Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mechanisms of Astringency
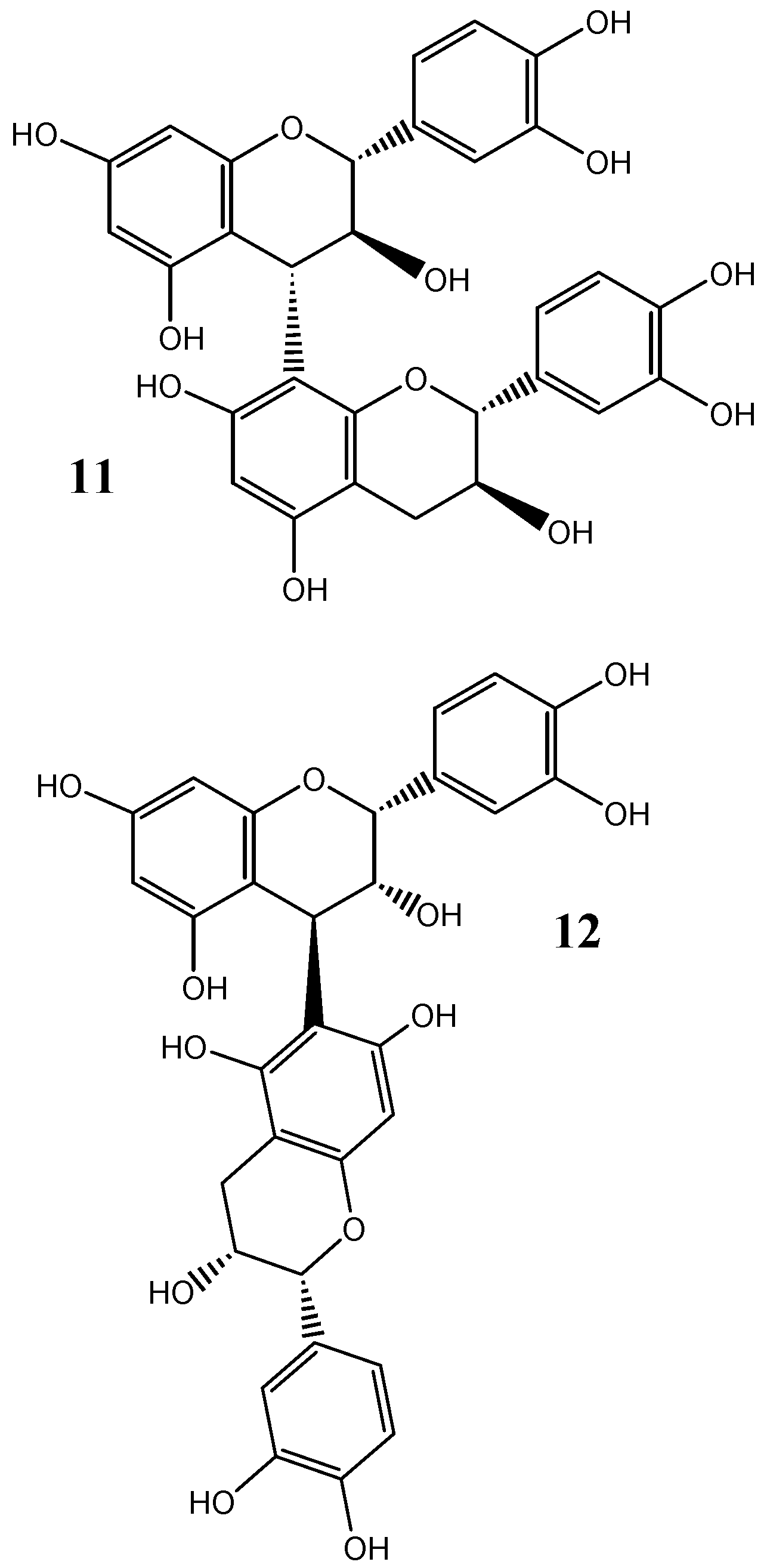
3. Red Wine Tannins
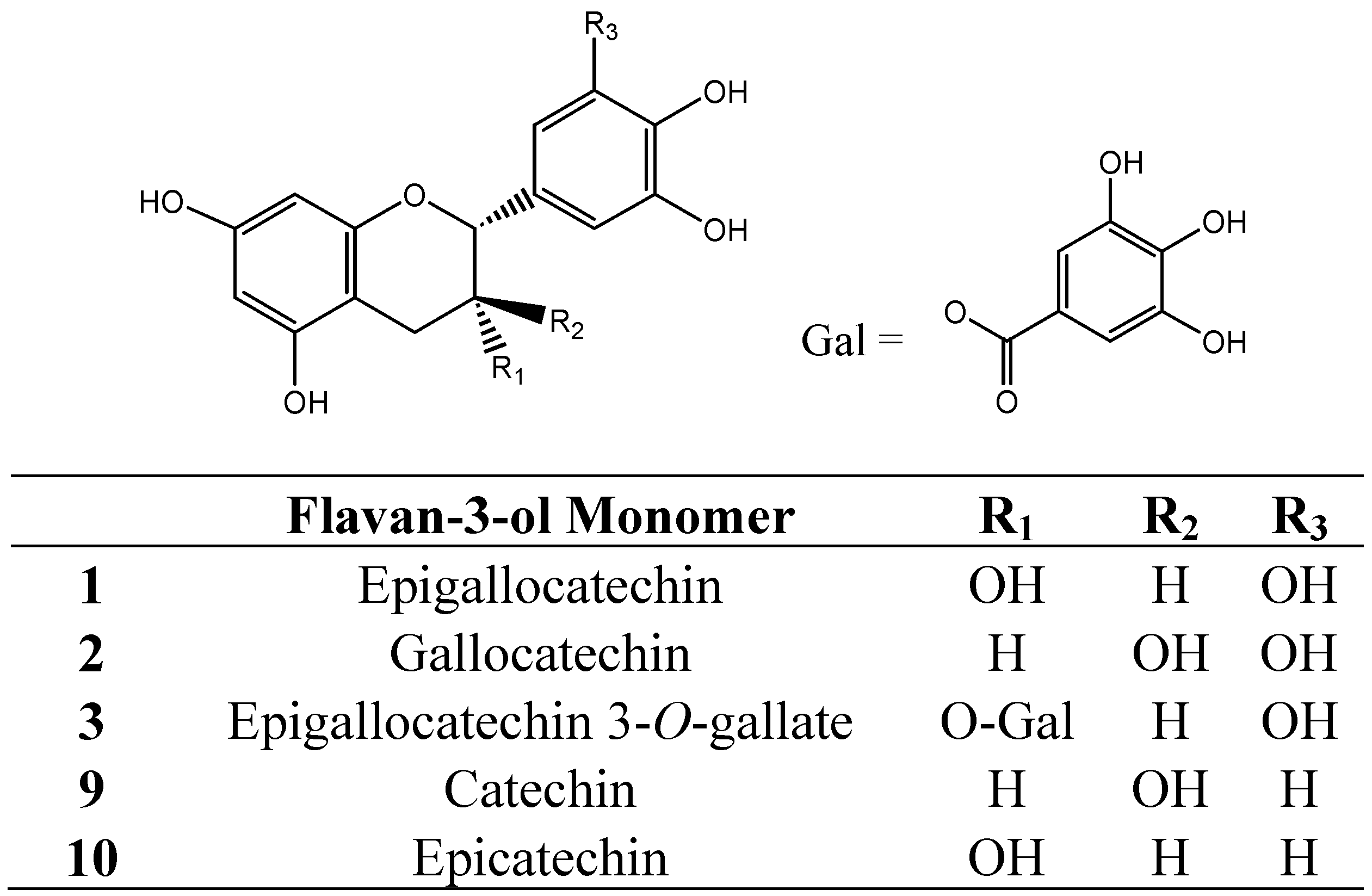
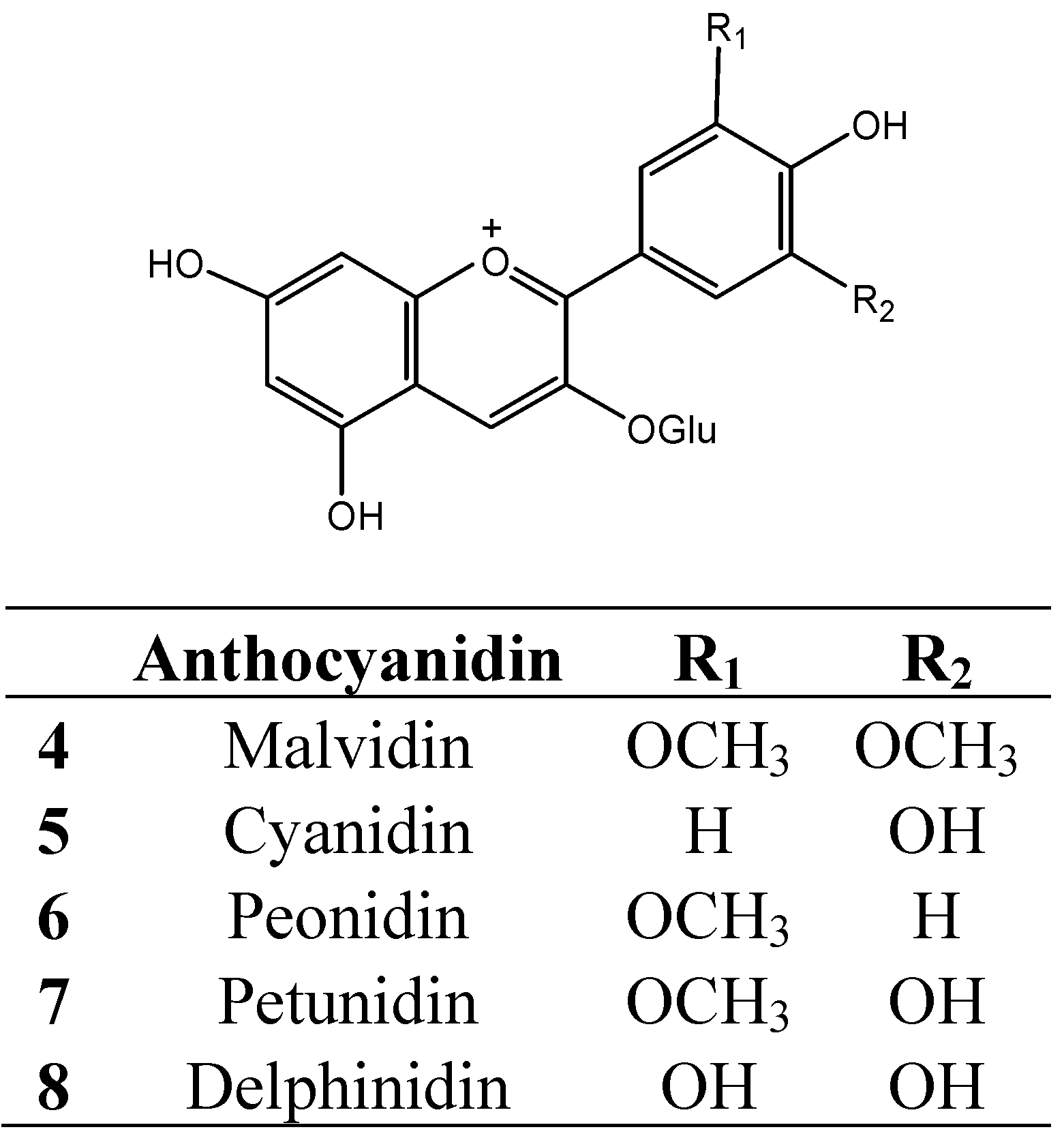
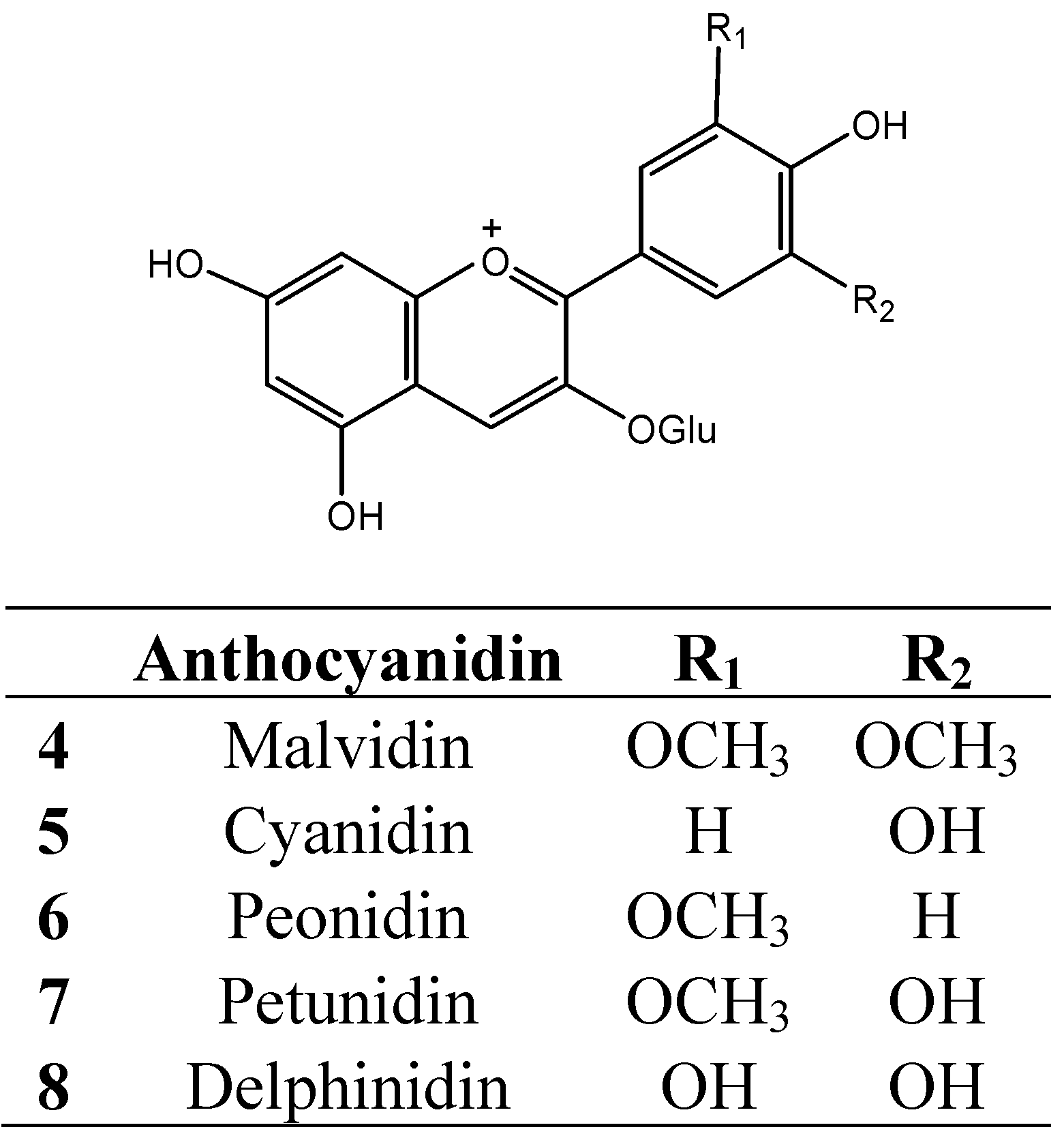
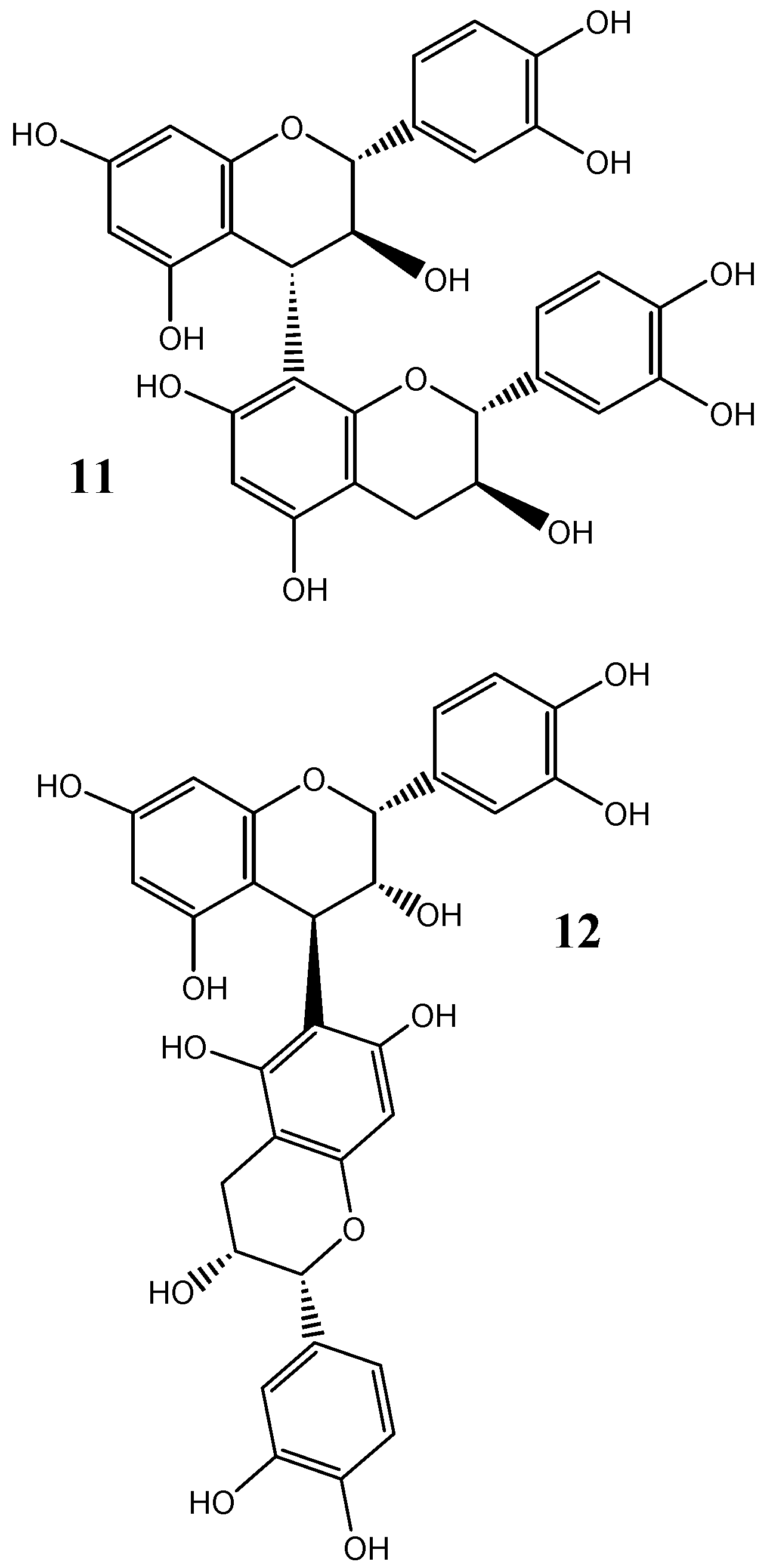
4. Protein-Tannin Interactions and Astringency
5. The Impact of the Wine Matrix on Astringency
6. Conclusions
Acknowledgements
References
- Lee, C.B.; Lawless, H.T. Time-course of astringent sensations. Chem. Senses 1991, 16, 225–238. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and wine phenolics: History and perspective. Am. J. Enol. Viticult. 2006, 57, 239–248. [Google Scholar]
- Mercurio, M.D.; Smith, P.A. Tannin quantification in red grapes and wine: Comparison of polysaccharide- and protein-based tannin precipitation techniques and their ability to model wine astringency. J. Agric. Food Chem. 2008, 56, 5528–5537. [Google Scholar] [CrossRef]
- Mercurio, M.D.; Dambergs, R.G.; Cozzolino, D.; Herderich, M.J.; Smith, P.A. Relationship between red wine grades and phenolics. 1. Tannin and total phenolics concentrations. J. Agric. Food Chem. 2010, 58, 12313–12319. [Google Scholar]
- Gawel, R.; Iland, P.G.; Francis, I.L. Characterizing the astringency of red wine: A case study. Food Qual. Prefer. 2001, 12, 83–94. [Google Scholar] [CrossRef]
- Kallithraka, S.; Bakker, J.; Clifford, M.N.; Vallis, L. Correlations between saliva protein composition and some T-I parameters of astringency. Food Qual. Prefer. 2001, 12, 145–152. [Google Scholar] [CrossRef]
- Kallithraka, S.; Bakker, J.; Clifford, M.N. Evaluation of bitterness and astringency of (+)-catechin and (-)-epicatechin in red wine and in model solution. J. Sens. Stud. 1997, 12, 25–37. [Google Scholar] [CrossRef]
- Demiglio, P.; Pickering, G.J. The influence of ethanol and pH on the taste and mouthfeel sensations elicited by red wine. J. Food Agric. Environ. 2008, 6, 143–150. [Google Scholar]
- Gawel, R.; Oberholster, A.; Francis, I.L. A 'mouth-feel wheel': terminology for communicating the mouth-feel characteristics of red wine. Aust. J. Grape Wine Res. 2000, 6, 203–207. [Google Scholar] [CrossRef]
- Kennedy, J.A. Grape and wine phenolics: Observations and recent findings. Cienc. Invest. Agrar. 2008, 35, 107–120. [Google Scholar]
- Boselli, E.; Boulton, R.B.; Thorngate, J.H.; Frega, N.G. Chemical and sensory characterization of doc red wines from Marche (Italy) related to vintage and grape cultivars. J. Agric. Food Chem. 2004, 52, 3843–3854. [Google Scholar] [CrossRef]
- Landon, J.L.; Weller, K.; Harbertson, J.F.; Ross, C.F. Chemical and sensory evaluation of astringency in Washington state red wines. Am. J. Enol. Viticult. 2008, 59, 153–158. [Google Scholar]
- Charlton, A.; Baxter, N.; Khan, M.; Moir, A.; Haslam, E.; Davies, A.; Williamson, M. Polyphenol/peptide binding and precipitation. J. Agric. Food Chem. 2002, 50, 1593–1601. [Google Scholar]
- Dinnella, C.; Recchia, A.; Fia, G.; Bertuccioli, M.; Monteleone, E. Saliva characteristics and individual sensitivity to phenolic astringent stimuli. Chem. Senses 2009, 34, 295–304. [Google Scholar] [CrossRef]
- Haslam, E. Natural polyphenols (vegetable tannins) as drugs: Possible modes of action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- Baxter, N.J.; Lilley, T.H.; Haslam, E.; Williamson, M.P. Multiple interactions between polyphenols and a salivary proline-rich protein repeat result in complexation and precipitation. Biochemistry 1997, 36, 5566–5577. [Google Scholar] [CrossRef]
