Protective Effects of Commiphora erythraea Resin Constituents Against Cellular Oxidative Damage


Abstract
:1. Introduction
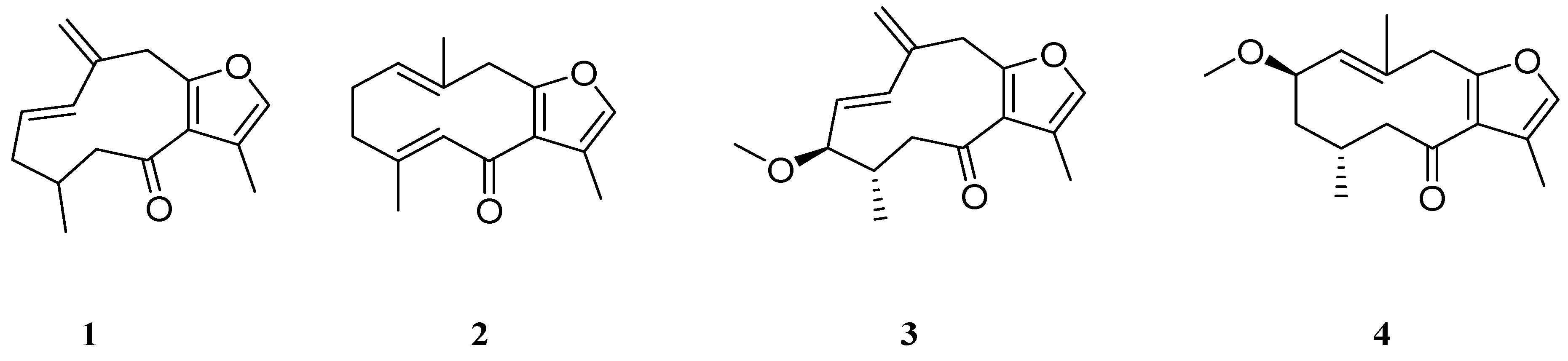
2. Results and Discussion
2.1. Lipid Peroxidation Inhibition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Lipid peroxidation inhibition a IC50 (μg/mL) | Lipid peroxidation inhibition a IC50 (μM) |
|---|---|---|
| HD b | 15.120 ± 0.520 | = |
| D c | 14.258 ± 1.560 | = |
| SF1 d | 17.893 ± 1.090 | = |
| SF2 e | 23.757 ± 4.860 | = |
| H f | 0.743 ± 0.040 | = |
| H1 | 56.848 ± 8.360 | = |
| H2 | 1.365 ± 0.200 | = |
| H3 | 0.756 ± 0.060 | = |
| 1 | 0.091 ± 0.003 | |
| 2 | 0.087 ± 0.001 | |
| 3 | 3.381 ± 0.079 | |
| 4 | 3.263 ± 0.057 | |
| BHT | 3.860 ± 0.850 | 17.517 ± 0.850 |
| Caffeic acid | 5.764 ± 0.480 | 31.993 ± 0.480 |
| Ascorbic acid | 18.630 ± 1.310 | 105.780 ±1.310 |
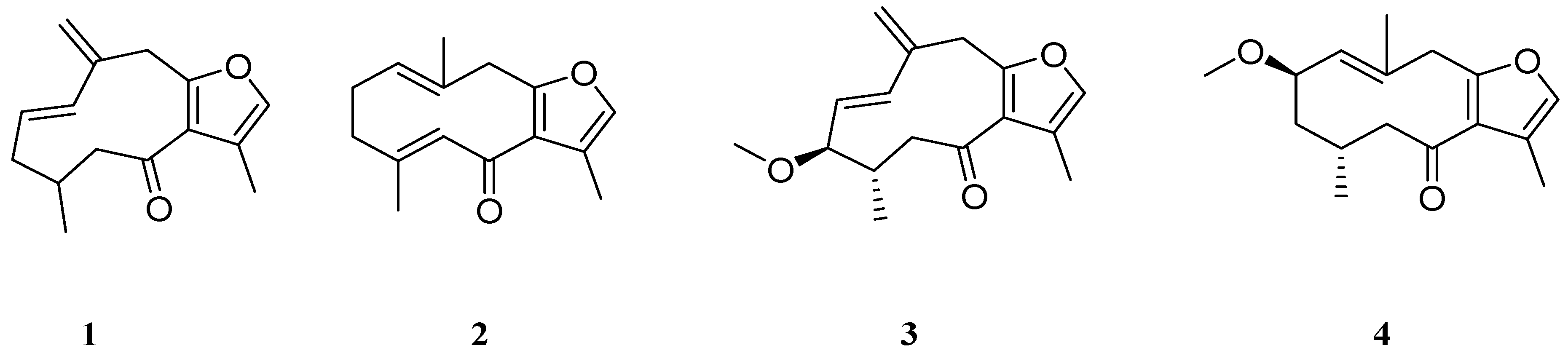
2.2. Inhibition of RNS formation
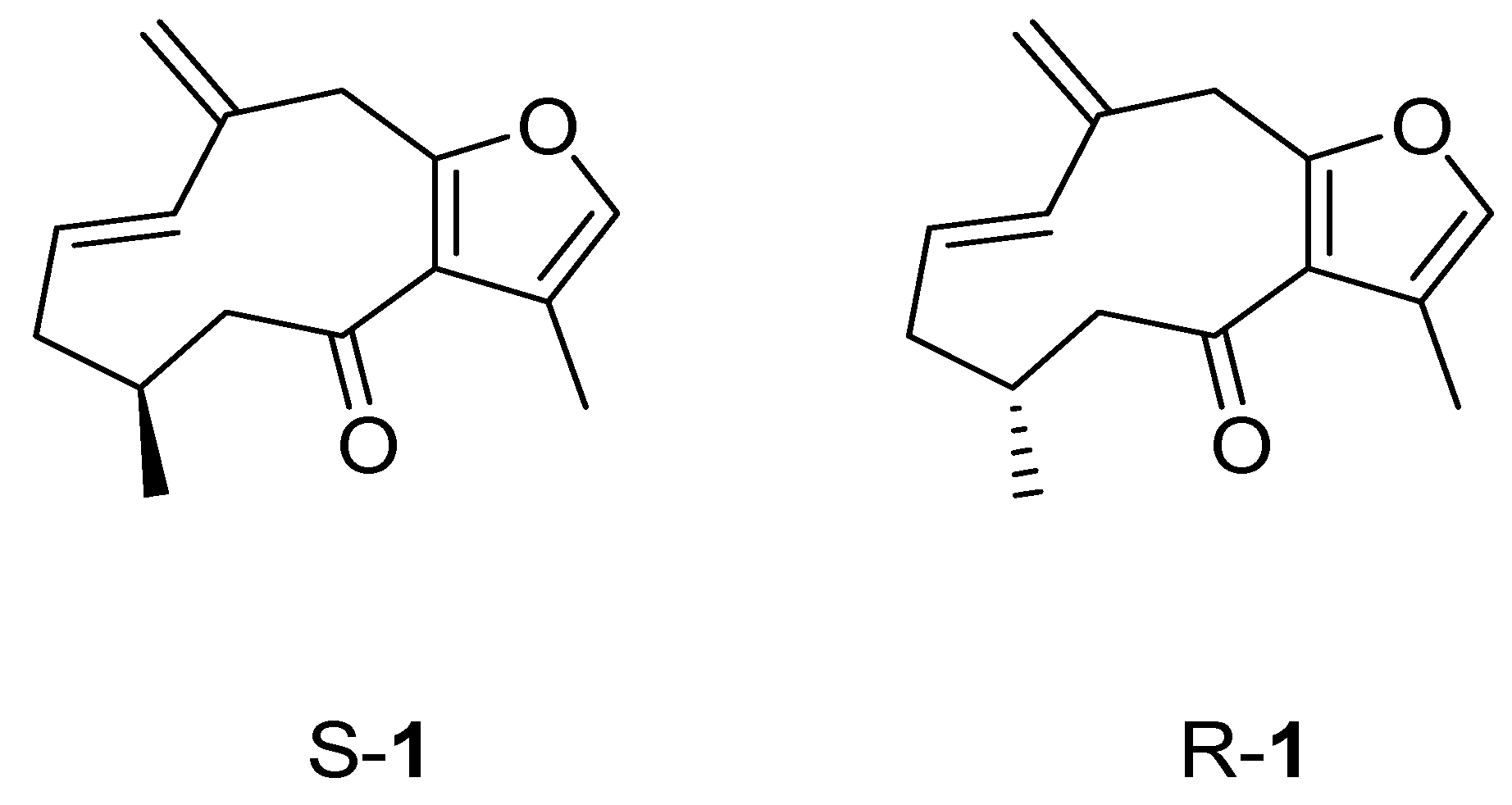
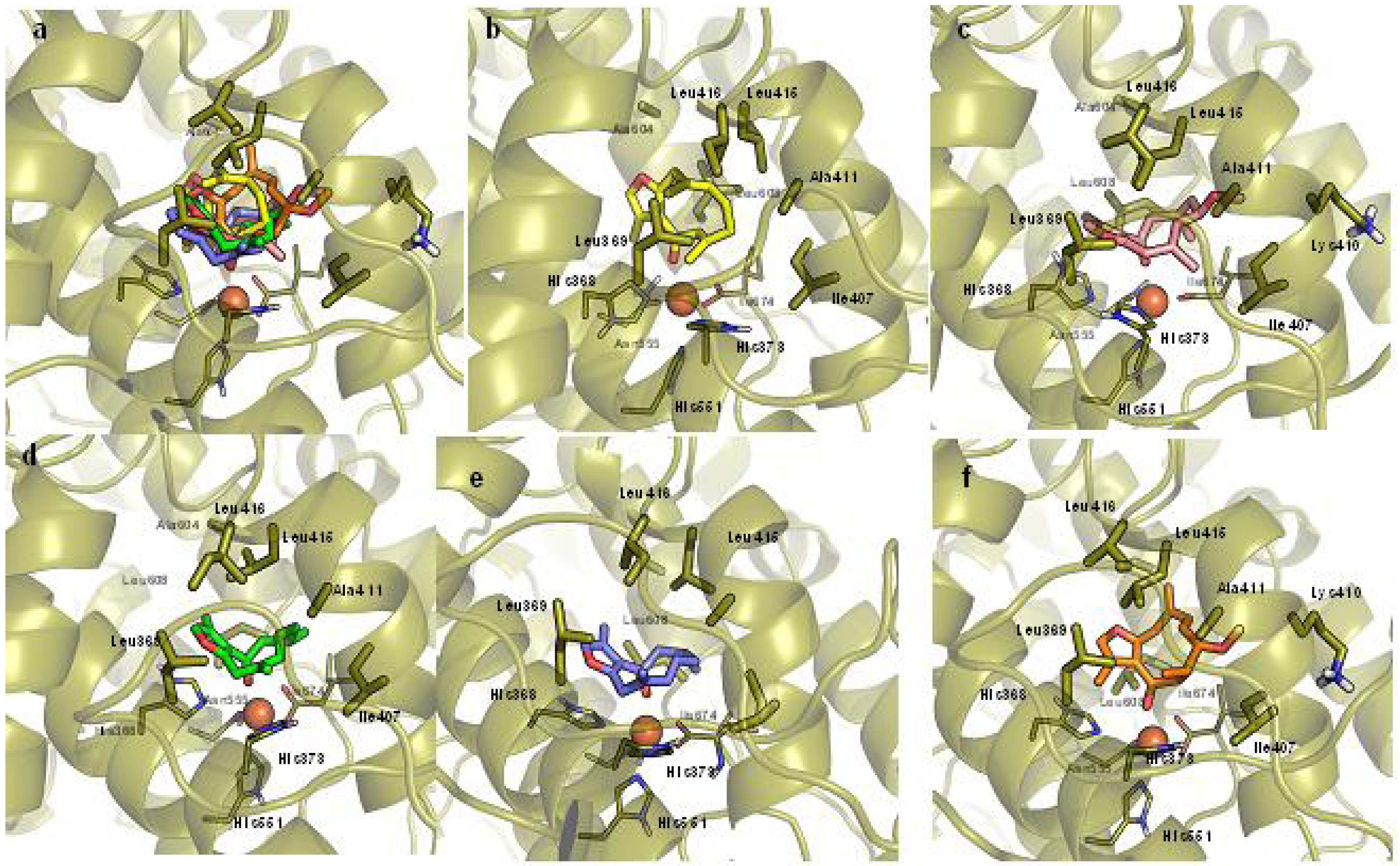
2.3. Molecular Docking
| Code | IC50 (μM) | Glide Score (kcal/mol) | IFD Score |
|---|---|---|---|
| S-1 | 0.091 ± 0.003 a | −8.01 | −1067.53 |
| R-1 | 0.091 ± 0.003 a | −7.73 | −756.54 |
| 2 | 0.087 ± 0.001 | −9.39 | −757.63 |
