Abstract
Accumulating evidence indicates that consumption of tea, especially green tea, is good for preventing cancer. To elucidate the cancer preventive mechanisms of green tea, much effort has been devoted to investigating the anticancer effects of (-)-epigallocatechin-3-gallate (EGCG), the major component of green tea. It has been revealed that EGCG restrained carcinogenesis in a variety of tissues through inhibition of mitogen-activated protein kinases (MAPK), growth factor-related cell signaling, activation of activator protein 1 (AP-1) and nuclear factor-B (NF-κB), topoisomerase I, matrix metalloproteinases and other potential targets. Therefore, EGCG is a multipotent anticancer agent, which not only provides solid evidence to support the anticancer potential of green tea, but also offers new clues for discovering multiple-targeted anticancer drugs.
1. Introduction
Tea is one of the most popular beverages consumed in the world. Generally, it is divided into three types: green tea (non-fermented), oolong tea (semi-fermented), and black tea (fermented). Consumption of tea, especially green tea, has been associated with many healthy benefits including cancer prevention [1]. Some epidemiological observations have revealed that there was an inverse correlation between increased green tea intake and relative risk for cancers. A prospective cohort study with 8,552 subjects from Saitama Prefecture in Japan indicated that green tea had a potentially preventive effect against cancers in many organs including stomach, lung, colorectum and liver [2]. Green tea drinking was also found to protect against stomach cancers in residents of Nagoya in Japan [3], pancreatic and colorectal cancers in Shanghai residents [4] and breast cancers in Saitama, Japan [5].
The benefits of green tea with regards to cancer prevention have been attributed, in a large part, to the green tea polyphenols, especially catechins. A cohort study (Iowa Women’s Health Study) examined 34,651 postmenopausal women for 12 years and found that food-derived catechin intake was inversely associated with a rectal cancer incidence [6]. The green tea catechins mainly consist of (-)-epicatechin (EC), (-)-epigallocatechin (EGC), (-)-epicatechin gallate (ECG) and (-)-epigallocatechin-3-gallate (EGCG) (Figure 1), of which EGCG may be the most effective chemopreventive agent and has been extensively studied with different human cancer cell lines and several cancer animal models. It has been revealed that EGCG inhibited carcinogenesis in a variety of tissues including lung, bladder, skin, small intestine, prostate and breast [7,8,9,10,11]. As to the molecular mechanisms, EGCG has the potential to inhibit the multiple targets implicated in the initiation, promotion and progression stage of cancers. Therefore, EGCG is a multiple-targeted anticancer agent with diverse activities, which stimulated our interest to summarize the current knowledge on this topic in this review.
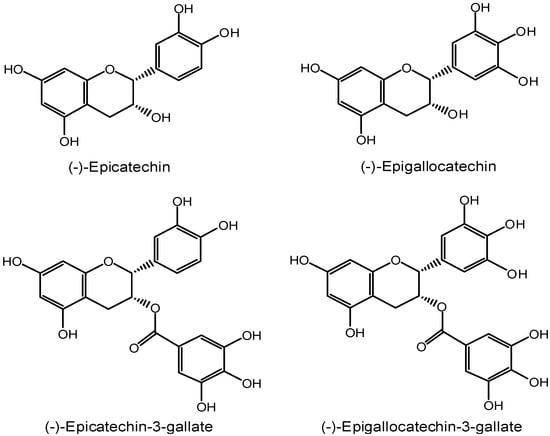
Figure 1.
Main catechin components of green tea polyphenols.
Figure 1.
Main catechin components of green tea polyphenols.
2. Inhibition of the cancer initiation stage by EGCG
It is well known that oncogene mutation and reactive oxygen species (ROS) play important roles in the cancer initiation stage. Oncogene mutation leads to procarcinogen activation by activating some phase I enzymes such as the cytochrome P450s. ROS actively participate in the metabolic activation of procarcinogens. EGCG can neutralize these procarcinogens by inhibiting the activity of cytochrome P450 enzymes and modulating ROS.
Wang et al. [12] investigated the interaction of EGCG with rat hepatic microsomal P450 and found that EGCG significantly inhibited NADPH-cytochrome c reductase activity. An examination of the structure-activity relationships of epicatechin derivatives suggested that the inhibitory effect on the microsomal enzyme system might arise from the galloyl or hydroxyl groups in the molecule. Mukhtar et al. [13] also claimed that EGCG could interact with hepatic cytochrome P450 and inhibit the P450-dependent mixed-function oxidase enzymes in skin and liver.
Considerable evidence has demonstrated that EGCG is a powerful antioxidant. The ROS-scavenging effects of EGCG were superior to those of ascorbic acid and α-tocopherol in many cases [14]. Besides, the pyrogallol structure of EGCG also confers the molecule with strong metal-chelating ability. As a result, EGCG can bind with transition metal ions and behave as a preventive antioxidant [15,16]. Its high affinity towards the lipid bilayers also facilitates the entry of EGCG into the nuclei of cancer cells [17].
3. Inhibition of the cancer promotion stage by EGCG
The cancer promotion stage is a reversible and a long-term process, in which some intracellular signaling pathways and proteins associated with cell cycle are involved. EGCG exerts its anticancer effect by interfering with many signaling pathways and modulating cell cycle.
3a. Interference with intracellular signaling pathway (Figure 2).