- Hufnagel, J.C.; Hofmann, T. Orosensory-directed identification of astringent mouthfeel and bitter-tasting compounds in red wine. J. Agric. Food Chem. 2008, 56, 1376–1386. [Google Scholar] [CrossRef]
- Gawel, R. Red wine astringency: A review. Aust. J. Grape Wine Res. 1998, 4, 74–95. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Ferrier, J.; Harbertson, J.F.; Gachons, C.P.D. Analysis of tannins in red wine using multiple methods: Correlation with perceived astringency. Am. J. Enol. Viticult. 2006, 57, 481–485. [Google Scholar]
- Vidal, S.; Francis, L.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, W.; Waters, E. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Carvalho, E.; Mateus, N.; Plet, B.; Pianet, I.; Dufourc, E.; de Freitas, V. Influence of wine pectic polysaccharides on the interactions between condensed tannins and salivary proteins. J. Agric. Food Chem. 2006, 54, 8936–8944. [Google Scholar] [CrossRef]
- Gawel, R.; Francis, L.; Waters, E.J. Statistical correlations between the in-mouth textural characteristics and the chemical composition of Shiraz wines. J. Agric. Food Chem. 2007, 55, 2683–2687. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Noble, A.; Kwiatkowski, M.; Cheynier, V.; Waters, E. Taste and mouth-feel properties of different types of tannin-like polyphenolic compounds and anthocyanins in wine. Anal. Chim. Acta 2004, 513, 57–65. [Google Scholar] [CrossRef]
- Fontoin, H.; Saucier, C.; Teissedre, P.-L.; Glories, Y. Effect of pH, ethanol and acidity on astringency and bitterness of grape seed tannin oligomers in model wine solution. Food Qual. Prefer. 2008, 19, 286–291. [Google Scholar] [CrossRef]
- Kallithraka, S.; Bakker, J.; Clifford, M.N. Red wine and model wine astringency as affected by malic and lactic acid. J. Food Sci. 1997, 62, 416–420. [Google Scholar] [CrossRef]
- Rossetti, D.; Bongaerts, J.H.H.; Wantling, E.; Stokes, J.R.; Williamson, A.M. Astringency of tea catechins: More than an oral lubrication tactile percept. Food Hydrocolloid. 2009, 23, 1984–1992. [Google Scholar] [CrossRef]
- de Wijk, R.A.; Prinz, J.F. The role of friction in perceived oral texture. Food Qual. Prefer. 2005, 16, 121–129. [Google Scholar] [CrossRef]
- Payne, C.; Bowyer, P.K.; Herderich, M.; Bastian, S.E.P. Interaction of astringent grape seed procyanidins with oral epithelial cells. Food Chem. 2009, 115, 551–557. [Google Scholar] [CrossRef]
- Brossaud, F.; Cheynier, V.; Noble, A.C. Bitterness and astringency of grape and wine polyphenols. Aust. J. Grape Wine Res. 2001, 7, 33–39. [Google Scholar] [CrossRef]
- Smith, A.K.; June, H.; Noble, A.C. Effects of viscosity on the bitterness and astringency of grape seed tannin. Food Qual. Prefer. 1996, 7, 161–166. [Google Scholar] [CrossRef]
- Bajec, M.R.; Pickering, G. Astringency: Mechanisms and perception. Crit. Rev. Food Sci. Nutr. 2008, 48, 858–875. [Google Scholar] [CrossRef]
- Luck, G.; Liao, H.; Murray, N.J.; Grimmer, H.R.; Warminski, E.E.; Williamson, M.P.; Lilley, T.H.; Haslam, E. Polyphenols, astringency and proline-rich proteins. Phytochemistry 1994, 37, 357–371. [Google Scholar]
- Zanchi, D.; Poulain, C.; Konarev, P.; Tribet, C.; Svergun, D.I. Colloidal stability of tannins: astringency, wine tasting and beyond. J. Phys. Condens. Matter 2008, 20, 494224. [Google Scholar]
- Obreque-Slier, E.; López-Solís, R.; Peña-Neira, A.; Zamora-Marín, F. Tannin-protein interaction is more closely associated with astringency than tannin-protein precipitation: experience with two oenological tannins and a gelatin. Int. J. Food Sci. Technol. 2010, 45, 2629–2636. [Google Scholar] [CrossRef]
