Phage Display for the Generation of Antibodies for Proteome Research, Diagnostics and Therapy


{kind=link}
Abstract
:1. Polyclonal and Monoclonal Antibodies
2. Antibody Phage Display
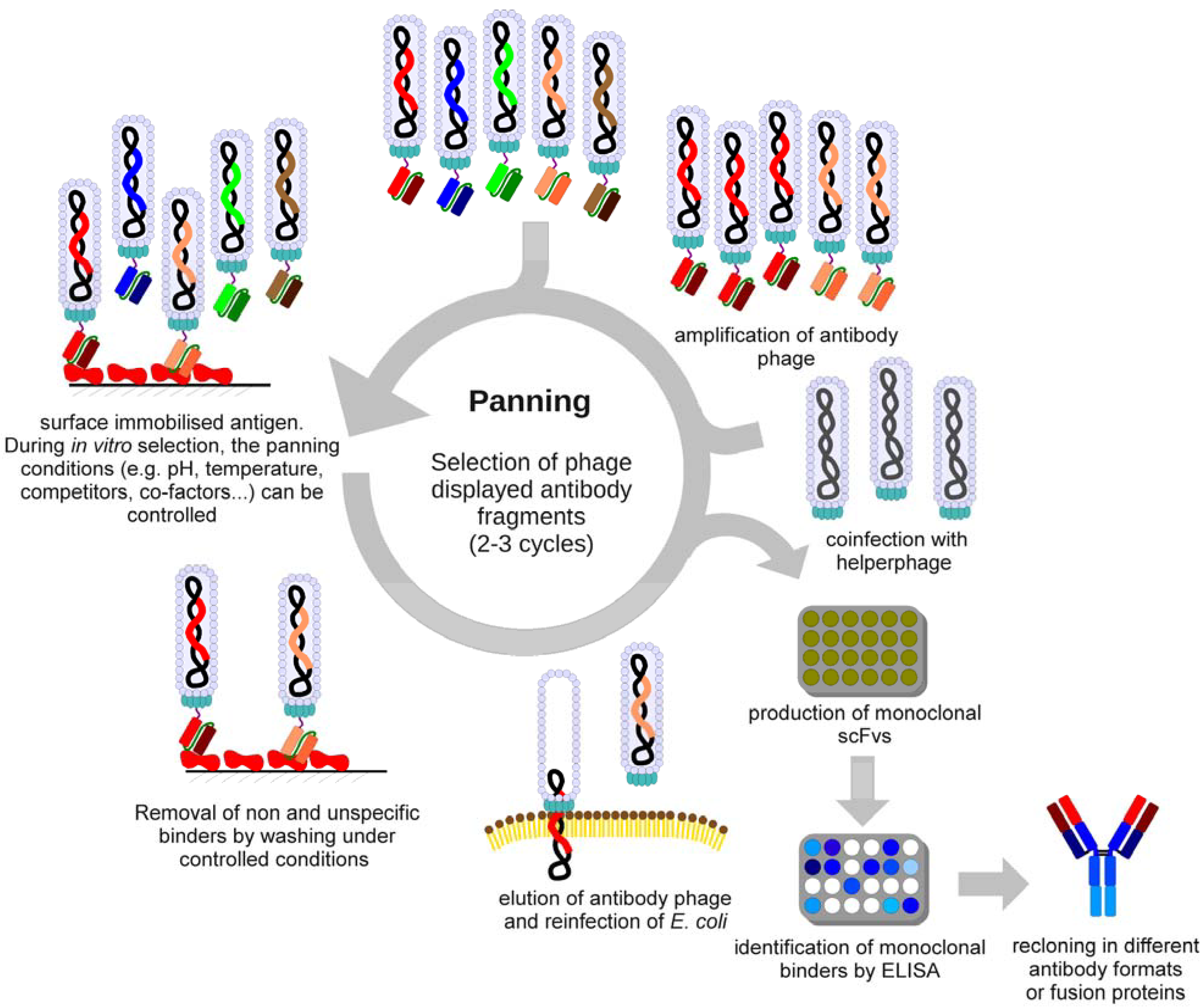
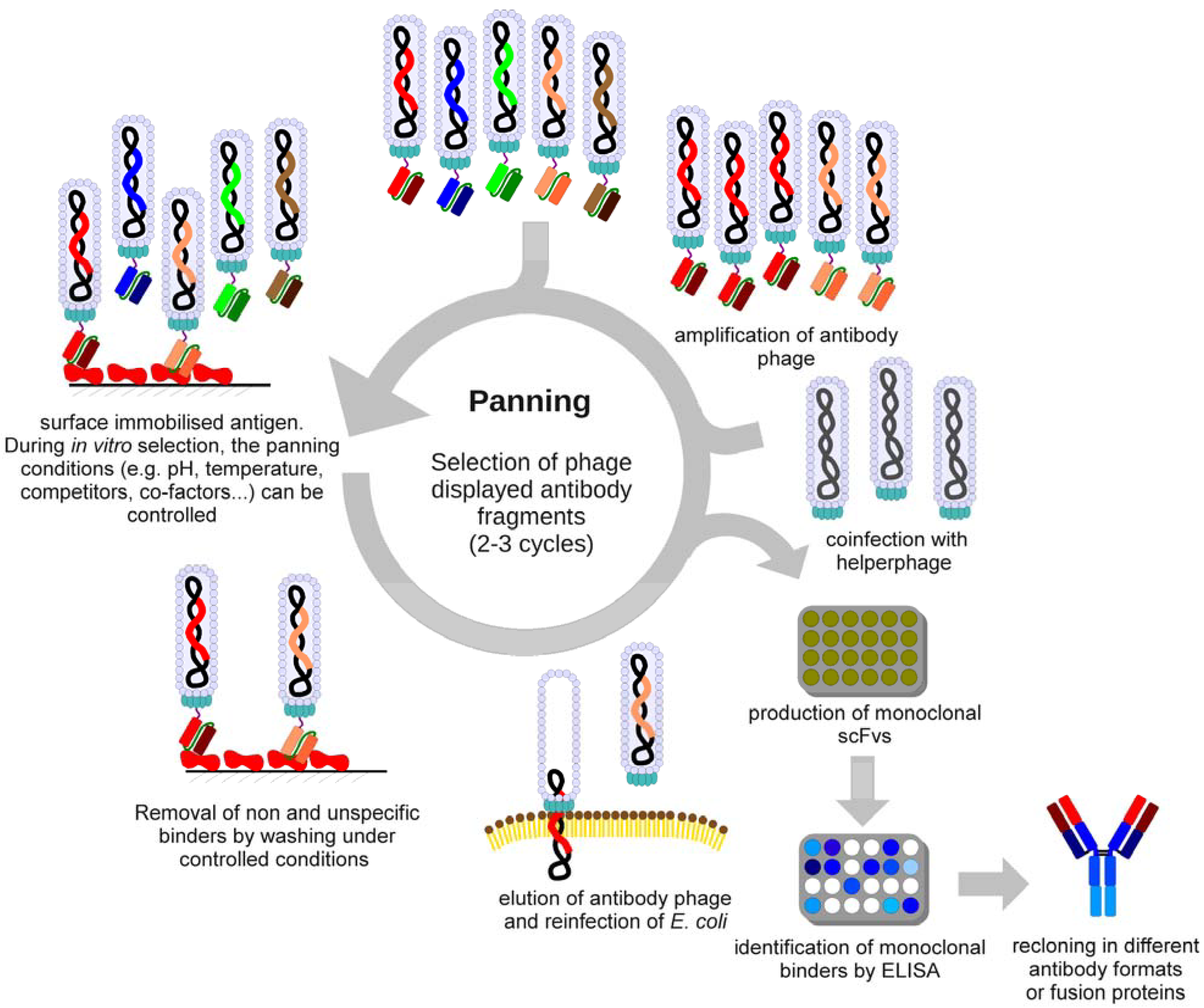
3. Selection of Antibodies
4. Antibody Gene Libraries
5. Antibodies for Basic Research
6. Antibodies for Diagnostics
7. Antibodies for Therapeutical Applications
Acknowledgements
References
- von Behring, E.; Kitasato, S. Über das Zustandekommen der Diphtherie-Immunität und der Tetanus-Immunität bei Thieren. Deut. Med. Wochenzeitschr. 1890, 16, 1113–1114. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar]
- Winter, G.; Milstein, C. Man-made antibodies. Nature 1991, 349, 293–299. [Google Scholar] [CrossRef]
- Courtenay-Luck, N.S.; Epenetos, A.A.; Moore, R.; Larche, M.; Pectasides, D.; Dhokia, B.; Ritter, M.A. Development of primary and secondary immune responses to mouse monoclonal antibodies used in the diagnosis and therapy of malignant neoplasms. Cancer Res. 1986, 46, 6489–6493. [Google Scholar]
- Tjandra, J.J.; Ramadi, L.; McKenzie, I.F. Development of human anti-murine antibody (HAMA) response in patients. Immunol. Cell Biol. 1990, 68, 367–376. [Google Scholar] [CrossRef]
- Almagro, J.C.; Fransson, J. Humanization of antibodies. Front. Biosci. 2008, 13, 1619–1633. [Google Scholar]
- Hwang, W.Y.K.; Foote, J. Immunogenicity of engineered antibodies. Methods 2005, 36, 3–10. [Google Scholar] [CrossRef]
- Kashmiri, S.V.S.; De Pascalis, R.; Gonzales, N.R.; Schlom, J. SDR grafting—a new approach to antibody humanization. Methods 2005, 36, 25–34. [Google Scholar] [CrossRef]
- Studnicka, G.M.; Soares, S.; Better, M.; Williams, R.E.; Nadell, R.; Horwitz, A.H. Human-engineered monoclonal antibodies retain full specific binding activity by preserving non-CDR complementarity-modulating residues. Protein Eng. 1994, 7, 805–814. [Google Scholar] [CrossRef]
- Osbourn, J.; Groves, M.; Vaughan, T. From rodent reagents to human therapeutics using antibody guided selection. Methods 2005, 36, 61–68. [Google Scholar] [CrossRef]
- Fishwild, D.M.; O'Donnell, S.L.; Bengoechea, T.; Hudson, D.V.; Harding, F.; Bernhard, S.L.; Jones, D.; Kay, R.M.; Higgins, K.M.; Schramm, S.R.; Lonberg, N. High-avidity human IgG kappa monoclonal antibodies from a novel strain of minilocus transgenic mice. Nat. Biotechnol. 1996, 14, 845–851. [Google Scholar] [CrossRef]
- Jakobovits, A. Production of fully human antibodies by transgenic mice. Curr. Opin. Biotechnol. 1995, 6, 561–566. [Google Scholar] [CrossRef]
- Kuroiwa, Y.; Kasinathan, P.; Sathiyaseelan, T.; Jiao, J.; Matsushita, H.; Sathiyaseelan, J.; Wu, H.; Mellquist, J.; Hammitt, M.; Koster, J.; Kamoda, S.; Tachibana, K.; Ishida, I.; Robl, J.M. Antigen-specific human polyclonal antibodies from hyperimmunized cattle. Nat. Biotechnol. 2009, 27, 173–181. [Google Scholar]
- Lonberg, N.; Huszar, D. Human antibodies from transgenic mice. Int. Rev. Immunol. 1995, 13, 65–93. [Google Scholar] [CrossRef]
- Weiner, L.M. Fully human therapeutic monoclonal antibodies. J. Immunother. 2006, 29, 1–9. [Google Scholar] [CrossRef]
- Huse, W.D.; Sastry, L.; Iverson, S.A.; Kang, A.S.; Alting-Mees, M.; Burton, D.R.; Benkovic, S.J.; Lerner, R.A. Generation of a large combinatorial library of the immunoglobulin repertoire in phage lambda. Science 1989, 246, 1275–1281. [Google Scholar]
- Persson, M.A.; Caothien, R.H.; Burton, D.R. Generation of diverse high-affinity human monoclonal antibodies by repertoire cloning. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 2432–2436. [Google Scholar] [CrossRef]
