Autotoxicity and Allelopathy of 3,4-Dihydroxyacetophenone Isolated from Picea schrenkiana Needles

 ,
,
Abstract
:1. Introduction
2. Results
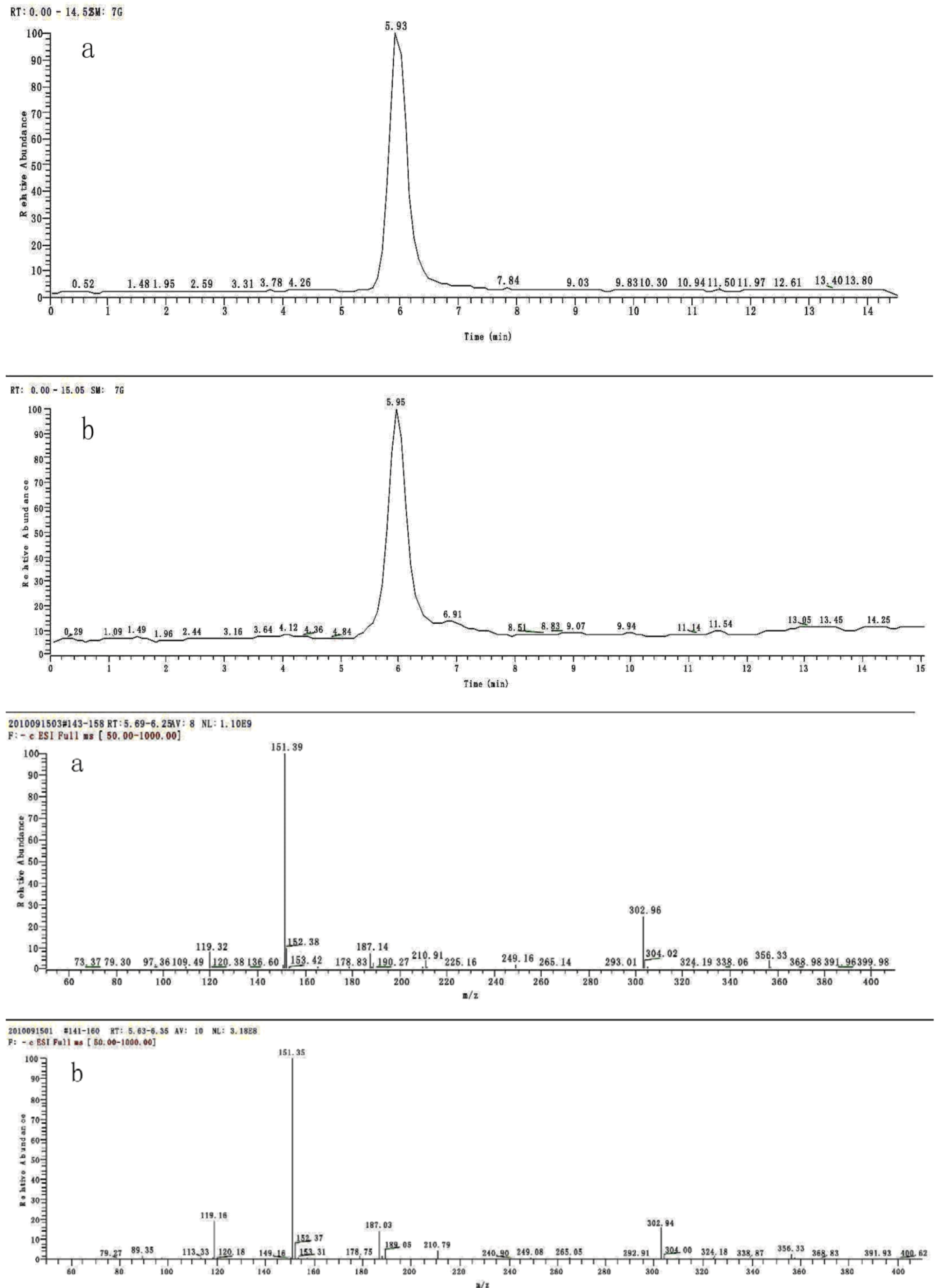
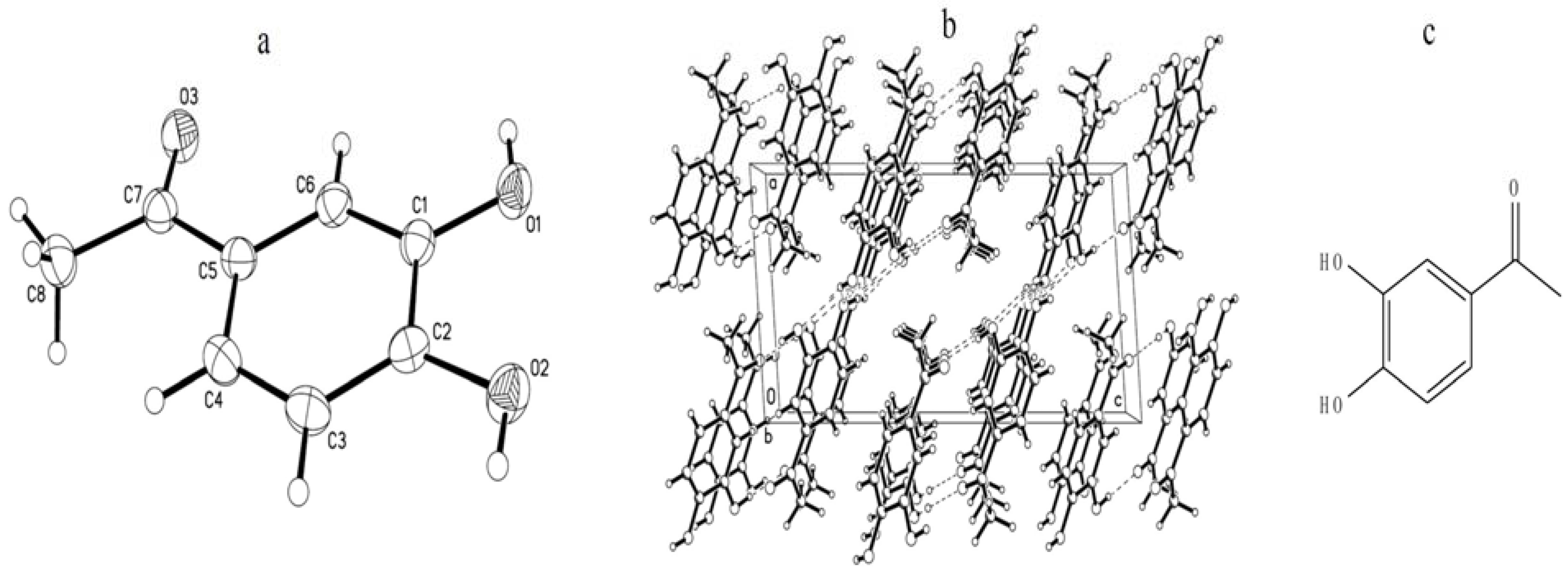
2.1. Identification and Quantitation Analysis of DHAP
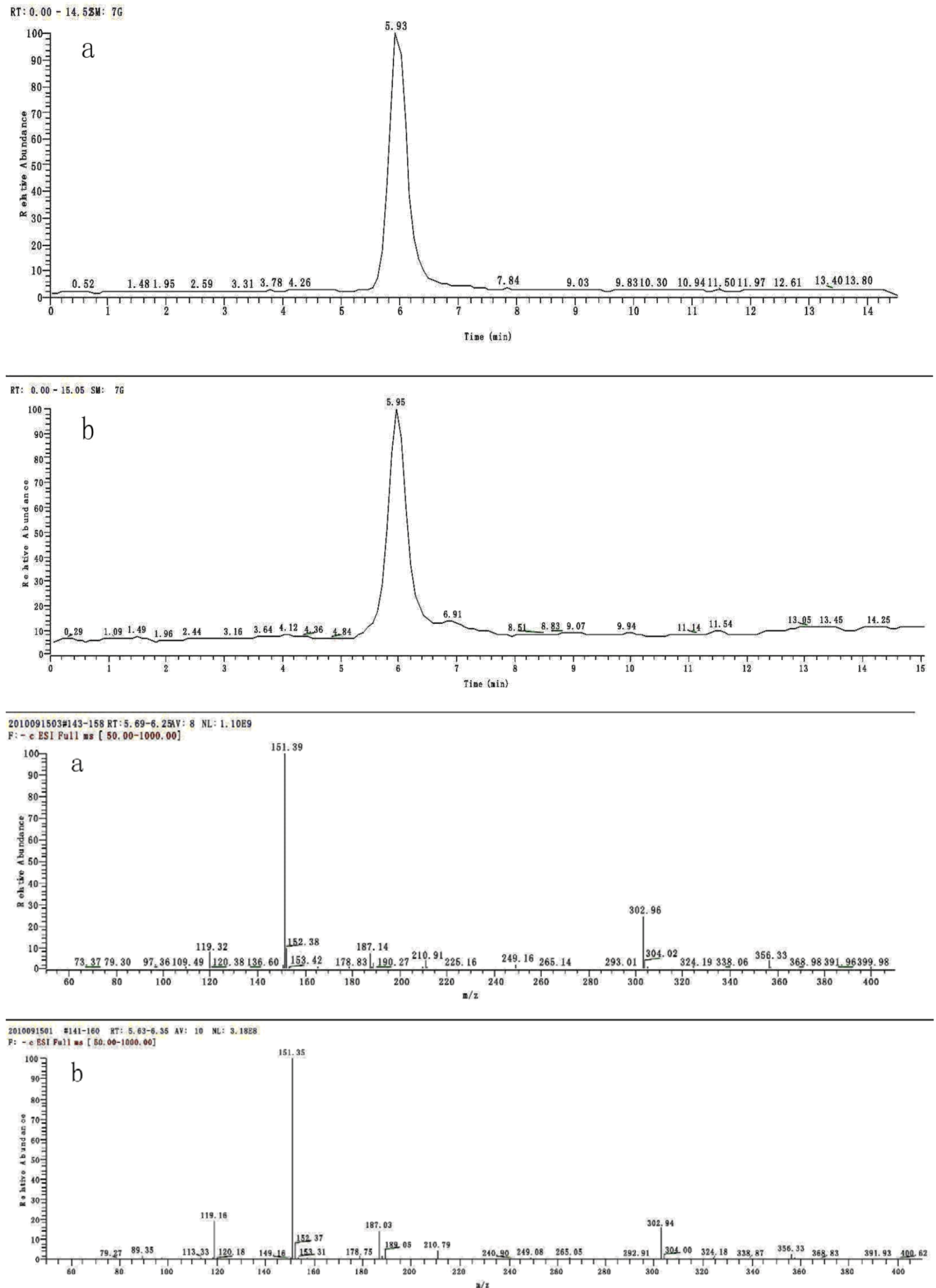
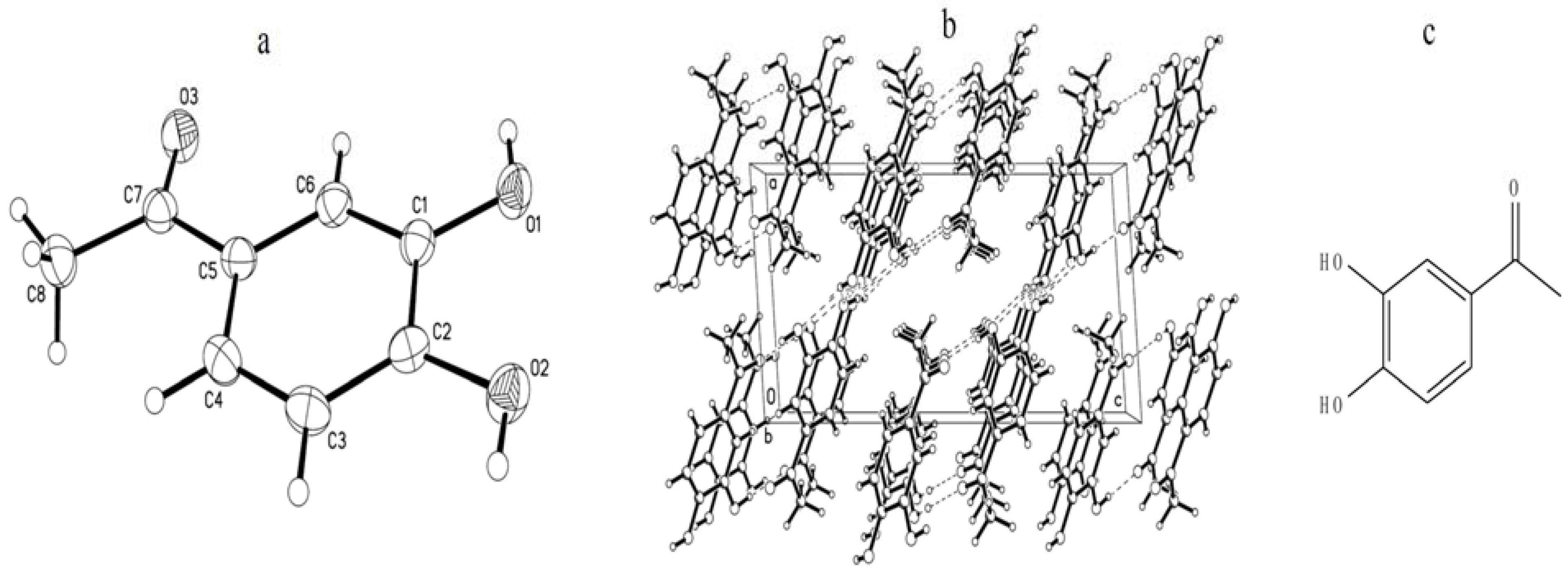
2.2. Effects of DHAP on Seed Germination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Concentration