Modulation of macrophage immune responses by Echinacea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
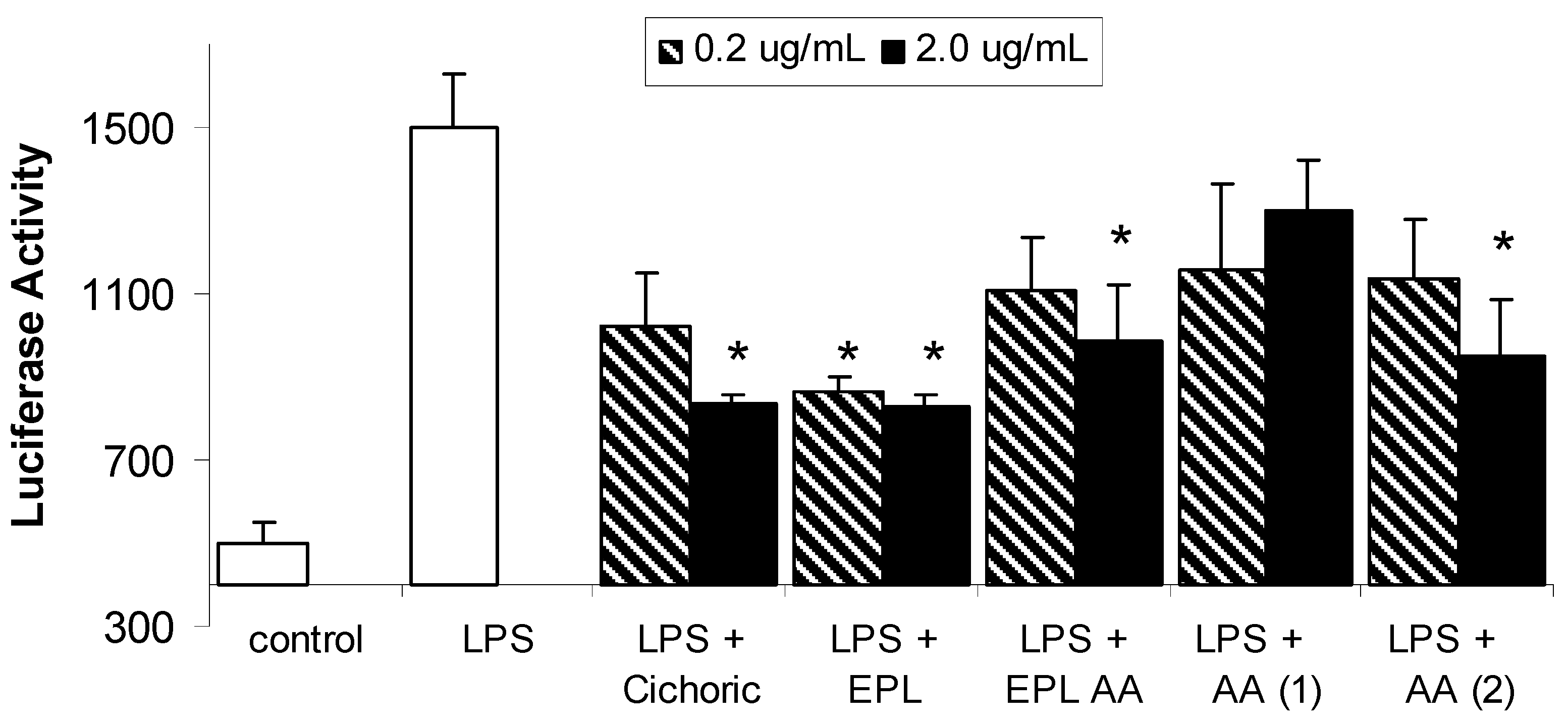
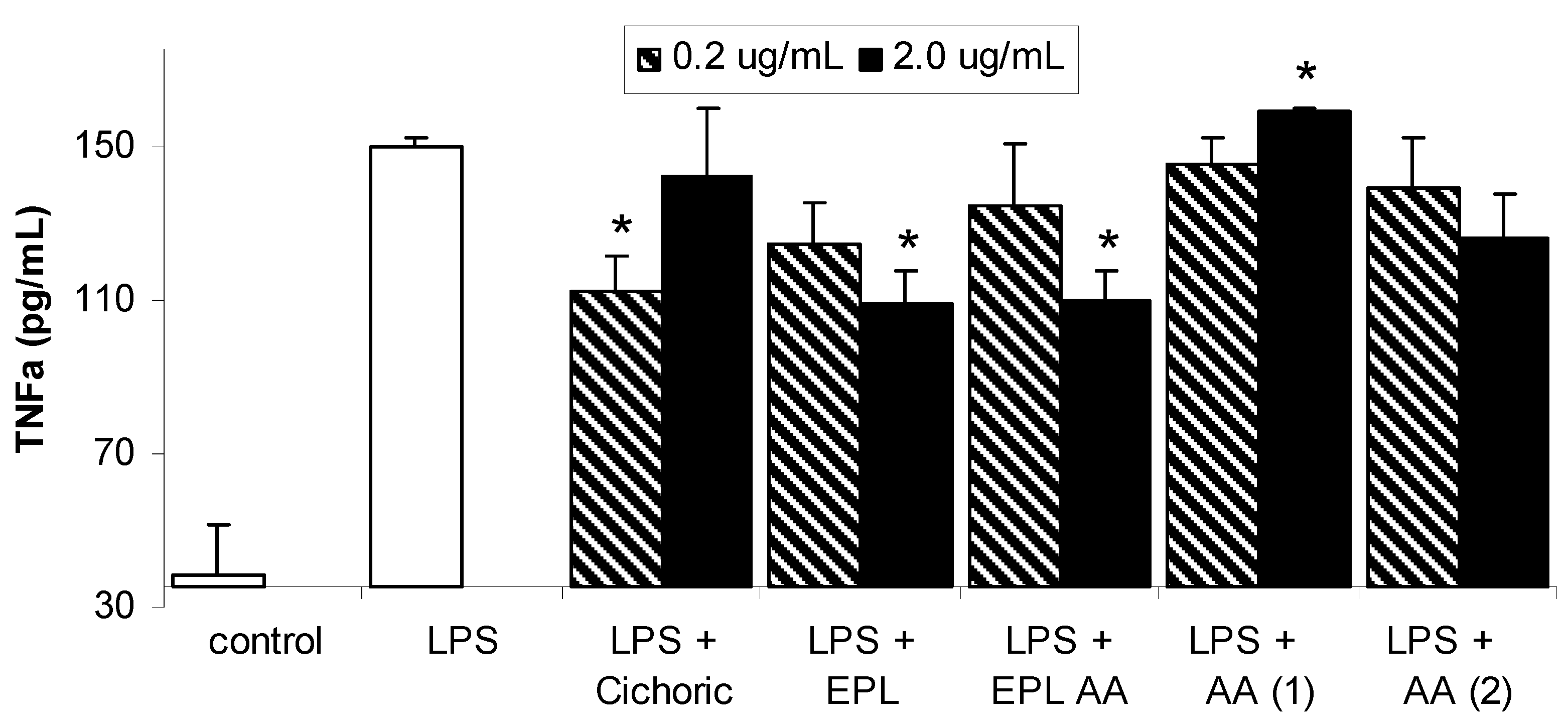
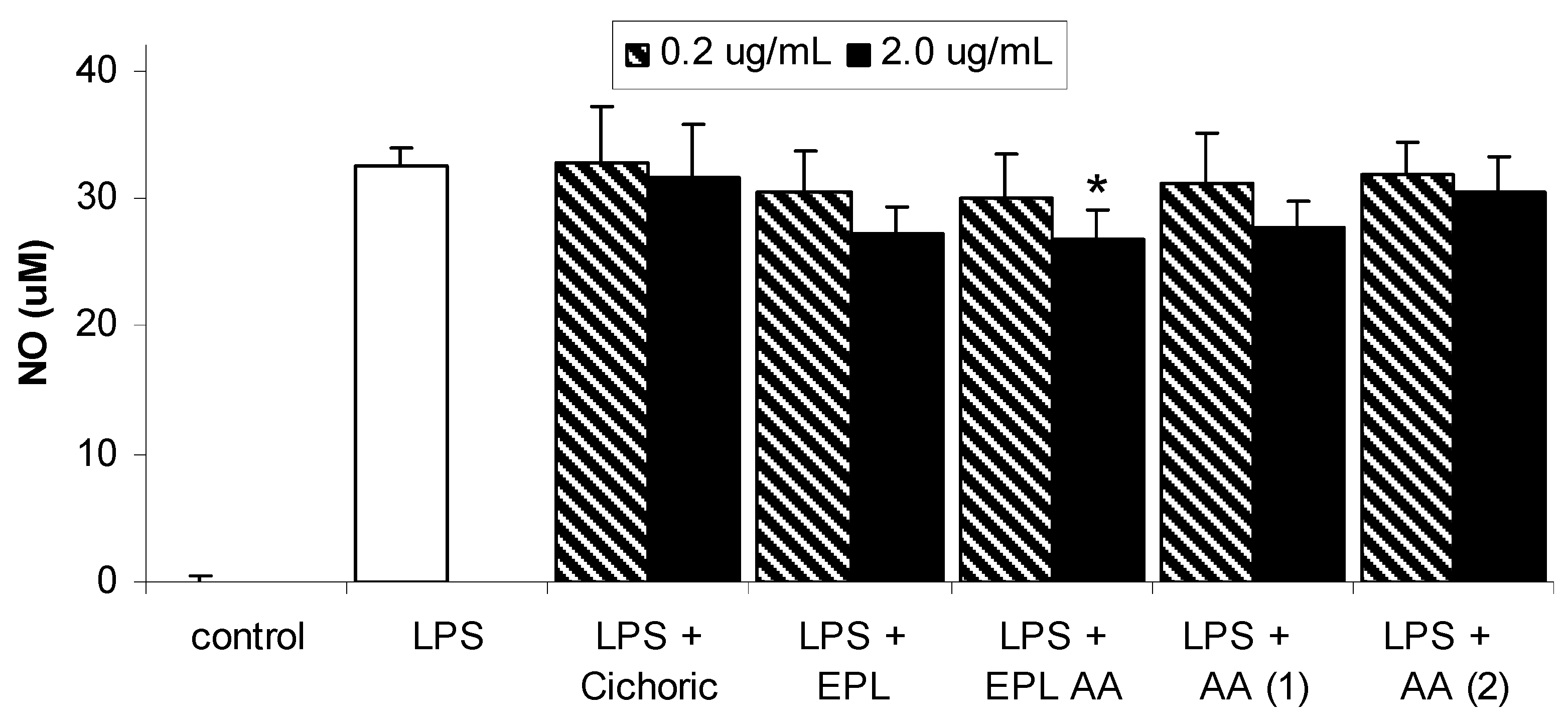
Results and Discussion
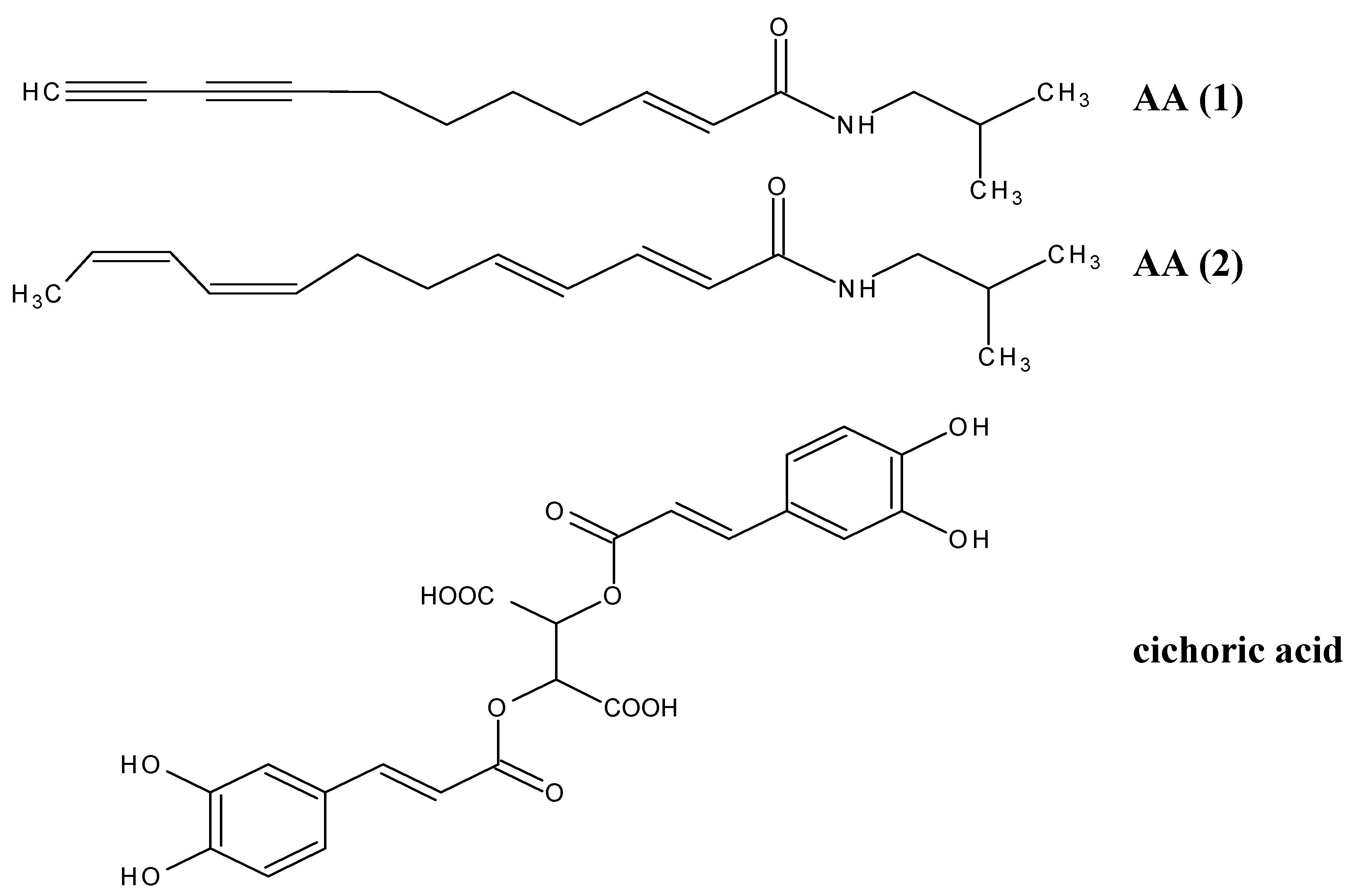
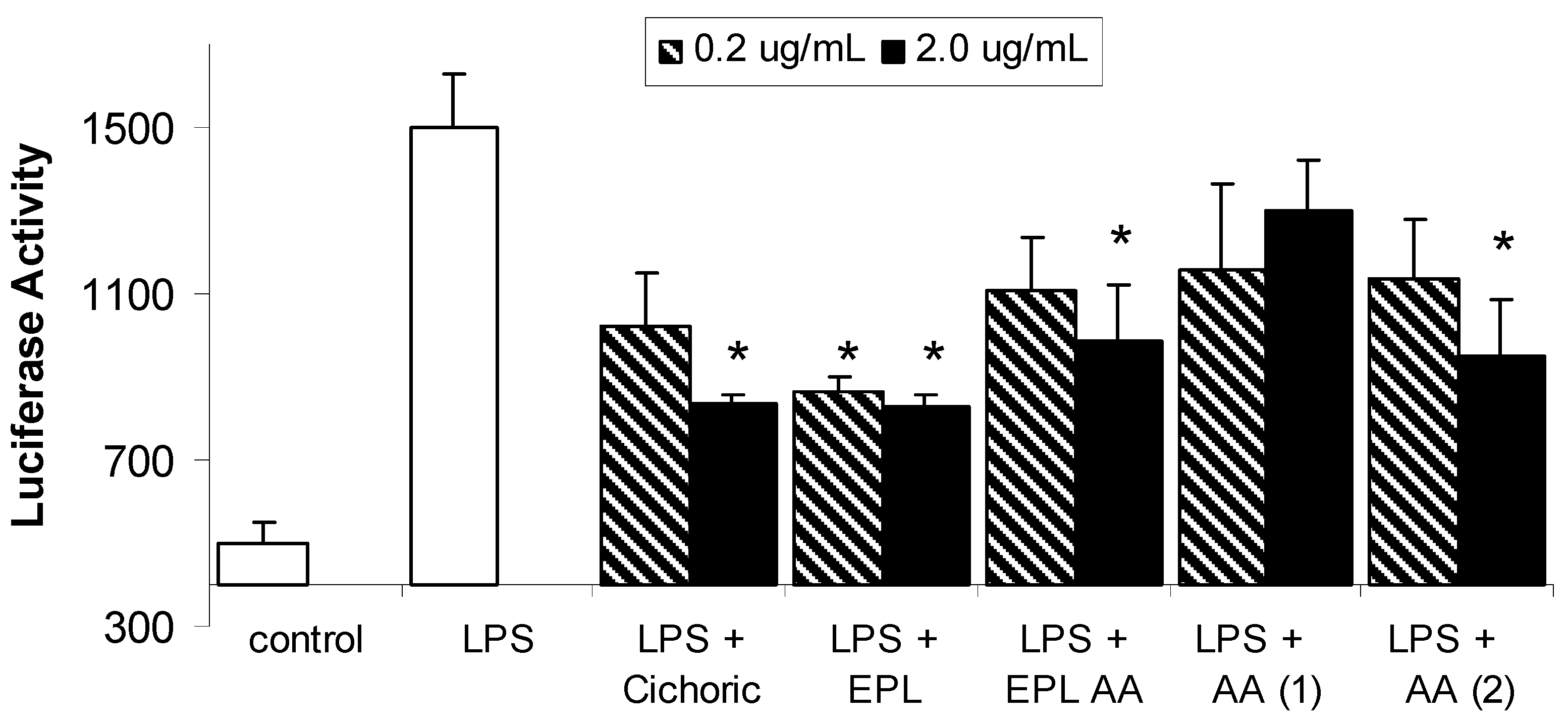
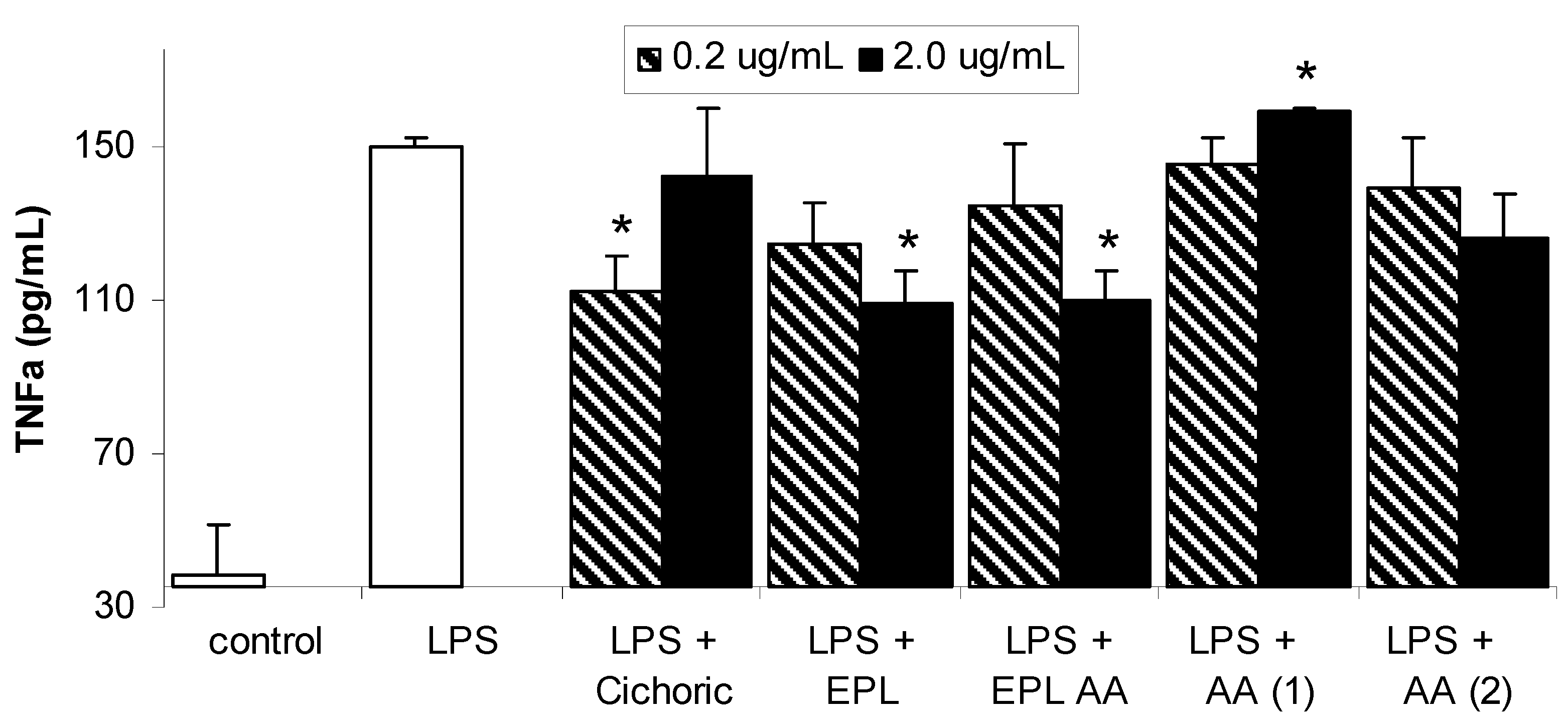
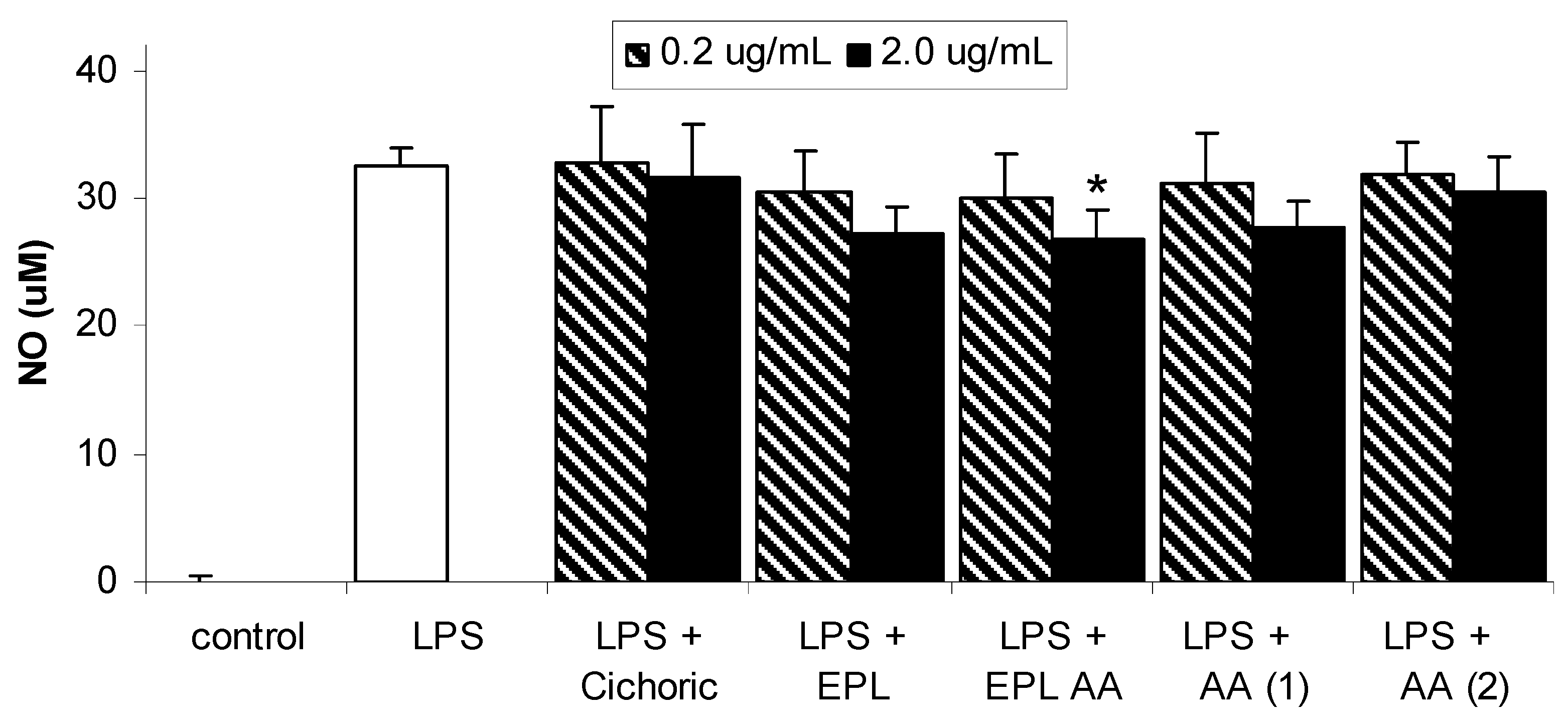
Conclusions
Acknowledgements
Experimental
Sample preparation
Cell Culture
NF-κB activity assay
TNF-α assay
Nitrite assay
Statistical Analysis