- Smith, G.P. Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar]
- Breitling, F.; Dübel, S.; Seehaus, T.; Klewinghaus, I.; Little, M. A surface expression vector for antibody screening. Gene 1991, 104, 147–153. [Google Scholar] [CrossRef]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar]
- Clackson, T.; Hoogenboom, H.R.; Griffiths, A.D.; Winter, G. Making antibody fragments using phage display libraries. Nature 1991, 352, 624–628. [Google Scholar] [CrossRef]
- Hoogenboom, H.R.; Griffiths, A.D.; Johnson, K.S.; Chiswell, D.J.; Hudson, P.; Winter, G. Multi-subunit proteins on the surface of filamentous phage: methodologies for displaying antibody (Fab) heavy and light chains. Nucleic Acids Res. 1991, 19, 4133–4137. [Google Scholar] [CrossRef]
- Barbas, C.F.; Kang, A.S.; Lerner, R.A.; Benkovic, S.J. Assembly of combinatorial antibody libraries on phage surfaces: the gene III site. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 7978–7982. [Google Scholar] [CrossRef]
- Holt, L.J.; Herring, C.; Jespers, L.S.; Woolven, B.P.; Tomlinson, I.M. Domain antibodies: proteins for therapy. Trends Biotechnol. 2003, 21, 484–490. [Google Scholar] [CrossRef]
- Hoet, R.M.; Cohen, E.H.; Kent, R.B.; Rookey, K.; Schoonbroodt, S.; Hogan, S.; Rem, L.; Frans, N.; Daukandt, M.; Pieters, H.; van Hegelsom, R.; Neer, N.C.; Nastri, H.G.; Rondon, I.J.; Leeds, J.A.; Hufton, S.E.; Huang, L.; Kashin, I.; Devlin, M.; Kuang, G.; Steukers, M.; Viswanathan, M.; Nixon, A.E.; Sexton, D.J.; Hoogenboom, H.R.; Ladner, R.C. Generation of high-affinity human antibodies by combining donor-derived and synthetic complementarity-determining-region diversity. Nat. Biotechnol. 2005, 23, 344–348. [Google Scholar]
- Hust, M.; Dübel, S. Phage display vectors for the in vitro generation of human antibody fragments. Methods Mol. Biol. 2005, 295, 71–96. [Google Scholar]
- Mazor, Y.; Van Blarcom, T.; Mabry, R.; Iverson, B.L.; Georgiou, G. Isolation of engineered, full-length antibodies from libraries expressed in Escherichia coli. Nat. Biotechnol. 2007, 25, 563–565. [Google Scholar] [CrossRef]
- Simmons, L.C.; Reilly, D.; Klimowski, L.; Raju, T.S.; Meng, G.; Sims, P.; Hong, K.; Shields, R.L.; Damico, L.A.; Rancatore, P.; Yansura, D.G. Expression of full-length immunoglobulins in Escherichia coli: rapid and efficient production of aglycosylated antibodies. J. Immunol. Methods 2002, 263, 133–147. [Google Scholar] [CrossRef]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef]
- Hawlisch, H.; Müller, M.; Frank, R.; Bautsch, W.; Klos, A.; Köhl, J. Site-specific anti-C3a receptor single-chain antibodies selected by differential panning on cellulose sheets. Anal. Biochem. 2001, 293, 142–145. [Google Scholar]
- Moghaddam, A.; Borgen, T.; Stacy, J.; Kausmally, L.; Simonsen, B.; Marvik, O.J.; Brekke, O.H.; Braunagel, M. Identification of scFv antibody fragments that specifically recognise the heroin metabolite 6-monoacetylmorphine but not morphine. J. Immunol. Methods 2003, 280, 139–155. [Google Scholar] [CrossRef]
- Hust, M.; Maiss, E.; Jacobsen, H.; Reinard, T. The production of a genus-specific recombinant antibody (scFv) using a recombinant potyvirus protease. J. Virol. Methods 2002, 106, 225–233. [Google Scholar] [CrossRef]
- Hoogenboom, H.R. Selecting and screening recombinant antibody libraries. Nat. Biotechnol. 2005, 23, 1105–1116. [Google Scholar] [CrossRef]
- Hust, M.; Dübel, S.; Schirrmann, T. Selection of recombinant antibodies from antibody gene libraries. Methods Mol. Biol. 2007, 408, 243–255. [Google Scholar] [CrossRef]
