Fluorescence Resonance Energy Transfer Assay for High-Throughput Screening of ADAMTS1 Inhibitors

Abstract
:1. Introduction
2. Results and Discussion
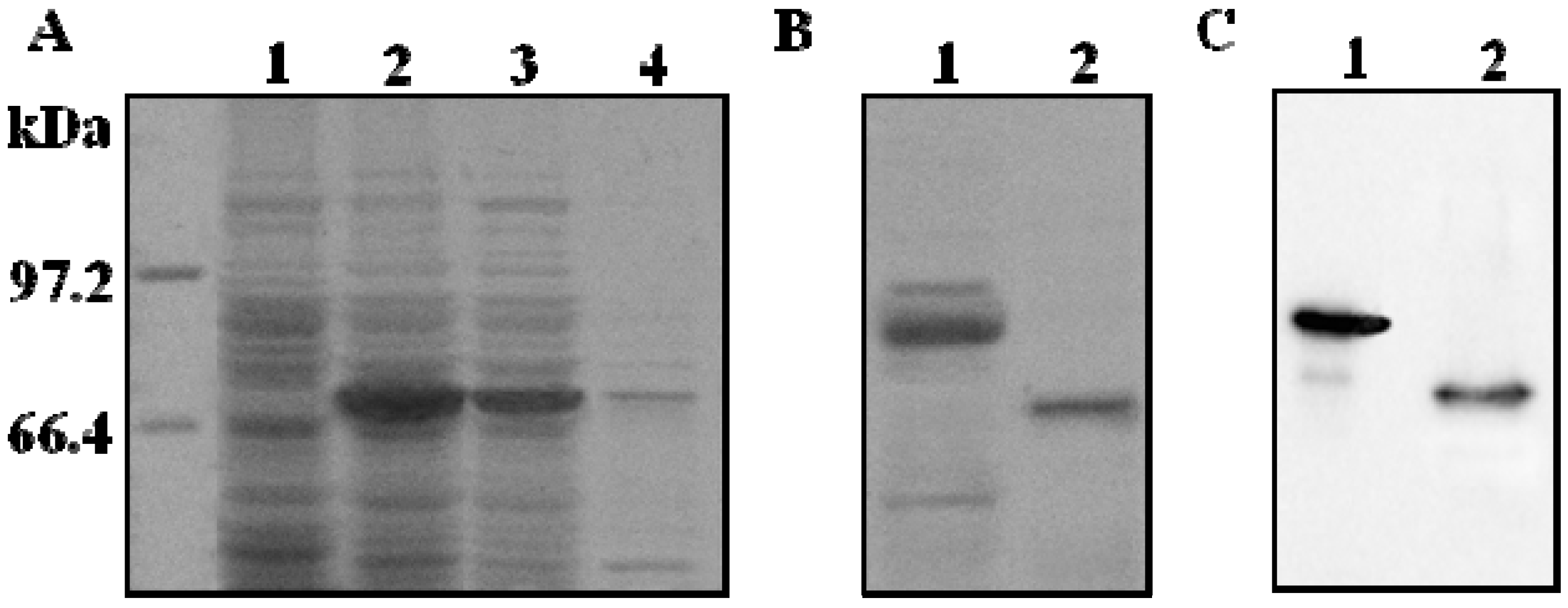
2.1. Expression of Recombinant ADAMTS1 in E. coli
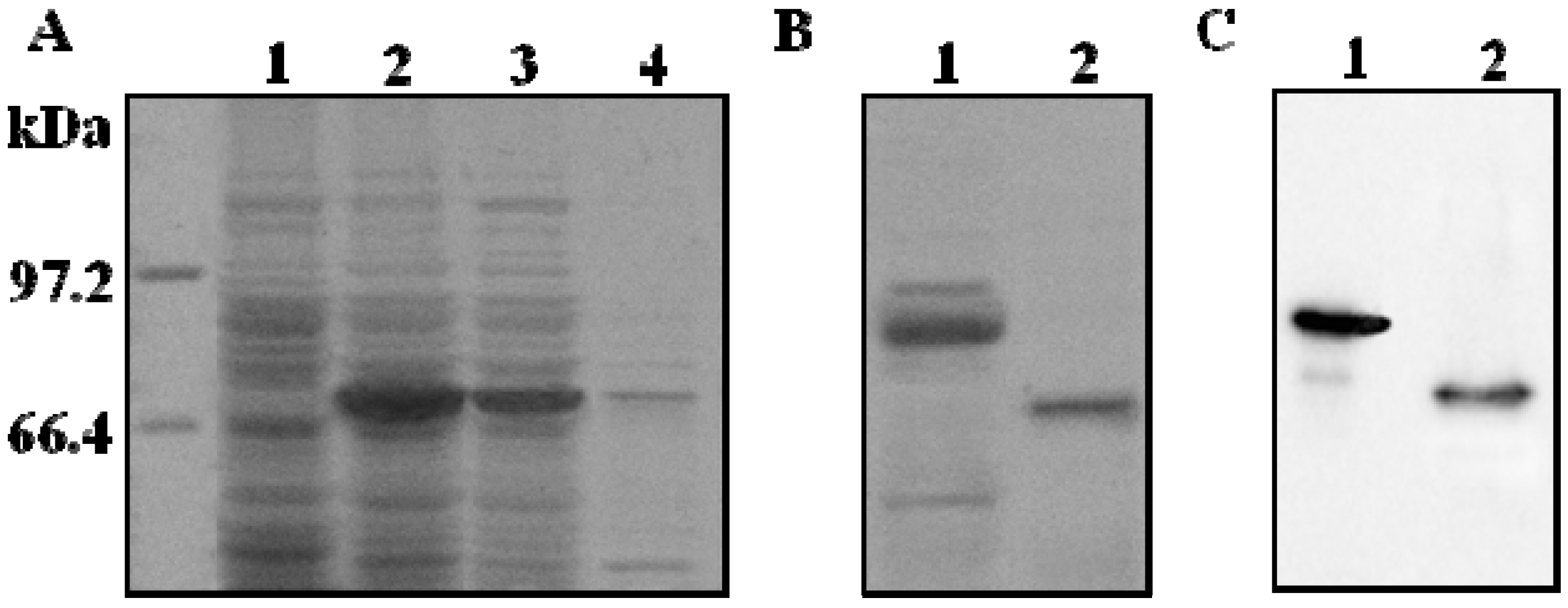
2.2. Purification and Proteolytic Cleavage of ADAMTS1 Fusion Protein
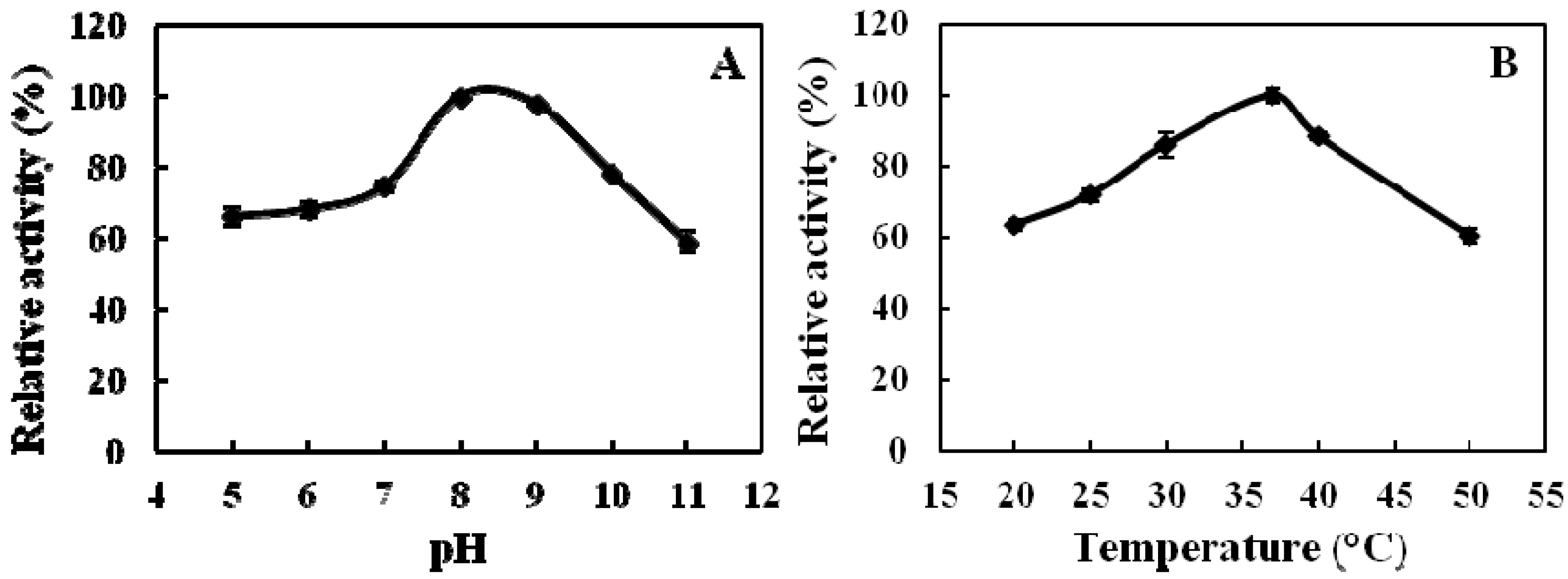
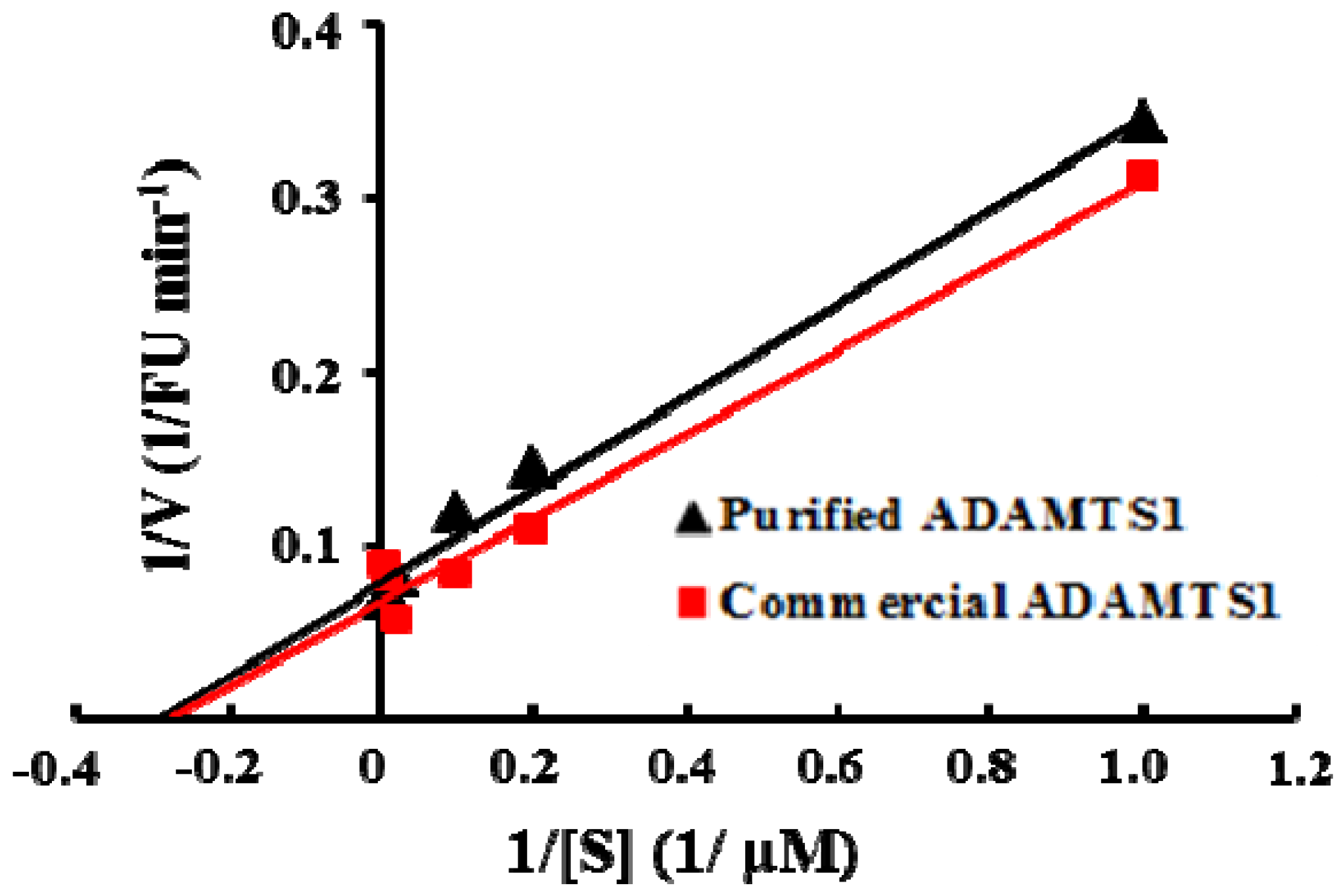
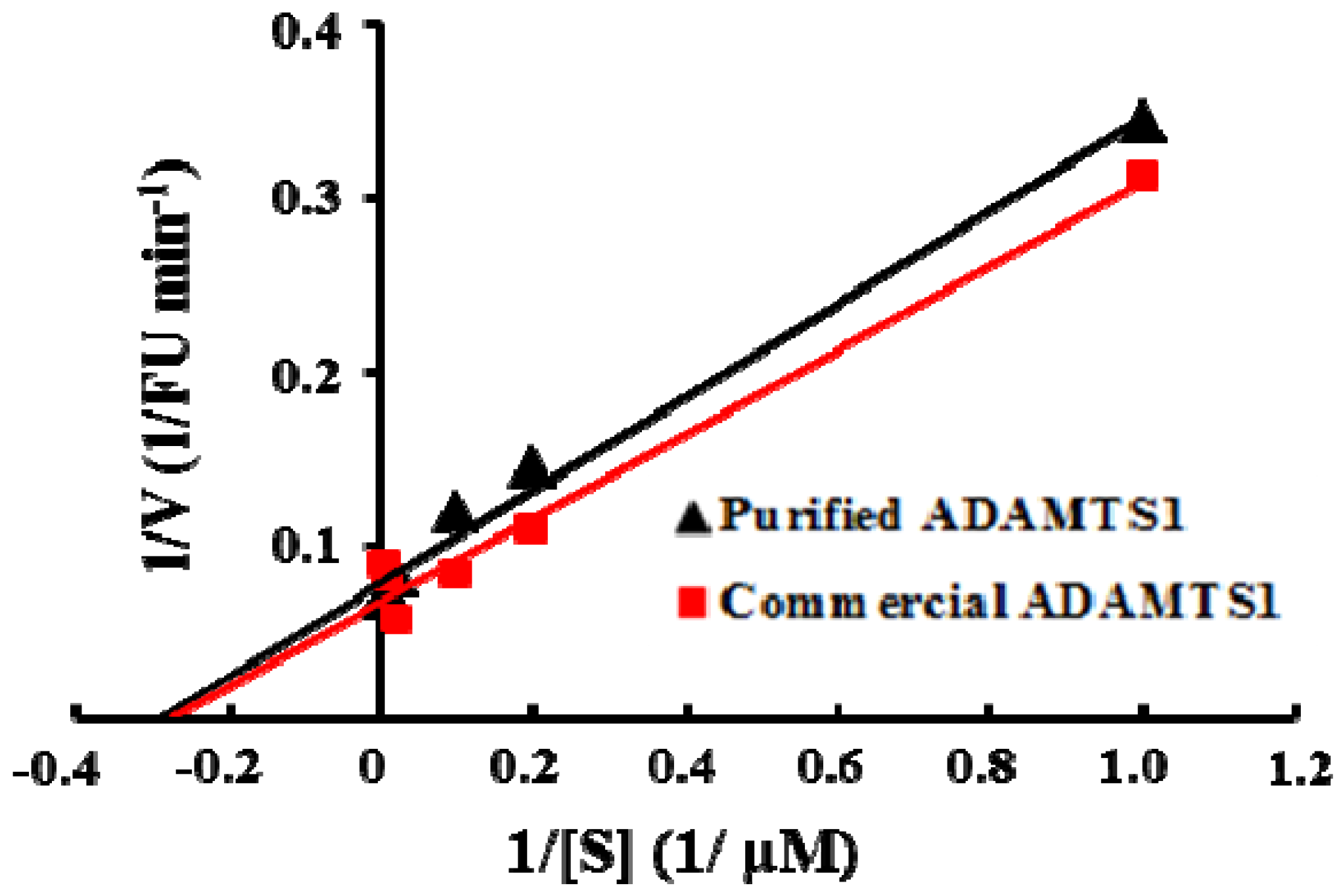
2.3. Properties of Recombinant ADAMTS1 Using the FRET Peptide
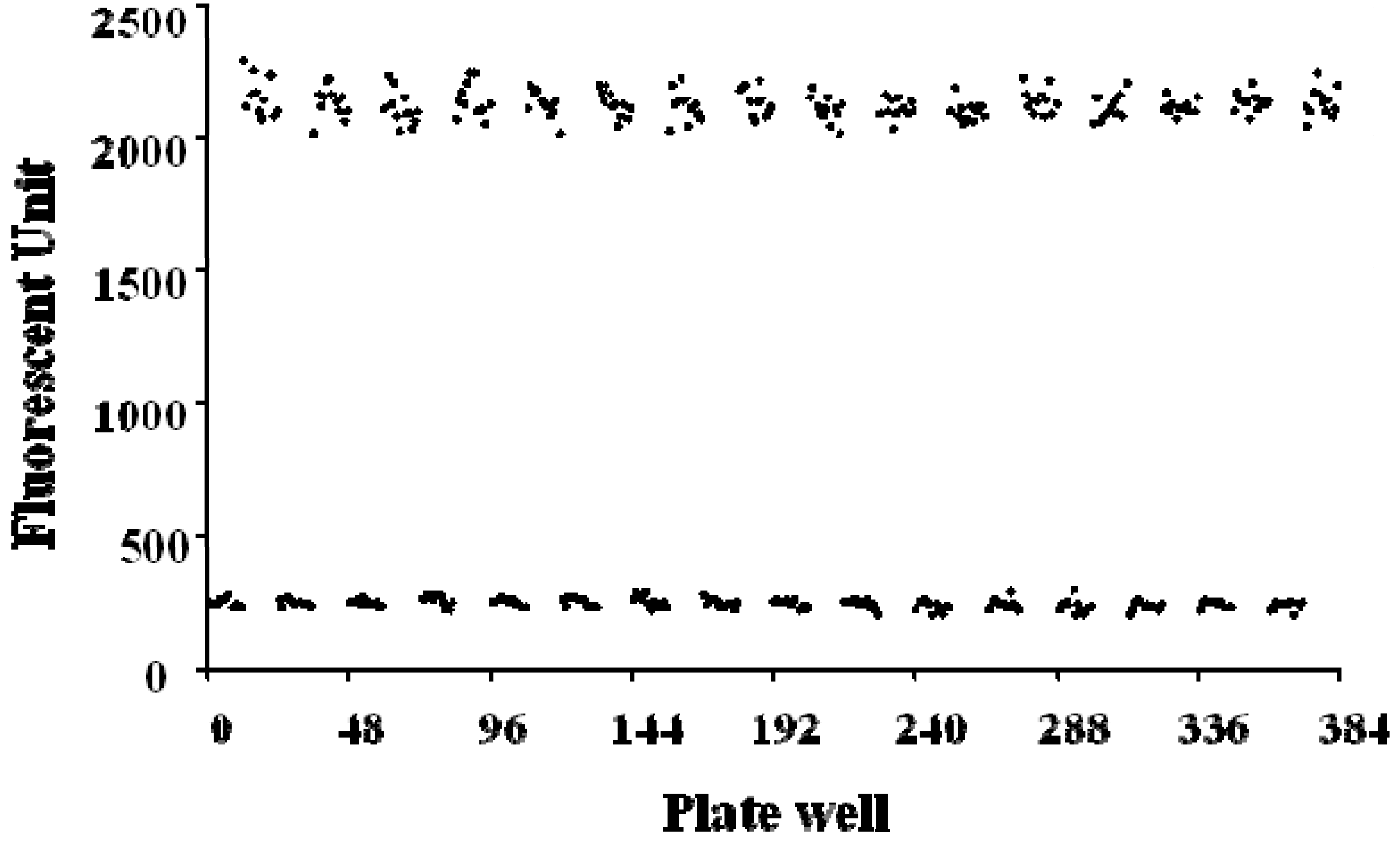
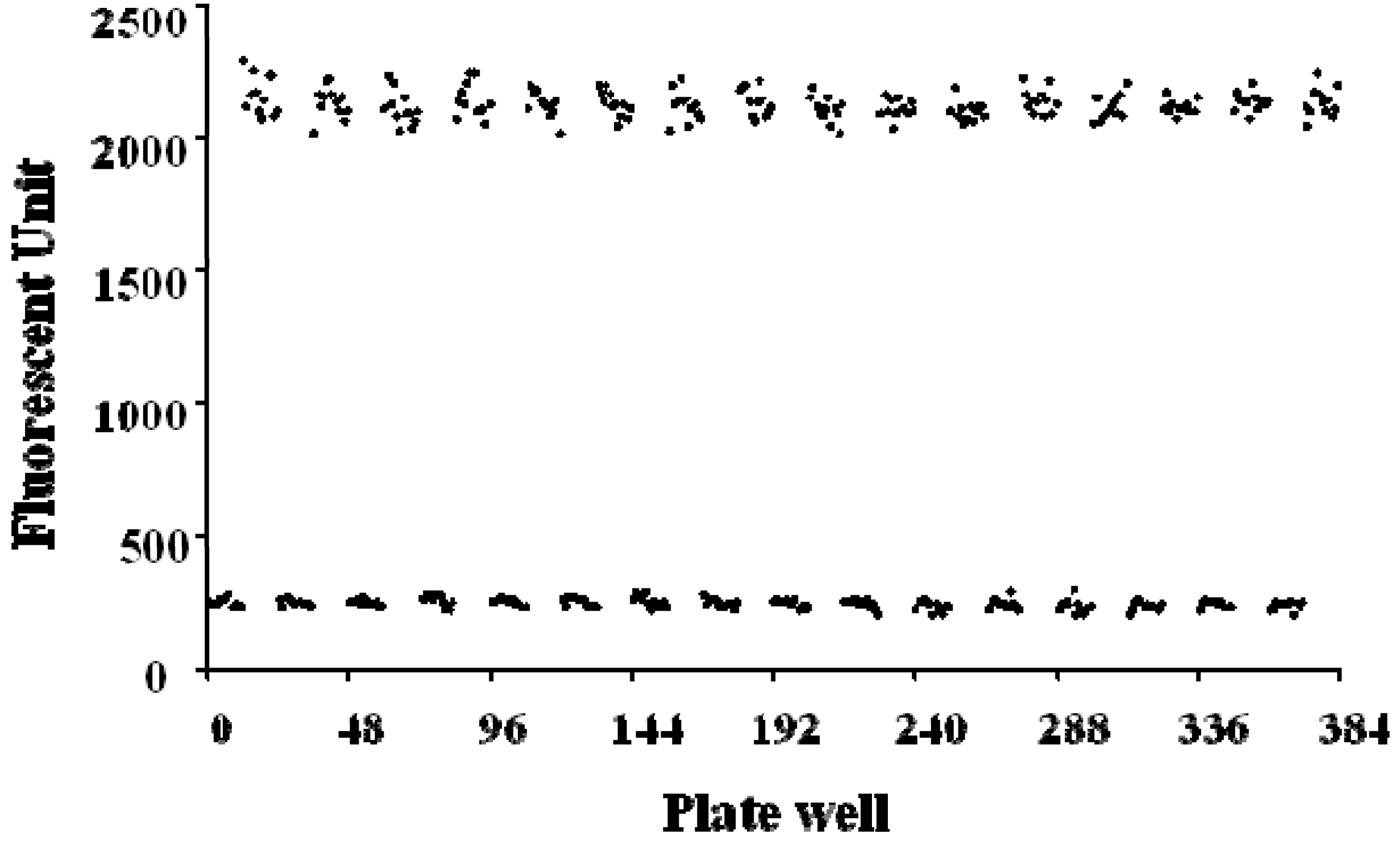
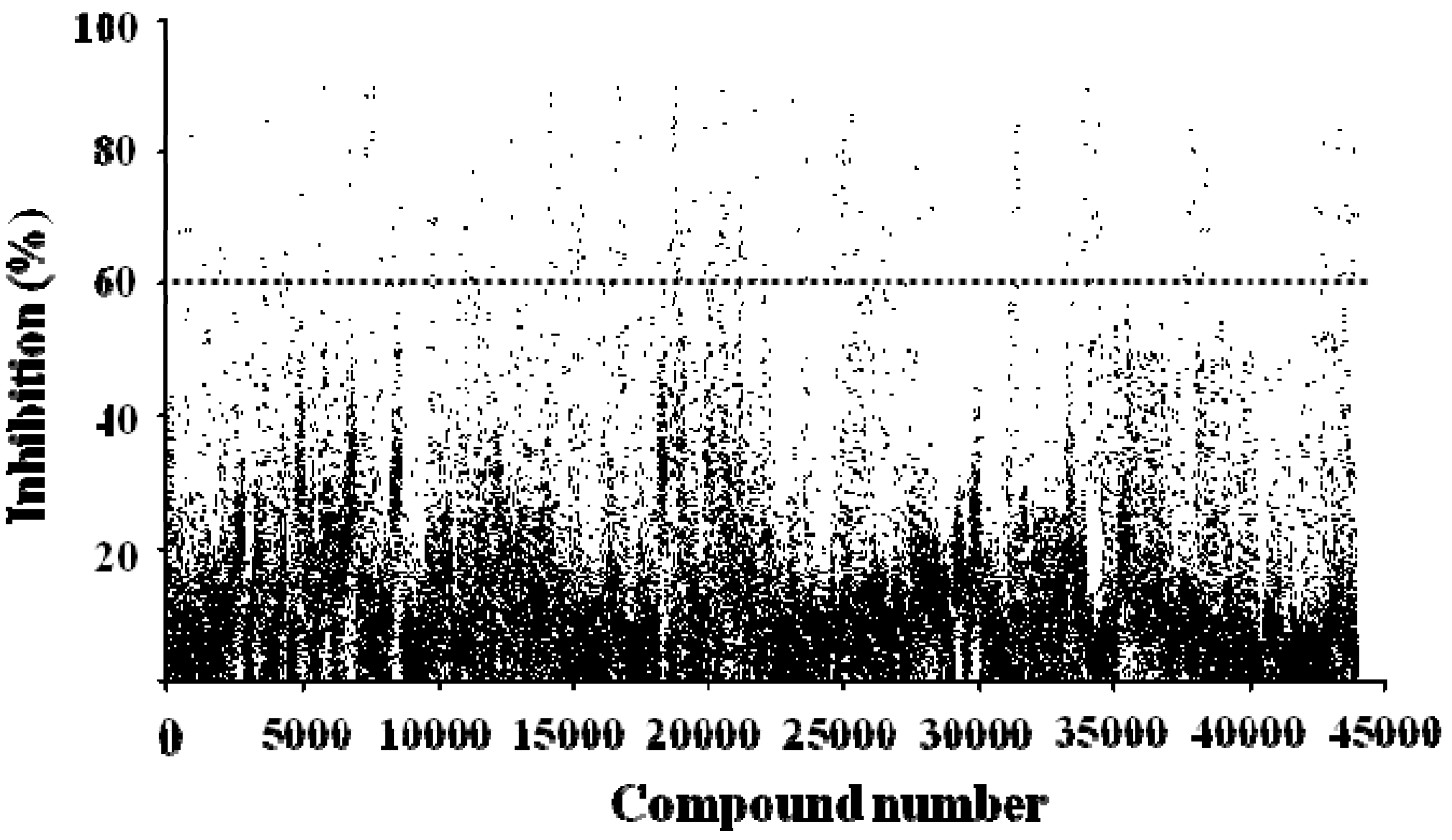
2.4. FRET-Based High-Throughput Drug Screening
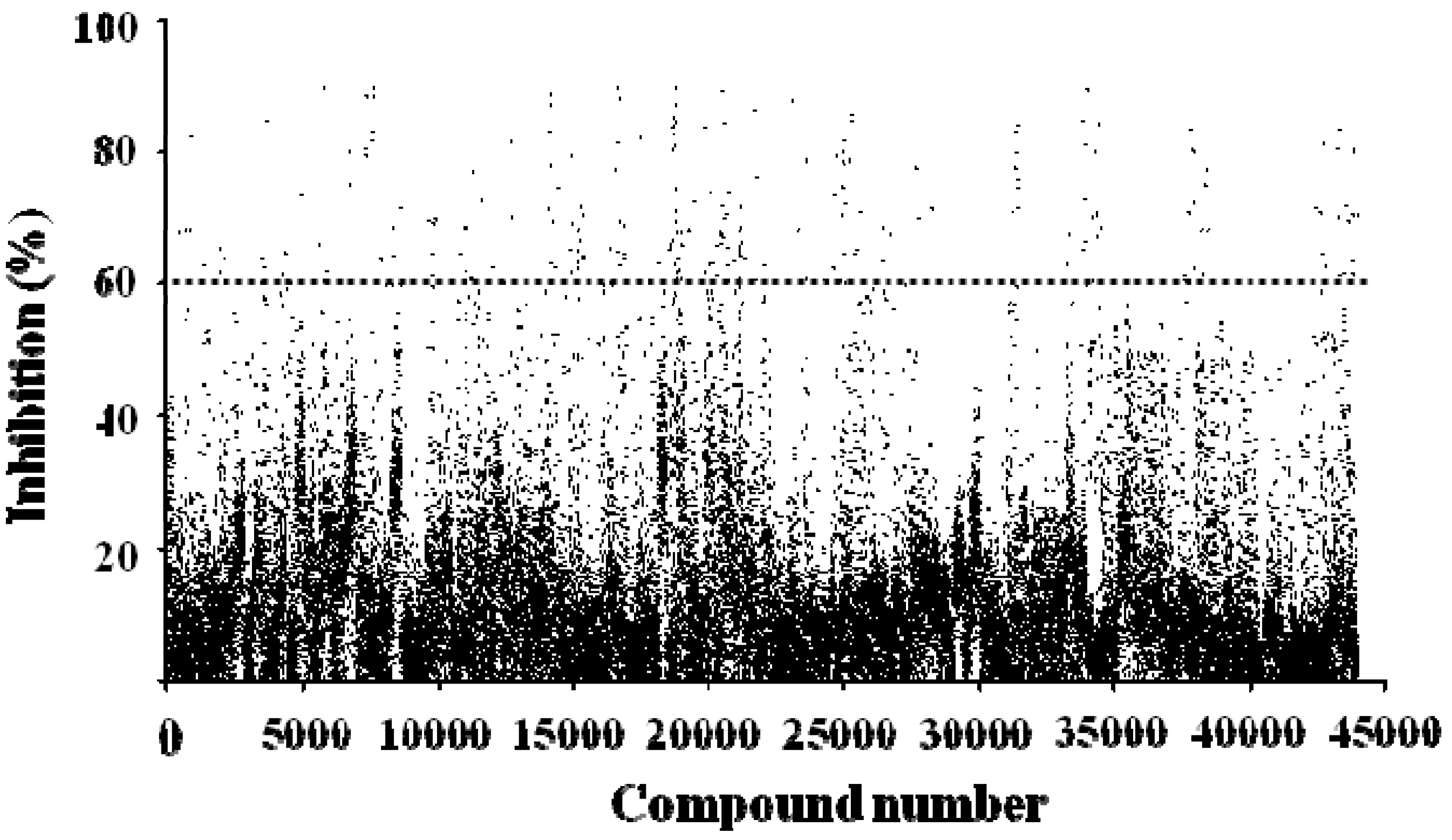
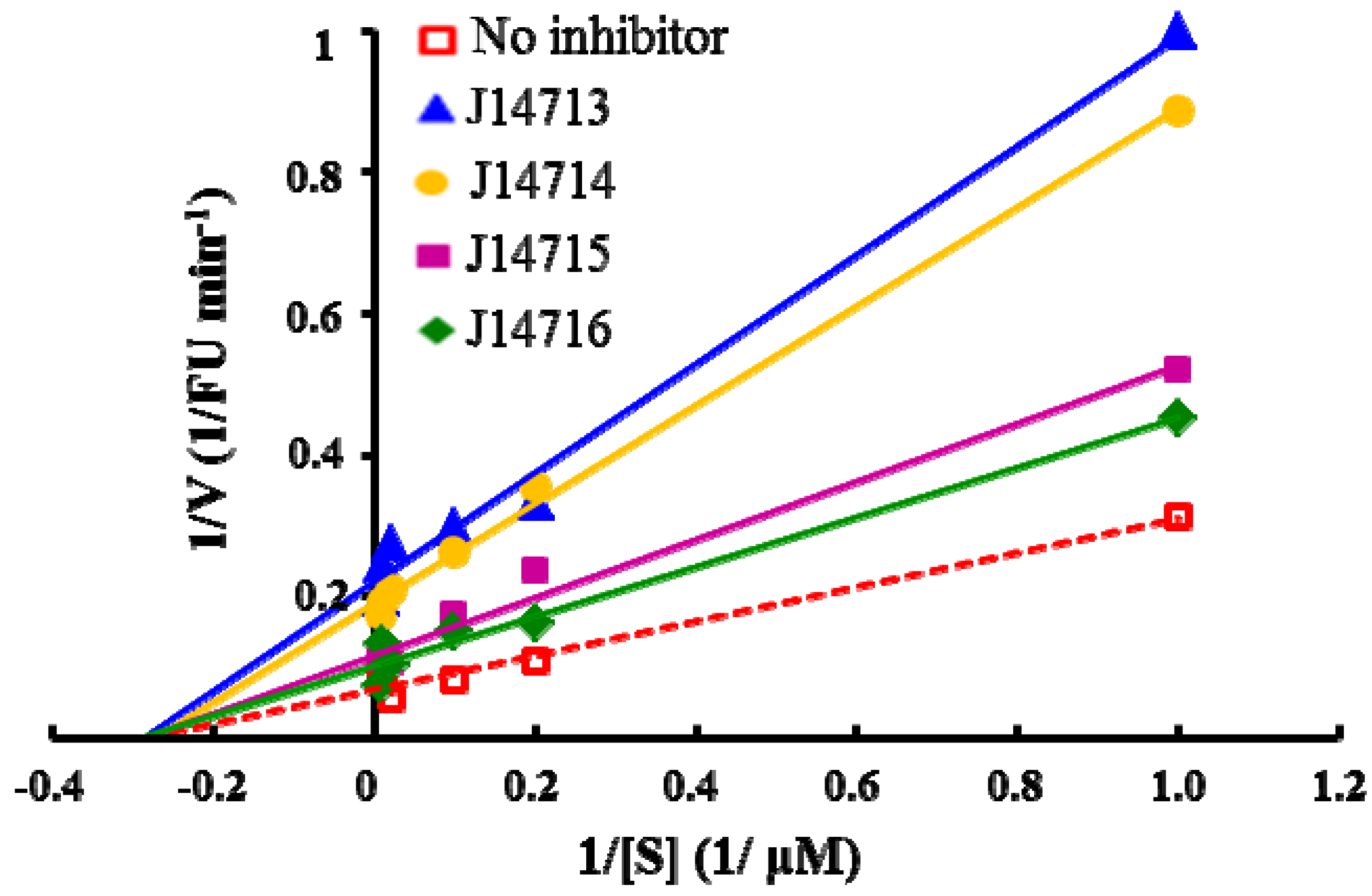
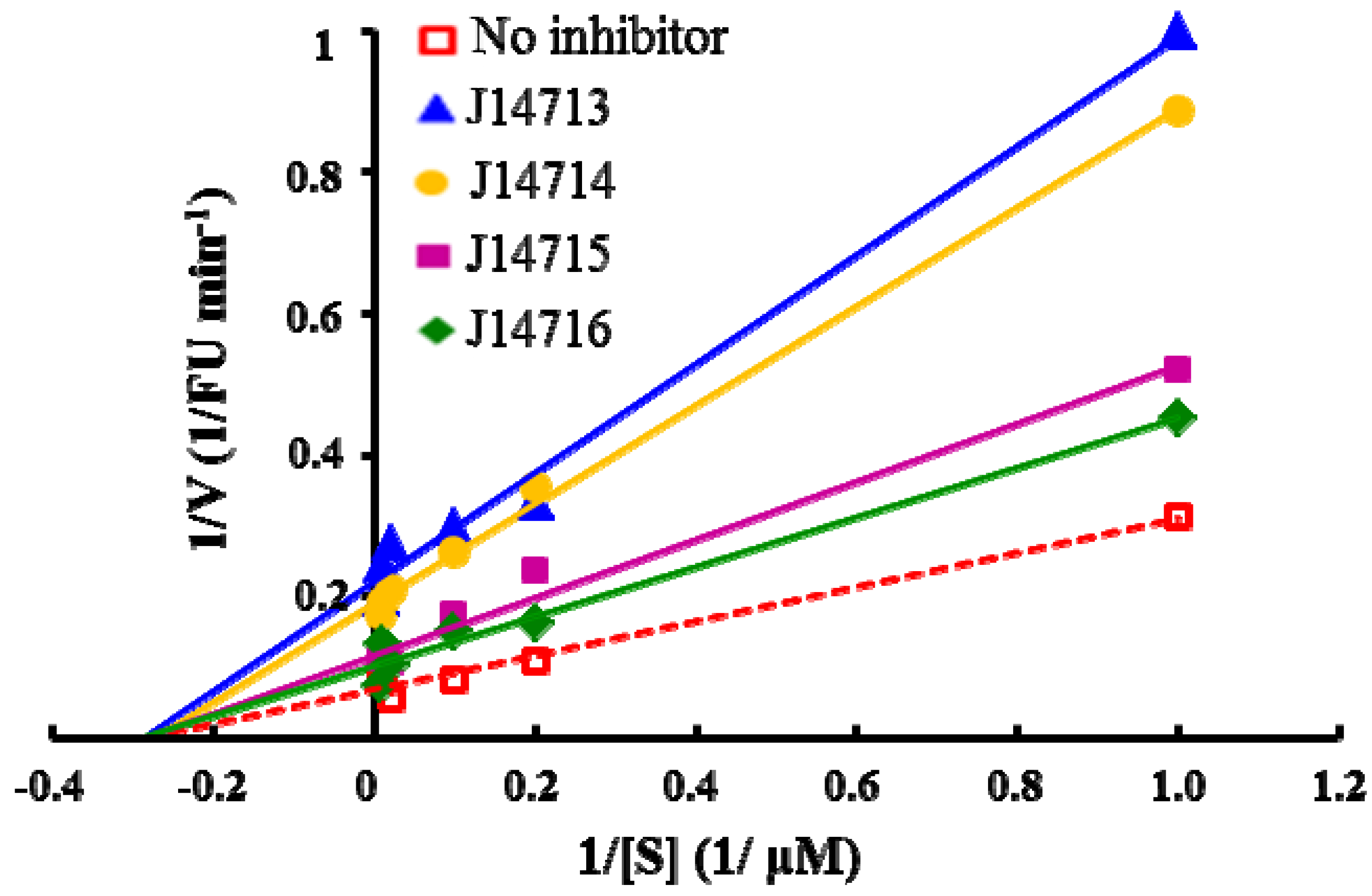
2.5. Evaluation of Four Hit compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Structure | IC50 (μM) * | IC50 (μM) # |
|---|---|---|---|
| J14713 Kuwanon P |  | 7.6 ± 0.6 | 5.5 ± 0.4 |