| 3 | 3.381 ± 0.079 | −9.55 | −1094.74 |
| 4 | 3.263 ± 0.057 | −8.59 | −789.2 |
| Code | Global Minimum(Kcal/mol) | Bioactive Conformation (Kcal/mol) | Conformational Gap Energy (Kcal/mol) |
|---|---|---|---|
| S-1 | 45.88 | 46.53 | +0.65 |
| R-1 | 45.88 | 49.69 | +3.81 |
| 2 | 66.98 | 66.99 | +0.01 |
| 3 | 41.99 | 53.53 | +11.54 |
| 4 | 47.34 | 50.45 | +3.11 |
3. Experimental
3.1. General Procedures
3.2. Material
3.3. Extraction, Isolation and Identification of Tested Compounds
3.4. Determination of Lipid Peroxidation Inhibitory Effect
3.5. Cell Cultures and Viability
3.6. Measurement of NO Production
3.7. Statistical Analysis
3.8. Molecular Modelling
4. Conclusions
Supplementary Materials
Acknowledgements
References and Notes
- Niki, E. Lipid peroxidation: Physiological levels and dual biological effects. Free Radic. Biol. Med. 2009, 47, 469–484. [Google Scholar] [CrossRef]
- Rubbo, H.; O'Donnell, V. Nitric oxide, peroxynitrite and lipoxygenase in atherogenesis: Mechanistic insights. Toxicology 2005, 208, 305–317. [Google Scholar] [CrossRef]
- Funk, C.D. Leukotriene inflammatory mediators meet their match. Sci. Transl. Med. 2011, 3, 66–ps3. [Google Scholar] [CrossRef]
