A Mangifera indica L. Extract Could Be Used to Treat Neuropathic Pain and Implication of Mangiferin

{kind=link}
Abstract
:Abbreviations
| sGC | soluble guanylyl cyclise |
| PLA2 | phospholipase A2 |
| AA | arachidonic acid |
| COX | cyclooxygenase |
| PKA | protein kinase A |
| P2X4 | purinergic P2 receptor |
| CX3CR1 | chemokine fractalkine receptor |
| NMDA | N-methyl-D-aspartate receptor |
| AMPA | α-amino-3-hydroxy-5-methylisoxazole-4-propionic |
| mGluRs | metabotropic glutamate receptors |
| NK1 | tachykinin NK1 receptor |
| TNF-R | TNF receptor |
1. Introduction
2. The Antinociceptive Action of MSBE in Formalin-induced Licking and Acetic acid-induced Writhing Models in Mice
3. The Effects of MSBE and Mangiferin in Different in Vitro and in Vivo Models of Inflammation
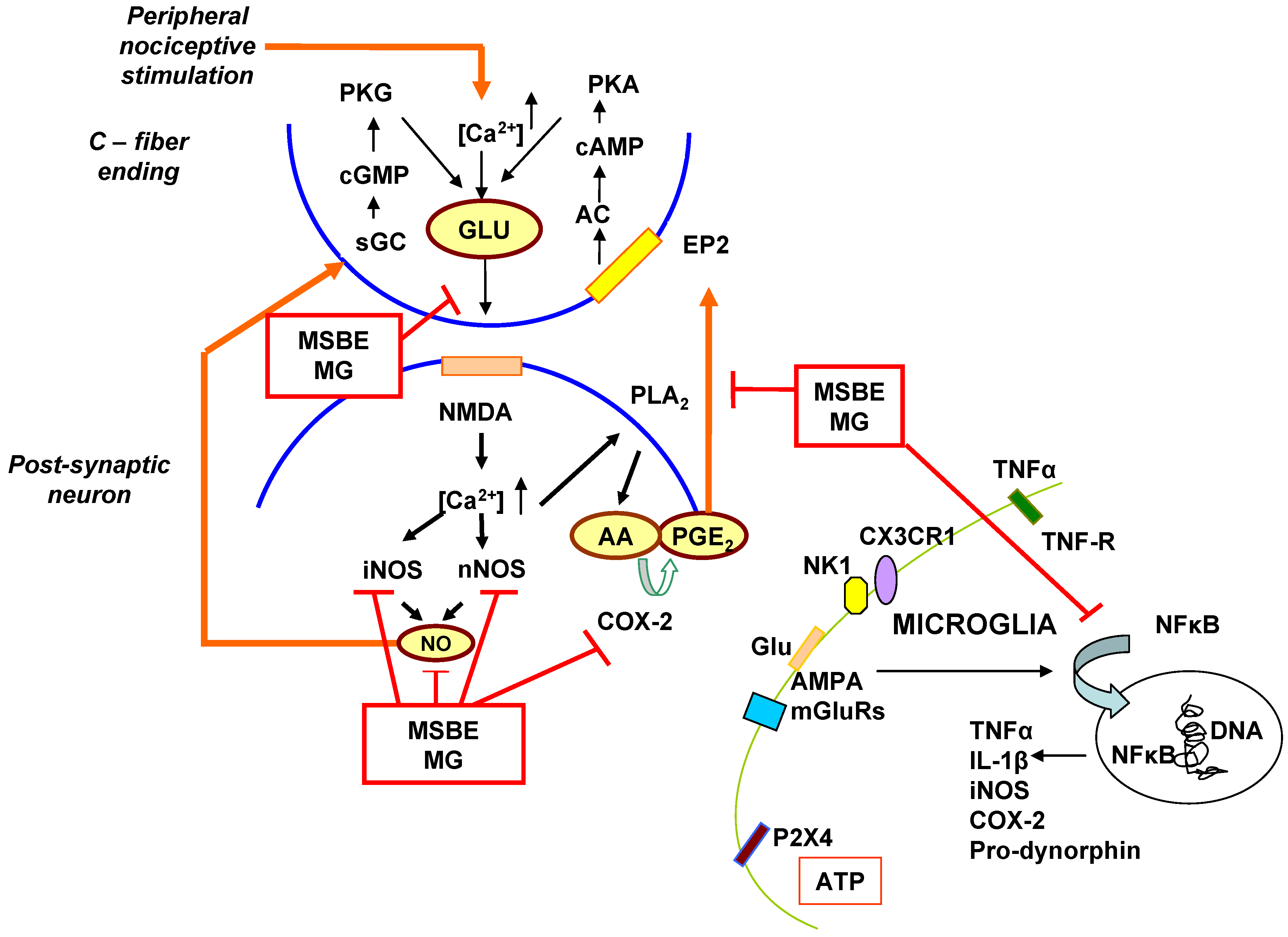
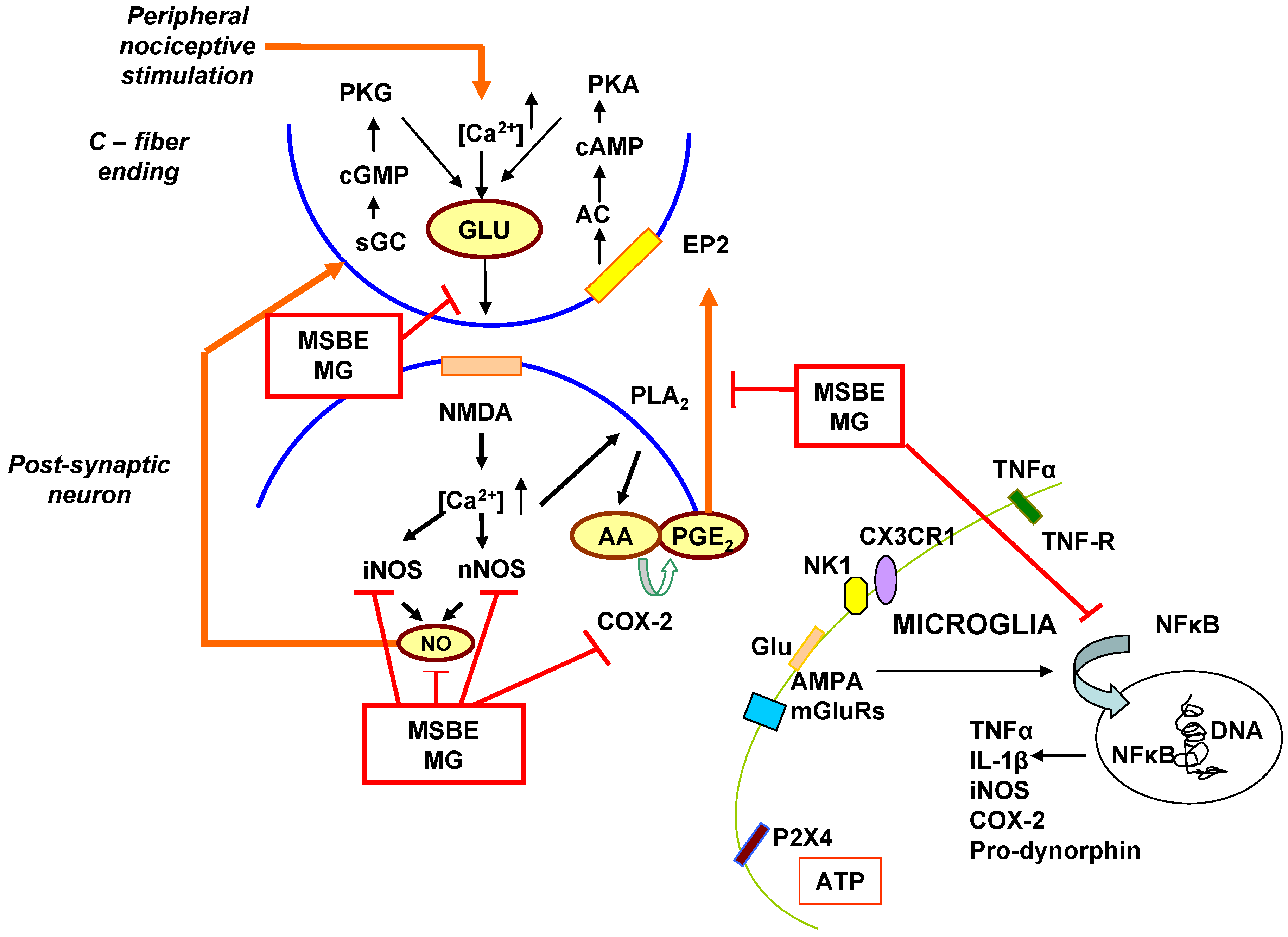
4. The Possible Role of MSBE and Mangiferin as Modulators of the Central Sensitization Mechanism
5. Conclusions
References
- Zebrowsky, M. Patient potential. In The Pain Market Outlook to 2011; Business Insights Ltd.: London, UK, 2006; pp. 16–18. [Google Scholar]
- Twining, C.M.; Sloane, E.M.; Milligan, E.D.; Chacur, M.; Martin, D.; Pooles, S.; Marsh, H.; Maier, S.F.; Watkins, L.R. Peri-sciatic proinflammatory cytokines, reactive oxygen species and complement induce mirror-image neuropathic pain in rats. Pain 2004, 110, 299–309. [Google Scholar] [CrossRef]
- Manning, D.C. The role of neuroimmune activation in chronic neuropathic pain and new targets for therapeutic intervention. In Emerging Strategies for the Treatment of Neuropathic Pain; Campbell, J.N., Basbaum, A.I., Dray, A., Dubner, R., Dworkin, R.H., Sang, Ch.N., Eds.; IASP Press: Seattle, WA, USA, 2006; pp. 161–192. [Google Scholar]
- De Leo, J.A.; Tawfik, V.L.; La Croix-Fralish, M.L. The tetrapartite synapse: Path to CNS sensitization and chronic pain. Pain 2006, 122, 17–21. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, S.K.; Zhou, J.L.; Taglialatela, G.; Chung, K.; Coggeshall, R.E.; Chung, J.M. Reactive oxygen species (ROS) play an important role in a rat model of neuropathic pain. Pain 2004, 111, 116–124. [Google Scholar] [CrossRef]