| J14714 Kuwanon X | 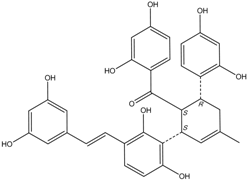 | 15.5 ± 0.8 | 16.2 ± 0.7 |
| J14715 Albafuran C | 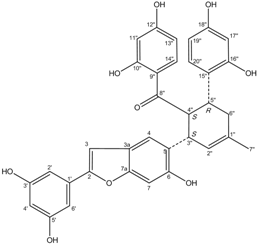 | 11.9 ± 0.7 | 10.7 ± 0.6 |
| J14716 Mulberrofuran J | 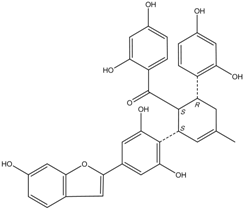 | 18.4 ± 1.4 | 25.4 ± 2.1 |
3. Experimental
3.1. Materials
3.2. Cloning, Expression and Purification of the Recombinant ADAMTS1
3.3. ADAMTS1 Activity Assay
3.4. Effects of pH, Temperature and DMSO Solvent on the ADAMTS1 Activity
3.5. Determination of the Kinetic Parameters Using the FRET Peptide
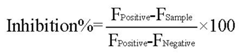
3.6. High-Throughput Drug Screening
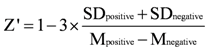
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Tang, B.L. ADAMTS: A novel family of extracellular matrix proteases. Int. J. Biochem. Cell Biol. 2001, 33, 33–44. [Google Scholar] [CrossRef]
- Mitani, H.; Takahashi, I.; Onodera, K.; Bae, J.W.; Sato, T.; Takahashi, N.; Sasano, Y.; Igarashi, K.; Mitani, H. Comparison of age-dependent expression of aggrecan and ADAMTSs in mandibular condylar cartilage, tibial growth plate, and articular cartilage in rats. Histochem. Cell Biol. 2006, 126, 371–380. [Google Scholar] [CrossRef]
