Antiradical and Cytoprotective Activities of Several C-Geranyl-substituted Flavanones from Paulownia tomentosa Fruit

Abstract
:1. Introduction
2. Results and Discussion

| Compound / substituent | R1 | R2 | R3 | R4 | R5 |
|---|---|---|---|---|---|
| Diplacone (1) | OH | OH | H | H | 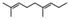 |
| Mimulone (2) | H | OH | H | H | |
| 3´-O-methyldiplacone (3) | OMe | OH | H | H | |
| Tomentodiplacone (4) | OMe | OH | H | H | 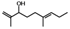 |
| 3´-O-methyldiplacol (5) | OMe | OH | H | OH | |
| 3´-O-methyl-5´-OH-diplacone (6) | OMe | OH | OH | H | |
| 3´-O-methyl-5´-O-methyldiplacone (7) | OMe | OH | OMe | H | |
| Tomentodiplacone B (8) | OMe | OH | H | H | 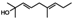 |
| Schizolaenone C (9) | OH | H | OH | H | |
| 6-isopentenyl-3´-O-methyltaxifolin (10) | OMe | OH | H | OH | 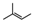 |
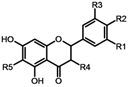
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.1. ABTS, DPPH, FRAP, the inhibition of peroxynitrite induced tyrosine nitration
| ABTS | DPPH | FRAP | Inhibition of. peroxynitrite induced tyrosine nitration | Superoxide scavenging activity | ||
|---|---|---|---|---|---|---|
| Enzymatic | Non-enzymatic | |||||
| 1 | 3.2 ± 0.01 | 1.06 ± 0.04 | 0.522 ± 0.01 | 0.84 ± 0.01 | 45.2 | 25.9 |
| 2 | 1.7 ± 0.01 | 0.02 ± 0.01 | 0.051 ± 0.00 | 0.09 ± 0.01 | -a | -a |