References
- Goel, V.; Chang, C.; Slama, J.; Barton, R.; Bauer, R.; Gahler, R.; Basu, T.K. Echinacea stimulates macrophage function in the lung and spleen of normal rats. J. Nutr. Biochem. 2002, 13, 487–492. [Google Scholar] [CrossRef]
- Randolph, R.K.; Gellenbeck, K.; Stonebrook, K.; Brovelli, E.; Qian, Y.; Bankaitis-Davis, D.; Cheronis, J. Regulation of human immune gene expression as influenced by a commercial blended Echinacea product: preliminary studies. Exp. Biol. Med. 2003, 228, 1051–1056. [Google Scholar]
- Forman, H.J.; Torres, M. Redox signalling in macrophages. Mol. Aspects Med. 2001, 22, 189–216. [Google Scholar] [CrossRef]
- Hobbs, A.J.; Higgs, A.; Moncada, S. Inhibition of nitric oxide synthase as a potential therapeutic agent. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Marsden, P.A. Nitric oxide synthases: gene structure and regulation. Adv. Pharmacol. 1995, 34, 71–90. [Google Scholar] [PubMed]
- Xie, Q.W.; Kashiwabara, Y.; Nathan, C. Role of NF<B/Rel in induction of nitric oxide synthase. J. Biol. Chem. 1994, 269, 4705–4708. [Google Scholar] [PubMed]
