Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate

Abstract
:Introduction
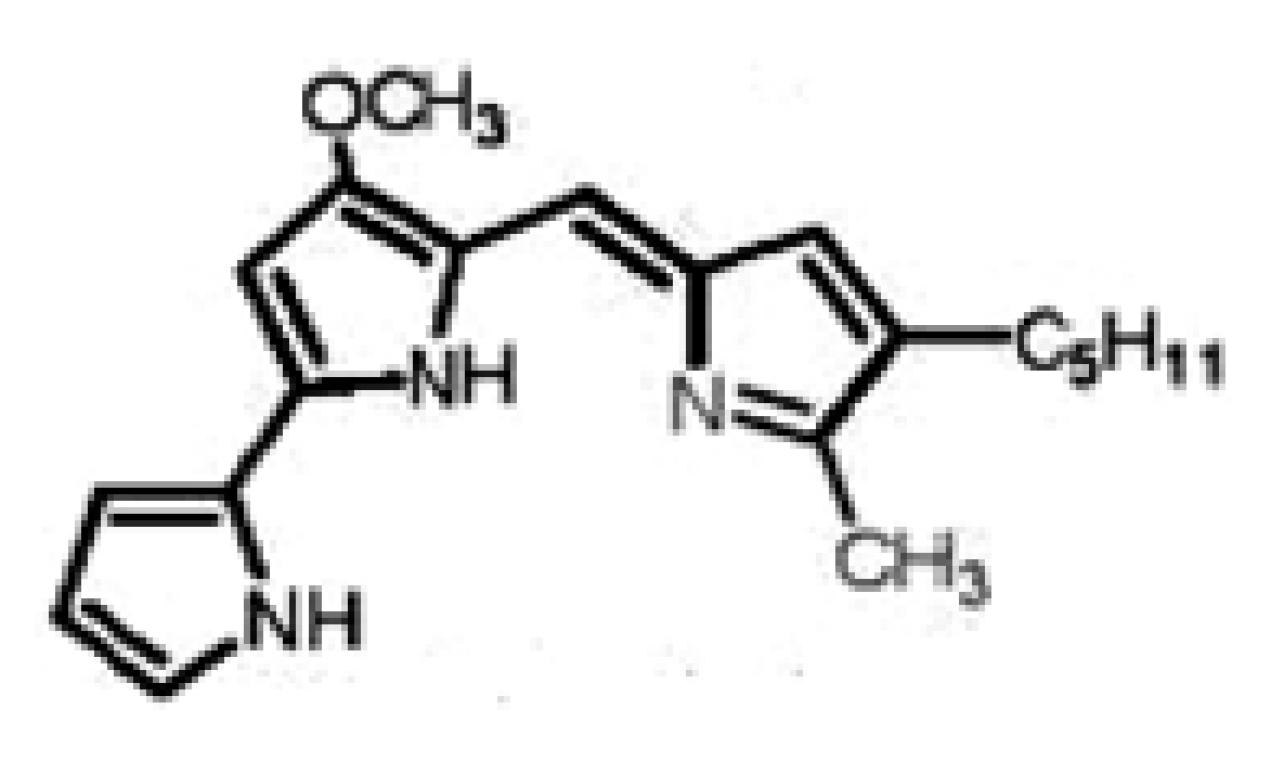
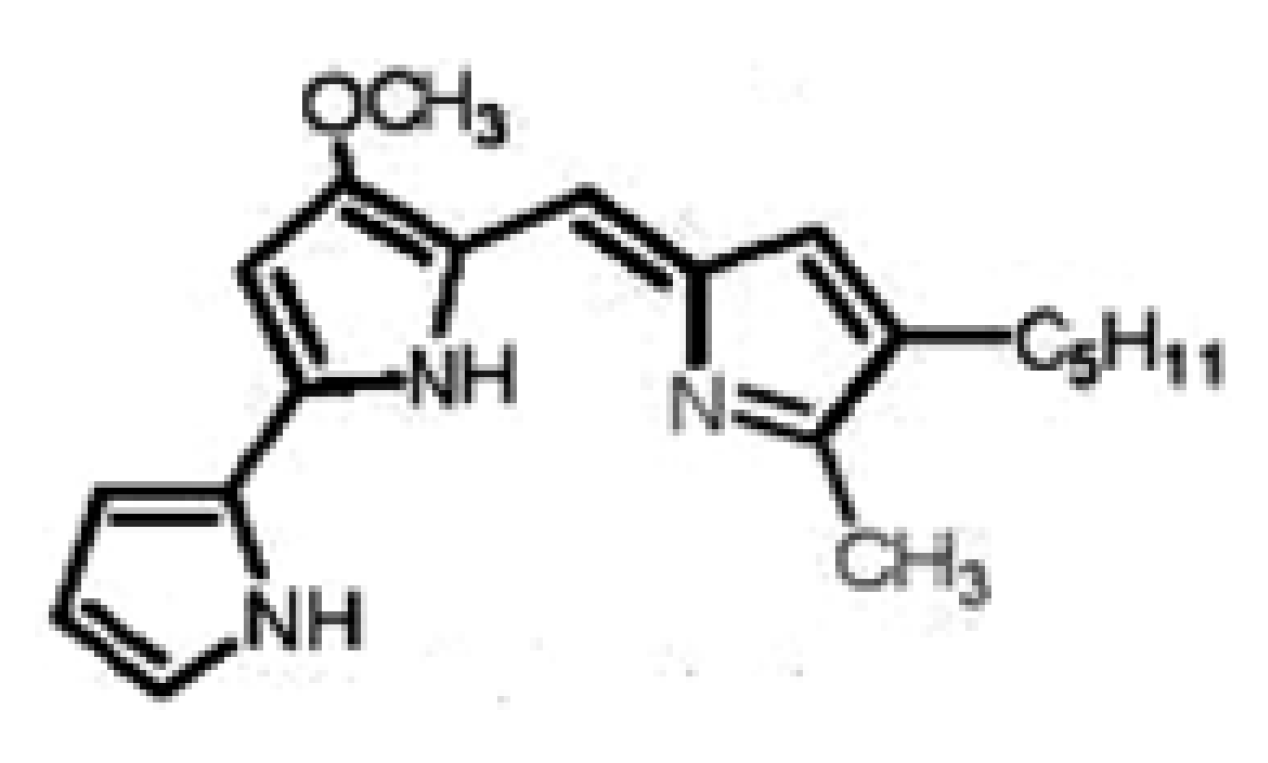
Results and Discussion
Effect of agroindustrial substrates on growth and pigment production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Colony color | Biomass (g/L-1) | Prodigiosin (mg/L) |
|---|---|---|---|
| Corn steep mannitol medium (CSMM) | Orange | 5.679 | 37500 |
| Mannitol medium (MM) | Dark red | 5.463 | 34000 |
| Corn steep medium (CSM) | Yellow | 3.465 | 25600 |
| Cassava waste mannitol medium (CWMM) | Red | 7.562 | 49500 |
| Cassava waste medium (CWM) | Light red | 3.654 | 27000 |
| Luria Bertani glucose medium (LBGM) | Rose | 2.865 | 13500 |