| 3 | 1.4 ± 0.00 | 0.12 ± 0.02 | 0.118 ± 0.00 | 0.80 ± 0.03 | -a | -a |
| 4 | 1.61 ± 0.01 | 0.14 ± 0.04 | -a | 0.82 ± 0.01 | -a | -a |
| 5 | 1.62 ± 0.01 | 0.10 ± 0.00 | 0.127 ± 0.01 | 0.74 ± 0.01 | -a | -a |
| 6 | 1.66 ± 0.01 | 0.98 ± 0.03 | 0.741 ± 0.01 | 0.84 ± 0.01 | 71.2 | 29.5 |
| 7 | 1.60 ± 0.01 | 0.29 ± 0.02 | 1.189 ± 0.06 | 0.83 ± 0.02 | -a | -a |
| 8 | 0.97 ± 0.03 | 0.12 ± 0.00 | -a | 0.82 ± 0.02 | -a | -a |
| 9 | -a | -a | -a | 0.93 ± 0.02 | -a | -a |
| Rutin | -a | -a | -a | -a | 50.2 | 43.6 |
2.2. Superoxide scavenging activity assay
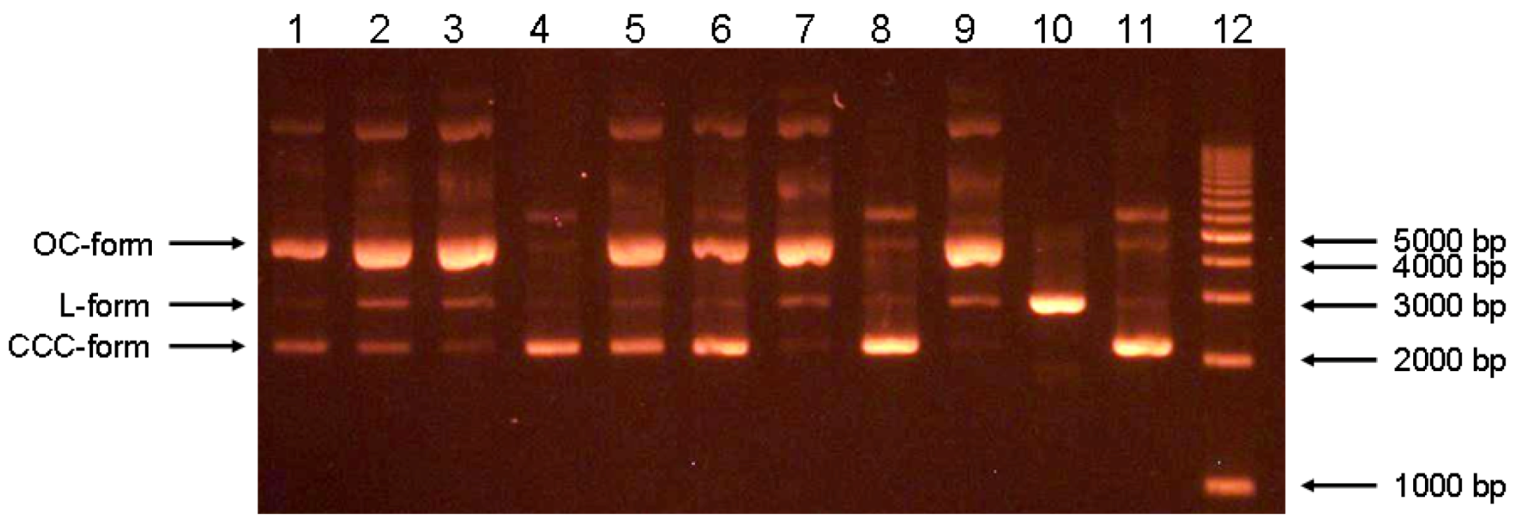
2.3. Inhibition of Fenton reaction assay
| Compound | AUC | Ratio CCC/ (OC+L) | Compound ratio/Rutin ratio | ||
|---|---|---|---|---|---|
| CCC | OC | L | |||
| 1 (50:1) | 569 | 1618 | 98 | 0.332 | 0.892 |
| 1 (10:1) | 453 | 3272 | 458 | 0.121 | 0.135 |
| 3 (50:1) | 708 | 3271 | 231 | 0.202 | 0.543 |
| 3 (10:1) | 229 | 3170 | 294 | 0.066 | 0.073 |
| 7 (50:1) | 514 | 2854 | 193 | 0.169 | 0.454 |
| 7 (10:1) | 564 | 2173 | 73 | 0.251 | 0.279 |
| Rutin (50:1) | 1288 | 3207 | 254 | 0.372 | 1 |
| Rutin (10:1) | 1942 | 1955 | 207 | 0.898 | 1 |
| Natural plasmid | 2790 | 386 | 193 | 4.819 | - |
| 2 (50:1) | 439 | 634 | - | 0.692 | 0.553 |
| 2 (10:1) | 152 | 1502 | - | 0.101 | 0.199 |
| 6 (50:1) | 1289 | 402 | - | 3.206 | 2.565 |
| 6 (10:1) | 656 | 1406 | - | 0.467 | 0.921 |
| 10 (50:1) | 194 | 1556 | 96 | 0.117 | 0.094 |
| 10 (10:1) | 189 | 1915 | 122 | 0.093 | 0.183 |