Mitogen-activated protein kinases (MAPKs) pathways are very common in various cells. They are composed of a group of Ser/Thr protein kinases that are activated as a cascade. Three classes of MAPKs are most well known, namely extracellular signal-regulated protein kinases (ERKs), c-Jun N-terminal kinases/stress activated protein kinases (JNKs/SAPKs) and p38 kinases. In general, ERKs are critical transducers of proliferation signals and are often activated by growth-inducing tumor promoters, including 12-O-tetradecanoyl-phorbol-13-acetate (TPA), epidermal growth factor (EGF) and platelet-derived growth factor (PDGF). In comparison, JNKs/SAPKs and p38 kinases are strongly activated by stress-related tumor promoters, such as ultraviolet (UV) irradiation and arsenic. Some studies have shown that EGCG can regulate the important molecules in MAPK pathway, which result in the inhibition of cancer cell survival.
Katiyar et al. [18] reported that treatment of H2O2 resulted in phosphorylation of ERK1/2, JNK, and p38 in human epidermal keratinocytes. When these cells were pretreated with EGCG, H2O2-induced phosphorylation of ERK1/2, JNK, and p38 were found to be significantly inhibited. Maeda-Yamamoto et al [19] also reported that EGCG inhibited the phosphorylation of extracellular signal-regulated kinases 1 and 2 (ERK1/2), and suppressed p38 MAPK activity in human fibrosarcoma HT1080 cells. These findings demonstrated that EGCG had potential of inhibiting oxidative stress-mediated phosphorylation of MAPK signaling pathways.
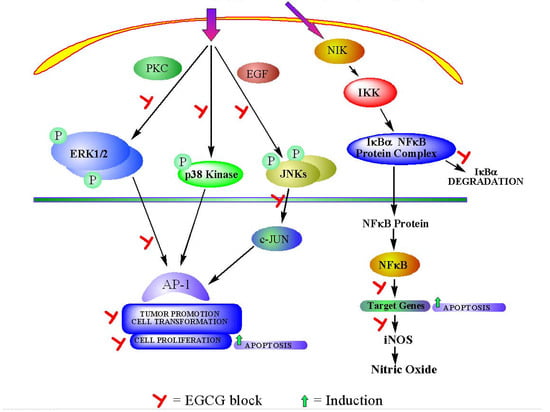
Figure 2.
Intracellular signaling pathways and its modulation by EGCG.
Figure 2.
Intracellular signaling pathways and its modulation by EGCG.
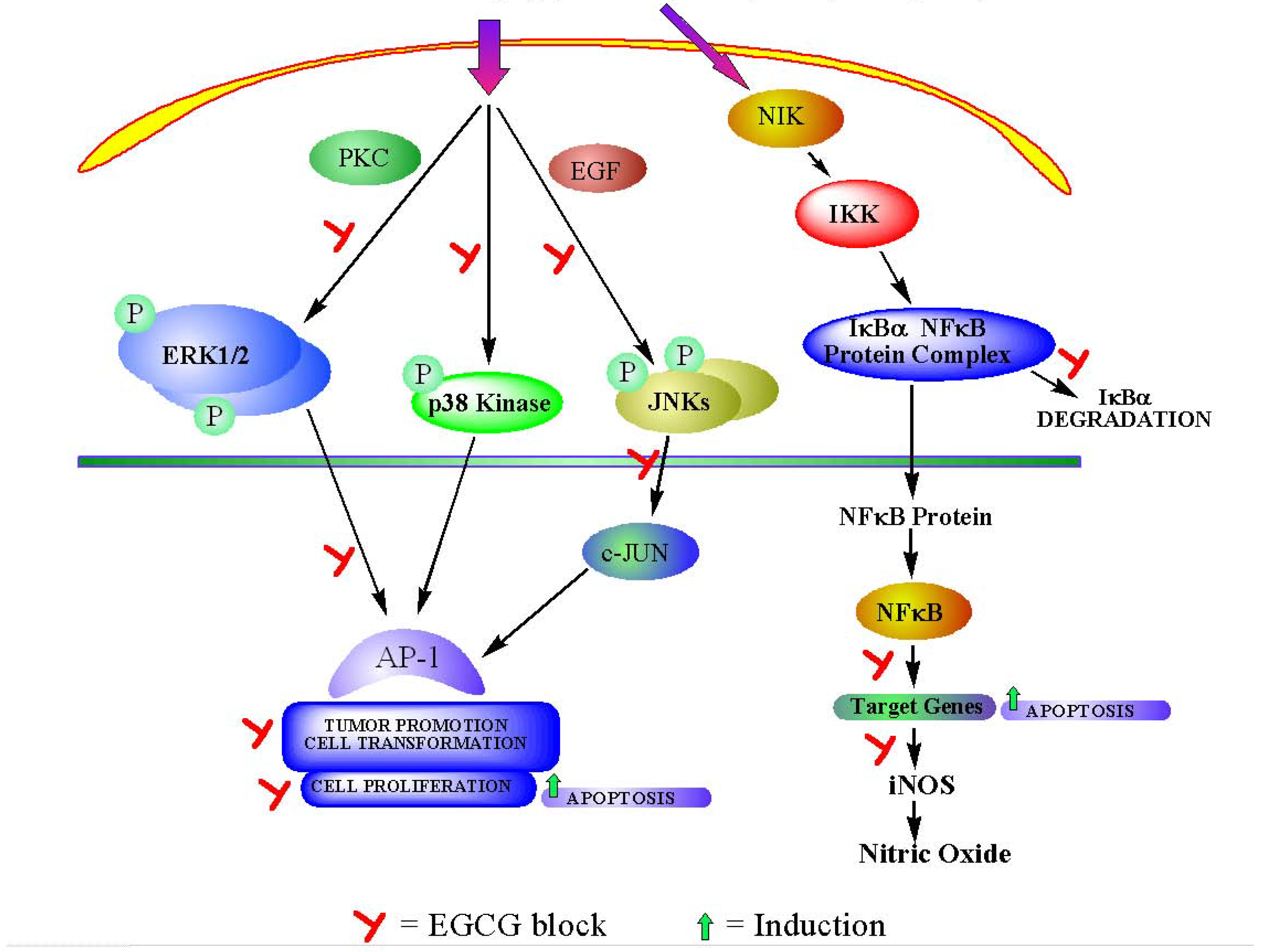
After activation, MAPKs translocate to the nucleus to activate numerous transcription factors. Two major transcription factors in eukaryotic cells are nuclear factor-kappa B (NF-κB) and activator protein-1 (AP-1) [20,21]. NF-κB is a pleiotropic transcription factor that exists in all cell types and plays an important role in regulating the expression of multiple genes. NF-κB signaling pathway is composed of about a dozen different dimers comprising five structurally related DNA-binding proteins, namely NF-κB1/p50, NF-κB2/p52, RelA/p65, Rel/c-Rel and RelB. It has been well accepted that NF-κB signaling pathway plays a critical role in the control of cell growth and apoptosis [22]. Although there are other important molecules, such as IκB and IKK, within NF-κB signaling pathway, NF-κB is the key factor in this pathway.