- Naik, A.K.; Tandan, S.K.; Dudhgaonkar, S.P.; Jadhav, S.H.; Kataria, M.; Prakash, V.R.; Kumar, D. Role of oxidative stress in pathophysiology of peripheral neuropathy and modulation by N-acetyl-L-cysteine in rats. Eur. J. Pain 2006, 10, 573–579. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, J.H.; Gao, X.; Zhou, J.L.; Lee, I.; Chung, K.; Chung, J.M. Analgesic effect of vitamin E is mediated by reducing central sensitization in neuropathic pain. Pain 2006, 122, 53–62. [Google Scholar] [CrossRef]
- Gao, X.; Kim, H.K.; Chung, J.M.; Chung, K. Reactive oxygen species (ROS) are involved in enhancement of NMDA-receptor phosphorylation in animal models of pain. Pain 2007, 131, 262–271. [Google Scholar] [CrossRef]
- Xu, J.T.; Xin, W.J.; Zhang, Y.; Wu, Ch.Y.; Liu, X.G. The role of the tumor necrosis factor-alpha in the neuropathic pain induced by Lumbar 5 ventral root transection in rat. Pain 2006, 123, 306–321. [Google Scholar] [CrossRef]
- Zelenka, M.; Schäfers, M.; Sommer, C. Intraneural injection of interleukin-1β and TNFα into rat sciatic nerve at physiological doses induces signs of neuropathic pain. Pain 2005, 116, 257–263. [Google Scholar] [CrossRef]
- Quintão, N.L.M.; Balz, D.; Santos, A.R.S.; Campos, M.M.; Calixto, J.B. Long-lasting neuropathic pain induced by brachial plexus injury in mice: Role triggered by pro-inflammatory cytokine, tumor necrosis factor α. Neuropharmacology 2006, 50, 614–620. [Google Scholar] [CrossRef]
- Lin, Y.L.; Zhou, L.J.; Hu, N.W.; Xu, J.T.; Chang, Y.W.; Zhang, T.; Li, Y.Y.; Liu, X.G. Tumor necrosis factor α induces long-term potentiation of C-fiber evoked field potentials in spinal dorsal horn in rats with nerve injury: the role of NF-kappa B, JNK and p38 MAPK. Neuropharmacology 2007, 52, 708–715. [Google Scholar] [CrossRef]
- Wei, X.H.; Zhang, Y.; Wu, C.Y.; Xu, J.T.; Xin, W.J.; Liu, X.G. Pri-sciatic administration of recombinant rat TNF-α induces mechanical allodynia via upregulation of TNF-α in dorsal root ganglia and in spinal dorsal horn: The role of NF-kappa B pathway. Exp. Neurol. 2007, 205, 471–484. [Google Scholar] [CrossRef]
- Milligan, E.D.; Ledeboer, A.; Sloane, E.M.; Maier, S.M.; Busha, D.A.; Watkins, L.R. Glially driven enhancement of pain and its control by anti-Inflammatory cytokines. In Immune and Glial Regulation of Pain; De Leo, J.A., Sorkin, L.S., Watkins, L.R., Eds.; IASP Press: Seattle, WA, USA, 2007; pp. 407–415. [Google Scholar]
- Núñez-Sellés, A.J.; Delgado-Hernández, R.; Garrido-Garrido, G.; González-Rivera, D.; Guevara-García, M.; Pardo-Andreu, G.L. The paradox of natural products as pharmaceutical. Experimental evidences of a mango stem bark extract. Pharmacol. Res. 2007, 55, 351–358. [Google Scholar] [CrossRef]