| Rutin (50:1) | 1055 | 843 | - | 1.251 | 1 |
| Rutin (10:1) | 806 | 1591 | - | 0.507 | 1 |
| Natural plasmid | 1507 | - | - | - | - |
| 5 (50:1) | 505 | 1379 | - | 0.366 | 0.491 |
| 5 (10:1) | - | 1373 | - | - | - |
| Rutin (50:1) | 1155 | 1493 | 57 | 0.745 | 1 |
| Rutin (10:1) | 535 | 1788 | 47 | 0.292 | 1 |
| Natural plasmid | 2166 | 258 | - | 8.395 | - |
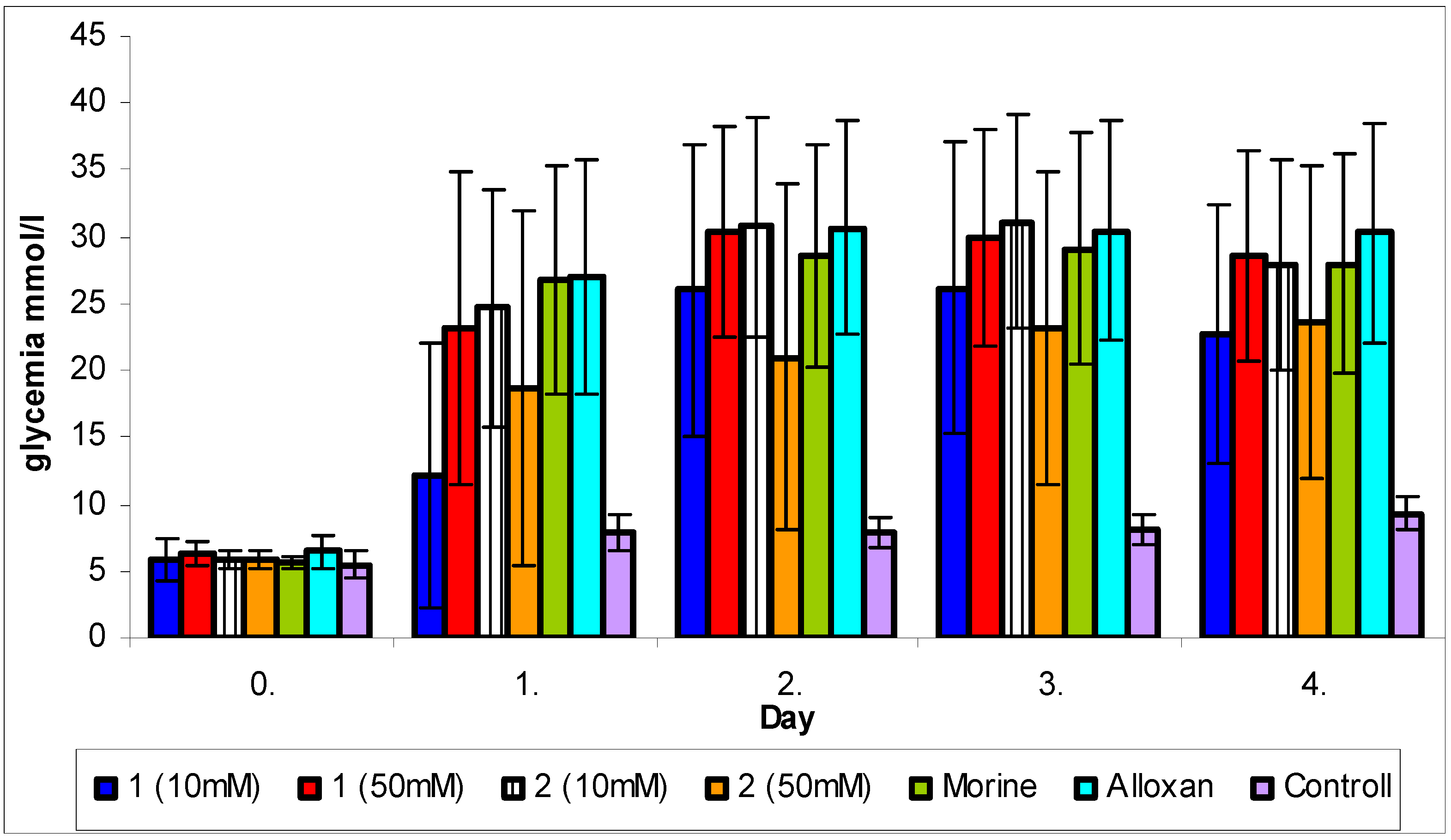
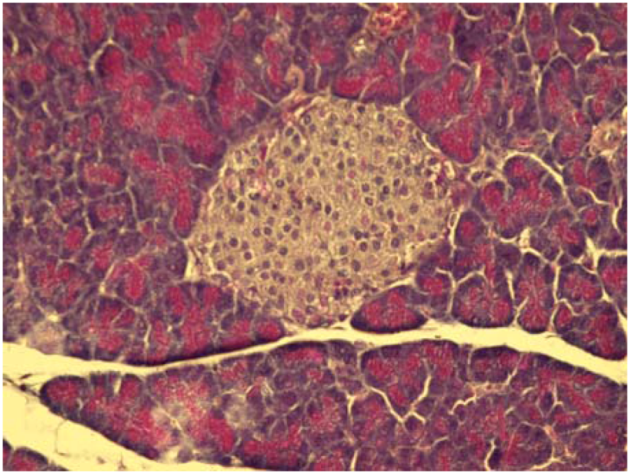
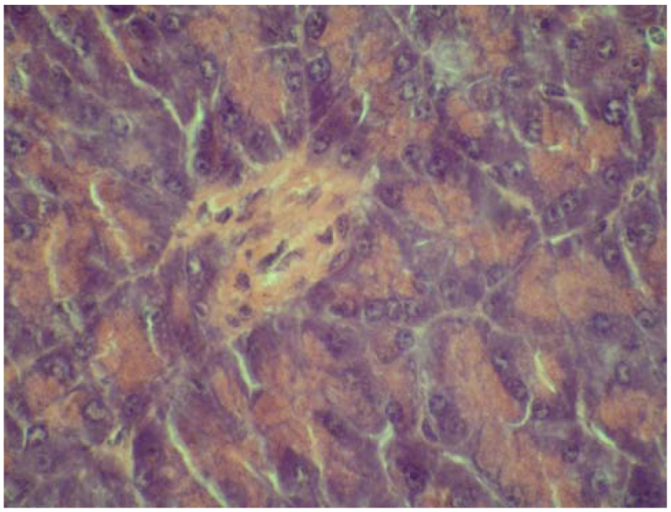
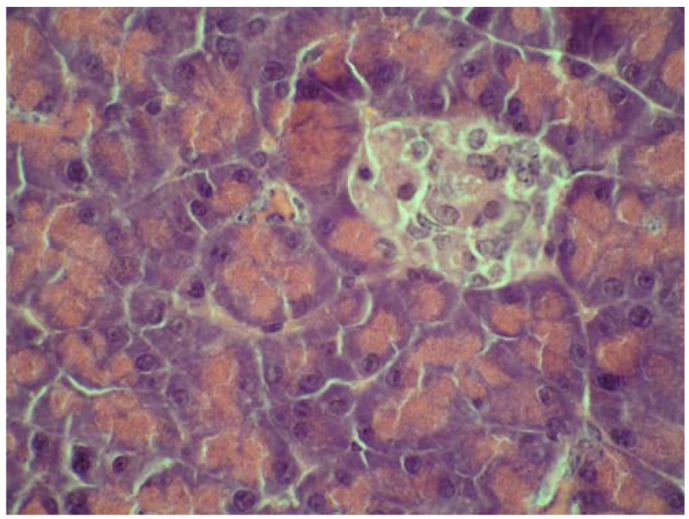
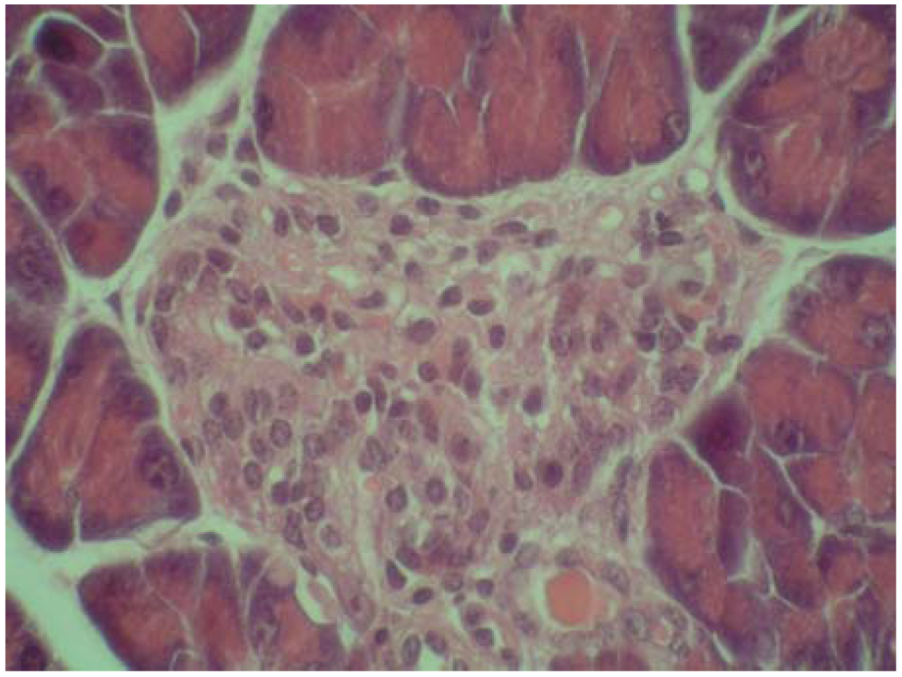
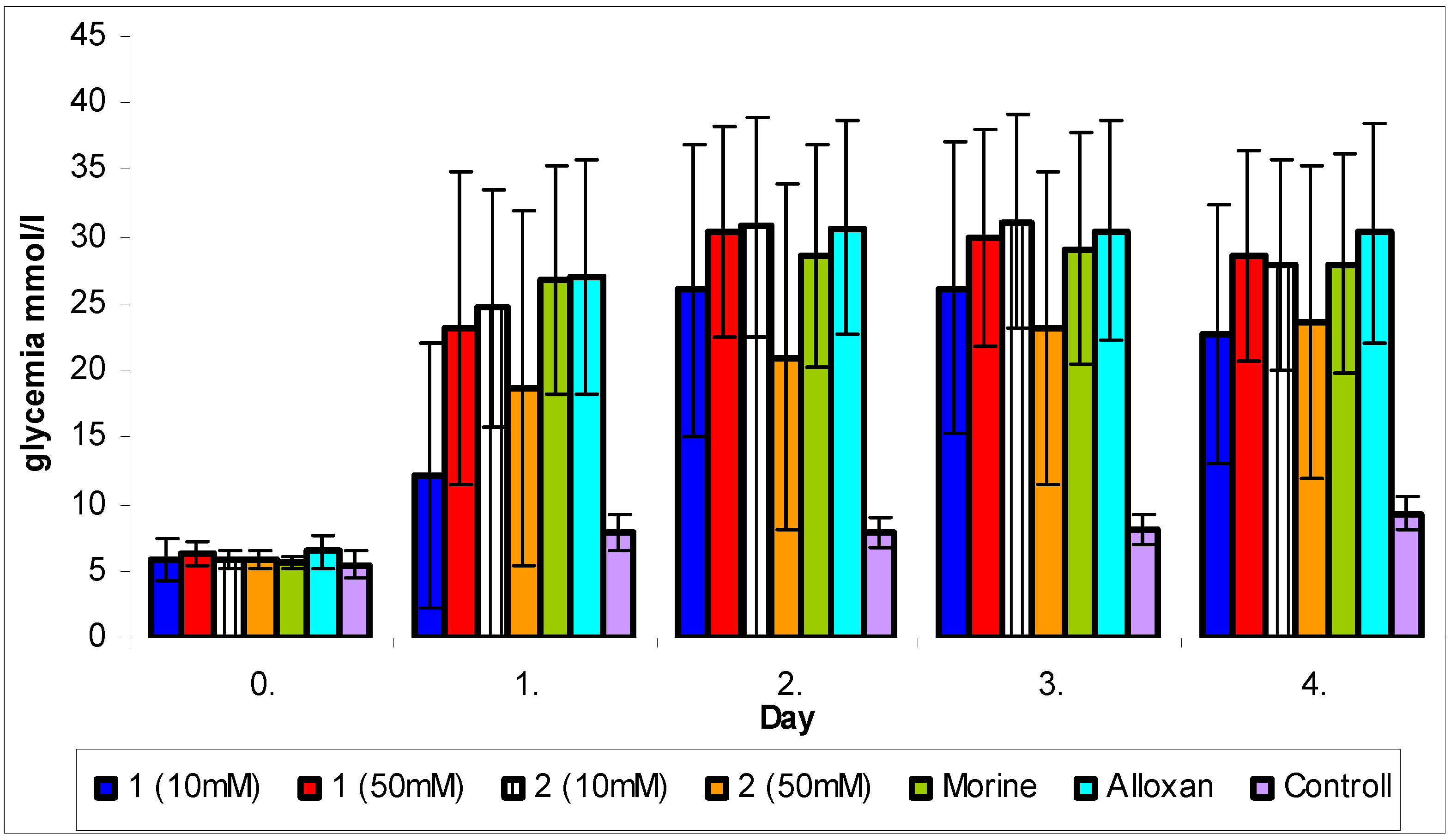
2.4. Cytoprotective effect against alloxan-induced diabetes
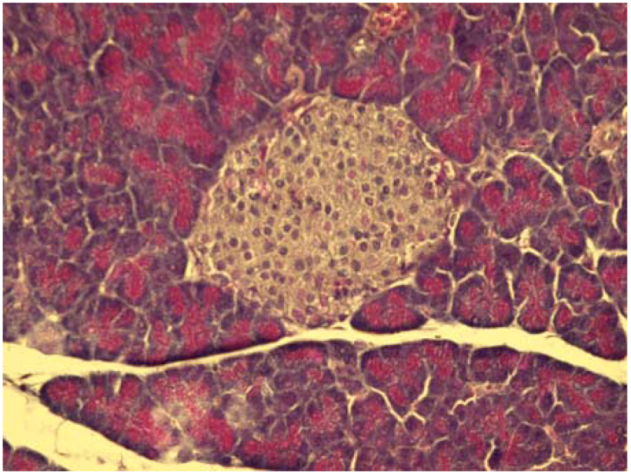
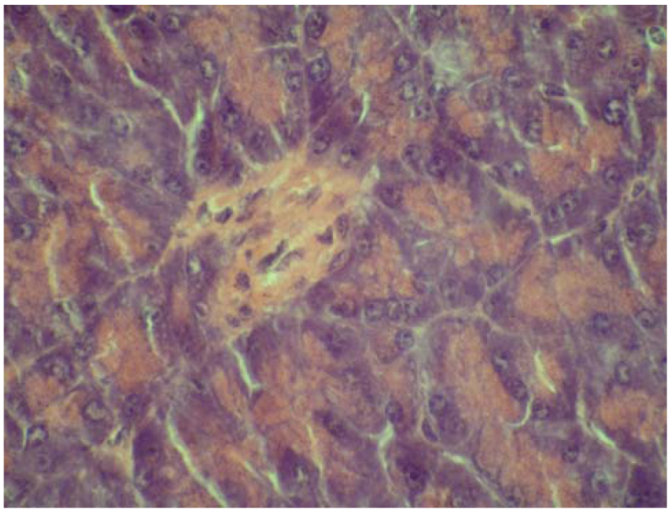
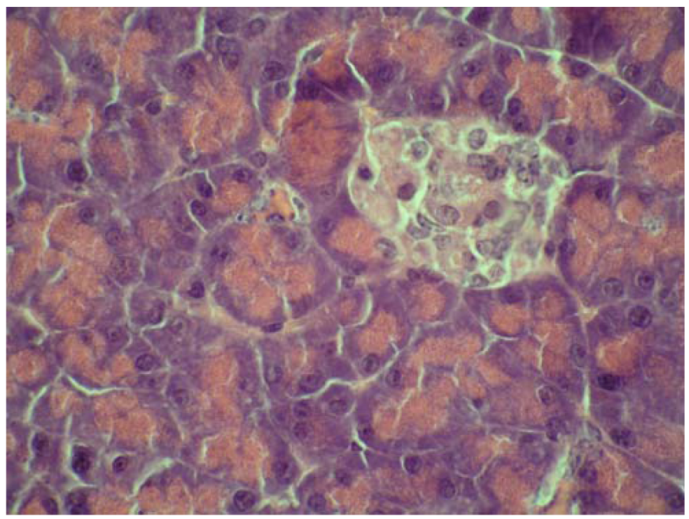
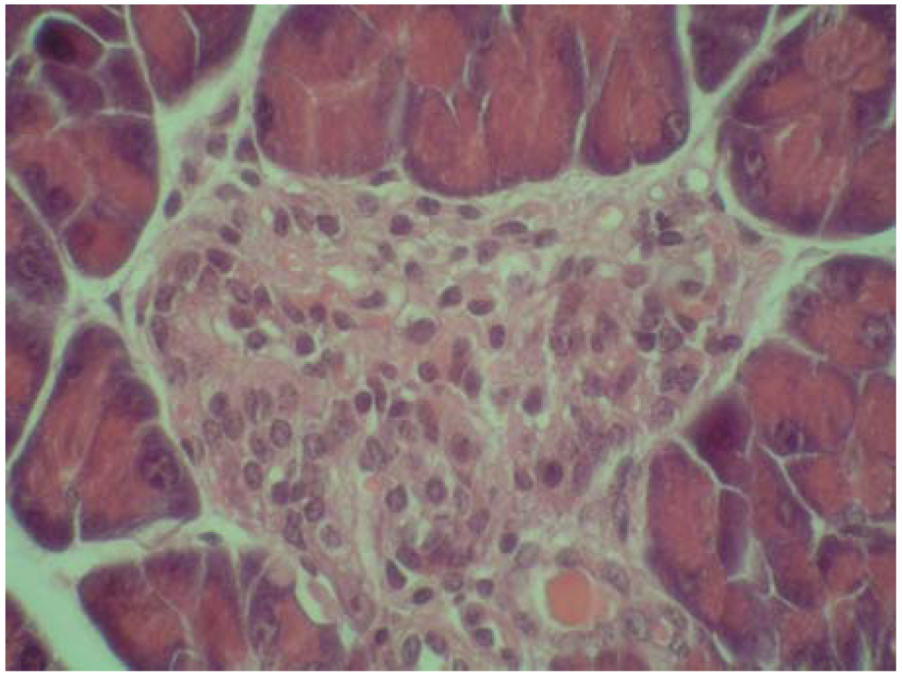
3. Experimental
3.1. Tested compounds
3.2. ABTS and DPPH scavenging activity
3.3.FRAP
3.4. Inhibition of peroxynitrite induced tyrosine nitration
3.5. Superoxide scavenging activity
3.5.1. Enzymatic assay
3.5.2. Non-enzymatic assay
3.6. Inhibition Fenton reaction assay
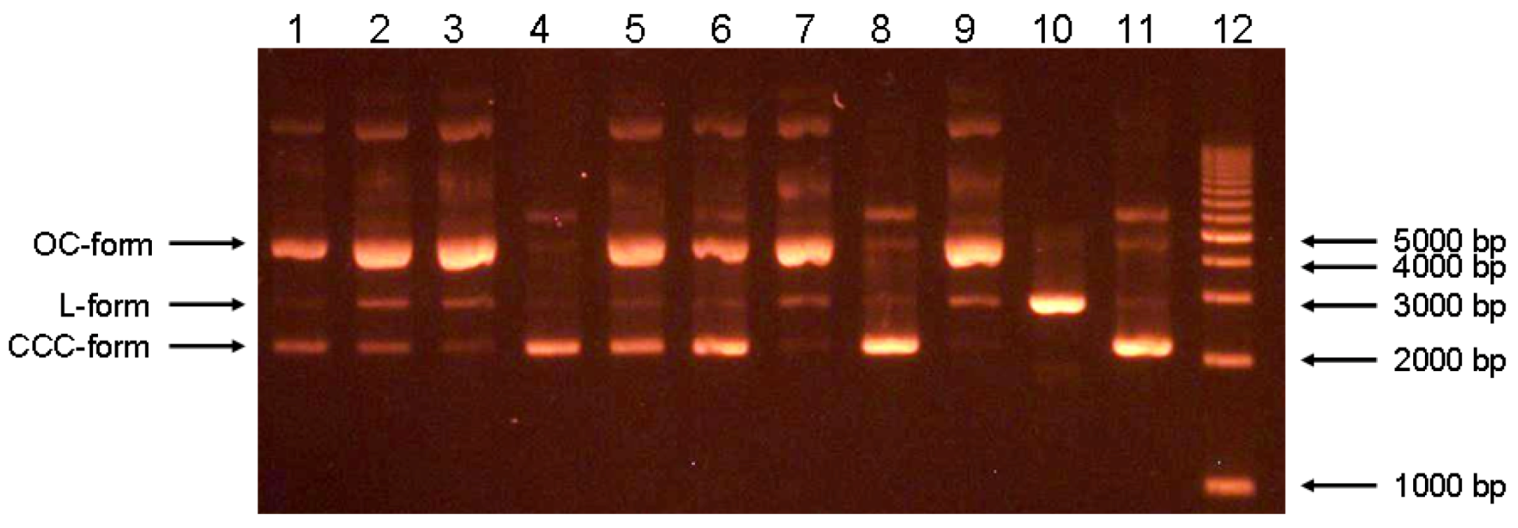
3.7. Cytoprotective effect against alloxan-induced diabetes
4. Conclusions
Acknowledgements
References and Notes
- Cadenas, E. Biochemistry of oxygen toxicity. Ann. Rev. Biochem. 1989, 58, 79–110. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar]