- Cheynier, V.; Duenas-Paton, M.; Salas, E.; Maury, C.; Souquet, J.M.; Sarni-Manchado, P.; Fulcrand, H. Structure and properties of wine pigments and tannins. Am. J. Enol. Viticult. 2006, 57, 298–305. [Google Scholar]
- Stark, T.; Bareuther, S.; Hofmann, T. Sensory-guided decomposition of roasted cocoa nibs (Theobroma cacao) and structure determination of taste-active polyphenols. J. Agric. Food Chem. 2005, 53, 5407–5418. [Google Scholar] [CrossRef]
- Condelli, N.; Dinnella, C.; Cerone, A.; Monteleone, E.; Bertuccioli, M. Prediction of perceived astringency induced by phenolic compounds II: Criteria for panel selection and preliminary application on wine samples. Food Qual. Prefer. 2006, 17, 96–107. [Google Scholar] [CrossRef]
- de Freitas, V.; Mateus, N. Structural features of procyanidin interactions with salivary proteins. J Agric. Food Chem. 2001, 49, 940–945. [Google Scholar] [CrossRef]
- Bennick, A. Interaction of plant polyphenols with salivary proteins. Crit. Rev. Oral Biol. Med. 2002, 13, 184–196. [Google Scholar] [CrossRef]
- Jöbstl, E.; O’Connell, J.; Fairclough, J.P.A.; Williamson, M.P. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar] [CrossRef]
- Dangles, O.; Dufour, C. Flavonoid-protein interactions. In Flavonoids - Chemistry, Biochemistry and Applications; Andersen, O.M., Markham, K.R., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 443–470. [Google Scholar]
- Croft, A.K.; Foley, M.K. Proline-rich proteins—deriving a basis for residue-based selectivity in polyphenolic binding. Org. Biomol. Chem. 2008, 6, 1594–1600. [Google Scholar] [CrossRef]
- Charlton, A.J.; Baxter, N.J.; Lilley, T.H.; Haslam, E.; McDonald, C.J.; Williamson, M.P. Tannin interactions with a full-length human salivary proline-rich protein display a stronger affinity than with single proline-rich repeats. FEBS Lett. 1996, 382, 289–292. [Google Scholar] [CrossRef]
- Pascal, C.; Paté, F.; Cheynier, V.; Delsuc, M. Study of the interactions between a proline-rich protein and a flavan-3-ol by NMR: Residual structures in the natively unfolded protein provides anchorage points for the ligands. Biopolymers 2009, 91, 745–756. [Google Scholar] [CrossRef]
- Canon, F.; Paté, F.; Meudec, E.; Marlin, T.; Cheynier, V.; Giuliani, A.; Sarni-Manchado, P. Characterization, stoichiometry, and stability of salivary protein–tannin complexes by ESI-MS and ESI-MS/MS. Anal. Bioanal. Chem. 2009, 395, 2535–2545. [Google Scholar] [CrossRef]
- Lu, Y.; Bennick, A. Interaction of tannin with human salivary proline-rich proteins. Arch. Oral Biol. 1998, 43, 717–728. [Google Scholar] [CrossRef]
- Boze, H.; Marlin, T.; Durand, D.; Pérez, J.; Vernhet, A.; Canon, F.; Sarni-Manchado, P.; Cheynier, V.; Cabane, B. Proline-rich salivary proteins have extended conformations. Biophys. J. 2010, 99, 656–665. [Google Scholar] [CrossRef]
- Bacon, J.R.; Rhodes, M.J.C. Binding affinity of hydrolyzable tannins to parotid saliva and to proline-rich proteins derived from it. J. Agric. Food Chem. 2000, 48, 838–843. [Google Scholar] [CrossRef]
- Pascal, C.; Poncet-Legrand, C.; Cabane, B.; Vernhet, A. Aggregation of a proline-rich protein induced by epigallocatechin gallate and condensed tannins: Effect of protein glycosylation. J. Agric. Food Chem. 2008, 56, 6724–6732. [Google Scholar]
- Sarni-Manchado, P.; Canals-Bosch, J.-M.; Mazerolles, G.; Cheynier, V. Influence of the glycosylation of human salivary proline-rich proteins on their interactions with condensed tannins. J. Agric. Food Chem. 2008, 56, 9563–9569. [Google Scholar]