- Winter, G.; Griffiths, A.D.; Hawkins, R.E.; Hoogenboom, H.R. Making antibodies by phage display technology. Annu. Rev. Immunol. 1994, 12, 433–455. [Google Scholar] [CrossRef]
- Buckler, D.R.; Park, A.; Viswanathan, M.; Hoet, R.M.; Ladner, R.C. Screening isolates from antibody phage-display libraries. Drug Discov. Today 2008, 13, 318–324. [Google Scholar] [CrossRef]
- Hallborn, J.; Carlsson, R. Automated screening procedure for high-throughput generation of antibody fragments. Biotechniques 2002, 30–37. [Google Scholar]
- Konthur, Z.; Hust, M.; Dübel, S. Perspectives for systematic in vitro antibody generation. Gene 2005, 364, 19–29. [Google Scholar] [CrossRef]
- Schirrmann, T.; Al-Halabi, L.; Dübel, S.; Hust, M. Production systems for recombinant antibodies. Front. Biosci. 2008, 13, 4576–4594. [Google Scholar]
- Hust, M.; Meyer, T.; Voedisch, B.; Rülker, T.; Thie, H.; El-Ghezal, A.; Kirsch, M.I.; Schütte, M.; Helmsing, S.; Meier, D.; Schirrmann, T.; Dübel, S. A human scFv antibody generation pipeline for proteome research. J. Biotechnol. 2011. In Press. [Google Scholar]
- Thie, H.; Voedisch, B.; Dübel, S.; Hust, M.; Schirrmann, T. Affinity maturation by phage display. Methods Mol. Biol. 2009, 525, 309–322. [Google Scholar] [CrossRef]
- Kobayashi, N.; Oyama, H.; Kato, Y.; Goto, J.; Söderlind, E.; Borrebaeck, C.A.K. Two-step in vitro antibody affinity maturation enables estradiol-17beta assays with more than 10-fold higher sensitivity. Anal. Chem. 2010, 82, 1027–1038. [Google Scholar]
- Finlay, W.J.; Cunningham, O.; Lambert, M.A.; Darmanin-Sheehan, A.; Liu, X.; Fennell, B.J.; Mahon, C.M.; Cummins, E.; Wade, J.M.; O'Sullivan, C.M.; Tan, X.Y.; Piche, N.; Pittman, D.D.; Paulsen, J.; Tchistiakova, L.; Kodangattil, S.; Gill, D.; Hufton, S.E. Affinity maturation of a humanized rat antibody for anti-RAGE therapy: comprehensive mutagenesis reveals a high level of mutational plasticity both inside and outside the complementarity-determining regions. J. Mol. Biol. 2009, 388, 541–558. [Google Scholar] [CrossRef]
- Pelat, T.; Hust, M.; Laffly, E.; Condemine, F.; Bottex, C.; Vidal, D.; Lefranc, M.; Dübel, S.; Thullier, P. High-affinity, human antibody-like antibody fragment (single-chain variable fragment) neutralizing the lethal factor (LF) of Bacillus anthracis by inhibiting protective antigen-LF complex formation. Antimicrob. Agents Chemother. 2007, 51, 2758–2764. [Google Scholar]
- Pelat, T.; Hust, M.; Hale, M.; Lefranc, M.; Dübel, S.; Thullier, P. Isolation of a human-like antibody fragment (scFv) that neutralizes ricin biological activity. BMC Biotechnol. 2009, 9, 60. [Google Scholar] [CrossRef]
- Sun, L.; Lu, X.; Li, C.; Wang, M.; Liu, Q.; Li, Z.; Hu, X.; Li, J.; Liu, F.; Li, Q.; Belser, J.A.; Hancock, K.; Shu, Y.; Katz, J.M.; Liang, M.; Li, D. Generation, characterization and epitope mapping of two neutralizing and protective human recombinant antibodies against influenza A H5N1 viruses. PloS One 2009, 4, e5476. [Google Scholar]
- Hust, M.; Dübel, S. Mating antibody phage display with proteomics. Trends Biotechnol. 2004, 22, 8–14. [Google Scholar] [CrossRef]
- de Haard, H.J.; van Neer, N.; Reurs, A.; Hufton, S.E.; Roovers, R.C.; Henderikx, P.; de Bruïne, A.P.; Arends, J.W.; Hoogenboom, H.R. A large non-immunized human Fab fragment phage library that permits rapid isolation and kinetic analysis of high affinity antibodies. J. Biol. Chem. 1999, 274, 18218–18230. [Google Scholar]
- Griffiths, A.D.; Williams, S.C.; Hartley, O.; Tomlinson, I.M.; Waterhouse, P.; Crosby, W.L.; Kontermann, R.E.; Jones, P.T.; Low, N.M.; Allison, T.J. Isolation of high affinity human antibodies directly from large synthetic repertoires. EMBO J. 1994, 13, 3245–3260. [Google Scholar]