- Mates, J.M.; Perez-Gomez, C.; De Castro, I.N. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- McCall, M.R.; Frei, B. Can antioxidant vitamins materially reduce oxidative damage in humans? Free Radical Biol. Med. 1999, 26, 1034–1053. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Šmejkal, K.; Holubová, P.; Zima, A; Muselík, J.; Dvorská, M. Antiradical activity of Paulownia tomentosa (Scrophulariaceae) extracts. Molecules 2007, 12, 1210–1219. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure–antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Šmejkal, K.; Grycová, L.; Marek, R.; Jankovská, D.; Forejtníková, H.; Vančo, J.; Suchý, V. C-geranyl compounds from Paulownia tomentosa fruits. J. Nat. Prod. 2007, 70, 1244–1248. [Google Scholar] [CrossRef]
- Šmejkal, K.; Chudík, S.; Klouček, P.; Marek, R.; Cvačka, J.; Urbanová, M.; Julínek, O.; Kokoška, L.; Šlapetová, T.; Holubová, P.; Zima, A.; Dvorská, M. Antibacterial C-geranyl flavonoids from Paulownia tomentosa fruits. J. Nat. Prod. 2008, 71, 706–709. [Google Scholar]
- Asai, T.; Hara, N.; Kobayashi, S.; Kohshima, S.; Fujimoto, Y. Geranylated flavanones from the secretion on the surface of the immature fruits of Paulownia tomentosa. Phytochemistry 2008, 69, 1234–1241. [Google Scholar] [CrossRef]