Based on the many functions of NF-κB target genes, a close relationship between NF-κB and cancer has been proposed. Some studies have confirmed this hypothesis. Treatment with EGCG (10-40 µmol/L) in a dose- and time-dependent manner was found to inhibit UVB-mediated activation of NF-κB in normal human epidermal keratinocytes [23]. Gupta et al. [24] have identified NF-κB /p65 component of the NF-κB complex as a target for specific cleavage by caspases during EGCG-mediated apoptosis.
Cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) are important enzymes that mediate inflammatory processes. Improper up-regulation of COX-2 and/or iNOS has been associated with pathophysiology of certain types of human cancers. Surh et al. [25] have shown that EGCG could down-regulate the expression of COX-2 and iNOS by suppressing NF-κB activity. These data strongly suggested that NF-κB and its components are potential targets for EGCG cancer prevention. Since NF-κB is considered as a target for the management of cancer, based on recent studies, modulation of this pathway by EGCG could contribute to its chemopreventive potential.
AP-1 is another important eukaryotic transcription factor. It is a well-characterized transcription factor composed of members of the basic region leucine zipper protein superfamily, i.e., Jun, Fos and activating transcription factor proteins [26]. It regulates the transcription of various genes associated with cellular proliferation and apoptosis [27]. Strong evidence indicates that AP-1 plays a key role in cancer development and it is up-regulated during tumor promotion stage.
Dong et al. [28] found that EGCG could inhibit EGF- or TPA-induced cell transformation, as well as AP-1-induced transcriptional activity and DNA binding activity. This study also implicated that the inhibition of AP-1 activation occurred via the inhibition of a JNK-dependent pathway. Huang et al. [29] showed that EGCG inhibited arsenite-induced AP-1 transcriptional activation and AP-1 DNA binding activity. Thus, AP-1 serves as another potential target, besides NF-κB, for the cancer prevention effects of EGCG.
In addition, EGCG can also influence epidermal growth factor receptor–mediated signal transduction pathway. Over-expression of growth factor and growth factor receptors such as EGF-Receptor (EGFR), PDGF-Receptor (PDGFR) and others can result in a neoplastic phenotype in tumor cells. EGCG can interfere with cancer promotion by inhibiting growth factor-mediated cell signaling pathway.
Shimizu et al. [30] have reported that EGCG (at 10-20 µg/mL) was shown to inhibit the activation of the EGFR, HER2, and multiple downstream signaling pathways in colon cancer cell lines. Tachibana et al. [31] found that EGCG bound to a specific metastasis associated 67kDa laminin receptor that was expressed on a variety of tumor cells. It was shown using a subtraction cloning strategy involving cDNA libraries constructed from cells treated or untreated with all trans-retinoic acid that the anticancer action of EGCG is mediated by laminin receptor and it allows EGCG to bind to the cell surface. Based on these data, it was suggested that there exist a receptor for EGCG. This suggestion awaits follow-up and confirmation.
In addition, Jung et al. [32] reported that 30 µmol/L of EGCG inhibited serum starvation-induced vascular endothelial growth factor (VEGF) expression in HT29 colon cancer cells. This observation may account for the antitumor activity of EGCG against HT29 xenografts in athymic nude mice. The authors observed that EGCG not only decreased tumor growth (58%) and increased tumor cell apoptosis (1.9-fold) but also decreased tumor microvessel density (30%).
3b. Cell cycle modulation
Characterization of the cell cycle has made rapid progress in recent years. The relationship between disrupted cell cycle control and cancer development has been a focus in carcinogenetic mechanism studies. Some research has demonstrated that the anticancer functions of EGCG might be associated with cell cycle modulation.