- Pini, A.; Viti, F.; Santucci, A.; Carnemolla, B.; Zardi, L.; Neri, P.; Neri, D. Design and use of a phage display library. Human antibodies with subnanomolar affinity against a marker of angiogenesis eluted from a two-dimensional gel. J. Biol. Chem. 1998, 273, 21769–21776. [Google Scholar]
- Hayashi, N.; Welschof, M.; Zewe, M.; Braunagel, M.; Dübel, S.; Breitling, F.; Little, M. Simultaneous mutagenesis of antibody CDR regions by overlap extension and PCR. Biotechniques 1994, 17, 310–315. [Google Scholar]
- Knappik, A.; Ge, L.; Honegger, A.; Pack, P.; Fischer, M.; Wellnhofer, G.; Hoess, A.; Wölle, J.; Plückthun, A.; Virnekäs, B. Fully synthetic human combinatorial antibody libraries (HuCAL) based on modular consensus frameworks and CDRs randomized with trinucleotides. J. Mol. Biol. 2000, 296, 57–86. [Google Scholar] [CrossRef]
- Sblattero, D.; Bradbury, A. Exploiting recombination in single bacteria to make large phage antibody libraries. Nat. Biotechnol. 2000, 18, 75–80. [Google Scholar] [CrossRef]
- Bittner, M.; Kupferer, P.; Morris, C.F. Electrophoretic transfer of proteins and nucleic acids from slab gels to diazobenzyloxymethyl cellulose or nitrocellulose sheets. Anal. Biochem. 1980, 102, 459–471. [Google Scholar] [CrossRef]
- Renart, J.; Reiser, J.; Stark, G.R. Transfer of proteins from gels to diazobenzyloxymethyl-paper and detection with antisera: a method for studying antibody specificity and antigen structure. Proc. Natl. Acad. Sci. U.S.A. 1979, 76, 3116–3120. [Google Scholar] [CrossRef]
- Engvall, E.; Perlmann, P. Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Immunochemistry 1971, 8, 871–874. [Google Scholar] [CrossRef]
- Coons, A.H.; Creech, H.J.; Jones, R.N. Immunological properties of an antibody containing a fluorescence group. Proc. Soc. Exp. Biol. Med. 1941, 47, 200–202. [Google Scholar]
- Bonner, W.A.; Hulett, H.R.; Sweet, R.G.; Herzenberg, L.A. Fluorescence activated cell sorting. Rev. Sci. Instrum. 1972, 43, 404–409. [Google Scholar] [CrossRef]
- Porter, R.R.; Press, E.M. Immunochemistry. Annu. Rev. Biochem. 1962, 31, 625–652. [Google Scholar] [CrossRef]
- Silzel, J.W.; Cercek, B.; Dodson, C.; Tsay, T.; Obremski, R.J. Mass-sensing, multianalyte microarray immunoassay with imaging detection. Clin. Chem. 1998, 44, 2036–2043. [Google Scholar]
- Stoevesandt, O.; Taussig, M.J.; He, M. Protein microarrays: high-throughput tools for proteomics. Expert Rev. Proteomics 2009, 6, 145–157. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [CrossRef]
- Levy, S.; Sutton, G.; Ng, P.C.; Feuk, L.; Halpern, A.L.; Walenz, B.P.; Axelrod, N.; Huang, J.; Kirkness, E.F.; Denisov, G.; Lin, Y.; MacDonald, J.R.; Pang, A.W.C.; Shago, M.; Stockwell, T.B.; Tsiamouri, A.; Bafna, V.; Bansal, V.; Kravitz, S.A.; Busam, D.A.; Beeson, K.Y.; McIntosh, T.C.; Remington, K.A.; Abril, J.F.; Gill, J.; Borman, J.; Rogers, Y.; Frazier, M.E.; Scherer, S.W.; Strausberg, R.L.; Venter, J.C. The diploid genome sequence of an individual human. PLoS Biol. 2007, 5, e254. [Google Scholar]
- Wheeler, D.A.; Srinivasan, M.; Egholm, M.; Shen, Y.; Chen, L.; McGuire, A.; He, W.; Chen, Y.; Makhijani, V.; Roth, G.T.; Gomes, X.; Tartaro, K.; Niazi, F.; Turcotte, C.L.; Irzyk, G.P.; Lupski, J.R.; Chinault, C.; Song, X.; Liu, Y.; Yuan, Y.; Nazareth, L.; Qin, X.; Muzny, D.M.; Margulies, M.; Weinstock, G.M.; Gibbs, R.A.; Rothberg, J.M. The complete genome of an individual by massively parallel DNA sequencing. Nature 2008, 452, 872–876. [Google Scholar]