- Yan, Q.; Bennick, A. Identification of histatins as tannin-binding proteins in human saliva. Biochem. J. 1995, 311, 341–347. [Google Scholar]
- Naurato, N.; Wong, P.; Lu, Y.; Wroblewski, K.; Bennick, A. Interaction of tannin with human salivary histatins. J. Agric. Food Chem. 1999, 47, 2229–2234. [Google Scholar] [CrossRef]
- Sarneckis, C.; Dambergs, R.G.; Jones, P.; Mercurio, M.; Herderich, M.J.; Smith, P. Quantification of condensed tannins by precipitation with methyl cellulose: development and validation of an optimised tool for grape and wine analysis. Aust. J. Grape Wine Res. 2006, 12, 39–49. [Google Scholar] [CrossRef]
- Pocock, K.F.; Sefton, M.A.; Williams, P.J. Taste thresholds of phenolic extracts of French and American oakwood: The influence of oak phenols on wine flavor. Am. J. Enol. Viticult. 1994, 45, 429–434. [Google Scholar]
- Peyrot des Gachons, C.; Kennedy, J.A. Direct method for determining seed and skin proanthocyanidin extraction into red wine. J. Agric. Food Chem. 2003, 51, 5877–5881. [Google Scholar] [CrossRef]
- Herderich, M.J.; Smith, P.A. Analysis of grape and wine tannins: Methods, applications and challenges. Aust. J. Grape Wine Res. 2005, 11, 1–10. [Google Scholar]
- Busse-Valverde, N.; Gómez-Plaza, E.; López-Roca, J.M.; Gil-Muñoz, R.; Fernández- Fernández, J.I.; Bautista-Ortiń, A.B. Effect of different enological practices on skin and seed proanthocyanidins in three varietal wines. J. Agric. Food Chem. 2010, 58, 11333–11339. [Google Scholar]
- Vidal, S.; Courcoux, P.; Francis, L.; Kwiatkowski, M.; Gawel, R.; Williams, P.; Waters, E.; Cheynier, V. Use of an experimental design approach for evaluation of key wine components on mouth-feel perception. Food Qual. Prefer. 2004, 15, 209–217. [Google Scholar] [CrossRef]
- Cheynier, V.; Prieur, C.; Guyot, S.; Rigaud, J.; Moutounet, M. The structures of tannins in grapes and wines and their interactions with proteins. In Wine - Nutritional and Therapeutic Benefits; Watkins, T.R., Ed.; ACS: Washington, DC, USA, 1997; pp. 81–93. [Google Scholar]
- Kennedy, J.A.; Hayasaka, Y.; Vidal, S.; Waters, E.J.; Jones, G.P. Composition of grape skin proanthocyanidins at different stages of berry development. J. Agric. Food Chem. 2001, 49, 5348–5355. [Google Scholar] [CrossRef]
- Cosme, F.; Ricardo-Da-Silva, J.M.; Laureano, O. Tannin profiles of Vitis vinifera L. cv. red grapes growing in Lisbon and from their monovarietal wines. Food Chem. 2009, 112, 197–204. [Google Scholar] [CrossRef]
- Souquet, J.-M.; Cheynier, V.; Brossaud, F.; Moutounet, M. Polymeric proanthocyanidins from grape skins. Phytochemistry 1996, 43, 509–512. [Google Scholar] [CrossRef]
- Bindon, K.; Smith, P.; Kennedy, J.A. Interaction between grape-derived proanthocyanidins and cell wall material. 1. Effect on proanthocyanidin composition and molecular mass. J. Agric. Food Chem. 2010, 58, 2520–2528. [Google Scholar] [CrossRef]
- Monagas, M.; Bartolome, B.; Gomez-Cordoves, C. Updated knowledge about the presence of phenolic compounds in wine. Crit. Rev. Food Sci. Nutr. 2005, 45, 85–118. [Google Scholar] [CrossRef]
- Cheynier, V. Flavonoids in wine. In Flavonoids - Chemistry, Biochemistry and Applications; Andersen, O.M., Markham, K.R., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Riou, V.; Vernhet, A.; Doco, T.; Moutounet, M. Aggregation of grape seed tannins in model wine--effect of wine polysaccharides. Food Hydrocolloid. 2002, 16, 17–23. [Google Scholar] [CrossRef]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutounet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Bindon, K.A.; Smith, P.A.; Holt, H.; Kennedy, J.A. Interaction between grape-derived proanthocyanidins and cell wall material. 2. Implications for vinification. J. Agric. Food Chem. 2010, 58, 10736–10746. [Google Scholar] [CrossRef]