- Núñez-Sellés, A.J.; Páez-Betancourt, E.; Amaro-González, D.; Acosta-Esquijarosa, J.; Agüero- Agüero, J.; Capote-Hernández, R.; Gárciga-Hernández, M.R.; Morales-Lacárrere, I.G.; García-Pulpeiro, O.; Garrido-Garrido, G.; Martinez-Sánchez, G.; Angel Morales-Segura, M. Composiciones Farmacéuticas y suplementos nutricionales a partir de extractos de Mangifera indica L. Oficina Cubana de la Propiedad Industrial. Patent 203/98, 30 October 2002. [Google Scholar]
- Martínez, G.; Re, L.; Giuliani, A.; Núñez-Sellés, A.J.; Pérez, G.; León, O.S. Protective effects of Mangifera indica L. extract, mangiferin, and selected antioxidants against TPA-induced biomolecules oxidation and peritoneal macrophage activation in mice. Pharmacol. Res. 2000, 42, 565–573. [Google Scholar] [CrossRef]
- Garrido, G.; González, D.; Delporte, C.; Backhouse, N.; Quintero, G.; Núñez, A.J.; Morales, M.A. Analgesic and anti-inflammatory effects of Mangifera indica L. extract (Vimang). Phytother. Res. 2001, 15, 18–21. [Google Scholar] [CrossRef]
- Garrido, G.; González, D.; Lemus, Y.; García, D.; Lodeiro, L.; Quintero, G.; Delporte, C.; Núñez-Sellés, A.J.; Delgado, R. In vivo and in vitro anti-inflammatory activity of Mangifera indica L. extract (VIMANG®). Pharmacol. Res. 2004, 50, 143–149. [Google Scholar] [CrossRef]
- García, D.; Delgado, R.; Ubeira, F.M.; Leiro, J. Modulator effects of rat macrophage function by Mangifera indica L extract (Vimang) and mangiferin. Int. Immunopharmacol. 2002, 2, 797–806. [Google Scholar] [CrossRef]
- García, D; Leiro, J.; Delgado, R.; San Martín, M.L.; Ubeira, F.M. Mangifera indica L. extract (Vimang) and mangiferin modulate mouse humoral immune response. Phytother. Res. 2003, 17, 1182–1187. [Google Scholar] [CrossRef]
- Garrido, G.; Blanco, M.; Sancho, R.; Macho, A.; Delgado, R.; Muñoz, E. An aqueous stem bark extract of Mangifera indica (Vimang®) inhibits T cell proliferation and TNF-induced activation of nuclear transcription factor NF-kappa B. Phytother. Res. 2005, 19, 211–215. [Google Scholar] [CrossRef]
- Leiro, J.; García, D.; Arranz, J.A.; Delgado, R.; Sanmartín, M.L.; Orallo, F. An Anacardiaceae preparation reduces the expression of inflammation-related genes in murine macrophages. Int. Immunopharmacol. 2004, 4, 991–1003. [Google Scholar] [CrossRef]