- Van den Berg, R.; Haenen, G.R.M.M.; Van den Berg, H.; Bast, A. Applicability of an improved Trolox equivalent antioxidant capacity (TEAC) assay for evaluation of antioxidant capacity measurements of mixtures. Food Chem. 1999, 66, 511–517. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: structure-activity relationships. Free Radical Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Sekher Pannala, A.; Chan, T.S.; O’Brien, P.J.; Rice-Evans, C.A. Flavonoid B-ring chemistry and antioxidant activity: fast reaction kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef]
- Dugas, A.J., Jr.; Castaneda-Acosta, J.; Bonin, G.C.; Price, K.L.; Fischer, N.H.; Winston, G.W. Evaluation of the total peroxyl radical scavenging capacity of flavonoids: structure-activity relationships. J. Nat. Prod. 2000, 63, 327–331. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: chemistry, metabolism and structure activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Firuzi, O.; Lacanna, A.; Petrucci, R.; Marrosu, G.; Saso, L. Evaluation of the antioxidant activity of flavonoids by “ferric reducingantioxidant power” assay and cyclic voltammetry. Biochim. Biophys. Acta 2005, 1721, 174–184. [Google Scholar] [CrossRef]
- Muselík, J.; García-Alonso, M.; Martín-López, M.P.; Žemlička, M.; Rivas-Gonzalo, J.C. Measurement of antioxidant activity of wine catechins, procyanidins, anthocyanins and pyranoanthocyanins. Int. J. Mol. Sci. 2007, 8, 797–809. [Google Scholar] [CrossRef]
- Cos, P.; Li, Y.; Calomme, M.; Jia, P.H.; Cimanga, K.; Van Poel, B.; Pieters, L; Vlietinck, A.J.; Van den Berghe, D. Structure-Activity Relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef]