mM | Germination Rate (% of control) | Germination vigor (% of control) |
|---|---|---|---|
| P. schrenkiana | 0 | 100 a | 100 a |
| 0.1 | 98.13 ± 4.18 a | 91.39 ± 6.03 a | |
| 0.5 | 94.37 ± 6.25 a | 97.84 ± 1.53 a | |
| 1 | 96.24 ± 4.16 a | 86.03 ± 0.58 ab | |
| 2.5 | 83.56 ± 2.52 b | 83.87 ± 1.73 b | |
| 5 | 85.45 ± 7.02 b | 23.65 ± 0.58 c | |
| 10 | 74.18 ± 5.13 c | 0 ± 0 d | |
| Wheat | 0 | 100 a | 100 a |
| 0.1 | 99.33 ± 0.58 a | 98.66 ± 0.58 a | |
| 0.5 | 97.67 ± 0.58 ab | 96.65 ± 1.53 ab | |
| 1 | 95.67 ± 1.53 b | 92.30 ± 1.00 b | |
| 2.5 | 92.33 ± 4.93 cd | 90.30 ± 6.08 bc | |
| 5 | 94.00 ± 1.00 c | 91.30 ± 0 b | |
| 10 | 89.33 ± 2.08 d | 81.60 ± 5.51 c | |
| Lettuce | 0 | 100 a | 100 a |
| 0.1 | 98.08 ± 3.22 a | 88.10 ± 8.74 a | |
| 0.5 | 84.68 ± 0.58 b | 78.37 ± 4.51 bc | |
| 1 | 81.23 ± 1.07 b | 75.13 ± 8.51 c | |
| 2.5 | 81.23 ± 3.22 b | 71.35 ± 5.29 cd | |
| 5 | 78.93 ± 6.11 bc | 62.70 ± 4.62 de | |