- Harrison, P.M.; Kumar, A.; Lang, N.; Snyder, M.; Gerstein, M. A question of size: the eukaryotic proteome and the problems in defining it. Nucleic Acids Res. 2002, 30, 1083–1090. [Google Scholar] [CrossRef]
- Berglund, L.; Björling, E.; Oksvold, P.; Fagerberg, L.; Asplund, A.; Szigyarto, C.A.; Persson, A.; Ottosson, J.; Wernérus, H.; Nilsson, P.; Lundberg, E.; Sivertsson, A.; Navani, S.; Wester, K.; Kampf, C.; Hober, S.; Pontén, F.; Uhlén, M. A genecentric Human Protein Atlas for expression profiles based on antibodies. Mol. Cell. Proteomics 2008, 7, 2019–2027. [Google Scholar] [CrossRef]
- Dübel, S.; Stoevesandt, O.; Taussig, M.J.; Hust, M. Generating recombinant antibodies to the complete human proteome. Trends Biotechnol. 2010, 28, 333–339. [Google Scholar] [CrossRef]
- Taussig, M.J.; Stoevesandt, O.; Borrebaeck, C.A.K.; Bradbury, A.R.; Cahill, D.; Cambillau, C.; de Daruvar, A.; Dübel, S.; Eichler, J.; Frank, R.; Gibson, T.J.; Gloriam, D.; Gold, L.; Herberg, F.W.; Hermjakob, H.; Hoheisel, J.D.; Joos, T.O.; Kallioniemi, O.; Koegl, M.; Konthur, Z.; Korn, B.; Kremmer, E.; Krobitsch, S.; Landegren, U.; van der Maarel, S.; McCafferty, J.; Muyldermans, S.; Nygren, P.; Palcy, S.; Plückthun, A.; Polic, B.; Przybylski, M.; Saviranta, P.; Sawyer, A.; Sherman, D.J.; Skerra, A.; Templin, M.; Ueffing, M.; Uhlén, M. ProteomeBinders: planning a European resource of affinity reagents for analysis of the human proteome. Nat. Methods 2007, 4, 13–17. [Google Scholar]
- Wingren, C.; James, P.; Borrebaeck, C.A.K. Strategy for surveying the proteome using affinity proteomics and mass spectrometry. Proteomics 2009, 9, 1511–1517. [Google Scholar] [CrossRef]
- Mersmann, M.; Meier, D.; Mersmann, J.; Helmsing, S.; Nilsson, P.; Gräslund, S.; Structural Genomics Consortium; Colwill, K.; Hust, M.; Dübel, S. Towards proteome scale antibody selections using phage display. New Biotechnol. 2010, 27, 118–128. [Google Scholar] [CrossRef]
- Pershad, K.; Pavlovic, J.D.; Gräslund, S.; Nilsson, P.; Colwill, K.; Karatt-Vellatt, A.; Schofield, D.J.; Dyson, M.R.; Pawson, T.; Kay, B.K.; McCafferty, J. Generating a panel of highly specific antibodies to 20 human SH2 domains by phage display. Protein Eng. Des. Sel. 2010, 23, 279–288. [Google Scholar] [CrossRef]
- Björling, E.; Uhlén, M. Antibodypedia, a portal for sharing antibody and antigen validation data. Mol. Cell. Proteomics: MCP 2008, 7, 2028–2037. [Google Scholar] [CrossRef]
- Berglund, L.; Björling, E.; Oksvold, P.; Fagerberg, L.; Asplund, A.; Szigyarto, C.A.; Persson, A.; Ottosson, J.; Wernérus, H.; Nilsson, P.; Lundberg, E.; Sivertsson, A.; Navani, S.; Wester, K.; Kampf, C.; Hober, S.; Pontén, F.; Uhlén, M. A genecentric Human Protein Atlas for expression profiles based on antibodies. Mol. Cell. Proteomics 2008, 7, 2019–2027. [Google Scholar] [CrossRef]
- He, M.; Taussig, M.J. Selection of recombinant antibodies by eukaryotic ribosome display. Methods Mol. Biol. 2008, 484, 193–205. [Google Scholar] [CrossRef]
- Steiner, D.; Forrer, P.; Plückthun, A. Efficient selection of DARPins with sub-nanomolar affinities using SRP phage display. J. Mol. Biol. 2008, 382, 1211–1227. [Google Scholar] [CrossRef]
- Skerra, A. Alternative binding proteins: anticalins - harnessing the structural plasticity of the lipocalin ligand pocket to engineer novel binding activities. FEBS J. 2008, 275, 2677–2683. [Google Scholar] [CrossRef]
- Dussart, P.; Petit, L.; Labeau, B.; Bremand, L.; Leduc, A.; Moua, D.; Matheus, S.; Baril, L. Evaluation of Two New Commercial Tests for the Diagnosis of Acute Dengue Virus Infection Using NS1 Antigen Detection in Human Serum. PLoS Negl. Trop. Dis. 2008, 2, e280. [Google Scholar] [CrossRef]