- Borges, F.; Fernandes, E. Progress towards the discovery of XO inhibitors. Curr. Med. Chem. 2002, 24, 195–217. [Google Scholar] [CrossRef]
- Yao, H.; Liao, Z.X.; Wu, Q; Lei, G.Q.; Liu, Z.J.; Chen, D.F; Chen, J.K.; Zhou, T.S. Antioxidative flavanone glycosides from the branches and leaves of Viscum coloratum. Chem. Pharm. Bull. 2006, 54, 133–135. [Google Scholar] [CrossRef]
- Valentaõ, P.; Fernandes, E.; Carvalho, F.; Andrade, P.B.; Seabra, R.M.; Bastos, M.L. Antioxidant activity of Centaurium erythraea infusion evidenced by its superoxide radical scavenging and xanthine oxidase inhibitory activity. J. Agric. Food Chem. 2001, 49, 3476–3479. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C. Iron promoters of the Fenton reaction and lipid peroxidation can be released from haemoglobin by peroxides. FEBS Lett. 1986, 201, 291–295. [Google Scholar] [CrossRef]
- Kang, J.H. Oxidative damage of DNA by the reaction of amino acid with methylglyoxal in the presence of Fe(III). Int. J. Biol. Macromol. 2003, 33, 43–48. [Google Scholar] [CrossRef]
- Prakash, D.; Suri, S.; Upadhyay, G.; Singh, B.N. Total phenol, antioxidant and free radical scavenging activities of some medicinal plants. Int. J. Food Sci. Nutr. 2007, 58, 18–28. [Google Scholar] [CrossRef]
- Szkudelski, T. The Mechanism of Alloxan and Streptozocin Action in B Cells of the Rat Pancreas. Physiol. Res. 2001, 50, 536–546. [Google Scholar]