- Leiro, J.; Arranz, J.A.; Yáñez, M.; Ubeira, F.M.; Sanmartín, M.L.; Orallo, F. Expression profiles of genes involved in the mouse nuclear factor-kappa B signal transduction pathway are modulated by mangiferin. Int. Immunopharmacol. 2004, 4, 763–778. [Google Scholar] [CrossRef]
- Martínez, G.; Giuliani, A.; León, O.S.; Pérez, G.; Núñez, A.J. Effect of Mangifera indica L. extract (Vimang) on protein and hepatic microsome peroxidation. Phytother. Res. 2001, 15, 581–585. [Google Scholar] [CrossRef]
- Martínez, G.; Candelario, E.; Giuliani, A.; León, O.S.; Ram, S.; Delgado, R.; Núñez, A.J. Mangifera indica L. extract (Vimang) reduces ischaemia-induced neuronal loss and oxidative damage in gerbil brain. Free Radic. Res. 2001, 35, 465–473. [Google Scholar] [CrossRef]
- Campos-Esparza, M.R.; Sánchez-Gómez, M.V.; Matute, C. Molecular mechanisms of neuroprotection by two natural antioxidants polyphenols. Cell Calcium 2009, 45, 358–368. [Google Scholar] [CrossRef]
- Gottlied, M.; Leal-Campanario, R.; Campos-Esparza, M.R.; Sánchez-Gómez, M.V.; Alberdi, E.; Arranz, A.; Delgado-García, J.M.; Gruart, A.; Matute, C. Neuroprotection by two polyphenols following excitotoxicity and experimental ischemia. Neurobiol. Dis. 2006, 23, 374–386. [Google Scholar] [CrossRef]
- Pardo-Andreu, G.L.; Barrios, M.F.; Curti, C.; Hernandez, I.; Merino, N.; Lemus, Y.; Martínez, I.; Riaño, A.; Delgado, R. Protective effects of Mangifera indica L extract (Vimang), and its major component mangiferin, on iron-induced oxidative damage to rat serum and liver. Pharmacol. Res. 2008, 57, 79–86. [Google Scholar] [CrossRef]
- Bhatia, H.S.; Candelario-Jalil, E.; Pinheiro de Oliveira, A.C.; Olajide, O.A.; Martínez-Sánchez, G.; Fiebich, B.L. Mangiferin inhibits cyclooxygenase-2 expression and prostaglandin E2 production in activated rat microglial cells. Arch. Biochem. Biophys. 2008, 477, 253–258. [Google Scholar] [CrossRef]
- Salter, M.W. Signaling pathways in pain neuroplasticity in the spinal dorsal horn. In Emerging Strategies for the Treatment of Neuropathic Pain; Campbell, J.N., Basbaum, A.I., Dray, A., Dubner, R., Dworkin, R.H., Sang, Ch.N., Eds.; IASP Press: Seattle, WA, USA, 2006; pp. 161–183. [Google Scholar]
- Bennett, G.J. Experimental neuropathic pain in animals. Models and mechanisms. In Pain-2005 an Updated Review: Refresher Course Syllabus; Justins, D.M., Ed.; IAPS Press: Seattle, WA, USA, 2005; pp. 97–105. [Google Scholar]