- Lee, J.; Kennedy, J.A.; Devlin, C.; Redhead, M.; Rennaker, C. Effect of early seed removal during fermentation on proanthocyanidin extraction in red wine: A commercial production example. Food Chem. 2008, 107, 1270–1273. [Google Scholar] [CrossRef]
- Souquet, J.-M.; Labarbe, B.; Guernevé, C.L.; Cheynier, V.; Moutounet, M. Phenolic composition of grape stems. J. Agric. Food Chem. 2000, 48, 1076–1080. [Google Scholar] [CrossRef]
- Vivas, N.; Nonier, M.F.; Gaulejac, N.V.d. Structural characterization and analytical differentiation of grape seeds, skins, stems and Quebracho tannins. Bull. OIV 2004, 77, 643–659. [Google Scholar]
- Cala, O.; Pinaud, N.; Simon, C.; Fouquet, E.; Laguerre, M.; dufourc, E.J.; Pianet, I. NMR and molecular modeling of wine tannins binding to saliva proteins: revisiting astringency from molecular and colloidal prospects. FASEB J. 2010, 24, 1–10. [Google Scholar] [CrossRef]
- Fulcrand, H.; Duenas, M.; Salas, E.; Cheynier, V. Phenolic reactions during winemaking and aging. Am. J. Enol. Viticult. 2006, 57, 289–297. [Google Scholar]
- Drinkine, J.; Glories, Y.; Saucier, C. (+)-Catechin-aldehyde condensations: Competition between acetaldehyde and glyoxylic acid. J. Agric. Food Chem. 2005, 53, 7552–7558. [Google Scholar] [CrossRef]
- Es-Safi, N.E.; Fulcrand, H.; Cheynier, V.; Moutounet, M. Studies on the acetaldehyde-induced condensation of (-)-epicatechin and malvidin 3-O-glucoside in a model solution system. J. Agric. Food Chem. 1999, 47, 2096–2102. [Google Scholar] [CrossRef]
- Dallas, C.; Ricardo-da-Silva, J.M.; Laureano, O. Interactions of oligomeric procyanidins in model wine solutions containing malvidin-3- - glucoside and acetaldehyde. J. Sci. Food Agric. 1996, 70, 493–500. [Google Scholar] [CrossRef]
- Dallas, C.; Ricardo-da-Silva, J.M.; Laureano, O. Products formed in model wine solutions involving anthocyanins, procyanidin B2, and acetaldehyde. J. Agric. Food Chem. 1996, 44, 2402–2407. [Google Scholar] [CrossRef]
- Poncet-Legrand, C.; Cabane, B.; Bautista-Ortín, A.; Carrillo, S.; Fulcrand, H.; Pérez, J.; Vernhet, A. Tannin oxidation: Intra- versus intermolecular reactions. Biomacromolecules 2010, 11, 2376–2386. [Google Scholar] [CrossRef]
- McRae, J.M.; Falconer, R.J.; Kennedy, J.A. Thermodynamics of grape and wine tannin interaction with polyproline: Implications for red wine astringency. J. Agric. Food Chem. 2010, 58, 12510–12518. [Google Scholar] [CrossRef]
- Haslam, E. In Vino Veritas - Oligomeric procyanidins and the aging of red wines. Phytochemistry 1980, 19, 2577–2582. [Google Scholar] [CrossRef]
- Mateus, N.; Oliveira, J.; Pissarra, J.; Gonzalez-Paramas, A.M.; Rivas-Gonzalo, J.C.; Santos-Buelga, C.; Silva, A.M.S.; de Freitas, V. A new vinylpyranoanthocyanin pigment occurring in aged red wine. Food Chem. 2006, 97, 689–695. [Google Scholar] [CrossRef]
- Mateus, N.; Silva, A.M.S.; Rivas-Gonzalo, J.C.; Santos-Buelga, C.; De Freitas, V. A new class of blue anthocyanin-derived pigments isolated from red wines. J. Agric. Food Chem. 2003, 51, 1919–1923. [Google Scholar] [CrossRef]
- Salas, E.; Atanasova, V.; Poncet-Legrand, C.; Meudec, E.; Mazauric, J.P.; Cheynier, V. Demonstration of the occurrence of flavanol-anthocyanin adducts in wine and in model solutions. Anal. Chim. Acta 2004, 513, 325–332. [Google Scholar] [CrossRef]