- Mergener, K.; Enzensberger, W.; Rübsamen-Waigmann, H.; von Briesen, H.; Doerr, H.W. Immunoglobulin class- and subclass-specific HIV antibody detection in serum and CSF specimens by ELISA and Western blot. Infection 1987, 15, 317–322. [Google Scholar] [CrossRef]
- Dong, L.; Masaki, Y.; Takegami, T.; Kawanami, T.; Itoh, K.; Jin, Z.; Huang, C.; Tong, X.; Fukushima, T.; Tanaka, M.; Sawaki, T.; Sakai, T.; Sugai, S.; Okazaki, T.; Hirose, Y.; Umehara, H. Cloning and expression of two human recombinant monoclonal Fab fragments specific for EBV viral capsid antigen. Int. Immunol. 2007, 19, 331–336. [Google Scholar] [CrossRef]
- Abu Elzein, E.M.; Crowther, J.R. Enzyme-labelled immunosorbent assay techniques in foot-and-mouth disease virus research. J. Hyg. 1978, 80, 391–399. [Google Scholar] [CrossRef]
- Molinelli, A.; Grossalber, K.; Führer, M.; Baumgartner, S.; Sulyok, M.; Krska, R. Development of qualitative and semiquantitative immunoassay-based rapid strip tests for the detection of T-2 toxin in wheat and oat. J. Agr. Food Chem. 2008, 56, 2589–2594. [Google Scholar] [CrossRef]
- Schweers, B.A.; Old, J.; Boonlayangoor, P.W.; Reich, K.A. Developmental validation of a novel lateral flow strip test for rapid identification of human blood (Rapid Stain Identification--Blood). Foren. Sci. Int. Genet. 2008, 2, 243–247. [Google Scholar] [CrossRef]
- Vignali, D.A. Multiplexed particle-based flow cytometric assays. J. Immunol. Methods 2000, 243, 243–255. [Google Scholar] [CrossRef]
- Sashihara, J.; Burbelo, P.D.; Savoldo, B.; Pierson, T.C.; Cohen, J.I. Human antibody titers to Epstein-Barr Virus (EBV) gp350 correlate with neutralization of infectivity better than antibody titers to EBV gp42 using a rapid flow cytometry-based EBV neutralization assay. Virology 2009, 391, 249–256. [Google Scholar] [CrossRef]
- Marrack, J.R. Sensitivy and specificity of methods of detecting antibodies. Brit. Med. Bull. 1963, 19, 178–182. [Google Scholar]
- Schlingemann, J.; Leijon, M.; Yacoub, A.; Schlingemann, H.; Zohari, S.; Matyi-Tóth, A.; Kiss, I.; Holmquist, G.; Nordengrahn, A.; Landegren, U.; Ekström, B.; Belák, S. Novel means of viral antigen identification: improved detection of avian influenza viruses by proximity ligation. J. Virol. Methods 2010, 163, 116–122. [Google Scholar] [CrossRef]
- Cai, J.; Liu, Z.; Wang, F.; Li, F. Phage display applications for molecular imaging. Curr. Pharm. Biotechnol. 2010, 11, 603–609. [Google Scholar] [CrossRef]
- Taussig, M.J. Bacteriophage linked assays for antibodies to protein antigens. Immunology 1970, 18, 323–330. [Google Scholar]
- Gao, C.; Mao, S.; Lo, C.H.; Wirsching, P.; Lerner, R.A.; Janda, K.D. Making artificial antibodies: a format for phage display of combinatorial heterodimeric arrays. Proc. Natl. Acad. Sci. U.S.A 1999, 96, 6025–6030. [Google Scholar] [CrossRef]
- Kirsch, M.; Hülseweh, B.; Nacke, C.; Rülker, T.; Schirrmann, T.; Marschall, H.; Hust, M.; Dübel, S. Development of human antibody fragments using antibody phage display for the detection and diagnosis of Venezuelan equine encephalitis virus (VEEV). BMC Biotechnol. 2008, 8, 66. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, Y.; Zhang, X.; Zhang, Z.; Qiao, Y.; Bi, L.; Wen, J.; Liang, M.; Zhang, J. Phage display mediated immuno-PCR. Nucleic Acids Res. 2006, 34, e62. [Google Scholar] [CrossRef]
- Meyer, T.; Stratmann-Selke, J.; Meens, J.; Schirrmann, T.; Gerlach, G.F.; Frank, R.; Dübel, S.; Strutzberg-Minder, K.; Hust, M. Isolation of scFv fragments specific to OmpD of Salmonella Typhimurium. Vet. Microbiol. 2011, 147, 162–169. [Google Scholar] [CrossRef]