- Rahimi, R.; Nikfar, S.; Larijani, B.; Abdollahi, M. A review on the role of antioxidants in the management of diabetes and its complications. Biomed. Pharmacother. 2005, 59, 365–373. [Google Scholar] [CrossRef]
- Soto, C.; Recoba, R.; Barron, H.; Alvarez, C.; Favari, L. Silymarin increases antioxidant enzymes in alloxan-induced diabetes in rat pancreas. Comp. Biochem. Physiol. - Part C Toxicol. Pharmacol. 2003, 136, 205–212. [Google Scholar]
- Rice-Evans, C; Packer, P. Flavonoids in Health and Disease, 2nd ed; CRC Press: New York, NY, USA, 2003; pp. 329–334. [Google Scholar]
- Šmejkal, K.; Svačinová, J.; Šlapetová, T.; Schneiderová, K.; Dall’Acqua, S.; Innocenti, G.; Závalová, V.; Kollár, P.; Chudík, S.; Marek, R.; Julínek, O.; Urbanová, M.; Kartal, M.; Csöllei, M.; Doležal, K. Cytotoxic activities of several geranyl-substituted flavanones. J. Nat. Prod. 2010, 73, 568–572. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of „antioxidant power“: the FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar]
- Vančo, J.; Marek, J.; Trávníček, Z.; Račanská, E.; Muselík, H.; Švajnelová, O. Synthesis, structural characterization, antiradical and antidiabetic activities of copper(II) and zinc(II) Schiff base complexes derived from salicylaldehyde and β-alanine. J. Inorg. Biochem. 2008, 102, 595–605. [Google Scholar] [CrossRef]