- Manev, H.; Uz, T.; Sugaya, K.; Qu, T. Putative role of neuronal 5-lipoxygenase in an aging brain. FASEB J. 2000, 14, 1464–1469. [Google Scholar] [CrossRef]
- Manev, H.; Uz, T.; Manev, R.; Zhang, Z. Neurogenesis and neuroprotection in the adult brain. A putative role for 5-lipoxygenase? Ann. NY Acad. Sci. 2001, 939, 45–51. [Google Scholar]
- Rubin, P.; Mollison, K.W. Pharmacotherapy of diseases mediated by 5-lipoxygenase pathway eicosanoids. Prostag. Lipid Mediat. 2007, 83, 188–197. [Google Scholar] [CrossRef]
- Koeners, M.P.; Wesseling, S.; Ulu, A.; López Sepúlveda, R.; Morisseau, C.; Braam, B.; Hammock, B.D.; Joles, J.A. Soluble epoxide hydrolase in the generation and maintenance of high blood pressure in spontaneusly hypertensive rats. Am. J. Physiol. Endocrinol. Metab. 2011, 300, 691–698. [Google Scholar] [CrossRef]
- Puccio, S.; Chu, J.; Praticò, D. Involvement of 5-lipoxygenase in the corticosteroid-dependent amyloid beta formation: In vitro and in vivo evidence. PLoS One 2011, 6, e15163. [Google Scholar]
- Al-Shabrawey, M.; Mussell, R.; Kahook, K.; Tawfik, A.; Eladl, M.; Sarthy, V.; Nussbaum, J.; El-Marakby, A.; Park, S.Y.; Gurel, Z.; et al. Increased expression and activity of 12-lipoxygenase in oxygen-induced ischemic retinopathy and proliferative