| 10 | 63.60 ± 7.02 c | 42.16 ± 6.08 e | |
| Rice | 0 | 100 a | 100 a |
| 0.1 | 97.56 ± 1.86 a | 106.25 ± 2.65 c | |
| 0.5 | 97.06 ± 3.00 a | 112.50 ± 3.61 c | |
| 1 | 96.53 ± 4.36 a | 147.94 ± 0.55 b | |
| 2.5 | 97.76 ± 1.45 a | 131.25 ± 1.61 bc | |
| 5 | 96.76 ± 3.46 a | 104.19 ± 1.53 c | |
| 10 | 97.59 ± 3.00 a | 166.69 ± 2.08 a | |
| Radish | 0 | 100 a | 100 a |
| 0.1 | 98.97 ± 1.53 a | 96.83 ± 5.77 a | |
| 0.5 | 97.27 ± 0.58 a | 94.36 ± 2.08 a | |
| 1 | 97.60 ± 4.58 a | 97.18 ± 5.19 a | |
| 2.5 | 96.24 ± 2.31 a | 94.36 ± 3.22 a | |
| 5 | 100.00 ± 1.53 a | 97.53 ± 1.53 a | |
| 10 | 97.95 ± 1.53 a | 97.53 ± 0.58 a | |
| Cucumber | 0 | 100 a | 100 a |
| 0.1 | 96.10 ± 2.08 a | 96.83 ± 5.77 a | |
| 0.5 | 96.46 ± 4.51 a | 94.36 ± 2.08 a | |
| 1 | 98.59 ± 4.04 a | 97.18 ± 5.19 a | |
| 2.5 | 96.46 ± 2.52 a | 94.36 ± 3.22 a | |
| 5 | 97.52 ± 3.51 a | 97.53 ± 1.53 a | |
| 10 | 96.10 ± 4.51 a | 97.53 ± 0.58 a |