- Pockert, A.J.; Richardson, S.M.; Le Maitre, C.L.; Lyon, M.; Deakin, J.A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Modified expression of the ADAMTS enzymes and tissue inhibitor of metalloproteinases 3 during human intervertebral disc degeneration. Arthritis Rheum. 2009, 60, 482–491. [Google Scholar] [CrossRef]
- Song, R.H.; Tortorella, M.D.; Malfait, A.M.; Alston, J.T.; Yang, Z.; Arner, E.C.; Griggs, D.W. Aggrecan degradation in human articular cartilage explants is mediated by both ADAMTS-4 and ADAMTS-5. Arthritis Rheum. 2007, 56, 575–585. [Google Scholar] [CrossRef]
- Kuno, K.; Terashima, Y.; Matsushima, K. ADAMTS-1 is an active metalloproteinase associated with the extracellular matrix. J. Biol. Chem. 1999, 274, 18821–18826. [Google Scholar] [CrossRef]
- Kuno, K.; Kanada, N.; Nakashima, E.; Fujiki, F.; Ichimura, F.; Matsushima, K. Molecular cloning of a gene encoding a new type of metalloproteinase-disintegrin family protein with thrombospondin motifs as an inflammation associated gene. J. Biol. Chem. 1997, 272, 556–562. [Google Scholar]