diabetic retinopathy implications in retinal neovascolarization. Diabetes 2011, 60, 614–624. [Google Scholar]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Jiang, Z.D.; Ketchum, S.O.; Gerwick, W.H. 5-Lipoxygenase-derived oxylipins from the red alga Rhodymenia pertusa. Phytochemistry 2000, 53, 129–133. [Google Scholar]
- Benrezzouk, R.; Terencio, M.C.; Ferrandiz, M.L; Hernandez-Perez, M.; Rabanal, R.; Alcaraz, M.J. Inhibition of 5-lipoxygenase activity by the natural antiinflammatory compound aethiopinone. Inflammation Res. 2001, 50, 96–101. [Google Scholar] [CrossRef]
- Amagata, T.; Whitman, S.; Johnson, T.A.; Stessman, C.C.; Loo, C.P.; Lobkovsky, E.; Clardy, J.; Crews, P.; Holman, T.R. Exploring sponge-derived terpenoids for their potency and selectivity against 12-human, 15-human, and 15-soybean lipoxygenase. J. Nat. Prod. 2003, 66, 230–235. [Google Scholar] [CrossRef]
- Paraskeva, M.P.; van Vuuren, S.F.; van Zyl, R.L.; Davids, H.; Viljoen, A.M. The in vivo biological activity of selected South African Commiphora species. J. Ethnopharmacol. 2008, 119, 673–679. [Google Scholar] [CrossRef]
- Increasing People Opportunities. Available online: http://www.ipoassociazione.org/ (accessed on 13 December 2011).