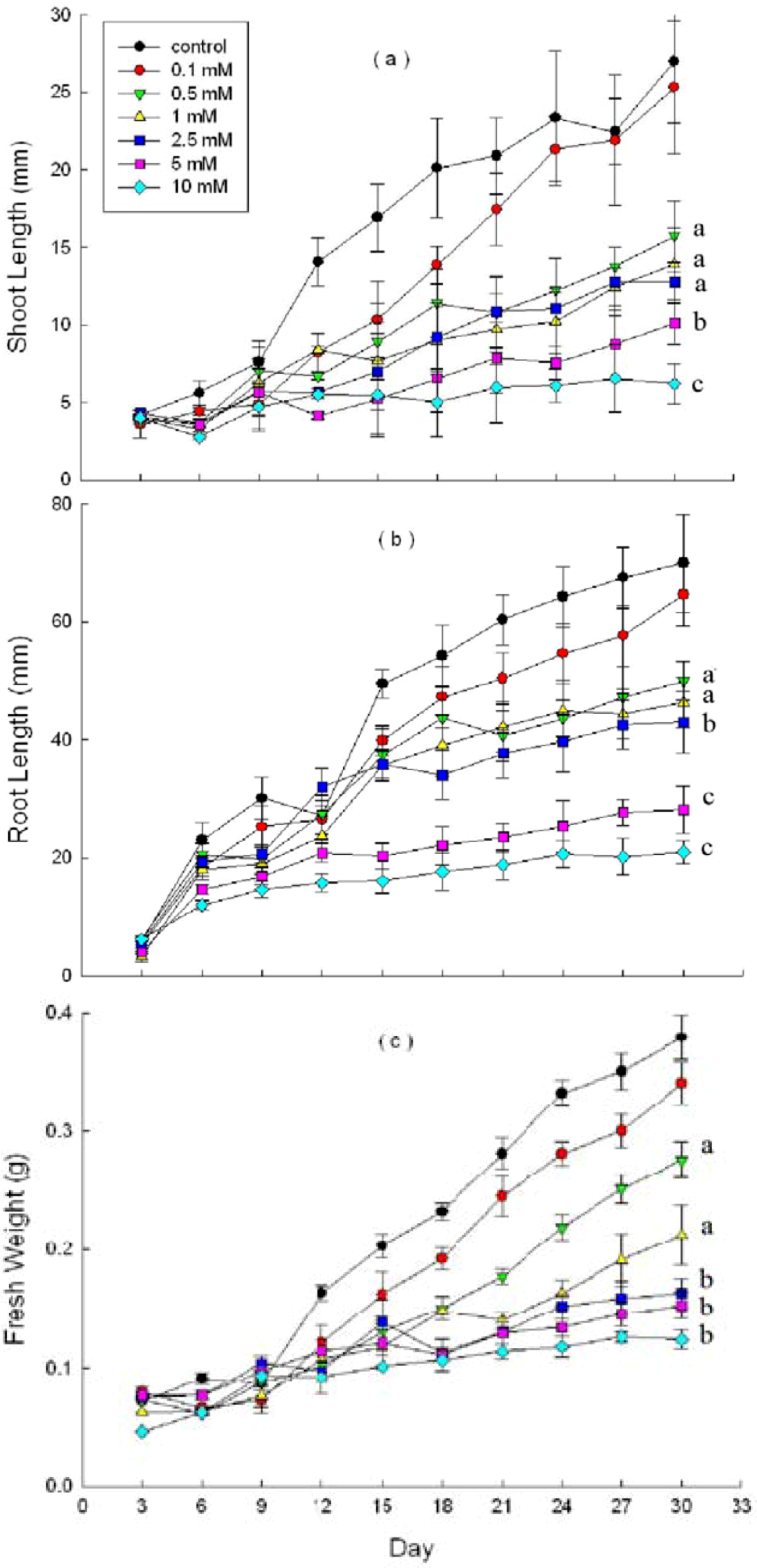
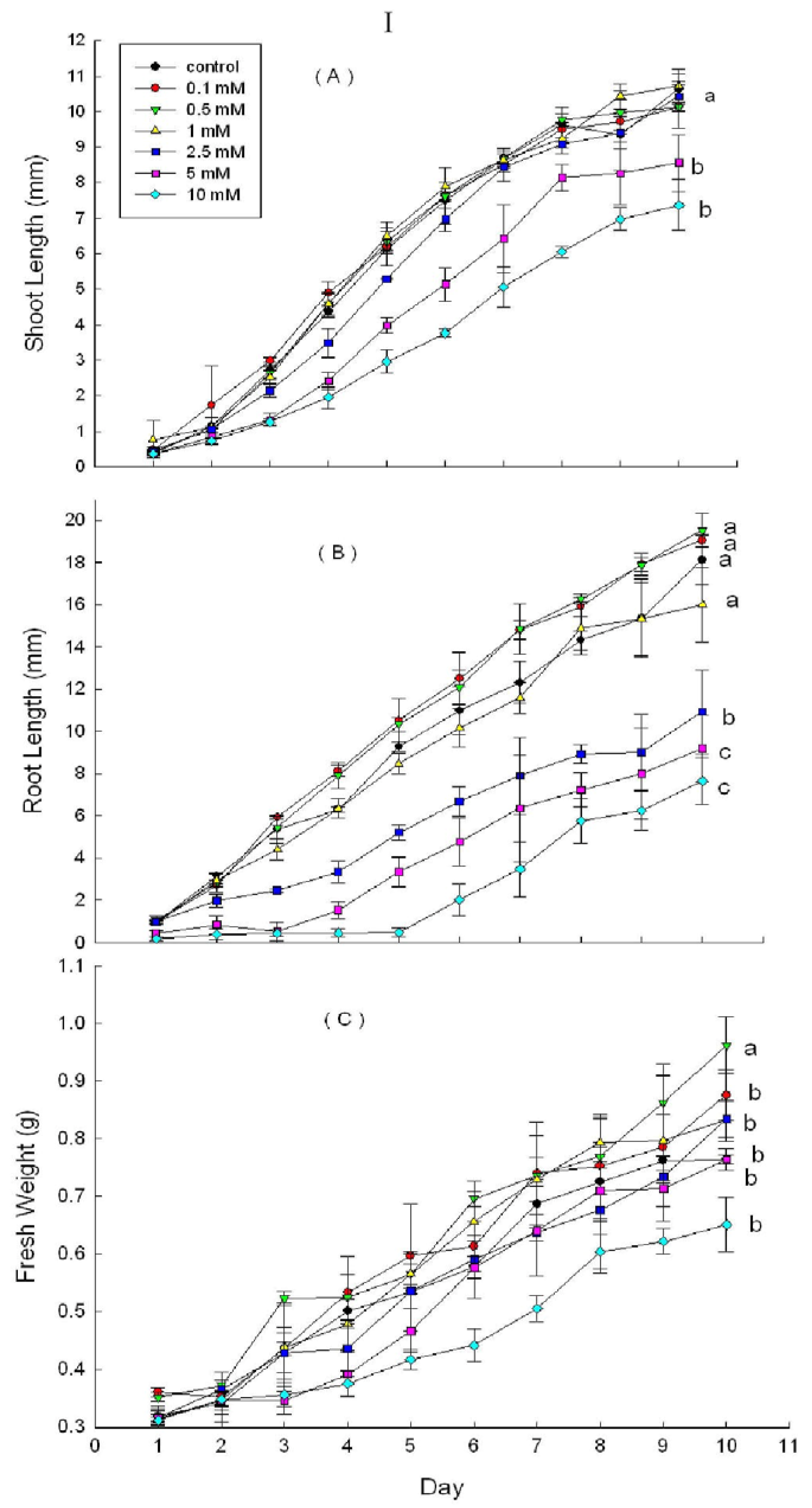
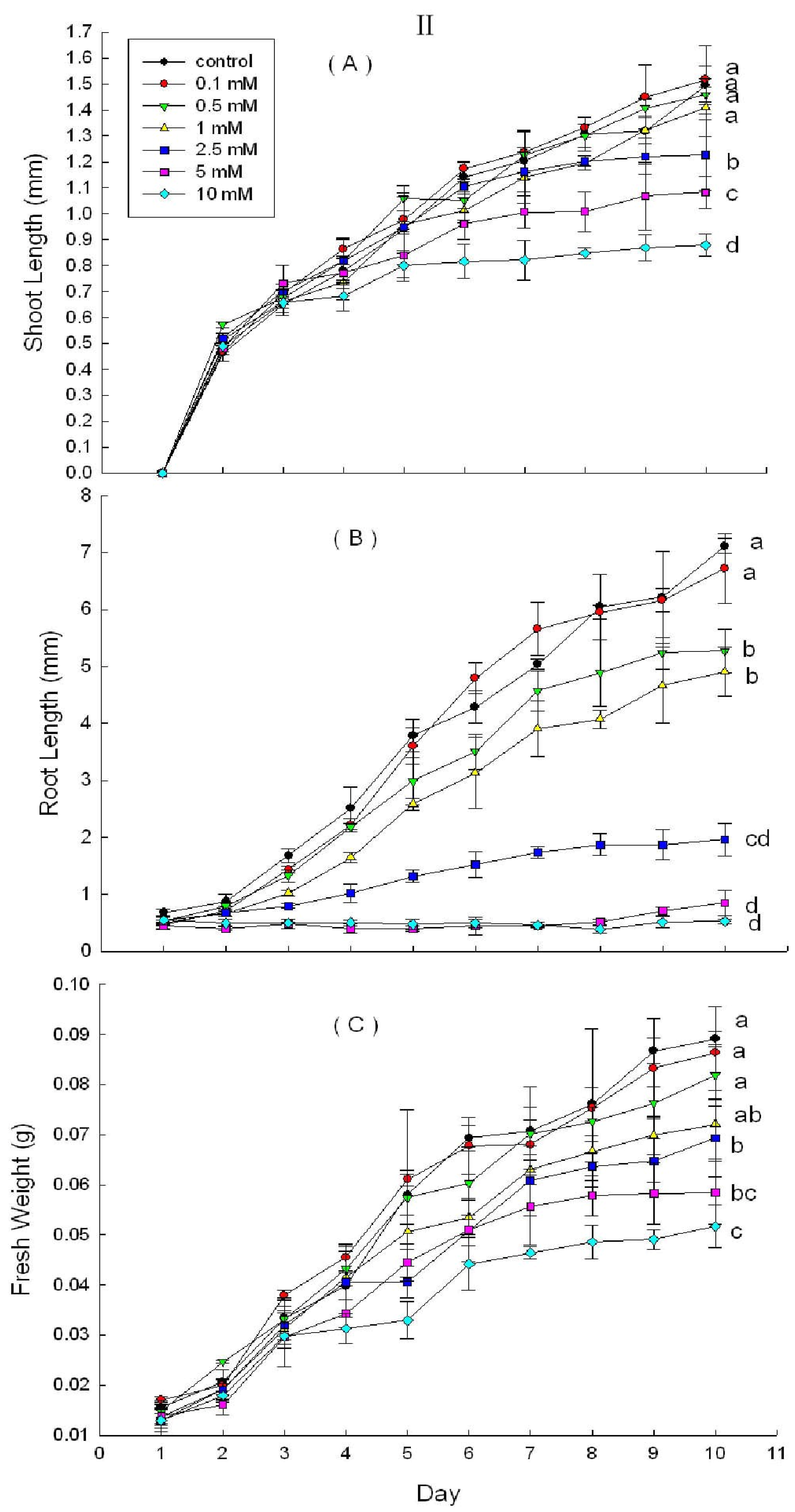
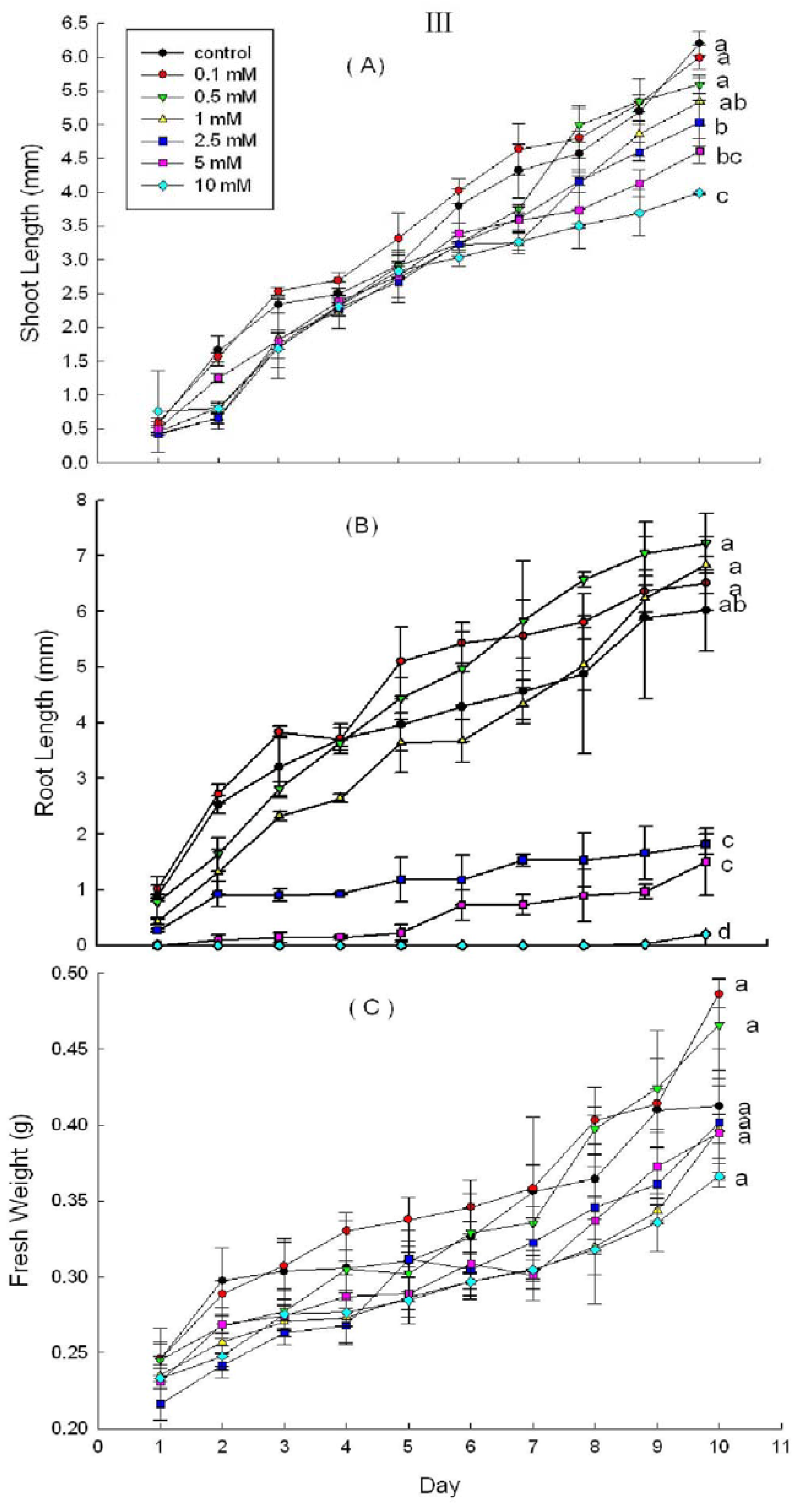
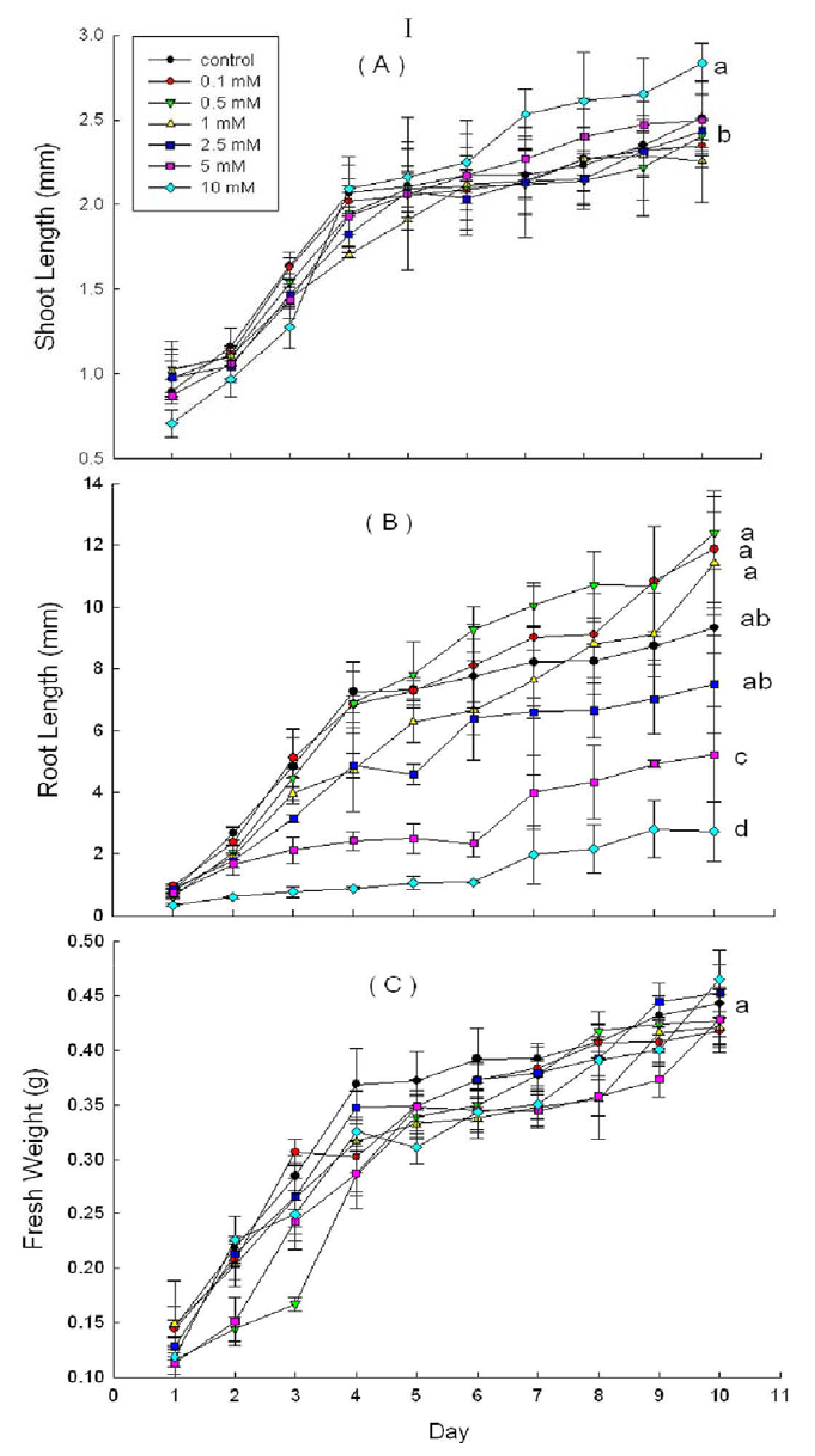
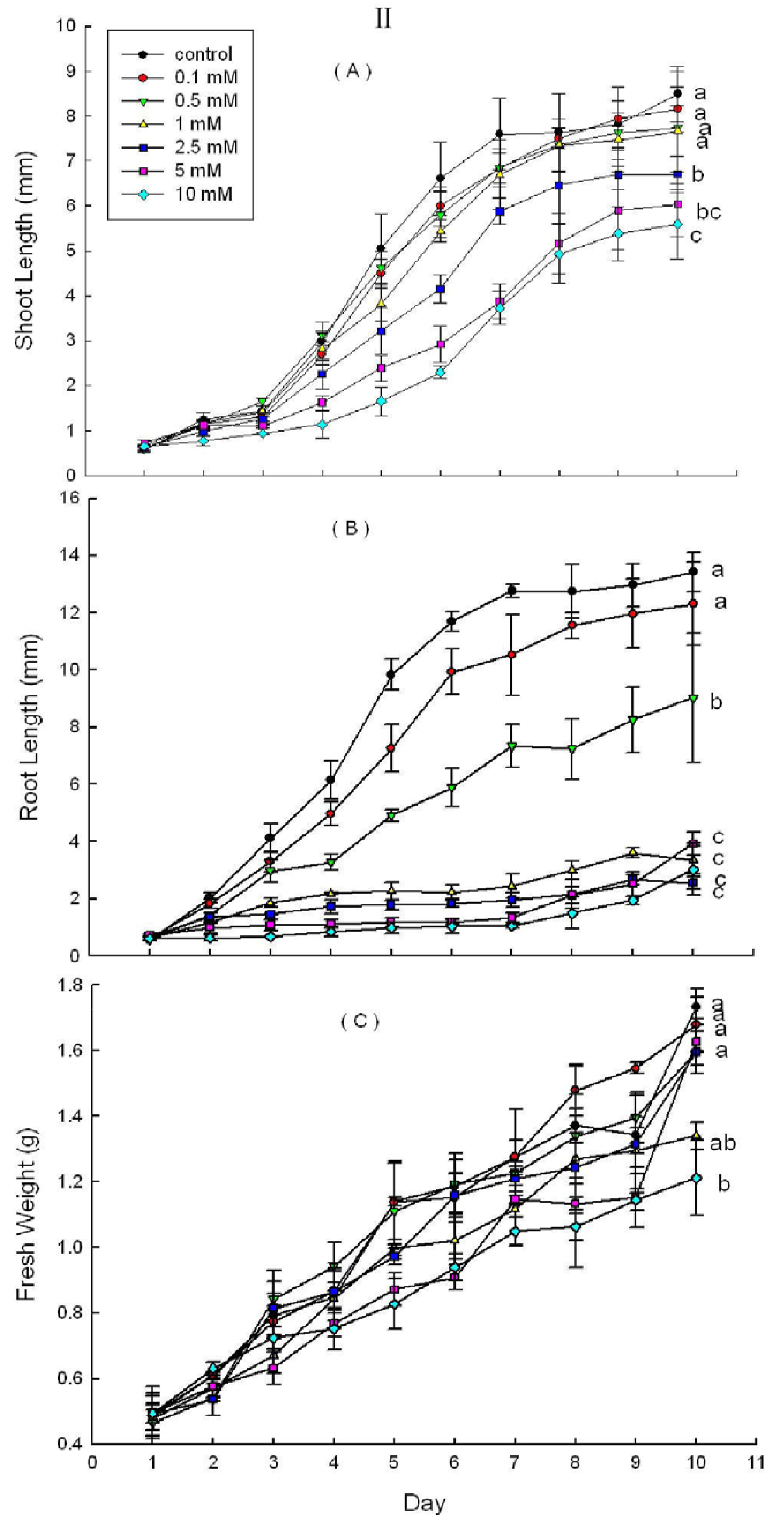
2.3. Effects of DHAP on Seedling Growth
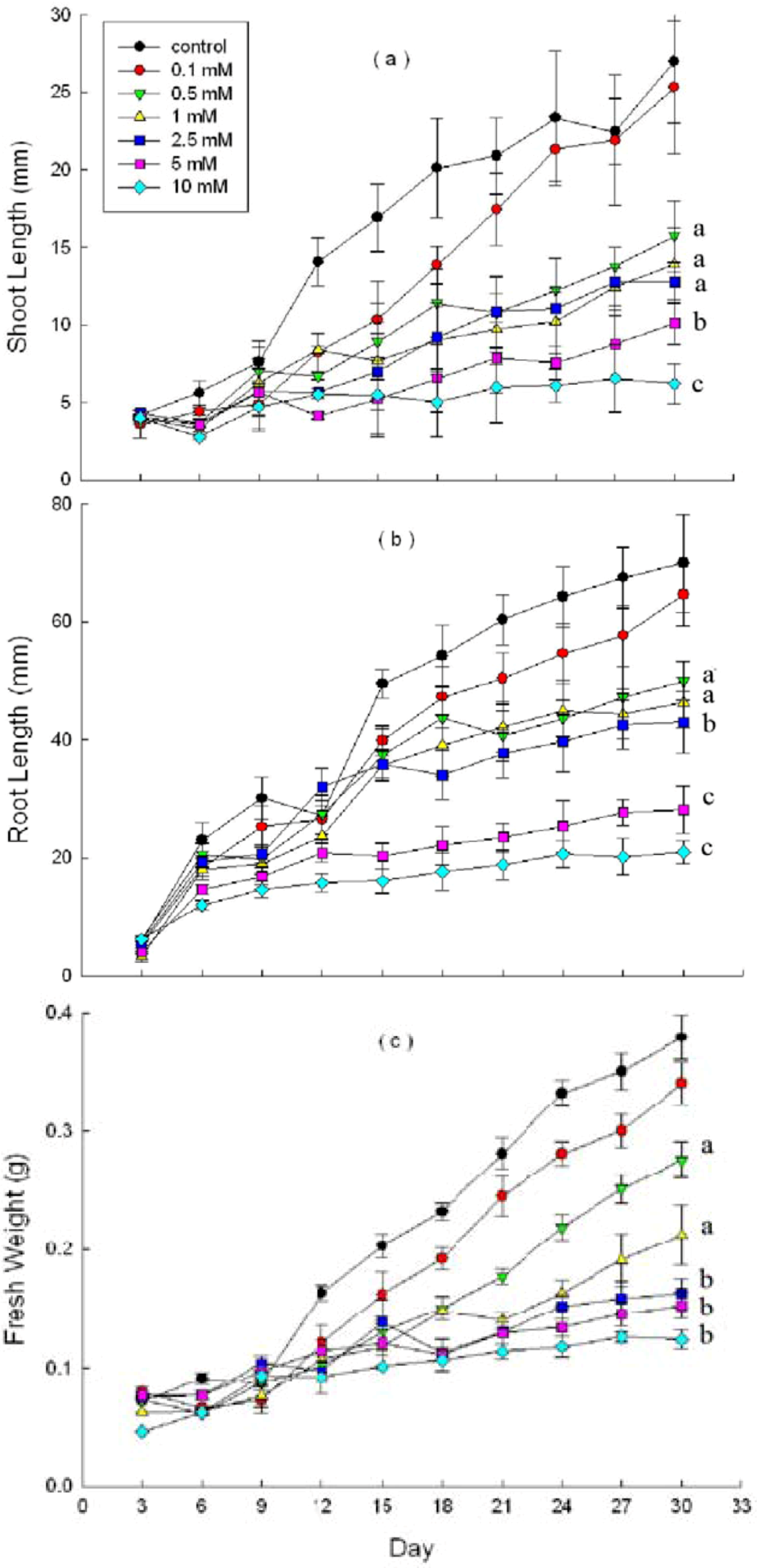


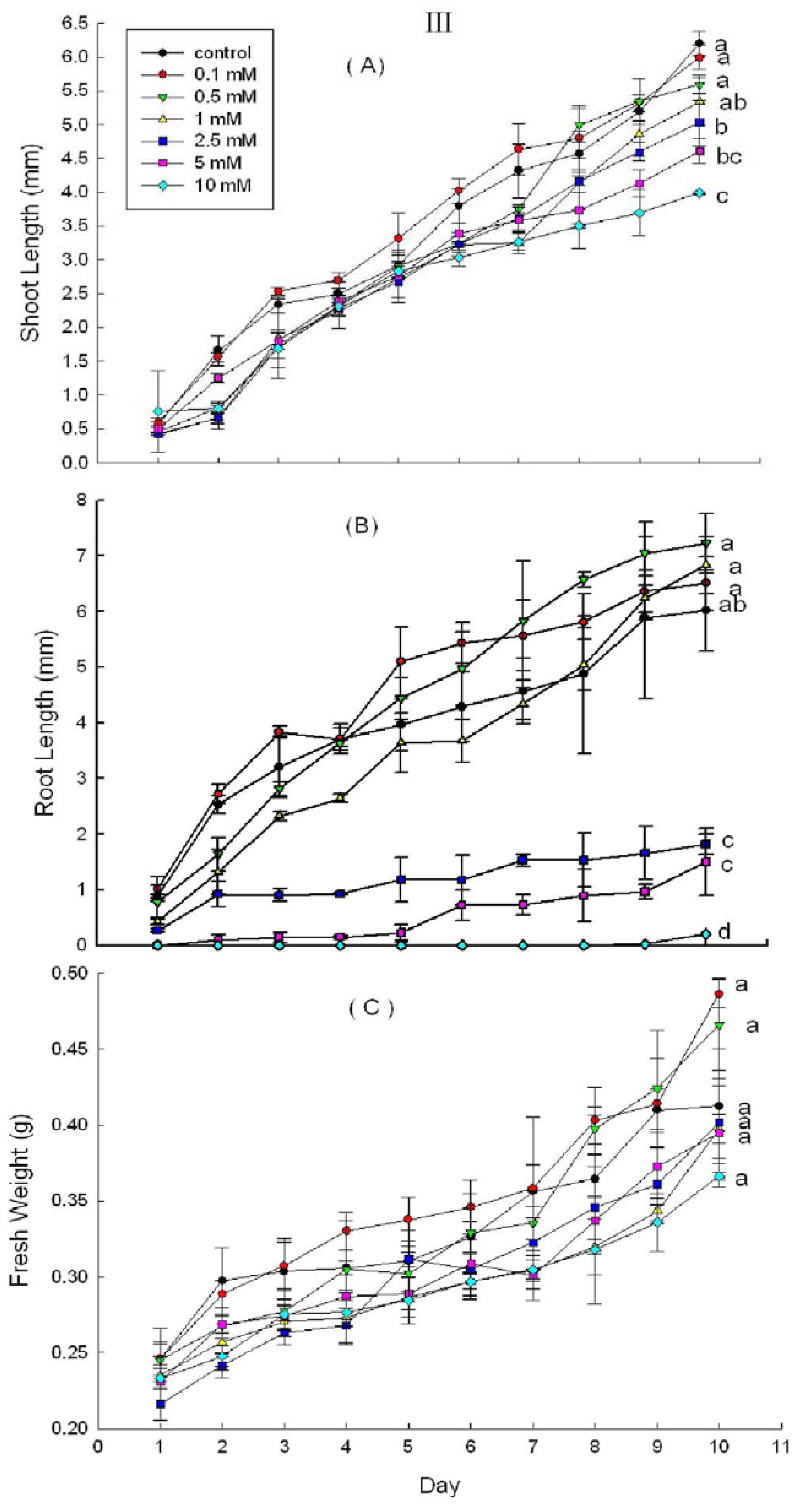
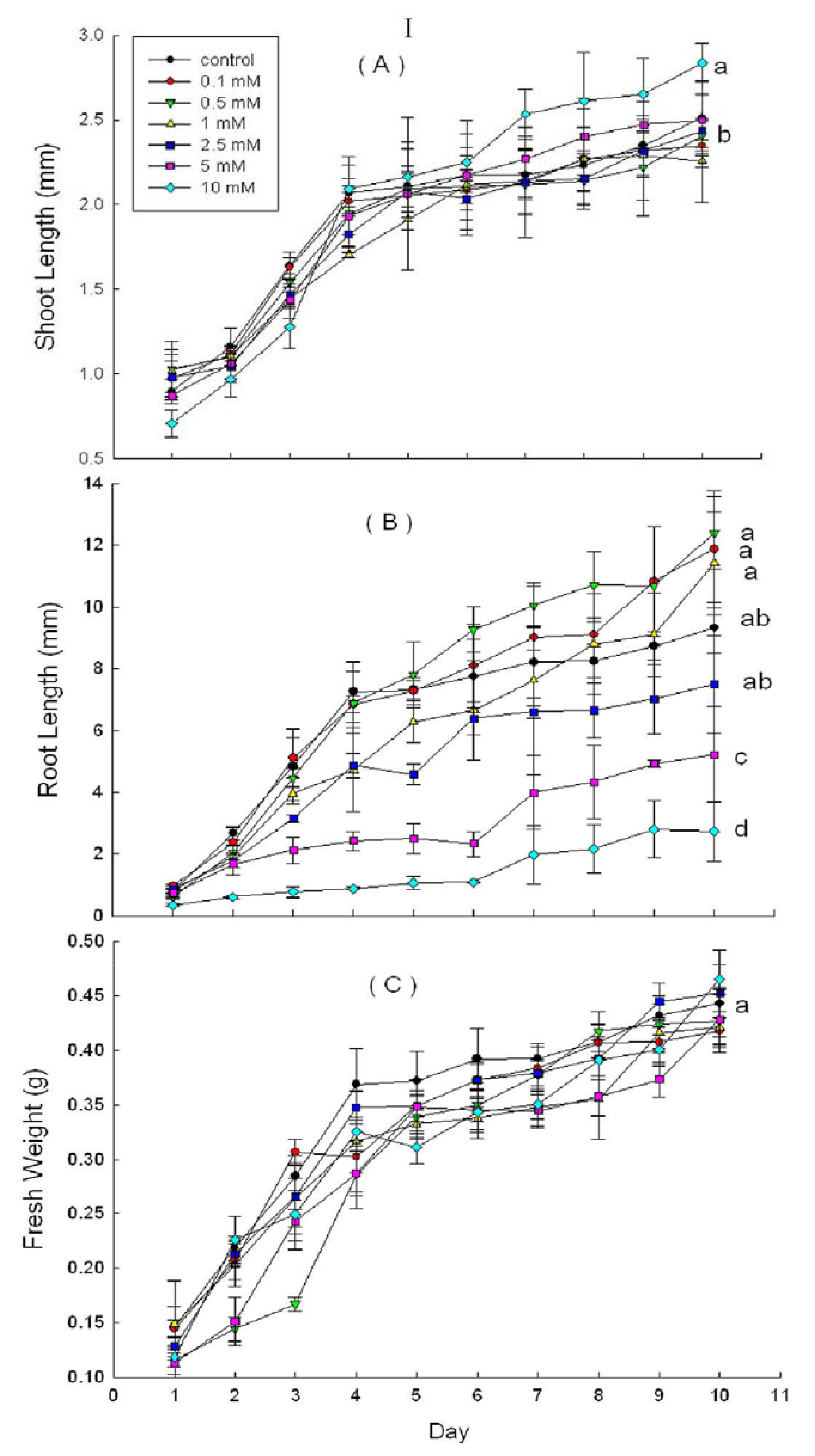
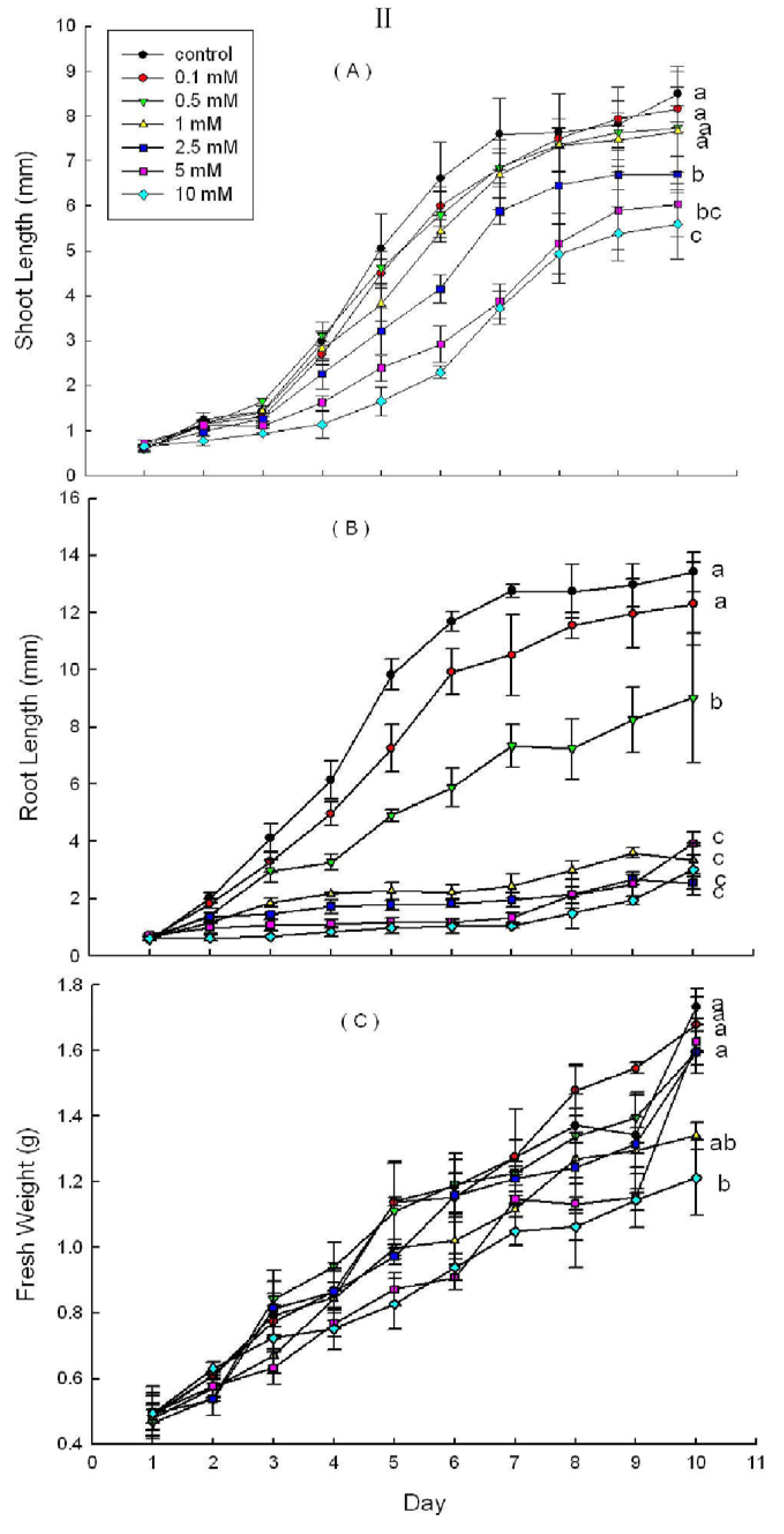