- Garrido-Suárez, B.; Rabí, M.C.; Bosch, F.; Garrido-Garrido, G.; Delgado-Hernández, R. Introducción de la suplementación con formulaciones Vimang en el Síndrome Doloroso Regional Complejo: experiencia en 15 pacientes. Rev. Soc. Esp. Dolor 2009, 16, 87–96. [Google Scholar]
- Alvarez, A.; Guevara, M.; Riaño, A.; Garrido, G.; Delgado, R. Eficacia del tratamiento con extracto de Mangifera indica L (VIMAG: crema 1,2% y tabletas 150 mg) sobre el Herpes Zoster. Presentación de un caso. Rev. Cubana Farm. 2002, 36, 171–173. [Google Scholar]
- Coderre, T.J.; Vaccarino, A.L.; Melzack, R. Central nervous system plasticity in the tonic pain response to subcutaneous formalin injection. Brain Res. 1990, 535, 155–158. [Google Scholar] [CrossRef]
- Hacimuftuoglu, A.; Handy, C.R.; Goettl, V.M.; Lin, C.G.; Dane, S.; Stephens, R.L., Jr. Antioxidants attenuate multiple phases of formalin-induced nociceptive response in mice. Behav. Brain Res. 2006, 173, 211–216. [Google Scholar] [CrossRef]
- Okuda, K.; Sakurada, Ch.; Takahashi, M.; Yamada, T.; Sakurada, T. Characterization of nociceptive responses and spinal releases of nitric oxide metabolites and glutamate evoked by different concentrations of formalin in rats. Pain 2001, 92, 107–115. [Google Scholar] [CrossRef]
- Vetter, G.; Geisslinger, G.; Tegeder, I. Release of glutamate, nitric oxide and prostaglandin E2 and metabolic activity in the spinal cord of rats following peripheral nociceptive stimulation. Pain 2001, 92, 213–218. [Google Scholar] [CrossRef]
- Xu, L.; Mabuchi, T.; Katano, T.; Matsumura, S.; Okuda-Ashitaka, E.; Sakimura, K.; Mishina, M.; Ito, S. Nitric oxide (NO) serves as a retrograde messenger to activate neuronal NO synthase in the spinal cord via NMDA receptors. Nitric Oxide 2007, 17, 18–24. [Google Scholar]
- Sawynok, J.; Reid, A. Modulation of formalin-induced behaviours and edema by local and systemic administration of dextromethorphan, memantine and ketamine. Eur. J. Pharmacol. 2002, 450, 153–162. [Google Scholar] [CrossRef]
- Lui, P.W.; Lee, Ch.H. Preemptive effects of intrathecal cyclooxygenase inhibitor or nitric oxide synthase inhibitor on thermal hypersensitivity following peripheral nerve injury. Life Sci. 2004, 75, 2527–2538. [Google Scholar] [CrossRef]