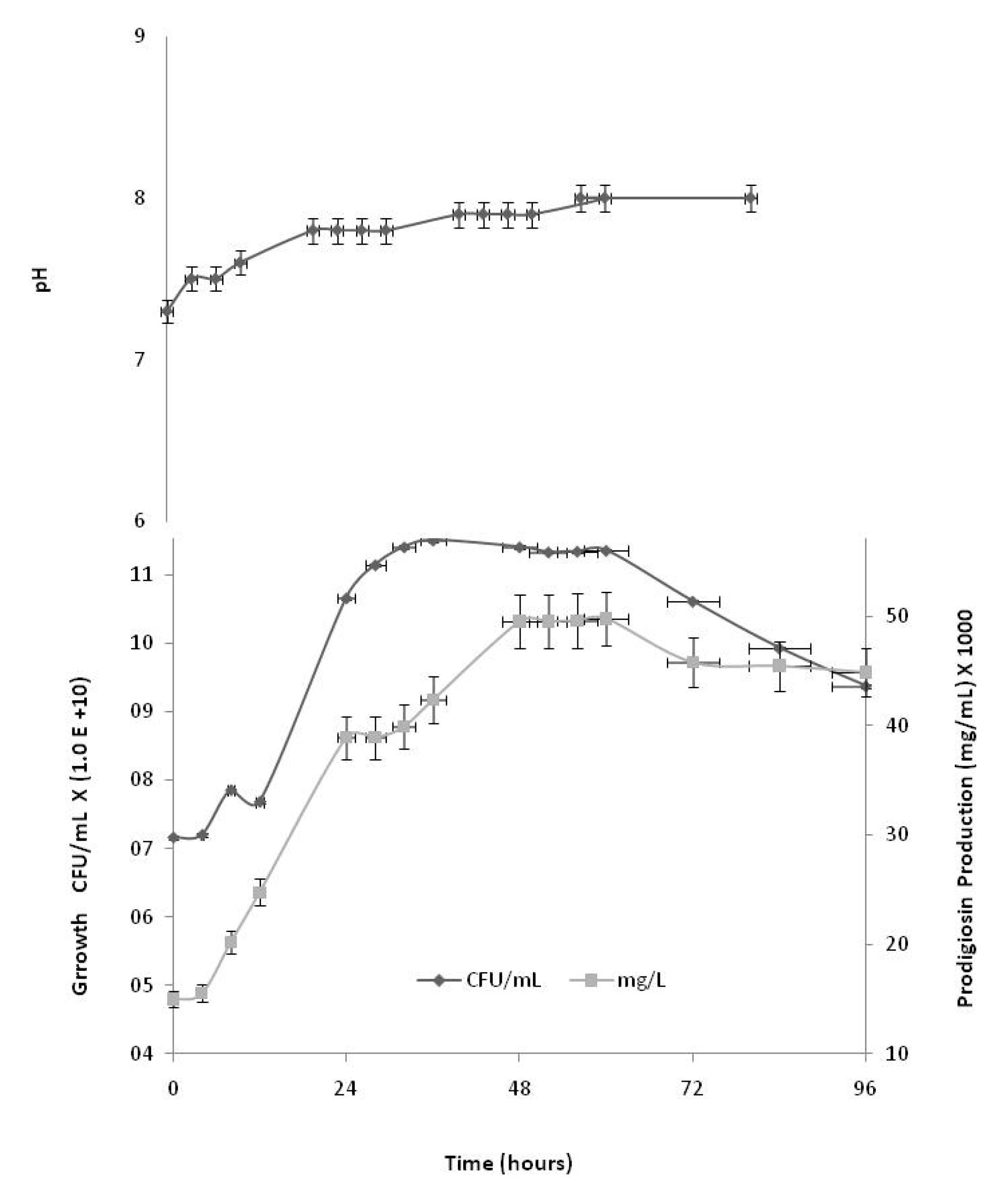
Kinetics of S. marcescens growth and prodigiosin production
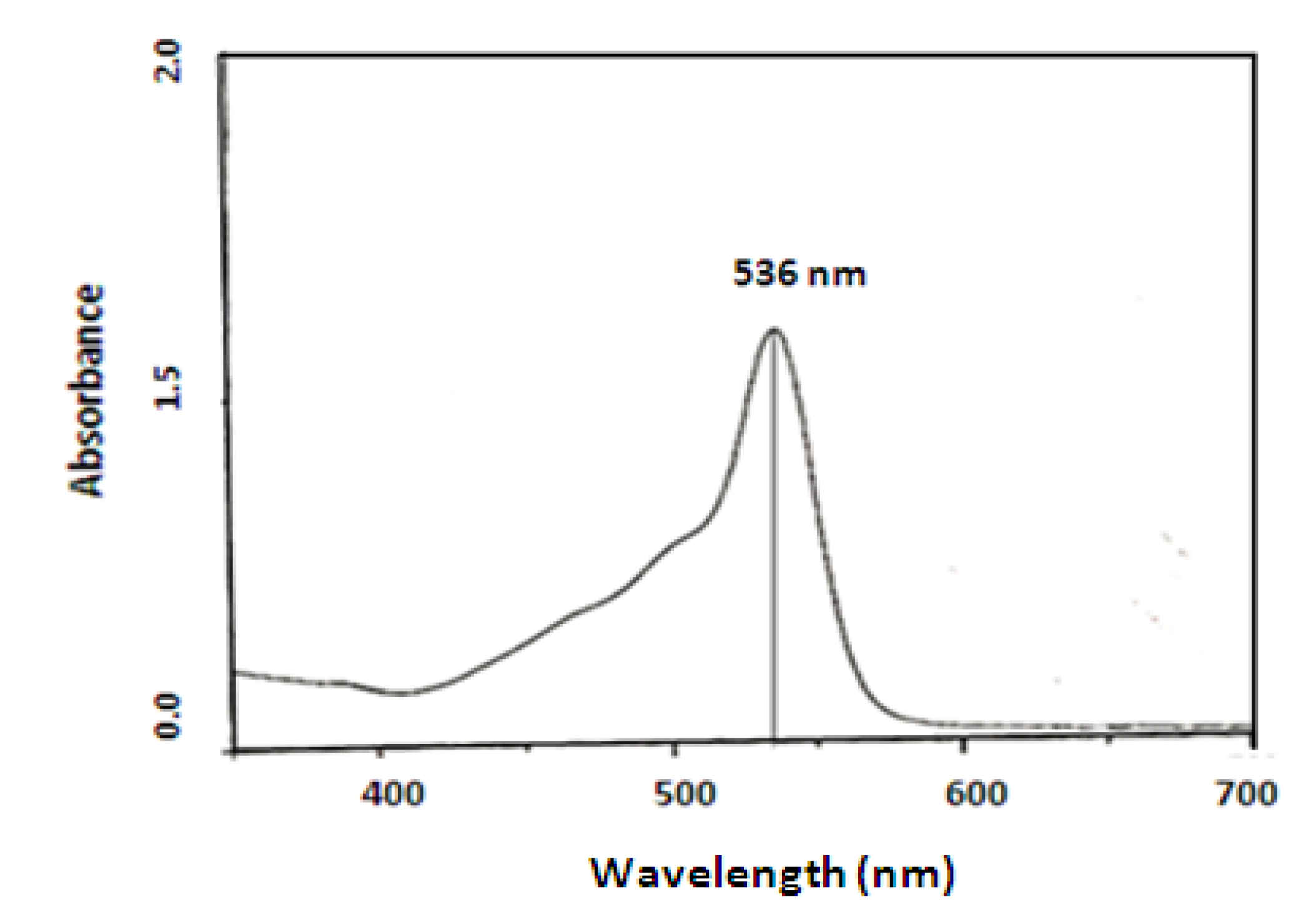
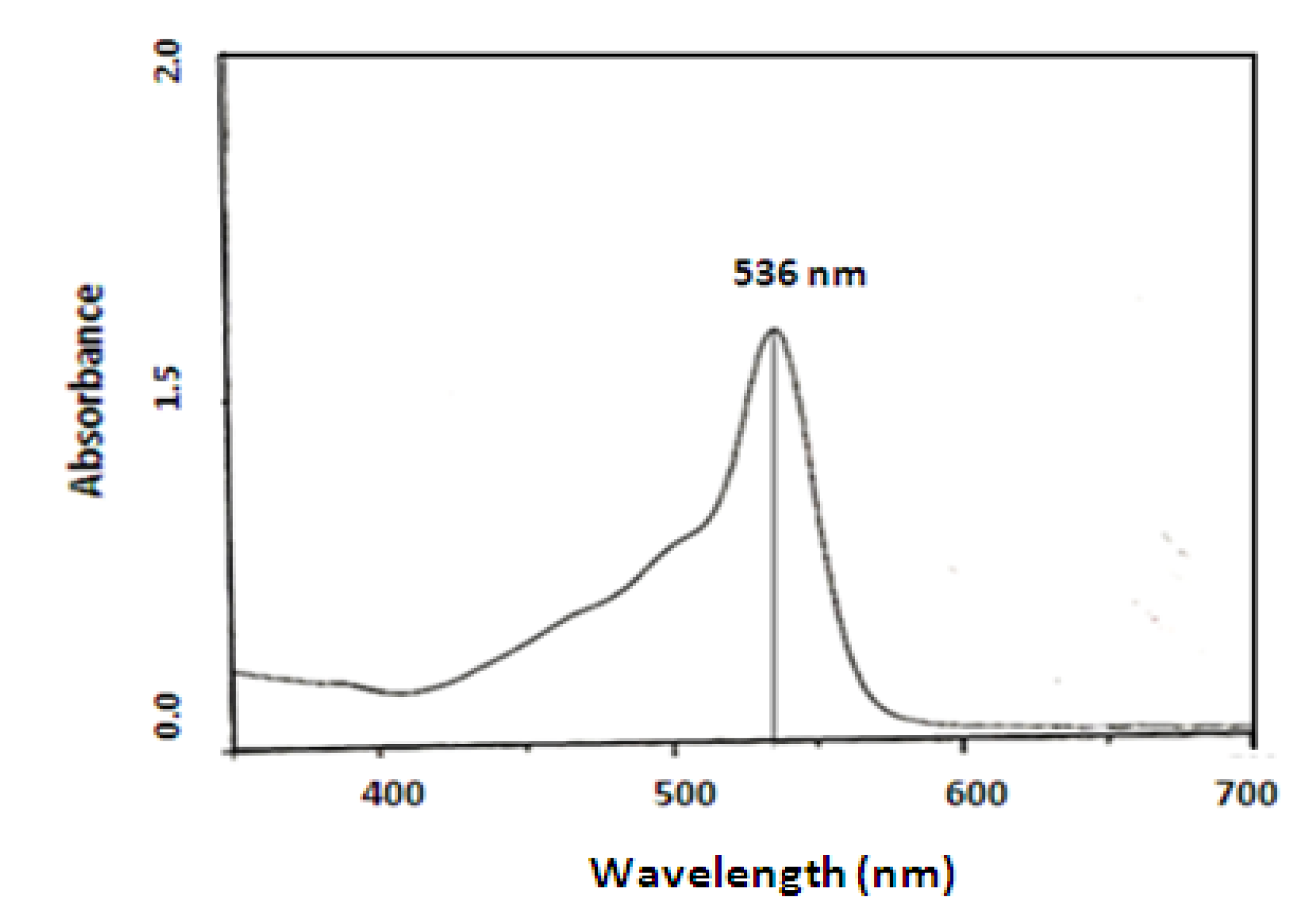
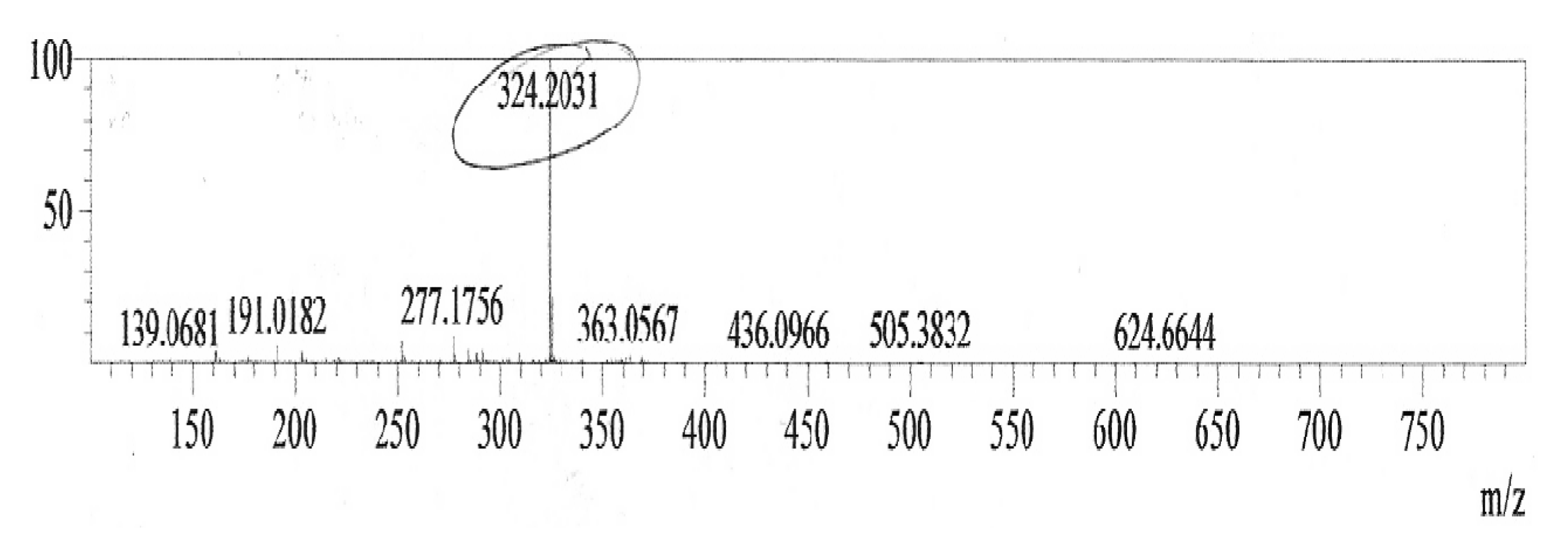
Prodigisiosin extraction and identification
Experimental
Microorganism and culture conditions
Pigment extraction and isolation
Kinetics of growth and prodigiosin production
Purification and chemical structure of the pigment
Conclusions
Acknowledgements
References
- Giri, A.V.; Anandkumar, N.; Muthukumaran, G.; Pennathur, G. A novel medium for the enhanced cell growth and production of prodigiosin from Serratia marcescens isolated from soil. BMC Microbiol. 2004, 4, 11–18. [Google Scholar] [CrossRef]