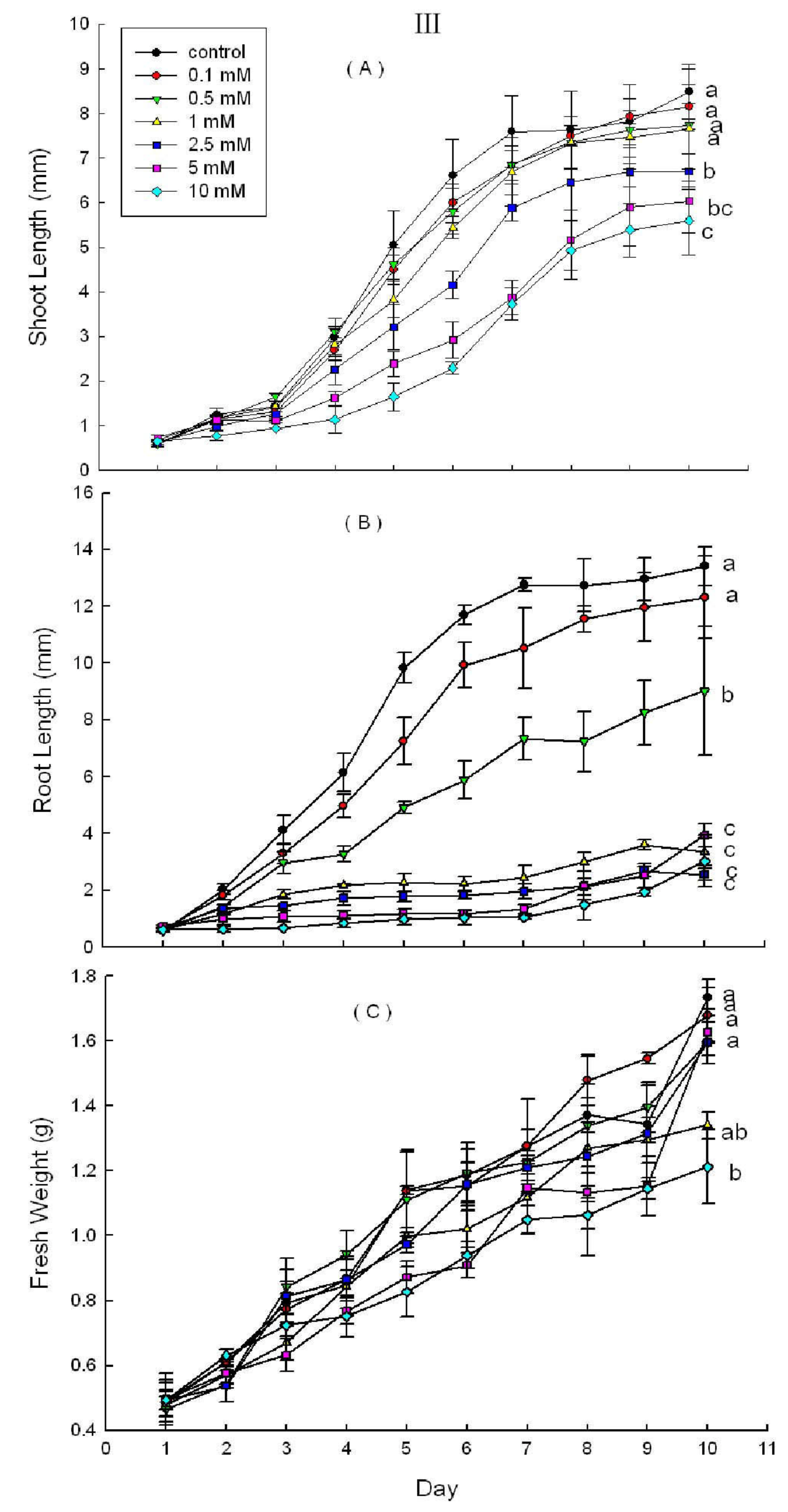
3. Discussion
4. Experimental
4.1. General
4.2. Plant Materials
4.3. Extraction and Isolation of the Active Compound
4.4. Bioassay
4.4.1. Effects of DHAP on Seed Germination
4.4.2. Effects of DHAP on Seedling Growth
4.5. Quantify DHAP Content in Needles and Soil
4.6. Statistical Analysis
5. Conclusions
Acknowledgements
- Sample Availability: Samples of the compounds DHAP are available from the authors.
