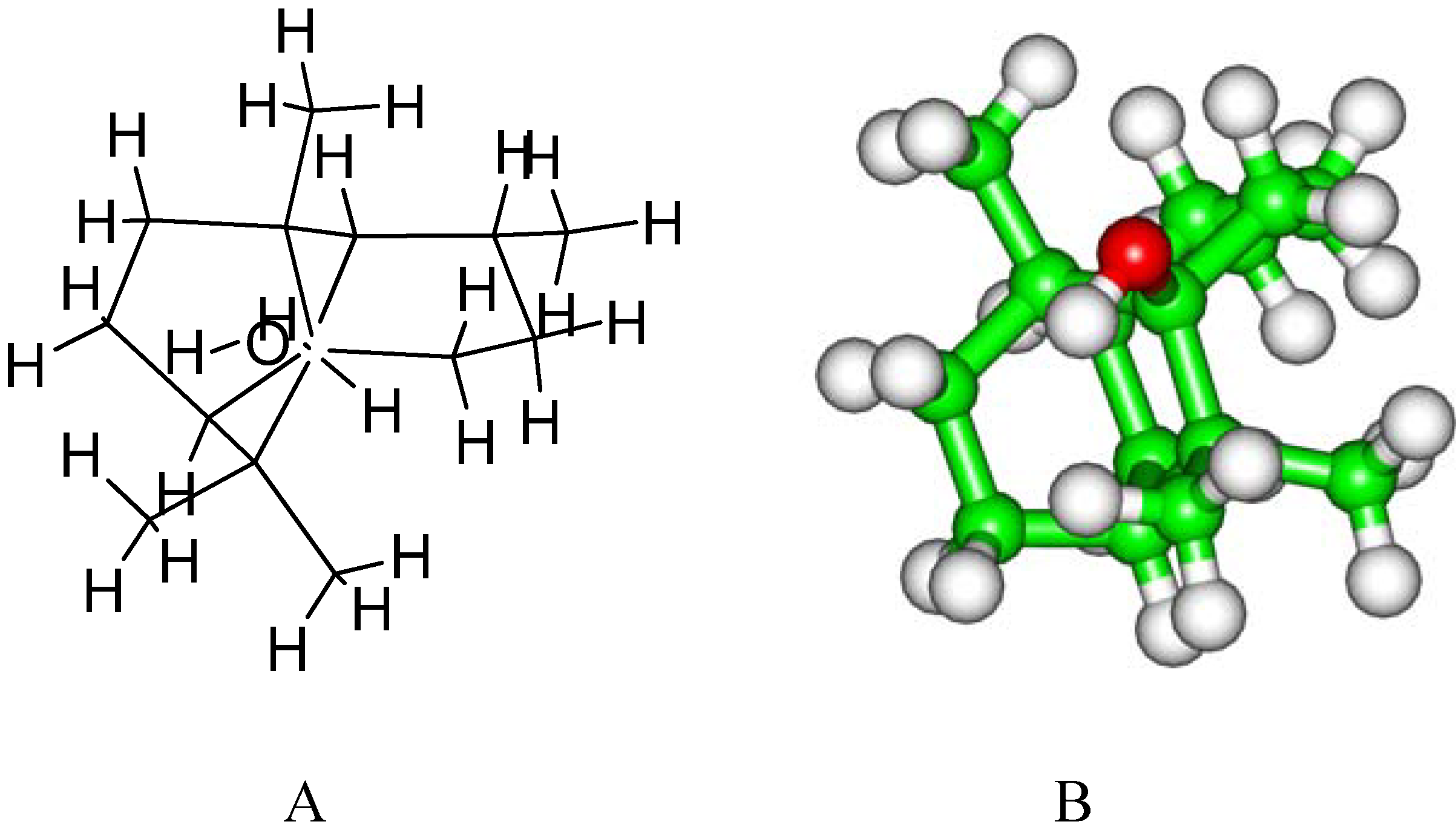
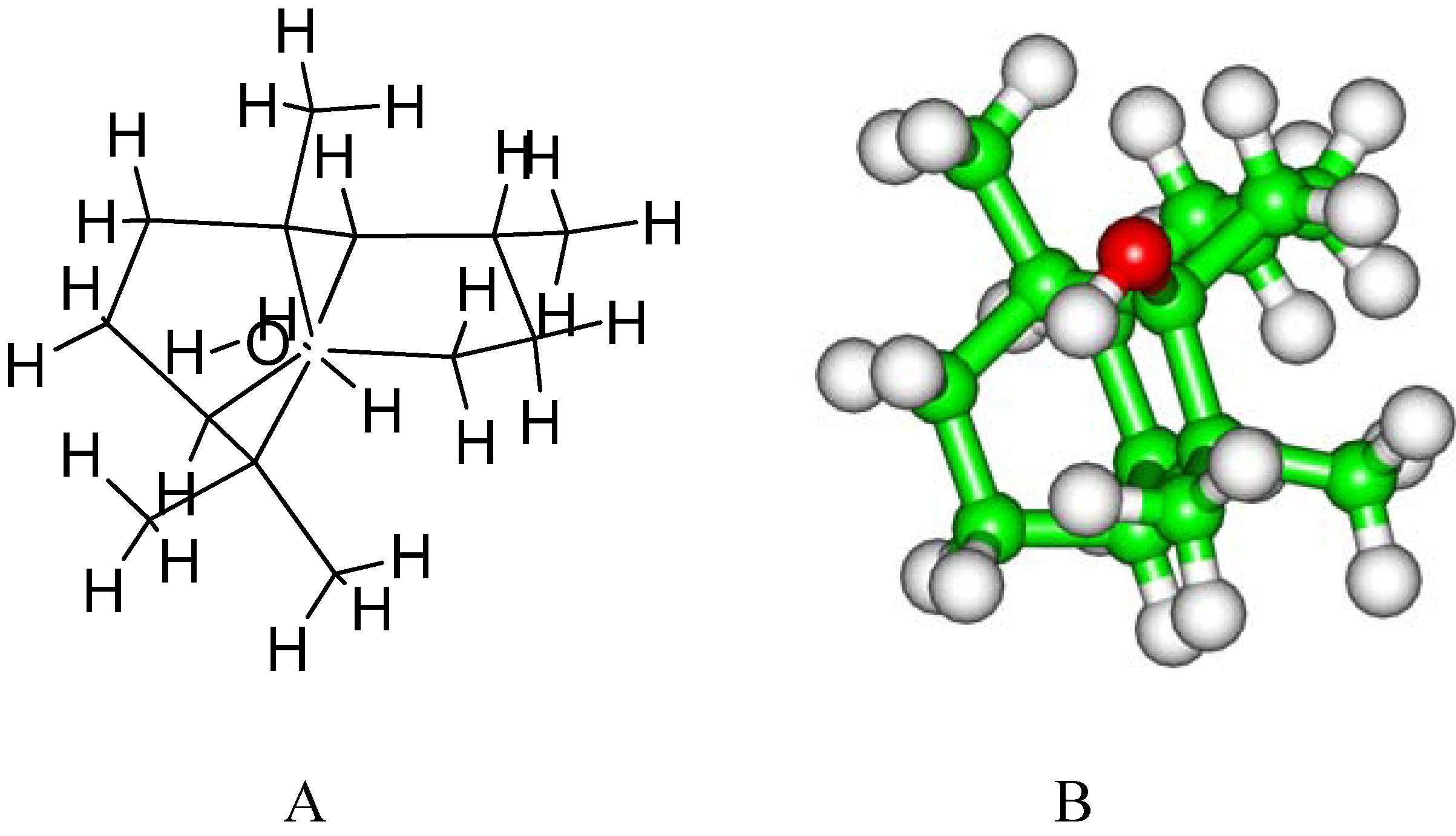
2. Results and Discussion
2.1. Effect of patchouli alcohol against influenza A (H2N2) virus by MTT assay in vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CC50a (µM) | Influenza A (A/Udorn/307/72) | |
|---|---|---|---|
| IC50b (µM) | SI c | ||
| Patchouli alcohol | >20.0 | 4.03 ± 0.23 * | >4.96 |
| Oseltamivir | >20.0 | 0.031 ± 0.012 | >645.16 |
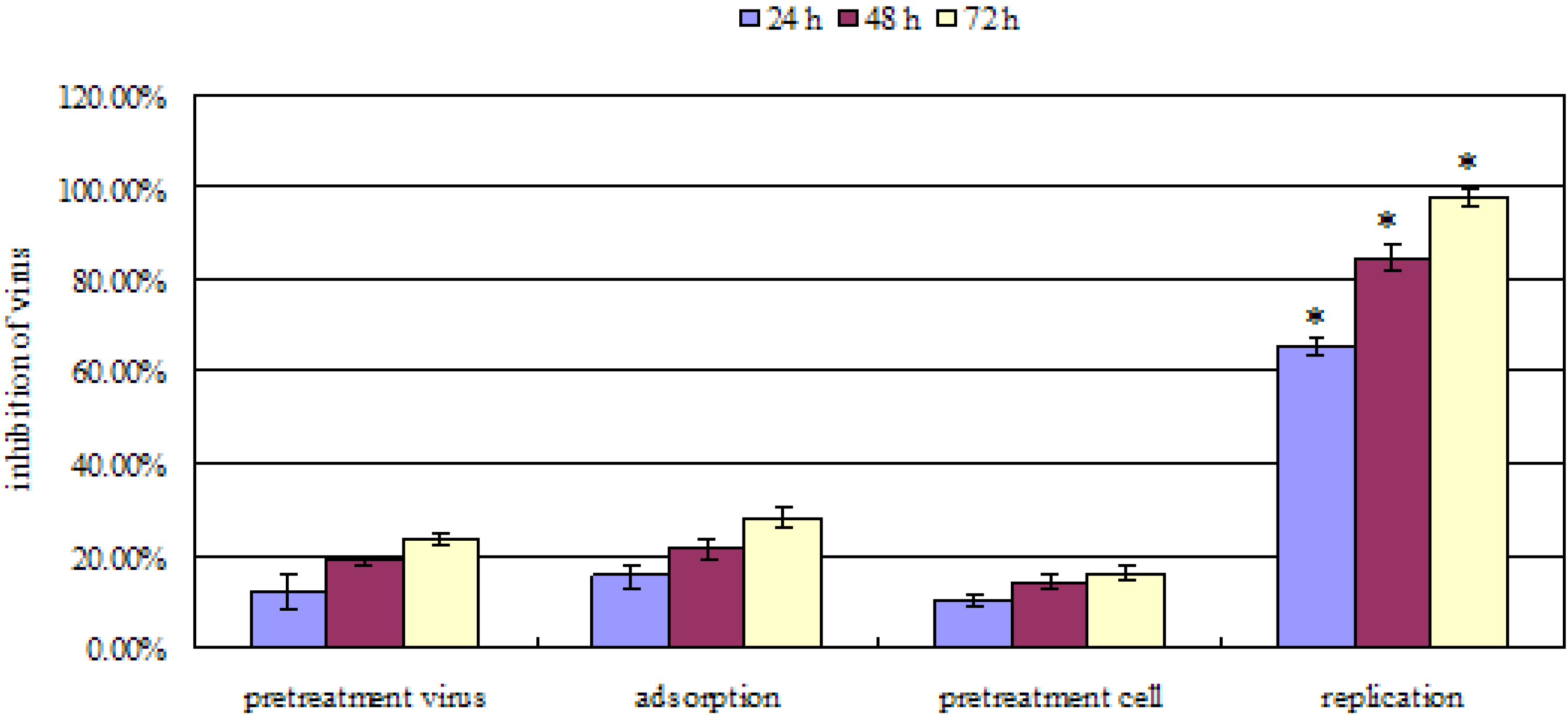
2.2. Mode of anti-influenza A (H2N2) activity
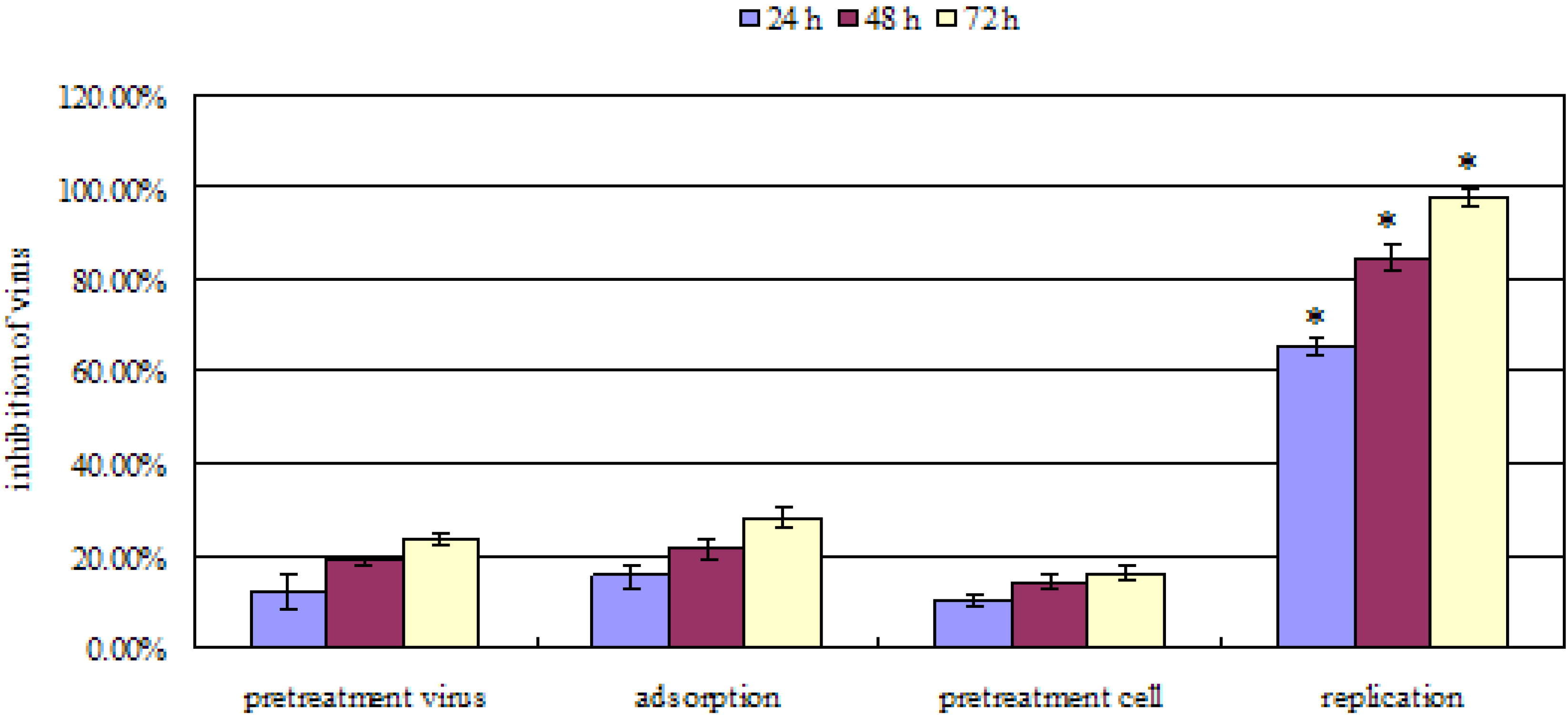
2.3. Anti-influenza A (H2N2) efficacy in mouse influenza model
| Group | Dose, mg/kg/day (q.d.) | Survivors/total | Mean day to death |
|---|---|---|---|
| Patchouli alcohol | 5 | 7/10 * | 11.8 ± 1.1 * |
| 1 | 2/10 * | 7.5 ± 1.8 * | |
| Oseltamivir | 1 | 5/10* | 9.1 ± 0.3 * |
| Control | - | 10/10 | - |
| H2N2 infected control | - | 0/10 | 5.2 ± 1.6 |
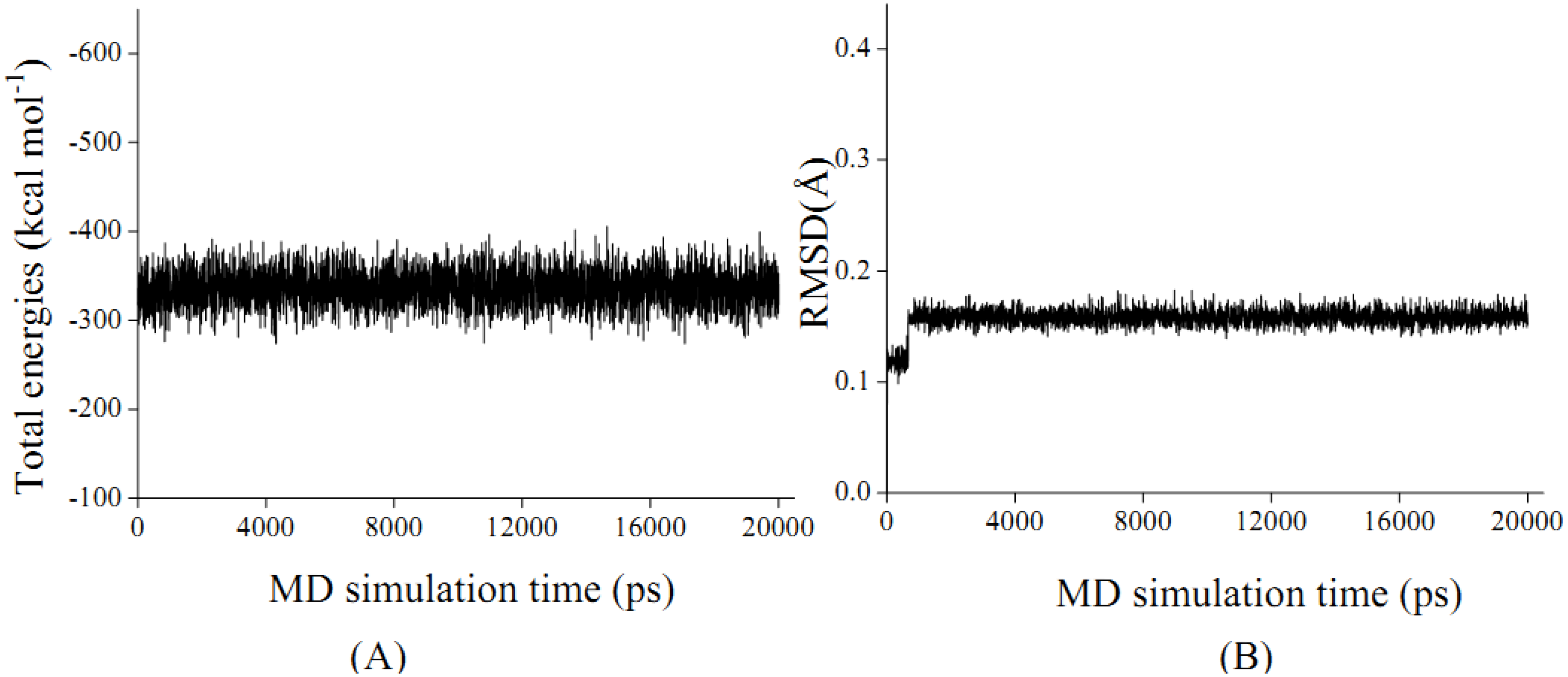
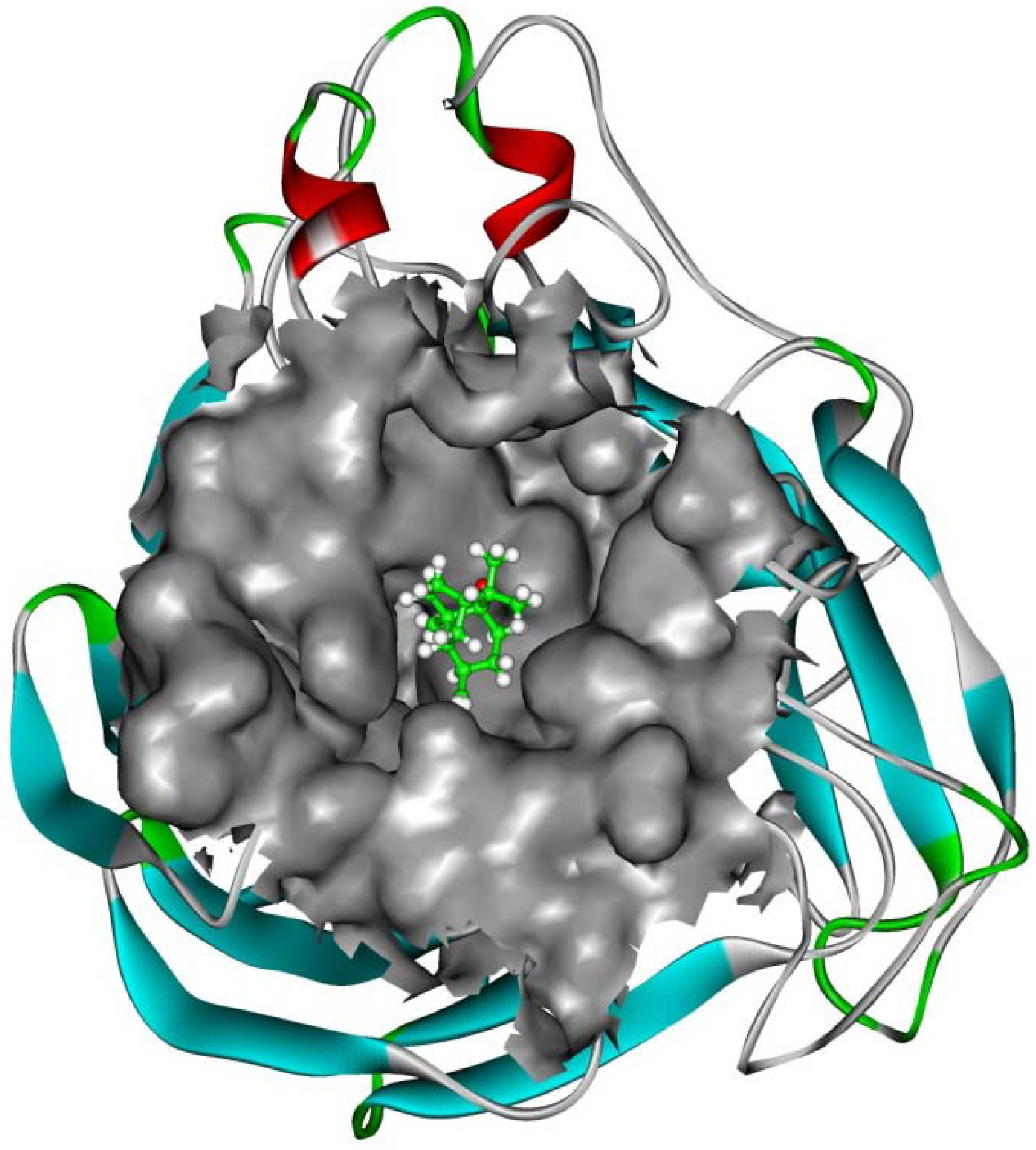
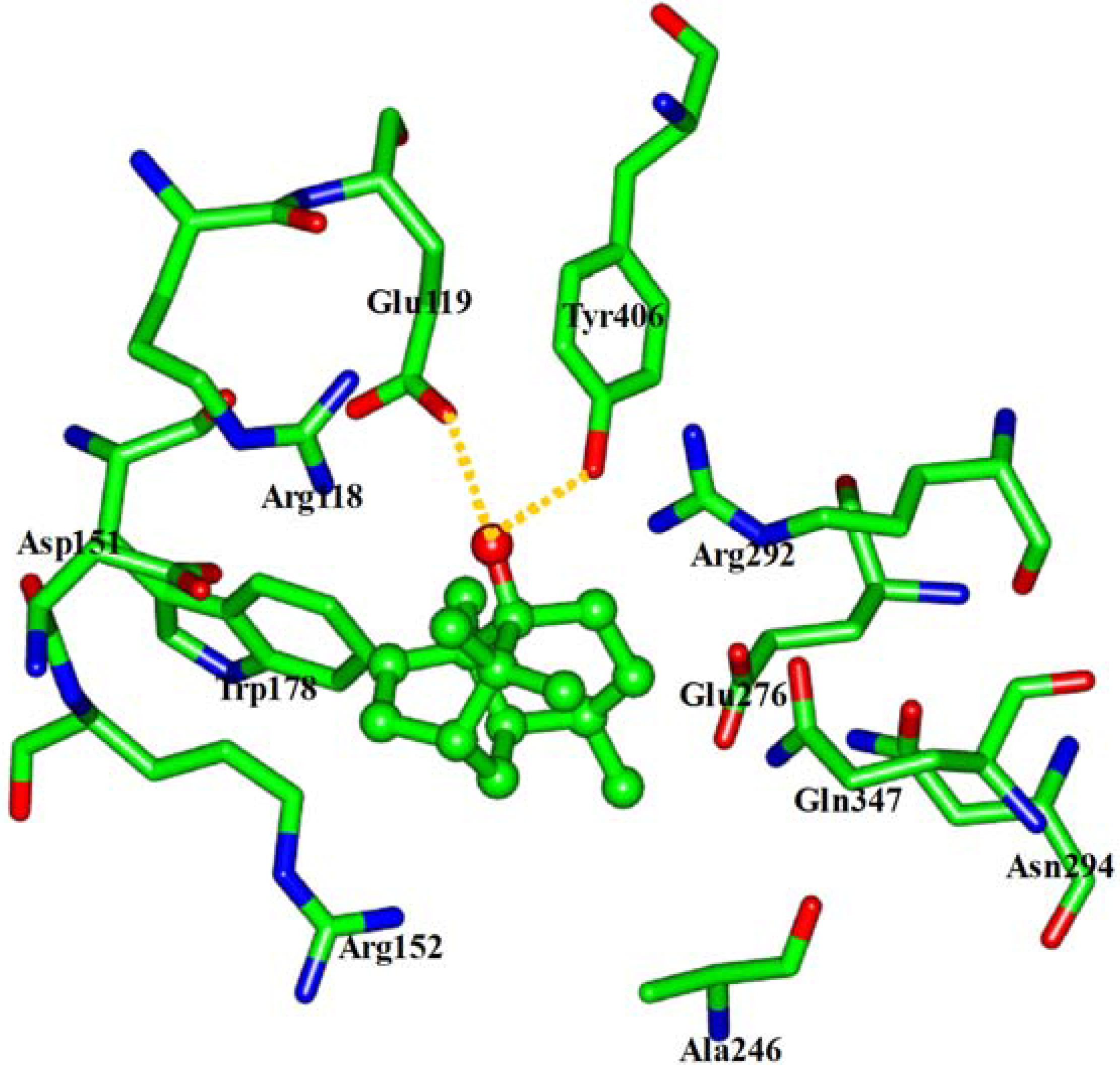
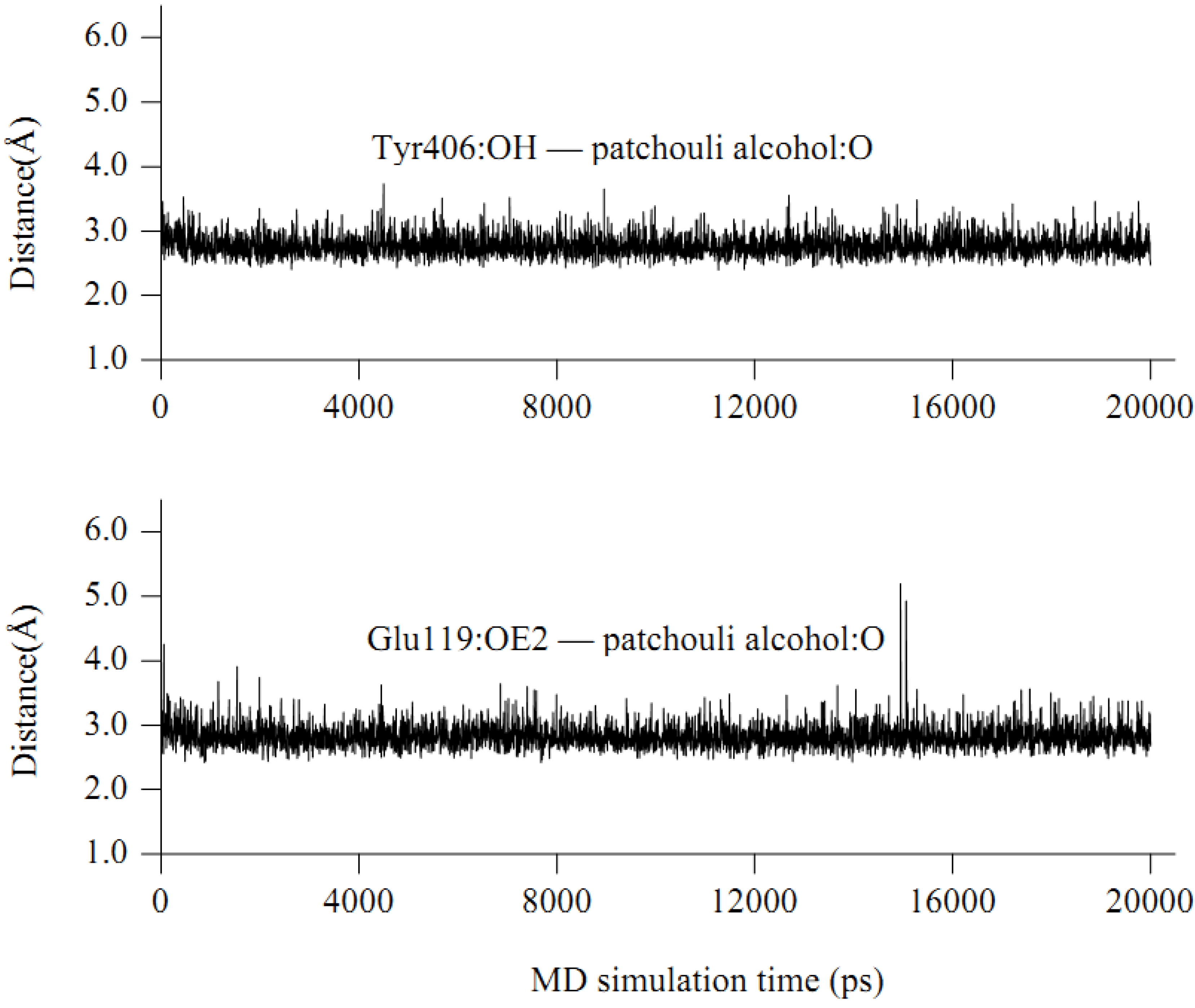
2.4. In silico inhibition mechanism of patchouli alcohol
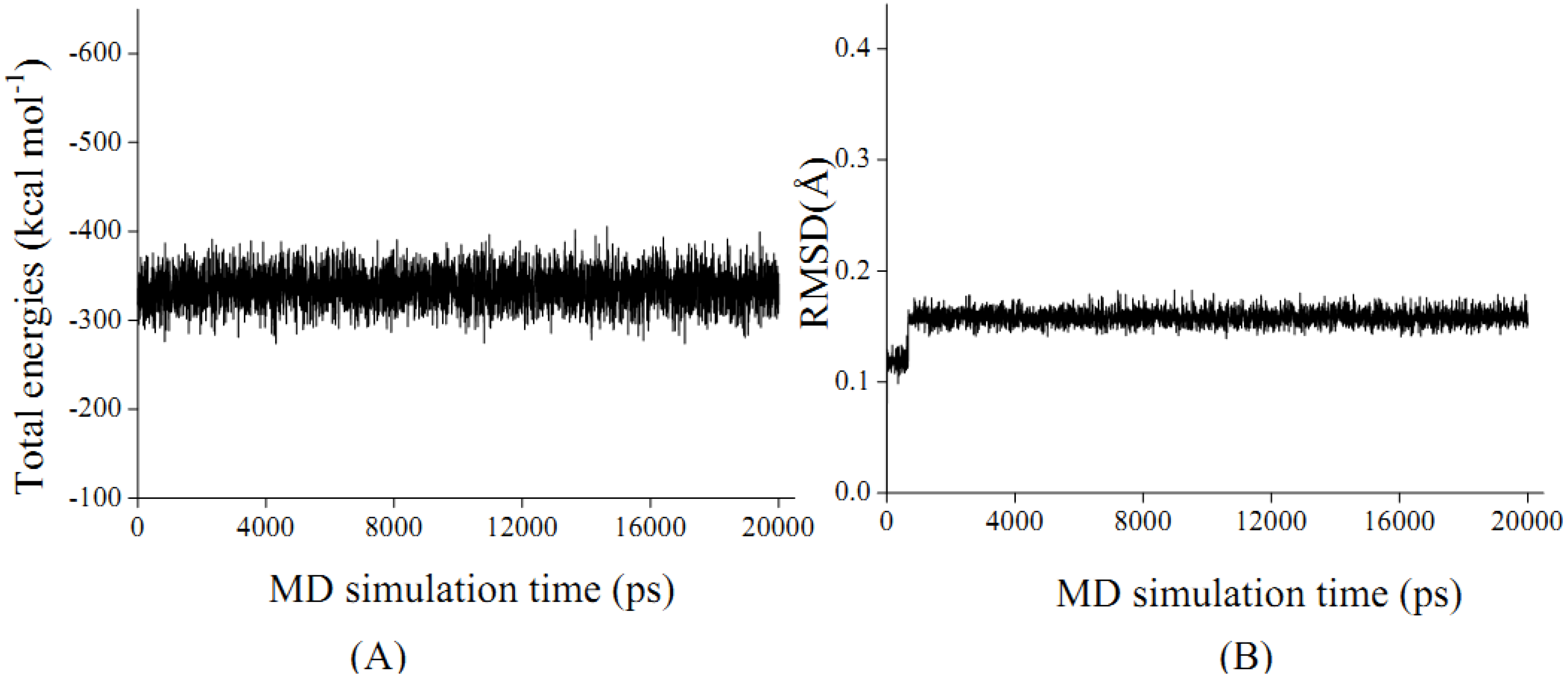
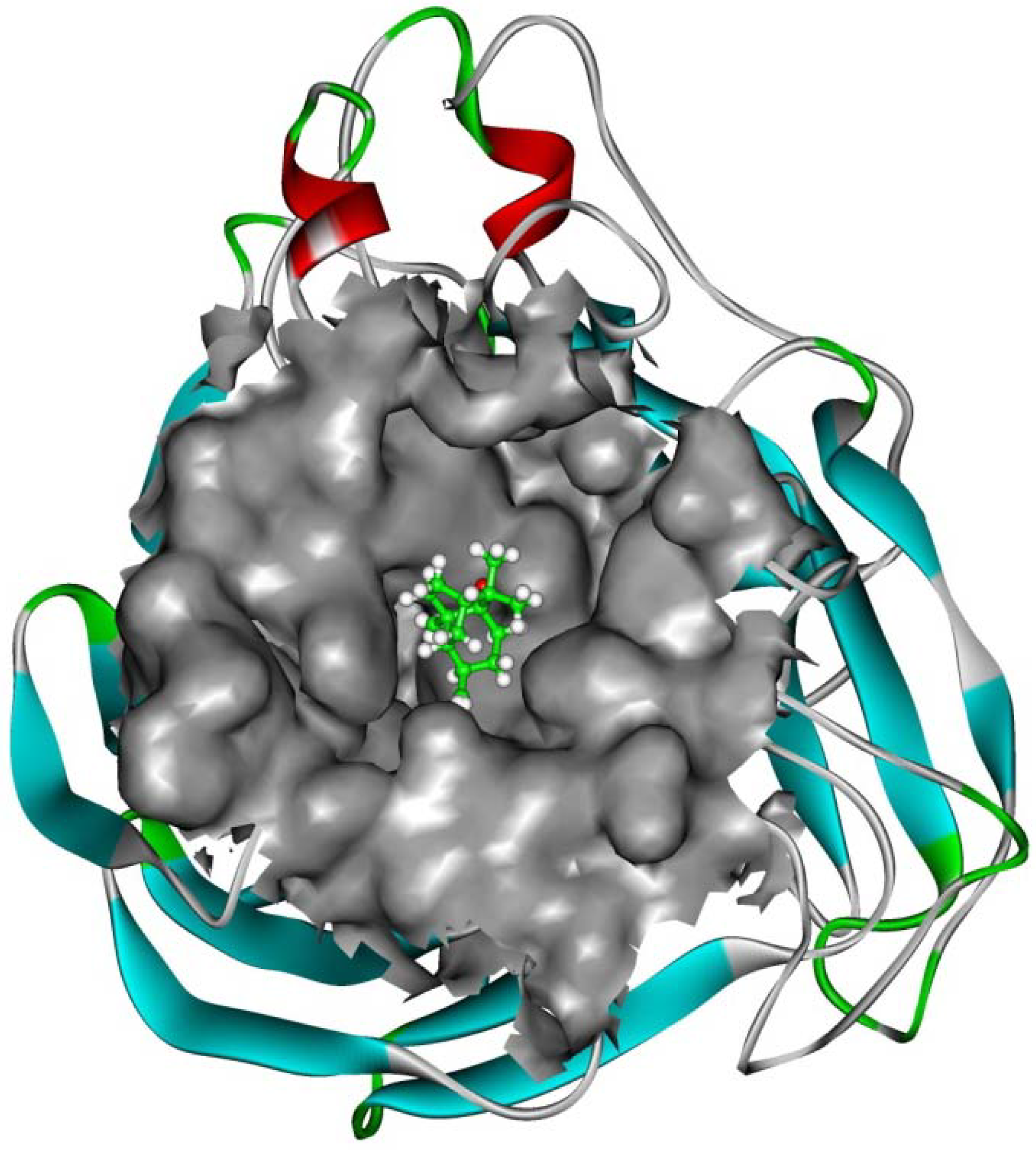
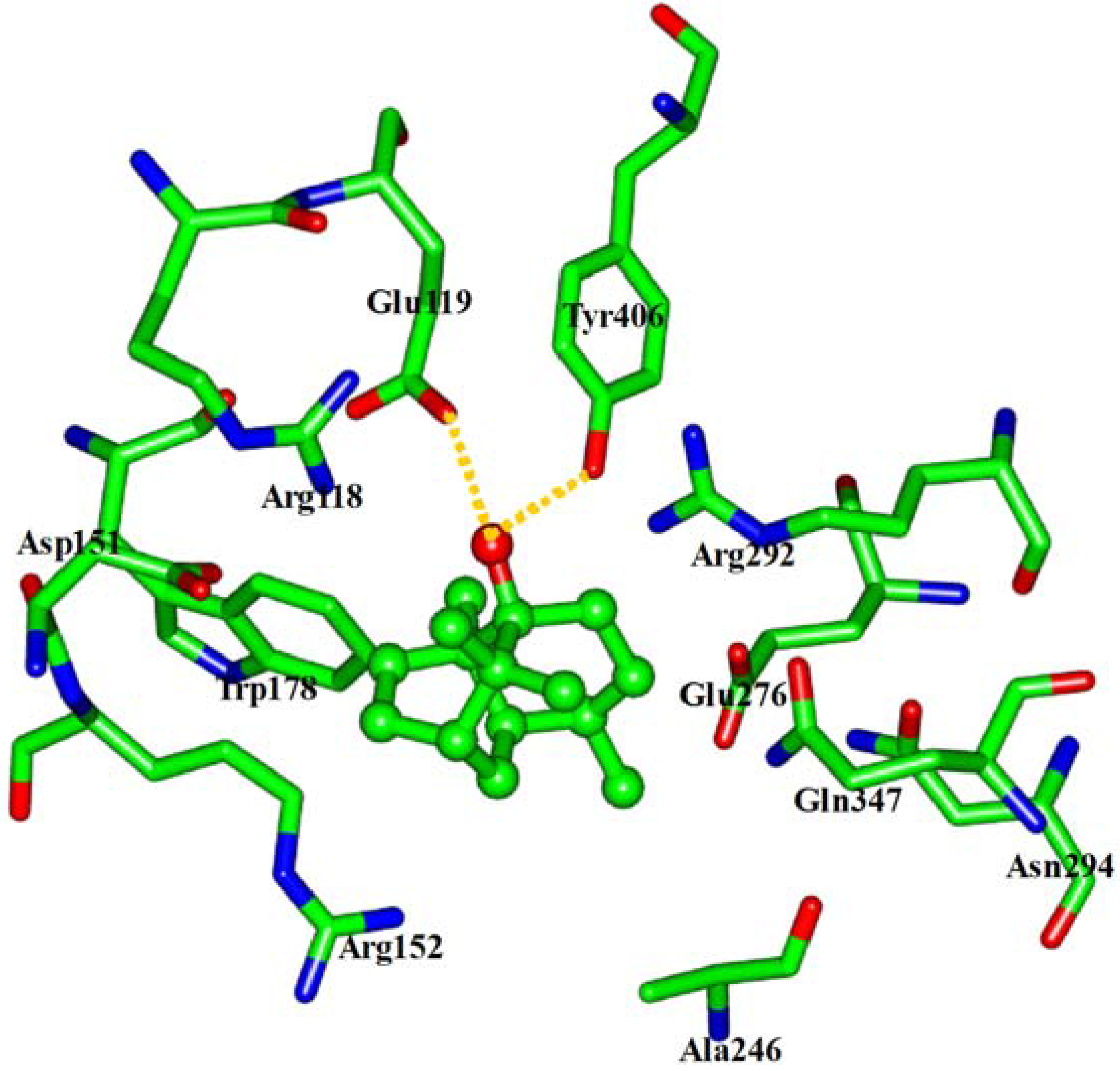
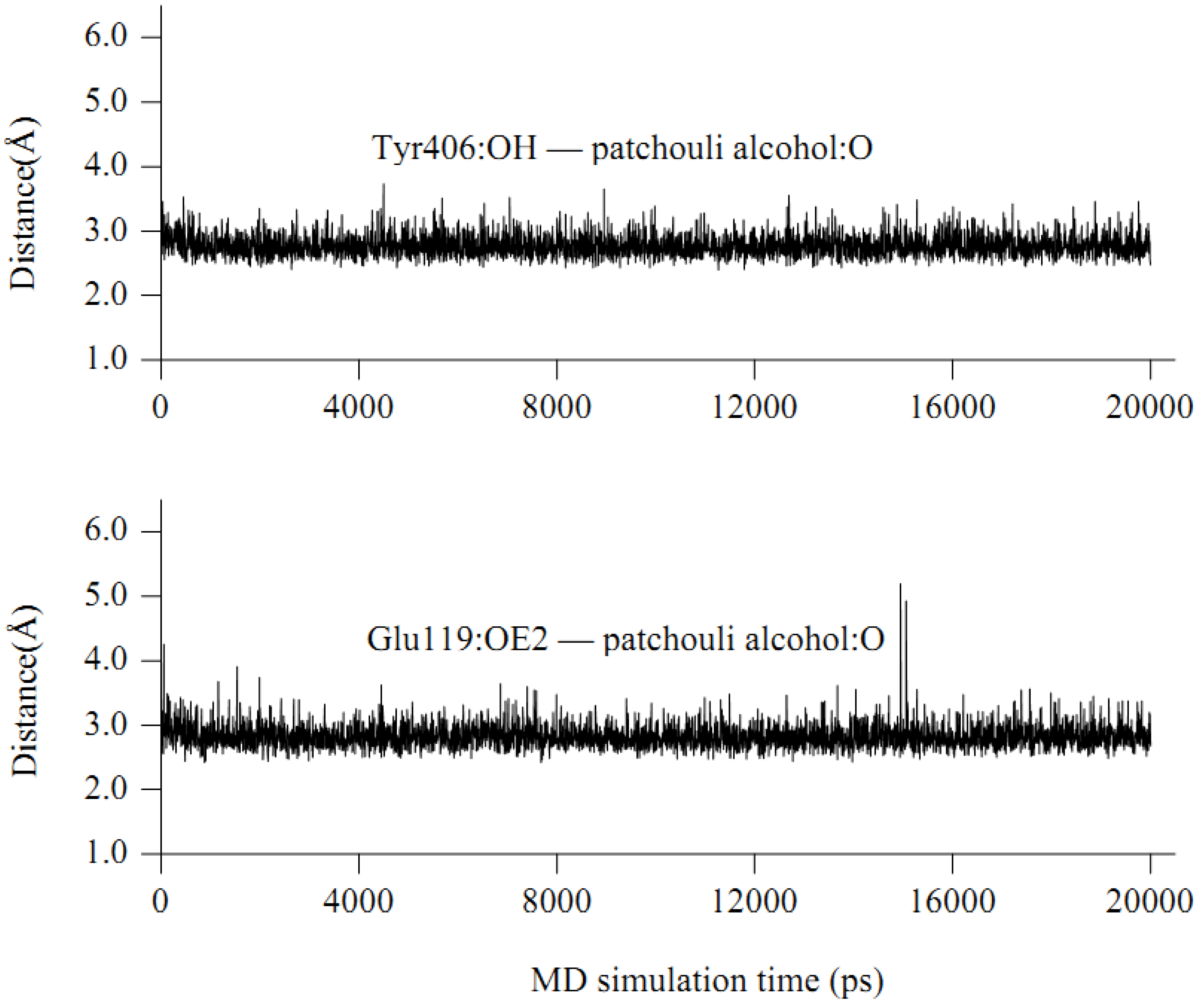
| Residue | E vdW | E ele | E sum |
|---|---|---|---|
| Arg118 | −2.03 | 0.63 | −1.40 |
| Glu119 | −2.18 | −6.28 | −8.46 |
| Asp151 | −2.08 | −0.53 | −2.61 |