- Kuno, K.; Matsushima, K. ADAMTS-1 protein anchors at the extracellular matrix through the thrombospondin type I motifs and its spacing region. J. Biol. Chem. 1998, 273, 13912–13917. [Google Scholar] [CrossRef]
- Rodriguez-Manzaneque, J.C.; Milchanowski, A.B.; Dufour, E.K.; Leduc, R.; Iruela-Arispe, M.L. Characterization of METH-1/ADAMTS1 processing reveals two distinct active forms. J. Biol. Chem. 2000, 275, 33471–33479. [Google Scholar]
- Liu, Y.J.; Xu, Y.; Yu, Q. Full-length ADAMTS-1 and the ADAMTS-1 fragments display pro- and antimetastatic activity, respectively. Oncogene 2006, 25, 2452–2467. [Google Scholar] [CrossRef]
- Kuno, K.; Okada, Y.; Kawashima, H.; Nakamura, H.; Miyasaka, M.; Ohno, H.; Matsushima, K. ADAMTS-1 cleaves a cartilage proteoglycan, aggrecan. FEBS Lett. 2000, 478, 241–245. [Google Scholar] [CrossRef]
- Rodriguez-Manzaneque, J.C.; Westling, J.; Thai, S.N.; Luque, A.; Knauper, V.; Murphy, G.; Sandy, J.D.; Iruela-Arispe, M.L. ADAMTS1 cleaves aggrecan at multiple sites and is differentially inhibited by metalloproteinase inhibitors. Biochem. Biophys. Res. Commun. 2002, 293, 501–508. [Google Scholar] [CrossRef]
- Gooljarsingh, L.T.; Lakdawala, A.; Coppo, F.; Luo, L.; Fields, G.B.; Tummino, P.J.; Gontarek, R.R. Characterization of an exosite binding inhibitor of matrix metalloproteinase 13. Protein Sci. 2008, 17, 66–71. [Google Scholar]
- Nagase, H.; Kashiwagi, M. Aggrecanases and cartilage matrix degradation. Arthritis Res. Ther. 2003, 5, 94–103. [Google Scholar] [CrossRef]
- Lee, N.V.; Rodriguez-Manzaneque, J.C.; Thai, S.N.; Twal, W.O.; Luque, A.; Lyons, K.M.; Argraves, W.S.; Iruela-Arispe, M.L. Fibulin-1 acts as a cofactor for the matrix metalloprotease ADAMTS-1. J. Biol. Chem. 2005, 280, 34796–34804. [Google Scholar]
- Tortorella, M.D.; Malfait, F.; Barve, R.A.; Shieh, H.S.; Malfait, A.M. A review of the ADAMTS family, pharmaceutical targets of the future. Curr. Pharm. Des. 2009, 15, 2359–2374. [Google Scholar] [CrossRef]