- Fukuoka, T.; Tokunaga, A.; Kondo, E.; Noguchi, K. The role of neighboring intact dorsal root ganglion neurons in a rat neuropathic pain model. In Progress in Pain Research and Management; Devor, M., Rowbotham, M., Wiesenfeld-Hallin, Z., Eds.; IASP Press: Seattle, WA, USA, 2000; pp. 137–146. [Google Scholar]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Kim, H.; Cui, L.; Kim, J.; Kim, S.J. Transient receptor potential vanilloid type 1 receptor regulates glutamatergic synaptic inputs to the spinothalamic tract neurons of the spinal cord deep dorsal horn. Neuroscience 2009, 160, 508–516. [Google Scholar] [CrossRef]
- Watson, C.P.; Tyler, K.L.; Bickers, D.R.; Millikan, L.E.; Smith, S.; Coleman, E.A. Randomized vehicle-controlled trial of topical capsaicin in the treatment of postherpetic neuralgia. Clin. Ther. 1993, 15, 510–526. [Google Scholar]
- Backonja, M.; Wallace, M.S.; Blonsky, E.R.; Cutler, B.J.; Malan, P., Jr.; Rauck, R.; Tobias, J. NGX-4010, a high concentration capsaicin patch, for the treatment of postherpetic neuralgia. A randomised, double-blind study. Lancet Neurol. 2008, 7, 1106–1112. [Google Scholar] [CrossRef]
- Nagy, I.; Sántha, P.; Jancsó, G.; Urbán, L. The role of the vanilloid (capsaicin) receptor (TRPV1) in physiology and pathology. Eur. J. Pharmacol. 2004, 500, 351–369. [Google Scholar] [CrossRef]
- Besson, M.; Brook, P.; Chizh, B.A.; Pickering, A.E. Tactile allodynia in patients with postherpetic neuralgia: Lack of change in skin blood flow upon dynamic stimulation. Pain 2005, 117, 154–161. [Google Scholar] [CrossRef]
- Garrido, G.; Delgado, R.; Lemus, Y.; Rodríguez, J.; García, D.; Núñez, A.J. Protection against septic shock and suppression of tumor necrosis factor alpha and nitric oxide production on macrophages and microglia by a standard aqueous extract of Mangifera indica L. (Vimang®): Role of mangiferin isolated from the extract. Pharmacol. Res. 2004, 50, 165–172. [Google Scholar] [CrossRef]
- Garrido, G.; González, D.; Lemus, Y.; Delporte, C.; Delgado, R. Protective effects of standard extract of Mangifera indica L (Vimang®) against mouse ear edema and its inhibition of eicosanoid production in J774 murine macrophages. Phytomedicine 2006, 13, 412–418. [Google Scholar] [CrossRef]
- Beltrán, A.E.; Ledón, N.; Romay, C.; Sironi, M.; Quintero, G.; Garrido, G.; Delgado, R. Extracto acuoso de Mangifera indica L. (VIMANG) y mangiferina inhiben la expresión de ICAM-1 en células endoteliales estimuladas con citocinas pro-inflamatorias. Rev. Cubana Invest. Biomed. 2003, 22, 164–172. [Google Scholar]