| Arg152 | −3.01 | 0.46 | −2.55 |
| Trp178 | −1.83 | 0.01 | −1.82 |
| Ala246 | −1.47 | 0.12 | −1.35 |
| Glu276 | −2.04 | −0.28 | −2.32 |
| Arg292 | −3.51 | 1.10 | −2.41 |
| Asn294 | −0.92 | 0.16 | −0.76 |
| Gln347 | −1.91 | 0.07 | −1.84 |
| Tyr406 | −3.07 | −4.30 | −7.37 |
3. Experimental
3.1. Materials
3.2. Cell cultures
3.3. Viruses
3.4. Cytotoxicity assay
3.5. Anti-influenza A (H2N2) activity in vitro
3.6. Mode of anti-influenza A (H2N2) activity
3.7. Anti-influenza A (H2N2) efficacy in mouse influenza model
3.8. Computational methods
3.9. Statistical analysis
4. Conclusions
Acknowledgements
References
- Cox, N.J.; Subbarao, K. Global epidemiology of influenza: Past and present. Annu. Rev. Med. 2000, 51, 407–421. [Google Scholar] [CrossRef]
- Fauci, A.S. Emerging and Re-Emerging Infectious Diseases: Influenza as a Prototype of the Host-Pathogen Balancing Act. Cell 2006, 124, 665–670. [Google Scholar] [CrossRef]
- Layne, S.P.; Monto, A.S.; Taubenberger, J.K. Pandemic influenza: An inconvenient mutation. Science 2009, 323, 1560–1561. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Kaiser, L.; Hayden, F.G. Influenza virus neuraminidase inhibitors. Lancet 2000, 355, 827–835. [Google Scholar]
- Moscona, A. Neuraminidase inhibitors for influenza. N. Engl. J. Med. 2005, 353, 1363–1373. [Google Scholar] [CrossRef]
- De Clercq, E.; Neyts, J. Avian influenza A (H5N1) infection: Targets and strategies for chemotherapeutic intervention. Trends Pharmacol. Sci. 2007, 28, 280–285. [Google Scholar] [CrossRef]