- Forstermann, U.; Kleinert, H. Nitric oxide synthase: expression and expressional control of the three isoforms. Naunyn Schmidebergs Arch. Pharmacol. 1995, 352, 351–364. [Google Scholar]
- Matthias, A.; Blanchfield, J.T.; Penman, K.G.; Toth, I.; Lang, C-S.; De Voss, J.J.; Lehmann, R.P. Permeability studies of alkylamides and caffeic acid conjugates from echinacea using a Caco-2 cell monolayer model. J. Clin. Pharm. Therap 2004, 29, 7–13. [Google Scholar] [CrossRef]
- Kitchens, R.L.; Munford, R.S. Enzymatically deacylated lipopolysaccharide (LPS) can antagonize LPS at multiple sites in the LPS recognition pathway. J. Biol. Chem. 1995, 270, 9904–9910. [Google Scholar] [PubMed]
- Goel, V.; Chang, C.; Slama, J.; Barton, R.; Bauer, R.; Gahler, R.; Basu, T.K. Alkylamides of Echinacea purpurea stimulate alveolar macrophage function in normal rats. Int. J. Immuno- pharmacol. 2002, 2, 381–387. [Google Scholar] [CrossRef]
- Gertsch, J.; Schoop, R.; Kuenzle, U.; Suter, A. Echinacea alkylamides modulate TNF-α gene expression via cannaboid receptor CB2 and multiple signal transduction pathways. FEBS Letters 2004, 577, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Aktan, F.; Henness, S; Roufogalis, B.D.; Ammit, A.J. Gypenosides derived from Gynostemma pentaphyllum suppress NO synthesis in murine macrophages by inhibiting iNOS enzymatic activity and attenuating NF- κB-mediated iNOS protein expression. Nitric Oxide 2003, 8, 235–242. [Google Scholar] [CrossRef]
- Sample Availability: Not available
© 2005 by MDPI (http:www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Stevenson, L.; Matthias, A.; Banbury, L.; Penman, K.; Bone, K.; Leach, D.; Lehmann, R. Modulation of macrophage immune responses by Echinacea. Molecules 2005, 10, 1279-1285. https://doi.org/10.3390/10101279
Stevenson L, Matthias A, Banbury L, Penman K, Bone K, Leach D, Lehmann R. Modulation of macrophage immune responses by Echinacea. Molecules. 2005; 10(10):1279-1285. https://doi.org/10.3390/10101279
Chicago/Turabian StyleStevenson, L., A. Matthias, L. Banbury, K. Penman, K. Bone, D. Leach, and R. Lehmann. 2005. "Modulation of macrophage immune responses by Echinacea" Molecules 10, no. 10: 1279-1285. https://doi.org/10.3390/10101279