- Song, M.J.; Bae, J.; Lee, D.S.; Kim, C.H.; Kim, J.S.; Kim, S.W.; Hong, S.I. Purification and Characterization of Prodigiosin Produced by Integrated Bioreactor from Serratia sp. KH-95. J. Biosci. Bioeng. 2006, 101, 157–161. [Google Scholar] [CrossRef]
- Williamson, N.R.; Fineram, P.C.; Leeper, F.J.; Salmond, G.P.C. The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 2006, 4, 887–899. [Google Scholar] [CrossRef]
- Harris, A.K.P.; Williamson, N.R.; Slater, H.; Cox, A.; Abbasi, S.; Foulds, I.; Simonsen, H.T.; Leeper, F.J.; Salmond, G.P.C. The Serratia gene cluster encoding biosynthesis of the red antibiotic, prodigiosin, shows speciesand strain-dependent genome context variation. Microbiol. 2004, 150, 3547–3560. [Google Scholar] [CrossRef]
- Khanafari, A.; Assadi, M.M.; Fakhr, F.A. Review of Prodigiosin, Pigmentation in Serratia marcescens. Online J. Biol. Sci. 2006, 1, 1–13. [Google Scholar]
- Grimont, P.A.D.; Grimont, F.; Dulong, H.L.C.; De Rosnay; Sneath, P.H.A. Taxonomy of the genus Serratia. J. Gen. Microbiol. 1977, 98, 39. [Google Scholar] [CrossRef]
- Wai, Y.H.; Chen, W.C. Enhaced production of prodigiosin-like pigment from Serratia marcescens SMAR by medium improvement and oil-supplementation strategies. J. Biosci. Bioeng. 2005, 99, 616–622. [Google Scholar] [CrossRef]
- Pandey, R.; Chander, R.; Sainis, K.B. Prodigiosin as anti cancer agents: Living upto their name. Curr. Pharm. Des. 2009, 15, 732–741. [Google Scholar] [CrossRef]