- Varga, A.; Bölcskei, K.; Szöke, É.; Almási, R.; Czéh, G.; Szolcsányi, J.; Pethö, G. Relative roles of protein kinase A and protein kinase C in modulation of Transient Receptor Potential Vanilloid type 1 receptor responsiveness in rat sensory neurons in vitro and peripheral nociceptors in vivo. Neuroscience 2006, 140, 645–657. [Google Scholar] [CrossRef]
- Rajkumar, S.V.; Fonseca, R.; Witzing, T.E. Complete resolution of reflex sympathetic dystrophy with thalidomide treatment. Arch. Intern. Med. 2003, 163, 1487–1488. [Google Scholar] [CrossRef]
- Lemus-Molina, Y.; Sánchez-Gómez, M.V.; Delgado-Hernández, R.; Matute, C. Mangifera indica L extract attenuates glutamate-induced neurotoxicity on rat cortical neurons. Neurotoxicology 2009, 30, 1053–1058. [Google Scholar] [CrossRef]
- Romano, A.; Freudenthal, R.; Merlo, E.; Routtenbert, A. Evolutionary-conserved role of the NF-κB transcription factor in neural plasticity and memory. Eur. J. Neurosci. 2006, 24, 1507–1516. [Google Scholar] [CrossRef]
- Laughlin, T.M.; Bethea, J.R.; Yezierski, R.P.; Wilcox, G.L. Cytokine involvement in dynorphin-induced allodynia. Pain 2000, 84, 159–167. [Google Scholar] [CrossRef]
- Fu, E.S.; Zhang, Y.P.; Sagen, J.; Candiotti, K.A.; Morton, P.D.; Liebl, D.J. Bethea, J.R.; Brambilla, R. Transgenic inhibition of glial NF-kappa B reduces pain behavior and inflammation after peripheral nerve injury. Pain 2010, 148, 509–518. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Garrido-Suárez, B.B.; Garrido, G.; Delgado, R.; Bosch, F.; Rabí, M.d.C. A Mangifera indica L. Extract Could Be Used to Treat Neuropathic Pain and Implication of Mangiferin. Molecules 2010, 15, 9035-9045. https://doi.org/10.3390/molecules15129035
Garrido-Suárez BB, Garrido G, Delgado R, Bosch F, Rabí MdC. A Mangifera indica L. Extract Could Be Used to Treat Neuropathic Pain and Implication of Mangiferin. Molecules. 2010; 15(12):9035-9045. https://doi.org/10.3390/molecules15129035
Chicago/Turabian StyleGarrido-Suárez, Bárbara B., Gabino Garrido, Rene Delgado, Fe Bosch, and María del C. Rabí. 2010. "A Mangifera indica L. Extract Could Be Used to Treat Neuropathic Pain and Implication of Mangiferin" Molecules 15, no. 12: 9035-9045. https://doi.org/10.3390/molecules15129035
APA StyleGarrido-Suárez, B. B., Garrido, G., Delgado, R., Bosch, F., & Rabí, M. d. C. (2010). A Mangifera indica L. Extract Could Be Used to Treat Neuropathic Pain and Implication of Mangiferin. Molecules, 15(12), 9035-9045. https://doi.org/10.3390/molecules15129035