- Vankemmelbeke, M.N.; Jones, G.C.; Fowles, C.; Ilic, M.Z.; Handley, C.J.; Day, A.J.; Knight, C.G.; Mort, J.S.; Buttle, D.J. Selective inhibition of ADAMTS-1, -4 and -5 by catechin gallate esters. Eur. J. Biochem. 2003, 270, 2394–2403. [Google Scholar] [CrossRef]
- Pasternak, B.; Aspenberg, P. Metalloproteinases and their inhibitors-diagnostic and therapeutic opportunities in orthopedics. Acta Orthop. 2009, 80, 693–703. [Google Scholar] [CrossRef]
- Li, X.Y.; Takasaki, C.; Satoh, Y.; Kimura, S.; Yasumoto, K.; Sogawa, K. Expression, purification and characterization of human PHD1 in Escherichia coli. J. Biochem. 2008, 144, 555–561. [Google Scholar] [CrossRef]
- Lind, T.; Birch, M.A.; McKie, N. Purification of an insect derived recombinant human ADAMTS-1 reveals novel gelatin (type I collagen) degrading activities. Mol. Cell. Biochem. 2006, 281, 95–102. [Google Scholar] [CrossRef]
- Takasugi, M.; Ishikawa, S.; Nagao, S.; Masamune, T. Studies on phytoalexins of the moraceae. Part 12. Albafuran-C, a natural diels-alder adduct of a dehydroprenyl-2-phenylbenzofuran with a chalcone from mulberry. Chem. Lett. 1982, 8, 1223–1224. [Google Scholar]