As an important family of positive cell cycle regulators, cyclins are frequently over-expressed in cancer cells, whereas the negative cell cycle regulators or Cyclin-Dependent Kinase Inhibitors (CKI), such as p16, p21, p27, p53 and P73, are under-expressed. Some investigators found that EGCG could prevent the cancer progression by up-regulating CKI, and inhibiting the activities of Cyclin-Dependent Kinase (CDK) such as CDK1 and CDK2.
Nihal et al. [33] found that EGCG treatment of human melanoma cells resulted in a significant, dose-dependent decrease in cyclin D1 and CDK2 protein levels and induction of p16, p21 and p27. Kavanagh et al. [34] found that p27, which could promote G1/S phase growth arrest, was induced in breast cancer cells by treatment with EGCG. Hastak et al. [35] have clearly demonstrated that EGCG activated growth arrest, primarily via a p53-dependent pathway that involved the function of both p21 and Bax such that down-regulation of either molecule conferred a growth advantage to prostate carcinoma cells. These results indicated that EGCG might exert its growth-inhibitory effects through modulation of cell cycle regulatory proteins.
Berger et al. [36] have demonstrated that EGCG selectively inhibited the activity of topoisomerase I (but not topoisomerase II), which plays a role in DNA replication, transcription, and chromosome condensation in human colon cancer cell lines. The doses of EGCG necessary for this inhibition (10-17 µmol/L) were lower than those necessary for inhibition of cell growth (IC50 = 10-90 µmol/L).
Telomerase is an enzyme essential for unlocking the proliferative capacity of cancer cells and it is lacking in normal somatic cells. It plays a salient role in the process of cancer. In most cancers, the maintenance of telomeres is achieved through the expression of telomerase, which stabilizes and elongates telomeres by the de novo synthesis of telomeric DNA. The role of telomerase in immortalization was confirmed recently by the findings that the ectopic expression of telomerase in various normal cells resulted in the extension of the life span of these cells [37,38]. Naasani et al. [39] demonstrated that EGCG after structural rearrangements at physiologically permissible conditions increased remarkably telomerase inhibition. In nude mice models bearing both telomerase-dependent- and independent xenograft tumors cloned from a single human cancer progeny, only the telomerase-dependent tumors responded to prolonged oral administration of EGCG. It is also reported that EGCG strongly inhibited telomerase activity, and thus induced senescence, limited the life span of cancer cells, in both leukemias and solid tumors [40]. Therefore, telomerase inhibition could be one of the mechanisms underlying the anticancer effects of EGCG.
4. Inhibition of the cancer progression stage by EGCG
During the complicated processes of cancer progression, apoptosis and some enzymes such as urokinase and matrix metalloproteinases (MMPs) play a key role. Accumulating evidence indicates that EGCG can inhibit the growth of malignant cells by inducing apoptosis and inhibiting the activity of some such enzymes.
Apoptosis is an active form of cell suicide controlled by a network of genes, in which the Bcl-2 family of proteins plays important roles in control of apoptosis via regulating mitochondrial permeability and releasing of cytochrome c, which activates the caspase cascade. Apoptosis is an essential process which plays a critical role in the pathogenesis of diseases including cancer. EGCG has been shown to induce the apoptosis in a number of cancer cells.
Hwang et al. [41] demonstrated that a treatment of chemoresistant HT-29 human colon cancer cells with 100 μmol/L EGCG inhibited cell proliferation by inducing apoptosis. Nishikawa et al. [42] showed that EGCG inhibited the growth of HLE cells (an undifferentiated human hepatocellular carcinoma cell line) in vitro and in vivo. The inhibition was caused by the induction of apoptosis as a result of the activations of caspase-8, -9 and -3. These caspases appeared to be activated by the down-regulation of Bcl-2α and Bcl-xl. Thangapazham et al. [10] found that EGCG treatment inhibited proliferation and induced apoptosis of MDA-MB-231 cells in vitro and in vivo. Lin et al. [43] clearly identified the inhibitory effect of EGCG on differentiation and the induction of apoptosis in adipocytes in vitro. Islam et al. [44] showed that EGCG displayed strong inhibitory effects on the proliferation and viability of HTB-94 human chondrosarcoma cells by inducing apoptosis.
Qin et al. [45] found that EGCG treatment induced apoptosis in the T24 human bladder cancer cell line by inhibiting phosphatidylinositol 3'-kinase/Akt activation that, in turn, resulted in modulation of Bcl-2 family proteins, leading to enhanced apoptosis of T24 cells. Recent studies find that CKI-p53, p73 play an important role for the anticancer function of EGCG.
Amin et al. [46] have identified SHP-2 (a kind of tyrosine phosphatase) as a protective factor of cells lacking functional p53 from EGCG-induced apoptosis. Moreover, they revealed a number of targets for EGCG-induced apoptosis which were expressed in the absence of functional p53. This had not previously been reported to be so induced. At the same time, they also identified a crucial role for p73 in EGCG-induced apoptosis and a number of previously unidentified p73 target genes. Manna et al. [47] suggested that in vivo EGCG could induce apoptosis in Sarcoma180 (S180) cells through alteration in G2/M phase of the cell cycle by up-regulation of p53, bax and down-regulation of c-myc, bcl-2 and U1B, U4-U6 UsnRNAs (uridylic acid rich small nuclear RNAs).
In addition, some researchers found that the pro-apoptotic activity of EGCG appeared to depend on the cell type and the dosage treated. Up-regulation of p57 was shown in both normal and oral cancer cells, but apoptosis was induced only in malignant cells, suggesting that EGCG affected a p57-mediated survival pathway in normal epithelial cells, while induced a pro-apoptotic pathway in oral cancer cells [48,49].