- Kim, S.J.; Lee, H.K.; Yim, J.H. Statistical optimization of medium components for the production of prodigiosin by Hahella chejuensis KCTC 2396. J. Microbiol. Biotechnol. 2009, 18, 1903–1907. [Google Scholar]
- Moraes, C.S.; Seabra, S.H.; Albuquerque-Cunha, J.M.; Castro, D.P.; Genta, F.A.; De Souza, W.; Brazil, R.P.; Garcia, E.S.; Azambuja, P. Prodigiosin is not a determinant factor in lysis of Leishmania (Viannia) braziliensis after interaction with Serratia marcescens d-mannose sensitive fimbriae. Exp. Parasitol. 2009, 122, 84–90. [Google Scholar] [CrossRef]
- D’Alessio, R.; Bargiotti, A.; Carlini, O.; Colotta, F.; Ferrari, M.; Gnocchi, P.; Isetta, A.; Mongelli, N.; Motta, P.; Rossi, A.; Rossi, M.; Tibolla, M.; Vanotti, E. Synthesis and immunosuppressive activity of novel prodigiosin derivatives. J. Med. Chem. 2000, 43, 2557–2565. [Google Scholar] [CrossRef]
- Montaner, B.; Navarro, S.; Piqué, M.; Vilaseca, M.; Martinell, M.; Giralt Gil, J.; Pérez-Tomás, R. Prodigiosin from the supernatant of Serratia marcescens induces apoptosis in haematopoietic cancer cell lines. Br. J. Pharmacol. 2000, 131, 585–593. [Google Scholar] [CrossRef]
- Nakamura, K.; Kitamura, K. Process for preparation of prodigiosin. U.S. Patent 4,266,028, 5 May 1981. [Google Scholar]
- Carvalho, M.F.; Santos, A.; Egler, P.G. Avaliação da atividade poluidora da manipueira na bacia do rio Santa Rita, em Vitória da Conquista, Bahia. In XI Congresso Brasileiro de Mandioca, Campo Grande – Mato Grosso de Sul, 25-28 October 2005.