References
- Inderjit, S.; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 1–8. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Autotoxicity: Concept, organisms, and ecological significance. Crit. Rev. Plant Sci. 1999, 18, 757–772. [Google Scholar] [CrossRef]
- Weidenhamer, J.D.; Hartnett, D.C.; Romeo, J.T. Density dependent phytotoxicity: distinguishing resource competition and allelopathic interference in plants. J. Appl. Ecol. 1989, 26, 613–624. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive plant versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Yu, J.Q.; Shou, Y.; Qian, R.; Hu, W.H. Autotoxic potential in cucurbit crops. Plant Soil 2000, 223, 147–151. [Google Scholar]
- Thibault, J.R.; Fortin, J.A.; Smirnoff, W.A. In vitro allelopathic inhibition of nitrification by balsam poplar (Populus balsamifera) and balsam fir (Abies balsamea). Am. J. Bot. 1982, 28, 478–485. [Google Scholar]
- Zhang, Q. Potential role of allelopathy in the soil and the decomposing root of Chinese fir replanted woodland. Plant Soil 1993, 15, 212–224. [Google Scholar]
- Gallet, C. Allelopathic potential in bilberry-spruce forests: Influence of phenolic compounds on spruce seedlings. J. Chem. Ecol. 1994, 20, 1009–1024. [Google Scholar] [CrossRef]
- Mallik, A.U.; Newton, P.F. Inhibition of black spruce seedling growth by forest-floor substances of central Newfoundland. For. Ecol. Manag. 1988, 23, 273–283. [Google Scholar] [CrossRef]
- Fernandez, C.; Voiriot, S.; Mévy, J.P.; Vila, B.; Ormeno, E.; Dupouyet, S.; Bousquet-Mélou, A. Regeneration failure of Pinus halepensis Mill.: The role of autotoxicity and some abiotic environmental parameters. For. Ecol. Manag. 2008, 255, 2928–2936. [Google Scholar] [CrossRef]
- Fernandez, C.; Monnier, Y.; Ormeno, E.; Baldy, V.; Greff, S.; Pasqualini, V.; Mévy, J.P.; Bousquet-Mélou, A. Variations in allelochemicals composition of leachates of different organs and maturity stages of Pinus halepensis. J. Chem. Ecol. 2009, 35, 970–979. [Google Scholar] [CrossRef]
- Alrababah, M.A.; Tadros, M.J.; Samarah, A.H.; Ghosheh, H. Allelopathic effects of Pinus halepensis and Quercus coccifera on the germination of Mediterranean crop seeds. New For. 2009, 38, 261–272. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Seki, T.; Shigemori, H. Allelopathy and allelopathic substance in the moss Rhynchostegium pallidifolium. J. Plant Phys. 2010, 167, 468–471. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M. Seasonal fluctuations in soil phenolics of coniferous forest: Effects of seed germination of different coniferous species. Plant Soil 2006, 284, 305–318. [Google Scholar] [CrossRef]
- Alias, J.C.; Sosa, T.; Escudero, J.C.; Chaves, N. Autotoxicity against germination and seedling emergence in Cistus ladanifer L. Plant Soil 2006, 282, 327–332. [Google Scholar] [CrossRef]
- Mallik, A. Conifer regeneration problems in boreal and temperate forests with ericaceous understory: Role of disturbance, seedbed limitation, and keystone species change. Crit. Rev. Plant Sci. 2003, 22, 341–366. [Google Scholar] [CrossRef]
- Weir, T.L.; Bais, H.P.; Vivanco, J.M. Intraspecific and interspecific interactions mediated by a phytotoxin, (−)-catechin, secreted by the roots of Centaurea maculosa (spotted knapweed). J. Chem. Ecol. 2003, 29, 2937–2412. [Google Scholar]
- Wang, Q.; Ruan, X.; Pan, C.D.; Xu, N.Y.; Luo, X.; Huang, M.M. Need for sustainability policy—A case study of the National Forest Conservation Program (NFCP) in the western region of Tianshan Mountain, China. For. Chron. 2006, 82, 31–39. [Google Scholar]
- Li, Z.H.; Wang, Q.; Lia, J.J.; Ruan, X.; Pan, C.D.; Jiang, D.A.; Luo, C.C. Elementary identification of potential autotoxins from Picea schrenkiana litters. Chin. J. Anal. Chem. 2009, 6, 888–892. [Google Scholar]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Zhang, J.C.; Jiang, D.A.; Wang, G.G. Biological activity and quantification of potential autotoxins from Picea schrenkiana leaves. Allelopathy J. 2011, 27, 245–262. [Google Scholar]
- Lodhi, M.A.K.; Killingbeck, K.T. Effects of pine-produced chemicals on selected understory species in a Pinus ponderosa community. J. Chem. Ecol. 1982, 8, 275–283. [Google Scholar] [CrossRef]
- Kil, B.S.; Yim, Y.J. Allelopathic effects of pinus-densiflora on undergrowth of red pine forest. J. Chem. Ecol. 1983, 9, 1135–1151. [Google Scholar] [CrossRef]
- Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Role of root-mediated interactions in phytotoxic interference of Ageratum conyzoides with rice (Oryza sativa). Flora 2008a, 204, 388–395. [Google Scholar]
- Rudrappa, T.; Bonsall, J.; Gallagher, J.L.; Seliskar, D.M.; Bais, H.P. Root-secreted allelochemical in the noxious weed Phragmites Australis deploys a reactive oxygen species response and microtubule assembly disruption to execute rhizotoxicity. J. Chem. Ecol. 2007, 33, 1898–1918. [Google Scholar] [CrossRef]
- Hiradate, S.; Morita, S.; Furubayashi, A.; Fujii, Y.; Harada, J. Plant growth inhibition by cis-cinnamoyl glucosides and cis-cinnamic acid. J. Chem. Ecol. 2005, 31, 591–601. [Google Scholar] [CrossRef]
- Beijing Pharmaceutical Research Institute. Research on the effective components of leaves of Tumaodongqing. Chin. Tradit. Herb. Drugs 1977, 8, 7–10.
- Kim, Y.J.; No, J.K.; Lee, J.S.; Kim, M.S.; Chung, H.Y. Antimelanogenic activity of 3,4-dihydroxyacetophenone: Inhibition of tyrosinase and MITF. Biosci. Biochenol. Biochem. 2006, 70, 532–534. [Google Scholar] [CrossRef]
- Yang, D.S.; Wu, X.R.; Ma, T.Y. Effects of 3,4-dihydroxyacetophenone on the biosynthesis of TXA2 and PGI2 in human placental villus and umbilical artery segments in vitro. Prostagladins 1989, 38, 497–504. [Google Scholar] [CrossRef]
- Blum, U. Allelopathic interactions involving phenolic acids. J. Nematol. 1996, 28, 259–267. [Google Scholar]
- Einhellig, F.A. Interactions involving allelopathy in cropping systems. Agronomy J. 1996, 88, 886–893. [Google Scholar] [CrossRef]
- Mizutani, J. Selected allelochemicals. Crit. Rev. Plant Sci. 1999, 18, 653–671. [Google Scholar] [CrossRef]
- Chen, L.C.; Wang, S.L.; Yu, X.J. Effects of phenolics on seedling growth and 15N nitrate absorption of Cunninghamia lanceolata. Allelopathy J. 2005, 15, 57–66. [Google Scholar]
- Huang, Z.Q.; Haig, T.; Wu, H.W.; An, M.; Pratley, J. Correlation between phytotoxicity on annual ryegrass (Lolium rigidum) and production dynamics of allelochemicals within root exudates of an allelopathic wheat. J. Chem. Ecol. 2003, 29, 2263–2279. [Google Scholar] [CrossRef]
- Zhang, Q.S.; Yu, X. Allelopathy in replant problem in forest soil. Allelopathy J. 2001, 8, 51–64. [Google Scholar]
- Pasqualini, V.; Robles, C.; Garzino, S.; Greff, S.; Bousquet-Melou, A.; Bonin, G. Phenolic compounds content in Pinus halepensis Mill. needles: A bioindicator of air pollution. Chemosphere 2003, 52, 239–248. [Google Scholar] [CrossRef]
- Vandermast, D.B.; Van Lear, D.H.; Clinton, B.D. American chestnut as an allelopathy in the southern Appalachians. For. Ecol. Manag. 2001, 165, 173–181. [Google Scholar]
- Zhang, J.H.; Mao, Z.Q.; Wang, L.Q.; Shu, H.R. Bioassay and identification of root exudates of three fruit tree species. J. Integr. Plant Biol. 2007, 49, 257–261. [Google Scholar] [CrossRef]
- Einhellig, F.A.; Schan, M.K.; Rasmussen, J.A. Synergistic effects of four cinnamic acid compounds on grain sorghom. Plant Growth Regul. 1983, 1, 251–258. [Google Scholar]
- Batish, D.R.; Singh, H.P.; Kaur, S.; Kohli, R.K.; Yadav, S.S. Caffeic acid affects early growth, and morphogenetic response of hypocotyl cuttings of mung bean (Phaseolus aureus). J. Plant Phys. 2008b, 165, 297–305. [Google Scholar] [CrossRef]
- Inderjit. Plant phenolics in allelopathy. Bot. Rev. 1996, 62, 186–202. [CrossRef]
- Chung, I.M.; Miller, D.A. Effect of alfalfa plant and soil extracts on germination and seedling growth. Agronomy J. 1995, 87, 762–767. [Google Scholar] [CrossRef]
- Charoenying, P.; Teerarak, M.; Laosinwattana, C. An allelopathic substance isolated from Zanthoxylum limonella Alston fruit. Sci. Hort. 2010, 125, 411–416. [Google Scholar] [CrossRef]
- ISTA, ISTA International Rules for Seed Testing; International Seed Testing Association: Bassersdorf, Switzerland, 1993; pp. 140–159.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ruan, X.; Li, Z.-H.; Wang, Q.; Pan, C.-D.; Jiang, D.-A.; Wang, G.G. Autotoxicity and Allelopathy of 3,4-Dihydroxyacetophenone Isolated from Picea schrenkiana Needles. Molecules 2011, 16, 8874-8893. https://doi.org/10.3390/molecules16108874
Ruan X, Li Z-H, Wang Q, Pan C-D, Jiang D-A, Wang GG. Autotoxicity and Allelopathy of 3,4-Dihydroxyacetophenone Isolated from Picea schrenkiana Needles. Molecules. 2011; 16(10):8874-8893. https://doi.org/10.3390/molecules16108874
Chicago/Turabian StyleRuan, Xiao, Zhao-Hui Li, Qiang Wang, Cun-De Pan, De-An Jiang, and G. Geoff Wang. 2011. "Autotoxicity and Allelopathy of 3,4-Dihydroxyacetophenone Isolated from Picea schrenkiana Needles" Molecules 16, no. 10: 8874-8893. https://doi.org/10.3390/molecules16108874
APA StyleRuan, X., Li, Z.-H., Wang, Q., Pan, C.-D., Jiang, D.-A., & Wang, G. G. (2011). Autotoxicity and Allelopathy of 3,4-Dihydroxyacetophenone Isolated from Picea schrenkiana Needles. Molecules, 16(10), 8874-8893. https://doi.org/10.3390/molecules16108874