Cancer invasion and metastasis are multifactorial processes and require the coordinated action of cell-secreted proteolytic enzymes and their inhibitors [50]. EGCG can also prevent the cancer progression stage by influencing urokinase and matrix metalloproteinases.
Urokinase is a kind of hydrolase. The urokinase plasminogen activator (uPA) is commonly overexpressed in many different human cancers [51]. Inhibition of urokinase-type plasminogen activator (uPA) activity can reduce tumor size or even cause complete remission of tumors in mice [52]. Jankun et al. [53] used computer-aided molecular modeling to demonstrate that EGGC bound to urokinase, blocked the histidine 57 and serine 195 residues of the urokinase catalytic triad and extended towards arginine 35 from a positively charged loop of urokinase. Binding of EGCG at such a location would interfere with the ability of uPA to recognize its substrates, thereby inhibiting its enzymatic activity. Kim et al. [54] indicated that the inhibition of uPA expression by EGCG was important for the anti-invasive function of EGCG.
MMPs are a family of tightly regulated zinc-dependent proteases that can degrade nearly all components of the extracellular matrix. A large body of experimental evidence suggested that MMPs essentially contributed to the maintenance of tumor growth in primary and metastatic sites [55,56,57]. Adhami et al. [58] have shown that p.o. administered green tea polyphenols (0.1% in drinking water) caused marked inhibition of MMP-2 and MMP-9 in the prostate in TRAMP mice. Fassina et al. [59] found that EGCG (25-100 µmol/L) could inhibit the MMP-2 and MMP-9 in endothelial cells. Annabi et al. [60] have shown that EGCG likewise inhibited the activity and expression of membrane-type matrix metalloproteinase 1-MMP (MT1-MMP), a protein responsible for the activation of MMP. Thus, it seems that EGCG could inhibit or delay cancer invasion, metastasis, and angiogenesis via modulations in MMPs.
Recently, by employing surface plasmon resonance assay (Biacore) and cold spray ionization-mass spectrometry, Kuzuhara et al. [61] claimed that DNA and RNA served as new binding targets of EGCG. Their results suggested that multiple binding sites of EGCG were present in DNA and RNA oligomers and revealed for the first time the link between catechins and polynucleotides. These findings will promote our understanding of the effects of catechins on DNA in terms of cancer prevention.
5. Conclusions
The above description illuminates the cancer preventive mechanisms of EGCG. Since the development of cancer is a dynamic and multistage process, in which a large number of genes and proteins are involved, to combat the disease, we will have to shift the drug-discovery paradigm from “one-drug, one-target” to “one-drug, multiple-targets”. Hence, the potential of EGCG in hitting multiple targets implicated in various stages of cancer development not just provides solid evidence to support the anticancer effects of green tea, but also offers new evidence to justify the multiple-targeted antitumor strategy. In addition, as the structure of EGCG holds the structural secrets of multipotent anticancer agents, EGCG is expected to serve as a promising starting point to derive novel anticancer drugs.
Acknowledgments
This study was supported by the National Basic Research Program of China (grant 2003CB114400) and the National Natural Science Foundation of China (grant 30100035 and 30570383).
References and Notes
- Fujiki, H.; Suganuma, M.; Imai, K.; Nakachi, K. Green tea: Cancer preventive beverage and/or drug. Cancer Lett. 2002, 188, 9–13. [Google Scholar] [CrossRef]
- Imai, K.; Suga, K.; Nakachi, K. Cancer-preventive effects of drinking green tea among a Japanese population. Prev. Med. 1997, 26, 769–775. [Google Scholar] [CrossRef]
- Inoue, M.; Tajima, K.; Hirose, K.; Hamajima, N.; Takezake, T.; Kuroishi, T.; Tominaga, S. Tea and coffee consumption and risk of digestive tract cancers: data from a comparative case-referent study in Japan. Cancer Causes Control 1998, 9, 209–216. [Google Scholar]
- Ji, B. T.; Chow, W. H.; Hsing, A. W.; McLaughlin, J. K.; Dai, Q.; Gao, Y. T.; Blot, W. J.; Fraumeni, J. F. Green tea consumption and the risk of pancreatic and colorectal cancers. Int. J. Cancer 1999, 70, 255–258. [Google Scholar]
- Nakachi, K.; Suemasu, K.; Suga, T.; Takeo, K.; Imai, K.; Higashi, Y. Influence of drinking green tea on breast cancer malignancy among Japanese patients. Jpn. J. Cancer Res. 1998, 89, 254–261. [Google Scholar] [CrossRef]