- Subash, S.; Subramanian, P. Morin a flavonoid exerts antioxidant potential in chronic hyperammonemic rats: a biochemical and histopathological study. Mol. Cell Biochem. 2009, 327, 153–161. [Google Scholar] [CrossRef]
- Sample Availability: Samples of compounds 1-8 are available from the authors.
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zima, A.; Hošek, J.; Treml, J.; Muselík, J.; Suchý, P.; Pražanová, G.; Lopes, A.; Žemlička, M. Antiradical and Cytoprotective Activities of Several C-Geranyl-substituted Flavanones from Paulownia tomentosa Fruit. Molecules 2010, 15, 6035-6049. https://doi.org/10.3390/molecules15096035
Zima A, Hošek J, Treml J, Muselík J, Suchý P, Pražanová G, Lopes A, Žemlička M. Antiradical and Cytoprotective Activities of Several C-Geranyl-substituted Flavanones from Paulownia tomentosa Fruit. Molecules. 2010; 15(9):6035-6049. https://doi.org/10.3390/molecules15096035
Chicago/Turabian StyleZima, Aleš, Jan Hošek, Jakub Treml, Jan Muselík, Pavel Suchý, Gabriela Pražanová, Ana Lopes, and Milan Žemlička. 2010. "Antiradical and Cytoprotective Activities of Several C-Geranyl-substituted Flavanones from Paulownia tomentosa Fruit" Molecules 15, no. 9: 6035-6049. https://doi.org/10.3390/molecules15096035
APA StyleZima, A., Hošek, J., Treml, J., Muselík, J., Suchý, P., Pražanová, G., Lopes, A., & Žemlička, M. (2010). Antiradical and Cytoprotective Activities of Several C-Geranyl-substituted Flavanones from Paulownia tomentosa Fruit. Molecules, 15(9), 6035-6049. https://doi.org/10.3390/molecules15096035