- Garman, E.; Laver, G. Controlling influenza by inhibiting the virus's neuraminidase. Curr. Drug. Targets 2004, 5, 119–136. [Google Scholar] [CrossRef]
- von Itzstein, M.; Wu, W.Y.; Kok, G.B.; Pegg, M.S.; Dyason, J.C.; Jin, B.; Van Phan, T.; Smythe, M.L.; White, H.F.; Oliver, S.W. Rational design of potent sialidase-based inhibitors of influenza virus replication. Nature 1993, 363, 418–423. [Google Scholar]
- Kim, C.U.; Lew, W.; Williams, M.A.; Liu, H.; Zhang, L.; Swaminathan, S.; Bischofberger, N.; Chen, M.S.; Mendel, D.B.; Tai, C.Y. Influenza neuraminidase inhibitors possessing a novel hydrophobic interaction in the enzyme active site: Design, synthesis, and structural analysis of carbocyclic sialic acid analogues with potent anti-influenza activity. J. Am. Chem. Soc. 1997, 119, 681–690. [Google Scholar]
- Varghese, J.N.; Smith, P.W.; Sollis, S.L.; Blick, T.J.; Sahasrabudhe, A.; McKimm-Breschkin, J.L.; Colman, P.M. Drug design against a shifting target: A structural basis for resistance to inhibitors in a variant of influenza virus neuraminidase. Structure 1998, 6, 735–746. [Google Scholar] [CrossRef]
- Atigadda, V.R.; Brouillette, W.J.; Duarte, F.; Babu, Y.S.; Bantia, S.; Chand, P.; Chu, N.; Montgomery, J.A.; Walsh, D.A.; Sudbeck, E.; et al. Hydrophobic benzoic acids as inhibitors of influenza neuraminidase. Bioorg. Med. Chem. 1999, 7, 2487–2497. [Google Scholar] [CrossRef]