- Arts, I. C.; Jacobs, D. R.; Gross, M.; Harnack, L. J.; Folsom, A. R. Dietary catechins and cancer incidence among postmenopausal women: the Iowa Women’s Health Study (United States). Cancer Cause Control 2002, 13, 373–382. [Google Scholar] [CrossRef]
- Mimoto, J.; Kiura, K.; Matsuo, K.; Yoshino, T.; Takata, I.; Ueoka, H.; Kataoka, M.; Harada, M. (-) -Epigallocatechin gallate can prevent cisplatin-induced lung tumorigenesis in A/J mice. Carcinogenesis 2000, 21, 915–919. [Google Scholar] [CrossRef]
- Chen, J. J.; Ye, Z. Q.; Koo, M. W. Growth inhibition and cell cycle arrest effects of epigallocatechin gallate in the NBT-II bladder tumour cell line. BJU Int. 2004, 93, 1082–1086. [Google Scholar] [CrossRef]
- Mantena, S. K.; Roy, A. M.; Katiyar, S. K. Epigallocatechin-3-gallate inhibits photocarcinogenesis through inhibition of angiogenic factors and activation of CD8+ T cells in tumors. Photochem. Photobiol. 2005, 81, 1174–1179. [Google Scholar] [CrossRef]
- Stuart, E. C.; Scandlyn, M. J.; Rosengren, R. J. Role of epigallocatechin gallate (EGCG) in the treatment of breast and prostate cancer. Life Sci. 2006, 79, 2329–2336. [Google Scholar]
- Thangapazham, R. L.; Singh, A. K.; Sharma, A.; Warren, J.; Gaddipati, J. P.; Maheshwari, R. K. Green tea polyphenols and its constituent epigallocatechin gallate inhibits proliferation of human breast cancer cells in vitro and in vivo. Cancer Lett. 2007, 245, 232–241. [Google Scholar] [CrossRef]
- Wang, Z. Y.; Das, M.; Bickers, D. R.; Mukhtar, H. Interaction of Epicatechins Derived from Green Tea with Hepatic CytochromP-450. Drug Metab. Dispos. 1988, 16, 98–103. [Google Scholar]
- Mukhtar, H.; Wang, Z. Y.; Katiqan, S. K.; Agarwal, R. Tea components: antimutagenic and antigagenic effects. Prev. Med. 1992, 21, 351–360. [Google Scholar] [CrossRef]
- Zhao, B. L.; Li, X. J.; He, R. G.; Cheng, S. J.; Xin, W. J. Scavenging effect of extracts of green tea and natural antioxidants on active oxygen radicals. Cell Biophys. 1989, 14, 175–185. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, B.; Li, M.; Shen, S.; Xin, W. Studies on protective mechanisms of four components of green tea polyphenols against lipid peroxidation in synaptosomes. Biochim. Biophys. Acta. 1996, 1304, 210–222. [Google Scholar] [CrossRef]
- Zhang, G.; Miura, Y.; Yagasaki, K. Suppression of adhesion and invasion of hepatoma cells in culture by tea compounds through antioxidative activity. Cancer Lett. 2000, 159, 169–173. [Google Scholar] [CrossRef]
- Okabe, S.; Suganuma, M.; Hayashi, M.; Sueoka, E.; Komori, A.; Fujiki, H. Mechanisms of growth inhibition of human lung cancer cell line, PC-9, by tea polyphenols. Jpn. J. Cancer Res. 1997, 88, 639–643. [Google Scholar] [CrossRef]
- Katiyar, S. K.; Afaq, F.; Azizuddin, K.; Mukhtar, H. Inhibition of UVB-induced oxidative stress-mediated phosphorylation of mitogen-activated protein kinase signaling pathways in cultured human epidermal keratinocytes by green tea polyphenol (-)-epigallocatechin-3-gallate. Toxicol. Appl. Pharmacol. 2001, 176, 110–117. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Suzuki, N.; Sawai, Y.; Miyase, T.; Sano, M.; Hashimoto-Ohta, A.; Isemura, M. Association of suppression of extracellular signal-regulated kinase phosphorylation by epigallocatechin gallate with the reduction of matrix metalloproteinase activities in human fibrosarcoma HT1080 cells. J. Agric. Food Chem. 2003, 51, 1858–1863. [Google Scholar] [CrossRef]
- Karin, M. The regulation of AP-1 activity by mitogen-activated protein kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef]
- Whitmarsh, A. J.; Davis, R. J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. 1996, 74, 589–607. [Google Scholar] [CrossRef]
- Lin, A.; Karin, M. NF-kappaB in cancer: a marked target. Semin. Cancer Biol. 2003, 13, 107–114. [Google Scholar]
- Afaq, F.; Adhami, V. M.; Ahmad, N.; Mukhtar, H. Inhibition of ultraviolet B-mediated activation of nuclear factor-κB in normal human epidermal keratinocytes by green tea constituent (-)-epigallocatechin-3-gallate. Oncogene 2003, 22, 1035–1044. [Google Scholar] [CrossRef]
- Gupta, S.; Hastak, K.; Afaq, F.; Ahmad, N.; Mukhtar, H. Essential role of caspases in epigallocatechin-3-gallate-mediated inhibition of nuclear factor-κB and induction of apoptosis. Oncogene 2004, 23, 2507–2522. [Google Scholar] [CrossRef]