- Salas, E.; Fulcrand, H.; Poncet-LeGrand, C.; Meudec, E.; Kohler, N.; Winterhalter, P.; Cheynier, V. Isolation of flavanol-anthocyanin adducts by countercurrent chromatography. J. Chromatogr. Sci. 2005, 43, 488–493. [Google Scholar]
- Fulcrand, H.; Atanasova, V.; Salas, E.; Cheynier, V. The fate of anthocyanins in wine: Are there determining factors? In Red Wine Color - Revealing the Mysteries; Waterhouse, A.L., Kennedy, J.A., Eds.; American Chemical Society: Washington, DC, USA, 2004; pp. 68–88. [Google Scholar]
- Mateus, N.; de Freitas, V. Evolution and stability of anthocyanin-derived pigments during port wine aging. J. Agric. Food Chem. 2001, 49, 5217–5222. [Google Scholar] [CrossRef]
- Simon, C.; Barathieu, K.; Laguerre, M.; Schmitter, J.-M.; Fouquet, E.; Pianet, I.; Dufourc, E.J. Three-dimensional structure and dynamics of wine tannin-saliva protein complexes, a multitechnique approach. Biochemistry 2003, 42, 10385–10395. [Google Scholar] [CrossRef]
- Wróblewski, K.; Muhandiram, R.; Chakrabartty, A.; Bennick, A. The molecular interaction of human salivary histatins with polyphenolic compounds. Eur. J. Biochem. 2001, 268, 4384–4397. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Rice, M.E.; Ritchard, N.T. Mechanisms of protein precipitation for two tannins, pentagalloyl glucose and epicatechin16 (4f8) catechin (procyanidin). J. Agric. Food Chem. 1998, 46, 2590–2595. [Google Scholar] [CrossRef]
- Kawamoto, H.; Nakatsubo, F. Effects of environmental factors on two-stage tannin-protein co-precipitation. Phytochemistry 1997, 46, 479–483. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Guyot, S.; Marnet, N.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E.J. The mouth-feel properties of grape and apple proanthocyanidins in a wine-like medium. J. Sci. Food Agr. 2003, 83, 564–573. [Google Scholar] [CrossRef]
- Hofmann, T.; Glabasnia, A.; Schwarz, B.; Wisman, K.N.; Gangwer, K.A.; Hagerman, A.E. Protein binding and astringent taste of a polymeric procyanidin, 1,2,3,4,6-penta-O-galloyl-beta-D-glucopyranose, castalagin, and grandinin. J. Agric. Food Chem. 2006, 54, 9503–9509. [Google Scholar] [CrossRef]
- Cheynier, V.; Duenas-Paton, M.; Salas, E.; Maury, C.; Souquet, J.M.; Sarni-Manchado, P.; Fulcrand, H. Structure and properties of wine pigments and tannins. Am. J. Enol. Viticult. 2006, 57, 298–305. [Google Scholar]
- Poncet-Legrand, C.; Edelmann, A.; Putaux, J.-L.; Cartalade, D.; Sarni-Manchado, P.; Vernhet, A. Poly(L-proline) interactions with flavan-3-ols units: Influence of the molecular structure and the polyphenol/protein ratio. Food Hydrocolloid. 2003, 20, 687–697. [Google Scholar]
- Deaville, E.R.; Green, R.J.; Mueller-Harvey, I.; Willoughby, I.; Frazier, R.A. Hydrolyzable tannin structures influence relative globular and random coil protein Binding Strengths. J. Agric. Food Chem. 2007, 55, 4554–4561. [Google Scholar]
- Richard, T.; Lefeuvre, D.; Descendit, A.; Quideau, S.; Monti, J.P. Recognition characters in peptide-polyphenol complex formation. BBA Gen. Subjects 2006, 1760, 951–958. [Google Scholar] [CrossRef]
- Poncet-Legrand, C.; Gautier, C.; Cheynier, V.; Imberty, A. Interactions between flavan-3-ols and poly(L-proline) studied by Isothermal Titration Calorimetry: Effect of the tannin structure. J. Agric. Food Chem. 2007, 55, 9235–9240. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Guyot, S.; Renard, C.M.G.C. Non-covalent interaction between procyanidins and apple cell wall material: Part I. Effect of some environmental parameters. BBA Gen. Subjects 2004, 1672, 192–202. [Google Scholar] [CrossRef]