- Zorloni, A. Evaluation of plants used for the control of animal ectoparasitoses in Southern Ethiopia (Oromiya and Somali regions). M.Sc. Thesis., University of Pretoria, Pretoria, South Africa, 2007. [Google Scholar]
- Fraternale, D.; Sosa, S.; Ricci, D.; Genovese, S.; Messina, F.; Tomasini, S.; Montanari, F.; Marcotullio, M.C. Marcotullio, M.C. Anti-inflammatory, antioxidant and antifungal furanosesquiterpenoids isolated from Commiphora erythraea (Ehrenb.) Engl. resin. Fitoterapia 2011, 82, 654–661. [Google Scholar] [CrossRef]
- Racine, P.; Auffray, B. Quenching of singlet molecular oxygen by Commiphora myrrha extracts and menthofuran. Fitoterapia 2005, 76, 316–323. [Google Scholar] [CrossRef]
- Koshihara, Y.; Neichi, T.; Murata, S.; Lao, A.; Fujimoto, Y.; Tatsuno, T. Caffeic acid is a selective inhibitor for leukotriene biosynthesis. Biochim. Biophys. Acta 1984, 792, 92–97. [Google Scholar] [CrossRef]
- Dekebo, A.; Dagne, E.; Sterner, O. Furanosesquiterpenes from Commiphora sphaerocarpa and related adulterants of true myrrh. Fitoterapia 2002, 73, 48–55. [Google Scholar] [CrossRef]
- Dekebo, A.; Dagne, E.; Hansen, L.K.; Gauntun, O.R.; Aasen, A.J. Crystal structures of two furanosesquiterpenes from Commiphora sphaerocarpa. Tetrahedron Lett. 2000, 41, 9875–9878. [Google Scholar] [CrossRef]
- Zhu, N.; Kikuzaki, H.; Shen, S.; Sang, S.; Rafi, M.M.; Wang, M.; Nakatani, N.; Di Paola, R.S.; Rosen, R.T.; Ho, C.-T. Furanosesquiterpenoids of Commiphora myrrha. J. Nat. Prod. 2001, 64, 1460–1462. [Google Scholar] [CrossRef]
- Pawate, S.; Shen, Q.; Fan, F.; Bhat, N.R. Redox regulation of glial inflammatory response to lipopolysaccharide and interferongamma. J. Neurosci. Res. 2004, 77, 540–551. [Google Scholar] [CrossRef]
- Aparoy, P.; Reddy, R.N.; Guruprasad, L.; Reddy, M.R.; Reddanna, P. Homology modeling of 5-lipoxygenase and hints for better inhibitor design. J. Comput-Aided Mol. Des. 2008, 22, 611–619. [Google Scholar] [CrossRef]
- Hazai, E.; Bikádi, Z.; Zsila, F.; Lockwood, S.F. Molecular modeling of the non-covalent binding of the dietary tomato carotenoids lycopene and lycophyll, and selected oxidative metabolites with 5-lipoxygenase. Bioorg. Med. Chem. 2006, 14, 6859–6867. [Google Scholar] [CrossRef]
- Charlier, C.; Hénichart, J.P.; Durant, F.; Wouters, J. Structural insights into human 5-lipoxygenase inhibition: Combined ligand-based and target-based approach. J. Med. Chem. 2006, 49, 186–195. [Google Scholar] [CrossRef]
- Tanaka, K.; Kuba, Y.; Ina, A.; Watanabe, H.; Komatsu, K. Prediction of cyclooxygenase inhibitory activity of Curcuma rhizome from chromatograms by multivariate analysis. Chem. Pharm. Bull. 2008, 56, 936–940. [Google Scholar] [CrossRef]
- Sud'ina, G.F.; Mirzoeva, O.K.; Pushkareva, M.A.; Korshunova, G.A.; Sumbatyan, N.V.; Varfolomeev, S.D. Caffeic acid phenethyl ester as a lipoxygenase inhibitor with antioxidant properties. FEBS Lett. 1993, 329, 21–24. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL Workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef]
- The UniProt Consortium. The Universal Protein Resource (UniProt) in 2010. Nucleic Acid. Res. 2010, 38, D142–D148. [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acid Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Oldham, M.L.; Brash, A.R.; Newcomer, M.E. Insights from the X-ray crystal structure of coral 8R-lipoxygenase: calcium activation via a C2-like domain and a structural basis of product chirality. J. Biol. Chem. 2005, 280, 39545–39552. [Google Scholar]
- Schrödinger Suite 2010. LLC: New York, NY, USA.