- Surh, Y. J.; Chun, K. S.; Cha, H. H. Molecular mechanisms underlying chemoprevention activities of anti-inflammatory phytochemicals: down-regulation of COX-2 and iNOS through suppression of NF-kappaB activation. Mutat. Res. 2001, 480-481, 243–268. [Google Scholar] [CrossRef]
- Karin, M.; Liu, Z. G.; Zandi, E. AP-1 function and regulation. Curr. Opin. Cell Biol. 1997, 9, 240–246. [Google Scholar] [CrossRef]
- Angel, P.; Karin, M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim. Biophys. Acta. 1991, 1072, 129–157. [Google Scholar]
- Dong, Z.; Ma, W.; Huang, C.; Yang, C. S. Inhibition of tumor promoter-induced activator protein 1 activation and cell transformation by tea polyphenols, (-)-epigallocatechin gallate, and theaflavins. Cancer Res. 1997, 57, 4414–4419. [Google Scholar]
- Huang, C.; Ma, W. Y.; Hanenberger, D.; Cleary, M. P.; Bowden, G. T.; Dong, Z. Inhibition of ultraviolet B-induced activator protein-1 (AP-1) activity by aspirin in AP-1-luciferase transgenic mice. J. Biol. Chem. 1997, 272, 26325–26331. [Google Scholar]
- Shimizu, M.; Deguchi, A.; Lim, J. T.; Moriwaki, H.; Kopelovich, L.; Weinstein, I. B. (-)-Epigallocatechin gallate and polyphenol E inhibit growth and activation of the epidermal growth factor receptor and human epidermal growth factor receptor-2 signaling pathways in human colon cancer cells. Clin. Cancer Res. 2005, 11, 2735–2746. [Google Scholar]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A receptor for green tea polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef]
- Jung, Y. D.; Kim, M. S.; Shin, B. A.; Chay, K. O.; Ahn, B. W.; Liu, W.; Bucana, C. D.; Gallick, G. E.; Ellis, L. M. EGCG, a major component of green tea, inhibits tumour growth by inhibiting VEGF induction in human colon carcinoma cells. Br. J. Cancer 2001, 84, 844–850. [Google Scholar] [CrossRef]
- Nihal, M.; Ahmad, N.; Mukhtar, H.; Wood, G. S. Anti-proliferative and proapoptotic effects of (-) -epigallocatechin-3-gallate on human melanoma: Possible implications for the chemoprevention of melanoma. Int. J. Cancer 2005, 11, 513–521. [Google Scholar]
- Kavanagh, K. T.; Hafer, L. J.; Kim, D. W.; Mann, K. K.; Sherr, D. H.; Rogers, A. E.; Sonenshein, G. E. Green tea extracts decrease carcinogen-induced mammary tumor burden in rats and rate of breast cancer cell proliferation in culture. J. Cell Biochem. 2001, 82, 387–398. [Google Scholar] [CrossRef]
- Hastak, K.; Agarwal, M. K.; Mukhtar, H.; Agarwal, M. L. Ablation of either p21 or Bax prevents p53-dependent apoptosis induced by green tea polyphenol epigallocatechin-3-gallate. FASEB J. 2005, 19, 789–791. [Google Scholar]
- Berger, S. J.; Gupta, S.; Belfi, C. A.; Gosky, D. M.; Mukhtar, H. Green tea constituent (-)-epigallocatechin-3-gallate inhibits topoisomerase I activity in human colon carcinoma cells. Biochem. Biophys. Res. Commun. 2001, 288, 101–105. [Google Scholar] [CrossRef]
- Bodnar, A. G.; Ouellette, M.; Frolkis, M.; Holt, S. E.; Chiu, C. P.; Morin, G. B.; Harley, C. B.; Shay, J. W.; Lichtsteiner, S.; Wright, W. E. Extension of life-span by introduction of telomerase into normal human cells. Science (Wash. DC) 1998, 279, 349–352. [Google Scholar] [CrossRef]
- Yang, J.; Chang, E.; Cherry, A. M.; Bangs, C. D.; Oei, Y.; Bodnar, A.; Bronstein, A.; Chiu, C. P.; Herron, G. S. Human endothelial cell life extension by telomerase expression. J. Biol. Chem. 1999, 274, 26141–26148. [Google Scholar] [CrossRef]
- Naasani, I.; Oh-hashi, F.; Oh-hara, T.; Feng, W. Y.; Johnston, J.; Chan, K.; Tsuruo, T. Blocking telomerase by dietary polyphenols is a major mechanism for limiting the growth of human cancer cells in vitro and in vivo. Cancer Res. 2003, 63, 824–830. [Google Scholar]
- Naasani, I.; Seimiya, H.; Tsuruo, T. Telomerase inhibition, telomerase shortening, and senescence of cancer cells by tea catechin. Biochem. Biophys. Res. Comm. 1998, 249, 391–396. [Google Scholar] [CrossRef]
- Hwang, J. T.; Ha, J.; Park, In-Ja.; Lee, S. K.; Baik, H. W.; Kim, Y. M.; Park, O. J. Apoptotic effect of EGCG in HT-29 colon cancer cells via AMPK signal pathway. Cancer Lett. 2007, 247, 115–121. [Google Scholar] [CrossRef]
- Nishikawa, T.; Nakajima, T.; Moriguchi, M.; Jo, M.; Sekoguchi, S.; Ishii, M.; Takashima, H.; Katagishi, T.; Kimura, H.; Minami, M.; Itoh, Y.; Kagawa, K.; Okanoue, T. A green tea polyphenol, epigalocatechin-3-gallate, induces apoptosis of human hepatocellular carcinoma, possibly through inhibition of Bcl-2 family proteins. J. Hepatol. 2006, 44, 1074–1082. [Google Scholar] [CrossRef]