- Tarascou, I.; Barathieu, K.; Simon, C.; Ducasse, M.A.; Andre, Y.; Fouquet, E.; Dufourc, E.J.; de Freitas, V.; Laguerre, M.; Pianet, I. A 3D structural and conformational study of procyanidin dimers in water and hydro-alcoholic media as viewed by NMR and molecular modeling. Magn. Reson. Chem. 2006, 44, 868–880. [Google Scholar] [CrossRef]
- Cala, O.; Fabre, S.; Fouquet, E.; Dufourc, E.J.; Pianet, I. NMR of human saliva protein/wine tannin complexes. Towards deciphering astringency with physico-chemical tools. C.R. Chim. 2010, 13, 449–452. [Google Scholar] [CrossRef]
- Vivar-Quintana, A.M.; Santos-Buelga, C.; Rivas-Gonzalo, J.C. Anthocyanin-derived pigments and colour of red wines. Anal. Chim. Acta 2002, 458, 147–155. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Calderón, F.; Suárez, J.A. Effects of pH, temperature and SO2 on the formation of pyranoanthocyanins during red wine fermentation with two species of Saccharomyces. Int. J. Food Microbiol. 2006, 106, 123–129. [Google Scholar] [CrossRef]
- Li, H.; Guo, A.; Wang, H. Mechanisms of oxidative browning of wine. Food Chem. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Waters, E.J.; Peng, Z.; Pocock, K.F.; Jones, G.P.; Clarke, P.; Williams, P.J. Solid-state 13C NMR investigation into insoluble deposits adhering to the inner glass surface of bottled red wine. J. Agric. Food Chem. 1994, 1994, 1761–1766. [Google Scholar]
- Herderich, M.; Birse, M.; Dambergs, R.; Holt, H.; Iland, P.; Lattey, K.; Smith, P.A. Grape and wine tannins - an overview on current research, emerging applications, and future challenges. In Advances in Tannin and Tannin Management, Proceedings of AWRI Roadshow seminar, Adelaide Convention Centre, Australia, 6 October 2005; Allen, M., Dundon, C., Francis, M., Howell, G.S., Wall, G., Eds.; Australian Society of VIticulture and Oenology: Adelaide, Australia, 2006; pp. 4–10. [Google Scholar]
- Parish, M.; Wollan, D.; Paul, R. Micro-oxygenation – A Review. Aust. N. Z. Grapegrower Winemaker 2000, 438a, 47–50. [Google Scholar]
- Pozo, A.G.-d.; Arozarena, I.; Noriega, M.-J.; Navarro, M.; Casp, A. Short- and long-term effects of micro-oxygenation treatments on the colour and phenolic composition of a Cabernet Sauvignon wine aged in barrels and/or bottles. Eur. Food Res. Technol. 2010, 231, 589–601. [Google Scholar] [CrossRef]
- González-Sanjosé, M.L.; Ortega-Heras, M.; Pérez-Magariño, S. Microoxygenation treatment and sensory properties of young red wines. Food Sci. Technol. Int. 2008, 14, 123–130. [Google Scholar] [CrossRef]
- Cano-Lopez, M.; Pardo-Minguez, F.; López-Roca, J.; Gómez-Plaza, E. Chromatic characteristics and anthocyanin profile of a micro-oxygenated red wine after oak or bottle maturation. Eur. Food Res. Technol. 2007, 225, 127–132. [Google Scholar] [CrossRef]
- Llaudy, M.D.; Canals, R.; Gonzalez-Manzano, S.; Canals, J.M.; Santos-Buelga, C.; Zamora, F. Influence of micro-oxygenation treatment before oak aging on phenolic compounds composition, astringency, and color of red wine. J. Agric. Food Chem. 2006, 54, 4246–4252. [Google Scholar] [CrossRef]
- Ross, C.F.; Weller, K. Effect of serving temperature on the sensory attributes of red and white wines. J. Sens. Stud. 2008, 23, 398–416. [Google Scholar] [CrossRef]
- Obreque-Slíer, E.; Peña-Neira, A.; López-Solís, R. Enhancement of both salivary protein-enological tannin interactions and astringency perception by ethanol. J. Agric. Food Chem. 2010, 58, 3729–3735. [Google Scholar] [CrossRef]