- Oboh, G.; Akindahunsi, A.A. Biochemical changes in Cassava products (flour and garri) subjected to Saccharomyces cerevisiae solid media fermentation. Appl. Trop. Agric. 2003, 82, 599–602. [Google Scholar]
- Oboh, G.; Akindahunsi, A.A. Chemical Changes in Cassava Peels Fermented with Mixed Culture of Aspergillus niger and Two species of Lactobacillus Integrated Bio-system. Appl. Trop. Agric. 2003, 2, 63–68. [Google Scholar]
- Barros, F.F.C.; Ponezi, A.N.; Pastore, G.M. Production of biosurfactant by Bacillus subtilis LB5a on a pilot scale using cassava wastewater as substrate. J. Ind. Microbiol. Biotechnol. 2008, 35, 1071–1078. [Google Scholar] [CrossRef]
- Andrade, R.F.S.; Luna, J.M.; Rufino, R.D.; Costa Albuquerque, C.D.; Sarubbo, L.A.; Campos Takaki, G.M. Surface Active Agent Produced by Candida lipolytica Using Cassava Flour Wastewater as Substrate. In Current Research Topics in Applied Microbiology and Microbial Biotchnology; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2009; pp. 751–756. [Google Scholar]
- Cang, S.; Sanada, M.; Johdo, O.; Ohta, S.; Nagamatsu, Y.; Yoshimoto, A. High production of prodigiosin by Serratia marcescens grown on ethanol. Biotchnol. Lett. 2000, 22, 1761–1765. [Google Scholar] [CrossRef]
- Harned, R.L. The production of Prodigiosin by submerged growth of Serratia marcescens. Appl. Environ. Microbiol. 1954, 2, 365–368. [Google Scholar]
- Araújo, H.W.C.; Ceballos, B.S.O.; Campos-Takaki, G.M. Biossurfactant production by Chromobacterium prodigiosum. In Current Research Topics in Applied Microbiology and Microbial Biotchnology; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2009; pp. 676–681. [Google Scholar]
- Venil, C.K.; Lakshmanaperumalsamy, P. Application of statistical design to the optimization of culture medium for prodigiosin production by Serratia marcescens SWML08. Malay. J. Microbiol. 2009, 5, 55–61. [Google Scholar]
- Chang, S.S.; Liu, C.H.; Chen, H.C. Cell arrangement and prodigiosin production of Serratia marcescens R-1. J. Fishery Soc. 1993, 20, 37–43. [Google Scholar]
- Melo, P.S.; Dura´, N.N.; Haun, M. Cytotoxicity of prodigiosin and benznidazole on V79 cells. Toxicol. Lett. 2000, 116, 237–242. [Google Scholar] [CrossRef]
- Nakashima, T.; Kurachi, M.; Kato, Y.; Yamaguchi, K.; Oda, T. Characterization of bacterium isolated from the sediment Coast area of Omura Bay in Japan and several biological activies of pigment produced by this isolated. Microbiol Immunol. 2005, 49, 407–415. [Google Scholar]
- Pirt, S.J. Principles of Microbe and cell cultivation, 1st ed.; Blackwell Scientific Publications: London, UK, 1975; pp. 115–117. [Google Scholar]
- Sample Availability: Contact the authors.
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Casullo de Araújo, H.W.; Fukushima, K.; Takaki, G.M.C. Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate. Molecules 2010, 15, 6931-6940. https://doi.org/10.3390/molecules15106931
Casullo de Araújo HW, Fukushima K, Takaki GMC. Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate. Molecules. 2010; 15(10):6931-6940. https://doi.org/10.3390/molecules15106931
Chicago/Turabian StyleCasullo de Araújo, Helvia W., K. Fukushima, and Galba M. Campos Takaki. 2010. "Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate" Molecules 15, no. 10: 6931-6940. https://doi.org/10.3390/molecules15106931
APA StyleCasullo de Araújo, H. W., Fukushima, K., & Takaki, G. M. C. (2010). Prodigiosin Production by Serratia marcescens UCP 1549 Using Renewable-Resources as a Low Cost Substrate. Molecules, 15(10), 6931-6940. https://doi.org/10.3390/molecules15106931