- Jorgensen, W.L.; Tirado-Rives, J. The OPLS Force Field for Proteins. Energy minimizations for crystals of cyclic peptides and crambin. J. Am. Chem. Soc. 1988, 110, 1657–1666. [Google Scholar] [CrossRef]
- Lüthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of protein models with three-dimensional profiles. Nature 1992, 356, 83–85. [Google Scholar]
- Chen, V.B.; Arendall, W.B., 3rd.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D. Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef]
- Still, W.C.; Tempczyk, A.; Hawley, R.C.; Hendrickson, T. Semianalytical treatment of solvation for molecular mechanics and dynamics. J. Am. Chem. Soc. 1990, 112, 6127–6129. [Google Scholar] [CrossRef]
- Saunders, M.; Houk, K.N.; Wu, Y.D.; Still, W.C.; Lipton, M.; Chang, G.; Guida, W.C. Conformations of cycloheptadecane. A comparison of methods for conformational searching. J. Am. Chem. Soc. 1990, 112, 1419–1427. [Google Scholar]
- Ponder, J.W.; Richards, F.M. An efficient Newton-like method for molecular mechanics energy minimization of large molecules. J. Comput. Chem. 1987, 8, 1016–1024. [Google Scholar] [CrossRef]
- InfarmaZone Onlus. Available online: http://www.infarmazone.org/wp/ (accessed on 13 December 2011).
- Sample Availability: Samples of the compounds 1, 2, 3 and 4 are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Marcotullio, M.C.; Messina, F.; Curini, M.; Macchiarulo, A.; Cellanetti, M.; Ricci, D.; Giamperi, L.; Bucchini, A.; Minelli, A.; Mierla, A.L.; et al. Protective Effects of Commiphora erythraea Resin Constituents Against Cellular Oxidative Damage. Molecules 2011, 16, 10357-10369. https://doi.org/10.3390/molecules161210357
Marcotullio MC, Messina F, Curini M, Macchiarulo A, Cellanetti M, Ricci D, Giamperi L, Bucchini A, Minelli A, Mierla AL, et al. Protective Effects of Commiphora erythraea Resin Constituents Against Cellular Oxidative Damage. Molecules. 2011; 16(12):10357-10369. https://doi.org/10.3390/molecules161210357
Chicago/Turabian StyleMarcotullio, Maria Carla, Federica Messina, Massimo Curini, Antonio Macchiarulo, Marco Cellanetti, Donata Ricci, Laura Giamperi, Anahi Bucchini, Alba Minelli, Anna Lisa Mierla, and et al. 2011. "Protective Effects of Commiphora erythraea Resin Constituents Against Cellular Oxidative Damage" Molecules 16, no. 12: 10357-10369. https://doi.org/10.3390/molecules161210357
APA StyleMarcotullio, M. C., Messina, F., Curini, M., Macchiarulo, A., Cellanetti, M., Ricci, D., Giamperi, L., Bucchini, A., Minelli, A., Mierla, A. L., & Bellezza, I. (2011). Protective Effects of Commiphora erythraea Resin Constituents Against Cellular Oxidative Damage. Molecules, 16(12), 10357-10369. https://doi.org/10.3390/molecules161210357