- Gubareva, L.V.; Kaiser, L.; Hayden, F.G. Influenza virus neuraminidase inhibitors. Lancet 2000, 355, 827–835. [Google Scholar] [CrossRef]
- Brouillette, W.J.; Bajpai, S.N.; Ali, S.M.; Velu, S.E.; Atigadda, V.R.; Lommer, B.S.; Finley, J.B.; Luo, M.; Air, G.M. Pyrrolidinobenzoic acid inhibitors of influenza virus neuraminidase: Modifications of essential pyrrolidinone ring substituents. Bioorg. Med. Chem. 2003, 11, 2739–2749. [Google Scholar]
- von Itzstein, M. The war against influenza: Discovery and development of sialidase inhibitors. Nat. Rev. Drug. Discov. 2007, 6, 967–974. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.; Fang, H.; Xu, W.; Liu, A.; Du, G. Design, synthesis, inhibitory activity, and SAR studies of hydrophobic p-aminosalicylic acid derivatives as neuraminidase inhibitors. Bioorg. Med. Chem. 2008, 16, 3839–3847. [Google Scholar] [CrossRef]
- Gubareva, L.V. Molecular mechanisms of influenza virus resistance to neuraminidase inhibitors. Virus Res. 2004, 103, 199–203. [Google Scholar] [CrossRef]
- Kiso, M.; Mitamura, K.; Sakai-Tagawa, Y.; Shiraishi, K.; Kawakami, C.; Kimura, K.; Hayden, F.G.; Sugaya, N.; Kawaoka, Y. Resistant influenza A viruses in children treated with oseltamivir: Descriptive study. Lancet 2004, 364, 759–765. [Google Scholar]