- Poncet-Legrand, C.; Cartalade, D.; Putaux, J.-L.; Cheynier, V.; Vernhet, A. Flavan-3-ol aggregation in model ethanolic solutions:Incidence of polyphenol structure, concentration, ethanol content, and ionic strength. Langmuir 2003, 19, 10563–10572. [Google Scholar]
- Zanchi, D.; Konarev, P.V.; Tribet, C.; Baron, A.; Svergun, D.I.; Guyot, S. Rigidity, conformation, and solvation of native and oxidized tannin macromolecules in water-ethanol solution. J. Chem. Phys. 2009, 130, 245103. [Google Scholar] [CrossRef]
- Zanchi, D.; Vernhet, A.; Poncet-Legrand, C.; Cartalade, D.; Tribet, C.; Schweins, R.; Cabane, B. Colloidal dispersions of tannins in water-ethanol solutions. Langmuir 2007, 23, 9949–9959. [Google Scholar] [CrossRef]
- Kallithraka, S.; Bakker, J.; Clifford, M.N. Effect of pH on astringency in model solutions and wines. J. Agric. Food Chem. 1997, 45, 2211–2216. [Google Scholar] [CrossRef]
- Borse, B.B.; Rao, L.J.M.; Ramalakshmi, K.; Raghavan, B. Chemical composition of volatiles from coconut sap (neera) and effect of processing. Food Chem. 2006, 101, 877–880. [Google Scholar]
- Escot, S.; Feuillat, M.; Dulau, L.; Charpentier, C. Release of polysaccharides by yeasts and the influence of released polysaccharides on colour stability and wine astringency. Aust. J. Grape Wine Res. 2001, 7, 153–159. [Google Scholar] [CrossRef]
- Taira, S.; Ono, M.; Matsumoto, N. Reduction of persimmon astringency by complex formation between pectin and tannins. Postharvest Biol. Technol. 1997, 12, 265–271. [Google Scholar] [CrossRef]
- McManus, J.P.; Davis, K.G.; Beart, J.E.; Gaffney, S.H.; Lilley, T.H.; Haslam, E. Polyphenol interactions. part 1. introduction; some observations on the reversible complexation of polyphenols with proteins and polysaccharides. J. Chem. Soc.-Perk. T 2 1985, 1429–1438. [Google Scholar]
- Soares, S.I.; Gonçalves, R.M.; Fernandes, I.; Mateus, N.; de Freitas, V. Mechanistic approach by which polysaccharides inhibit α-amylase/procyanidin aggregation. J. Agric. Food Chem. 2009, 57, 4352–4358. [Google Scholar] [CrossRef]
- Poncet-Legrand, C.; Doco, T.; Williams, P.; Vernhet, A. Inhibition of grape seed tannin aggregation by wine mannoproteins: Effect of polysaccharide molecular weight. Am. J. Enol. Viticult. 2007, 58, 87–91. [Google Scholar]
- Boselli, E.; Minardi, M.; Giomo, A.; Frega, N.G. Phenolic composition and quality of white d.o.c. wines from Marche (Italy). Anal. Chim. Acta 2006, 563, 93–100. [Google Scholar] [CrossRef]
- Ishikawa, I.; Noble, A.C. Temporal perception of astringency and sweetness in red wine. Food Qual. Prefer. 1995, 6, 27–33. [Google Scholar] [CrossRef]
- Sáenz-Navajas, M.P.; Tao, Y.S.; Dizy, M.; Ferreira, V.; Fernández-Zurbano, P. Relationship between nonvolatile composition and sensory properties of premium Spanish red wines and their correlation to quality perception. J. Agric. Food Chem. 2010, 58, 12407–12416. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
McRae, J.M.; Kennedy, J.A. Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research. Molecules 2011, 16, 2348-2364. https://doi.org/10.3390/molecules16032348
McRae JM, Kennedy JA. Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research. Molecules. 2011; 16(3):2348-2364. https://doi.org/10.3390/molecules16032348
Chicago/Turabian StyleMcRae, Jacqui M., and James A. Kennedy. 2011. "Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research" Molecules 16, no. 3: 2348-2364. https://doi.org/10.3390/molecules16032348
APA StyleMcRae, J. M., & Kennedy, J. A. (2011). Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research. Molecules, 16(3), 2348-2364. https://doi.org/10.3390/molecules16032348