- Lin, J.; Della-Fera, M. A.; Baile, C. A. Green tea polyphenol epigallocatechin gallate inhibits adipogenesis and induces apoptosis in 3T3-L1 adipocytes. Obes. Res. 2005, 13, 982–990. [Google Scholar] [CrossRef]
- Islam, S.; Islam, N.; Kermode, T.; Johnstone, B.; Mukhtar, H.; Moskowitz, R. W.; Goldberg, V. M.; Malemud, C. J.; Haqqi, T. M. Involvement of caspase-3 in epigallocatechin-3-gallate-mediated apoptosis of human chondrosarcoma cells. Biochem. Biophys. Res. Commun. 2000, 270, 793–797. [Google Scholar] [CrossRef]
- Qin, J.; Xie, L. P.; Zheng, X. Y.; Wang, Y. B.; Bai, Y.; Shen, H. F.; Li, L. C.; Dahiya, R. A component of green tea, (-)-epigallocatechin-3-gallate, promotes apoptosis in T24 human bladder cancer cells via modulation of the PI3K/Akt pathway and Bcl-2 family proteins. Biochem. Biophys. Res. Commun. 2007, 354, 852–857. [Google Scholar] [CrossRef]
- Amin, A. R.; Thakur, V. S.; Paul, R. K.; Feng, G. S.; Qu, C. K.; Mukhtar, H.; Agarwal, M. L. SHP-2 tyrosine phosphatase inhibits p73-dependent apoptosis and expression of a subset of p53 target genes induced by EGCG. Proc. Natl. Acad. Sci. USA 2007, 104, 5419–5424. [Google Scholar]
- Manna, S.; Banerjee, S.; Mukherjee, S.; Das, S.; Panda, C. K. Epigallocatechin gallate induced apoptosis in Sarcoma180 cells in vivo: mediated by p53 pathway and inhibition in U1B, U4-U6 UsnRNAs expression. Apoptosis 2006, 11, 2267–2276. [Google Scholar] [CrossRef]
- Hsu, S.; Lewis, J. B.; Borke, J. L.; Singh, B.; Dickinson, D. P.; Caughman, G. B.; Athar, M.; Drake, L.; Aiken, A. C.; Huynh, C. T.; Das, B. R.; Osaki, T.; Schuster, G. S. Chemopreventive effects of green tea polyphenols correlate with reversible induction of p57 expression. Anticancer Res. 2001, 21, 3743–3848. [Google Scholar]
- Hsu, S.; Singh, B. B.; Lewis, J. B.; Borke, J. L.; Dickinson, D. P.; Drake, L.; Caughman, G. B.; Schuster, G. S. Chemoprevention of oral cancer by green tea. Gen. Dent. 2002, 50, 140–146. [Google Scholar]
- Liotta, L. A. Tumor invasion and metastases: role of the extracellular matrix: Rhoads Memorial Award lecture. Cancer Res. 1986, 46, 1–7. [Google Scholar] [CrossRef]
- Conese, M.; Blasi, F. The urokinase/urokinase-receptor system and cancer invasion. Baillieres Clin. Haematol. 1995, 8, 365–389. [Google Scholar] [CrossRef]
- Jankun, J.; Keck, R. W.; Skrzypczak-Jankun, E.; Swiercz, R. Inhibitors of urokinase reduce size of prostate cancer xenografts in severe combined immunodeficient mice. Cancer Res. 1997, 57, 559–563. [Google Scholar]
- Jankun, J.; Selman, S. H.; Swiercz, R.; Skrzypczak-Jankun, E. Why drinking green tea could prevent cancer. Nature 1997, 387, 561. [Google Scholar] [CrossRef]
- Kim, M. H.; Jung, M. A.; Hwang, Y. S.; Jeong, M.; Kim, S. M.; Ahn, S. J.; Shin, B. A.; Ahn, B. W.; Jung, Y. D. Regulation of urokinase plasminogen activator by epigallocatechin-3-gallate in human fibrosarcoma cells. Eur. J. Pharmacol. 2004, 487, 1–6. [Google Scholar] [CrossRef]
- Nagase, H.; Woessner, J. F. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef]
- Foda, H. D.; Zucker, S. Matrix metalloproteinases in cancer invasion, metastasis and angiogenesis. Drug Discov. Today 2001, 6, 478–482. [Google Scholar] [CrossRef]
- Maeta, H.; Ohgi, S.; Terada, T. Protein expression of matrix metalloproteinases 2 and 9 and tissue inhibitors of metalloproteinase 1 and 2 in papillary thyroid carcinomas. Virchows Arch. 2001, 438, 121–128. [Google Scholar] [CrossRef]
- Adhami, V. M.; Siddiqui, I. A.; Ahmad, N.; Gupta, S.; Mukhtar, H. Oral consumption of green tea polyphenols inhibits insulin-like growth factor-I-induced signaling in an autochthonous mouse model of prostate cancer. Cancer Res. 2004, 6, 8715–8722. [Google Scholar]
- Fassina, G.; Vene, R.; Morini, M. Mechanisms of inhibition of tumor angiogenesis and vascular tumor growth by epigallocatechin-3-gallate. Clin. Cancer Res. 2004, 10, 4865–4873. [Google Scholar] [CrossRef]
- Annabi, B.; Lachambre, M. P.; Bousquet-Gagnon, N.; Page, M.; Gingras, D.; Beliveau, R. Green tea polyphenol (-)-epigallocatechin-3-gallate inhibits MMP-2 secretion and MT1-MMP-driven migration in glioblastoma cells. Biochim. Biophys. Acta. 2002, 1542, 209–220. [Google Scholar] [CrossRef]
- Kuzuhara, T.; Sei, Y.; Yamaguchi, K.; Suganuma, M.; Fujiki, H. DNA and RNA as new binding targets of green tea catechins. J. Biol. Chem. 2006, 281, 17446–17456. [Google Scholar] [CrossRef]
- Sample Availability: Not applicable.
© 2007 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.