- Pitaksajjakul, P.; Lekcharoensuk, P.; Upragarin, N.; Barbas, C.F.; Ibrahim, M.S.; Ikuta, K.; Ramasoota, P. Fab MAbs specific to HA of influenza virus with H5N1 neutralizing activity selected from immunized chicken phage library. Biochem. Biophys. Res. Commun. 2010, 395, 496–501. [Google Scholar] [CrossRef]
- Schütte, M.; Thullier, P.; Pelat, T.; Wezler, X.; Rosenstock, P.; Hinz, D.; Kirsch, M.I.; Hasenberg, M.; Frank, R.; Schirrmann, T.; Gunzer, M.; Hust, M.; Dübel, S. Identification of a putative Crf splice variant and generation of recombinant antibodies for the specific detection of Aspergillus fumigatus. PloS One 2009, 4, e6625. [Google Scholar]
- Kügler, J.; Nieswandt, S.; Gerlach, G.F.; Meens, J.; Schirrmann, T.; Hust, M. Identification of immunogenic polypeptides from a Mycoplasma hyopneumoniae genome library by phage display. Appl. Microbiol. Biotechnol. 2008, 80, 447–58. [Google Scholar] [CrossRef]
- Naseem, S.; Meens, J.; Jores, J.; Heller, M.; Dübel, S.; Hust, M.; Gerlach, G. Phage display-based identification and potential diagnostic application of novel antigens from Mycoplasma mycoides subsp. mycoides small colony type. Vet. Microbiol. 2010, 142, 285–292. [Google Scholar] [CrossRef]
- Miltiadou, D.R.; Mather, A.; Vilei, E.M.; Du Plessis, D.H. Identification of genes coding for B cell antigens of Mycoplasma mycoides subsp. mycoides Small Colony (MmmSC) by using phage display. BMC Microbiol. 2009, 9, 215. [Google Scholar] [CrossRef]
- Dübel, S. Recombinant therapeutic antibodies. Appl. Microbiol. Biotechnol. 2007, 74, 723–729. [Google Scholar] [CrossRef]
- Alonso-Ruiz, A.; Pijoan, J.I.; Ansuategui, E.; Urkaregi, A.; Calabozo, M.; Quintana, A. Tumor necrosis factor alpha drugs in rheumatoid arthritis: systematic review and metaanalysis of efficacy and safety. BMC Musculoskelet. Disord. 2008, 9, 52. [Google Scholar] [CrossRef]
- Peeters, M.; Price, T.; Van Laethem, J. Anti-epidermal growth factor receptor monotherapy in the treatment of metastatic colorectal cancer: where are we today? Oncologist 2009, 14, 29–39. [Google Scholar] [CrossRef]
- Chatenoud, L.; Bluestone, J.A. CD3-specific antibodies: a portal to the treatment of autoimmunity. Nat. Rev. Immunol. 2007, 7, 622–632. [Google Scholar] [CrossRef]
- Adams, G.P.; Weiner, L.M. Monoclonal antibody therapy of cancer. Nat. Biotechnol. 2005, 23, 1147–1157. [Google Scholar] [CrossRef]
- Moss, M.L.; Sklair-Tavron, L.; Nudelman, R. Drug insight: tumor necrosis factor-converting enzyme as a pharmaceutical target for rheumatoid arthritis. Nat. Clin. Pract. Rheumatol. 2008, 4, 300–309. [Google Scholar] [CrossRef]
- Dalle, S.; Thieblemont, C.; Thomas, L.; Dumontet, C. Monoclonal antibodies in clinical oncology. Anticancer Agents Med. Chem. 2008, 8, 523–532. [Google Scholar]
- Jones, S.E. Metastatic breast cancer: the treatment challenge. Clin. Breast Cancer 2008, 8, 224–233. [Google Scholar] [CrossRef]
- Moroni, M.; Veronese, S.; Benvenuti, S.; Marrapese, G.; Sartore-Bianchi, A.; Di Nicolantonio, F.; Gambacorta, M.; Siena, S.; Bardelli, A. Gene copy number for epidermal growth factor receptor (EGFR) and clinical response to antiEGFR treatment in colorectal cancer: a cohort study. Lancet Oncol. 2005, 6, 279–286. [Google Scholar] [CrossRef]
- Kay, J.; Matteson, E.L.; Dasgupta, B.; Nash, P.; Durez, P.; Hall, S.; Hsia, E.C.; Han, J.; Wagner, C.; Xu, Z.; Visvanathan, S.; Rahman, M.U. Golimumab in patients with active rheumatoid arthritis despite treatment with methotrexate: a randomized, double-blind, placebo-controlled, dose-ranging study. Arthritis Rheum. 2008, 58, 964–975. [Google Scholar] [CrossRef]