- Kiyohara, H.; Ichino, C.; Kawamura, Y.; Nagai, T.; Sato, N.; Yamada, H. Patchouli alcohol: In vitro direct anti-influenza virus sesquiterpene in Pogostemon cablin Benth. J. Nat. Med. 2011. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Shimizu, M.; Hiyama, Y.; Itoh, K.; Hashimoto, Y.; Nakayama, M.; Horie, T.; Morita, N. Antiviral activity of natural occurring flavonoids in vitro. Chem. Pharm. Bull. (Tokyo) 1985, 33, 3881–3886. [Google Scholar]
- Fan, H.; Ooi, A.; Tan, Y.W.; Wang, S.; Fang, S.; Liu, D.X.; Lescar, J. The nucleocapsid protein of coronavirus infectious bronchitis virus: Crystal structure of its N-terminal domain and multimerization properties. Structure 2005, 13, 1859–1868. [Google Scholar] [CrossRef]
- Tan, Y.W.; Fang, S.; Fan, H.; Lescar, J.; Liu, D.X. Amino acid residues critical for RNA-binding in the N-terminal domain of the nucleocapsid protein are essential determinants for the infectivity of coronavirus in cultured cells. Nucleic Acids Res. 2006, 34, 4816–4825. [Google Scholar]
- Jayaram, H.; Fan, H.; Bowman, B.R.; Ooi, A.; Jayaram, J.; Collisson, E.W.; Lescar, J.; Prasad, B.V. X-ray structures of the N- and C-terminal domains of a coronavirus nucleocapsid protein: implications for nucleocapsid formation. J. Virol. 2006, 80, 6612–6620. [Google Scholar] [CrossRef]
- Masukawa, K.M.; Kollman, P.A.; Kuntz, I.D. Investigation of Neuraminidase-Substrate Recognition Using Molecular Dynamics and Free Energy Calculations. J. Med. Chem. 2003, 46, 5628–5637. [Google Scholar]
- Bonnet, P.; Bryce, R.A. Molecular dynamics and free energy analysis of neuraminidase-ligand interactions. Protein Sci. 2004, 13, 946–957. [Google Scholar] [CrossRef]