- Hano, Y.; Takizawa, S.; Mizuno, E.; Nomura, T. Structure of kuwanon-P, a new diels-alder type adduct from the Root bark of the cultivated mulberry tree (morus-lhou (ser) koidz). Chem. Pharm. Bull. 1983, 31, 2936–2939. [Google Scholar] [CrossRef]
- Hirakura, K.; Hano, Y.; Fukai, T.; Nomura, T.; Uzawa, J.; Fukushima, K. Constituents of the cultivated mulberry tree. 21. Structures of 3 new natural diels-alder type adducts, kuwanon-P and kuwanon-X, and mulberrofuran-J, from the cultivated mulberry tree (morus-lhou koidz). Chem. Pharm. Bull. 1985, 33, 1088–1096. [Google Scholar] [CrossRef]
- Dai, S.J.; Mi, Z.M.; Ma, Z.B.; Li, S.; Chen, R.Y.; Yu, D.Q. Bioactive diels-alder type adducts from the stem bark of morus macroura. Planta Med. 2004, 70, 758–763. [Google Scholar] [CrossRef]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A Simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Qin, Y.; Lv, X.Q.; Wang, P.; Xu, T.Y.; Zhang, L.; Miao, C.Y. A fluorometric assay for high-throughput screening targeting nicotinamide phosphoribosyltransferase. Anal. Biochem. 2011, 412, 18–25. [Google Scholar]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peng, J.; Gong, L.; Si, K.; Bai, X.; Du, G. Fluorescence Resonance Energy Transfer Assay for High-Throughput Screening of ADAMTS1 Inhibitors. Molecules 2011, 16, 10709-10721. https://doi.org/10.3390/molecules161210709
Peng J, Gong L, Si K, Bai X, Du G. Fluorescence Resonance Energy Transfer Assay for High-Throughput Screening of ADAMTS1 Inhibitors. Molecules. 2011; 16(12):10709-10721. https://doi.org/10.3390/molecules161210709
Chicago/Turabian StylePeng, Jianhao, Lili Gong, Kun Si, Xiaoyu Bai, and Guanhua Du. 2011. "Fluorescence Resonance Energy Transfer Assay for High-Throughput Screening of ADAMTS1 Inhibitors" Molecules 16, no. 12: 10709-10721. https://doi.org/10.3390/molecules161210709
APA StylePeng, J., Gong, L., Si, K., Bai, X., & Du, G. (2011). Fluorescence Resonance Energy Transfer Assay for High-Throughput Screening of ADAMTS1 Inhibitors. Molecules, 16(12), 10709-10721. https://doi.org/10.3390/molecules161210709