- Church, L.D.; McDermott, M.F. Canakinumab: a human anti-IL-1β monoclonal antibody for the treatment of cryopyrin-associated periodic syndromes. Expert Rev. Clin. Immunol. 2010, 6, 831–841. [Google Scholar] [CrossRef]
- Kavanaugh, A.; Menter, A.; Mendelsohn, A.; Shen, Y.; Lee, S.; Gottlieb, A.B. Effect of ustekinumab on physical function and health-related quality of life in patients with psoriatic arthritis: a randomized, placebo-controlled, phase II trial. Curr. Med. Res. Opin. 2010, 26, 2385–2392. [Google Scholar] [CrossRef]
- Schirrmann, T.; Hust, M. Construction of human antibody gene libraries and selection of antibodies by phage display. Methods Mol. Biol. 2010, 651, 177–209. [Google Scholar] [CrossRef]
- Thie, H.; Meyer, T.; Schirrmann, T.; Hust, M.; Dübel, S. Phage display derived therapeutic antibodies. Curr. Pharm. Biotechnol. 2008, 9, 439–446. [Google Scholar] [CrossRef]
- Vaughan, T.J.; Williams, A.J.; Pritchard, K.; Osbourn, J.K.; Pope, A.R.; Earnshaw, J.C.; McCafferty, J.; Hodits, R.A.; Wilton, J.; Johnson, K.S. Human antibodies with sub-nanomolar affinities isolated from a large non-immunized phage display library. Nat. Biotechnol. 1996, 14, 309–314. [Google Scholar] [CrossRef]
- Baker, K.P.; Edwards, B.M.; Main, S.H.; Choi, G.H.; Wager, R.E.; Halpern, W.G.; Lappin, P.B.; Riccobene, T.; Abramian, D.; Sekut, L.; Sturm, B.; Poortman, C.; Minter, R.R.; Dobson, C.L.; Williams, E.; Carmen, S.; Smith, R.; Roschke, V.; Hilbert, D.M.; Vaughan, T.J.; Albert, V.R. Generation and characterization of LymphoStat-B, a human monoclonal antibody that antagonizes the bioactivities of B lymphocyte stimulator. Arthritis Rheum. 2003, 48, 3253–3265. [Google Scholar]
- Thie, H.; Toleikis, S.; Li, J.; von Wasielewsky, R.; Bastert, G.; Schirrmann, T.; Tourais Esteves, I.; Behrens, C.K.; Fournes, B.; Fournier, N.; de Romeuf, C.; Hust, M.; Dübel, S. Rise and fall of an anti-MUC1 specific antibody. PLoS One 2011. In Press. [Google Scholar]
- Wallace, D.J.; Stohl, W.; Furie, R.A.; Lisse, J.R.; McKay, J.D.; Merrill, J.T.; Petri, M.A.; Ginzler, E.M.; Chatham, W.W.; McCune, W.J.; Fernandez, V.; Chevrier, M.R.; Zhong, Z.J.; Freimuth, W.W. A phase II, randomized, double-blind, placebo-controlled, dose-ranging study of belimumab in patients with active systemic lupus erythematosus. Arthritis Rheum. 2009, 61, 1168–1178. [Google Scholar] [CrossRef]
- Mazumdar, S. Raxibacumab. mAbs 2009, 1, 531–538. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schirrmann, T.; Meyer, T.; Schütte, M.; Frenzel, A.; Hust, M. Phage Display for the Generation of Antibodies for Proteome Research, Diagnostics and Therapy. Molecules 2011, 16, 412-426. https://doi.org/10.3390/molecules16010412
Schirrmann T, Meyer T, Schütte M, Frenzel A, Hust M. Phage Display for the Generation of Antibodies for Proteome Research, Diagnostics and Therapy. Molecules. 2011; 16(1):412-426. https://doi.org/10.3390/molecules16010412
Chicago/Turabian StyleSchirrmann, Thomas, Torsten Meyer, Mark Schütte, André Frenzel, and Michael Hust. 2011. "Phage Display for the Generation of Antibodies for Proteome Research, Diagnostics and Therapy" Molecules 16, no. 1: 412-426. https://doi.org/10.3390/molecules16010412
APA StyleSchirrmann, T., Meyer, T., Schütte, M., Frenzel, A., & Hust, M. (2011). Phage Display for the Generation of Antibodies for Proteome Research, Diagnostics and Therapy. Molecules, 16(1), 412-426. https://doi.org/10.3390/molecules16010412