- Bonnet, P.; Bryce, R.A. Scoring binding affinity of multiple ligands using implicit solvent and a single molecular dynamics trajectory: Application to Influenza neuraminidase. J. Mol. Graph. Model. 2005, 24, 147–156. [Google Scholar] [CrossRef]
- Yang, Z.W.; Yang, G.; Zu, Y.G.; Fu, Y.J.; Zhou, L.J. The conformational analysis and proton transfer of the neuraminidase inhibitors: A theoretical study. Phys. Chem. Chem. Phys. 2009, 11, 10035–10041. [Google Scholar]
- Yang, Z.; Yang, G.; Zu, Y.; Fu, Y.; Zhou, L. Computer-Based De Novo Designs of Tripeptides as Novel Neuraminidase Inhibitors. Int. J. Mol. Sci. 2010, 11, 4932–4951. [Google Scholar] [CrossRef]
- Yang, Z.; Nie, Y.; Yang, G.; Zu, Y.; Fu, Y.; Zhou, L. Synergistic effects in the designs of neuraminidase ligands: Analysis from docking and molecular dynamics studies. J. Theor. Biol. 2010, 267, 363–374. [Google Scholar] [CrossRef]
- Oakley, A.J.; Barrett, S.; Peat, T.S.; Newman, J.; Streltsov, V.A.; Waddington, L.; Saito, T.; Tashiro, M.; McKimm-Breschkin, J.L. Structural and functional basis of resistance to neuraminidase inhibitors of influenza B viruses. J. Med. Chem. 2010, 53, 6421–6431. [Google Scholar]
- Stoll, V.; Stewart, K.D.; Maring, C.J.; Muchmore, S.; Giranda, V.; Gu, Y.-G.Y.; Wang, G.; Chen, Y.; Sun, M.; Zhao, C.; et al. Influenza neuraminidase inhibitors: Structure-based design of a novel inhibitor series. Biochemistry 2003, 42, 718–727. [Google Scholar]
- Aruksakunwong, O.; Malaisree, M.; Decha, P.; Sompornpisut, P.; Parasuk, V.; Pianwanit, S.; Hannongbua, S. On the lower susceptibility of oseltamivir to influenza neuraminidase subtype N1 than those in N2 and N9. Biophys. J. 2007, 92, 798–807. [Google Scholar] [CrossRef]
- Colman, P.M.; Varghese, J.N.; Laver, W.G. Structure of the catalytic and antigenic sites in influenza virus neuraminidase. Nature 1983, 303, 41–44. [Google Scholar] [CrossRef]
- Colman, P.M. Influenza virus neuraminidase: Structure, antibodies, and inhibitors. Protein Sci. 1994, 3, 1687–1696. [Google Scholar] [CrossRef]
- Jedrzejas, M.J.; Singh, S.; Brouillette, W.J.; Laver, W.G.; Air, G.M.; Luo, M. Structures of Aromatic Inhibitors of Influenza Virus Neuraminidase. Biochemistry 1995, 34, 3144–3151. [Google Scholar] [CrossRef]
- InisghtII Version 2005, 2005, Accelrys Inc.: San Diego, CA, USA, 2005.
- Head, J.D.; Zerner, M.C. A Broyden—Fletcher—Goldfarb—Shanno optimization procedure for molecular geometries. Chem. Phys. lett. 1985, 122, 264–270. [Google Scholar]
- Yang, Z.W.; Zu, Y.G.; Wu, X.M.; Liu, C.B.; Yang, G. A computational investigation on the interaction mechanisms of neuraminidases and 3-(3-pentyloxy)benzoic acid. Acta Chim. Sin. 2010, 14, 1370–1378. [Google Scholar]
- Yang, Z.W.; Wu, N.; Fu, F.J.; Yang, G.; Wang, W.; Zu, Y.G.; Efferth, T. Anti-infectious bronchitis virus (IBV) activity of 1,8-cineole: Effect on nucleocapsid (N) protein. J. Biomol. Struct. Dyn. 2010, 28, 323–330. [Google Scholar] [CrossRef]
- Affinity User Guide, Accelrys Inc.: San Diego, CA, USA, 2005.
- Adelman, S.A.; Doll, J.D. Generalized Langevin equation approach for atom/solid-surface scattering: General formulation for classical scattering off harmonic solids. J. Chem. Phys. 1976, 64, 2375–2388. [Google Scholar] [CrossRef]
- Sample Availability: Samples of patchouli alcohol are available on request from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, H.; Li, B.; Wang, X.; Jin, M.; Wang, G. Inhibitory Effect and Possible Mechanism of Action of Patchouli Alcohol against Influenza A (H2N2) Virus. Molecules 2011, 16, 6489-6501. https://doi.org/10.3390/molecules16086489
Wu H, Li B, Wang X, Jin M, Wang G. Inhibitory Effect and Possible Mechanism of Action of Patchouli Alcohol against Influenza A (H2N2) Virus. Molecules. 2011; 16(8):6489-6501. https://doi.org/10.3390/molecules16086489
Chicago/Turabian StyleWu, Huaxing, Beili Li, Xue Wang, Mingyuan Jin, and Guonian Wang. 2011. "Inhibitory Effect and Possible Mechanism of Action of Patchouli Alcohol against Influenza A (H2N2) Virus" Molecules 16, no. 8: 6489-6501. https://doi.org/10.3390/molecules16086489
APA StyleWu, H., Li, B., Wang, X., Jin, M., & Wang, G. (2011). Inhibitory Effect and Possible Mechanism of Action of Patchouli Alcohol against Influenza A (H2N2) Virus. Molecules, 16(8), 6489-6501. https://doi.org/10.3390/molecules16086489